
Tissue-Specific RNA-Seq Analysis of Cotton Roots’ Response to Compound Saline-Alkali Stress and the Functional Validation of the Key Gene GhERF2

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Transcriptome Sequencing and Data Quality Assessment
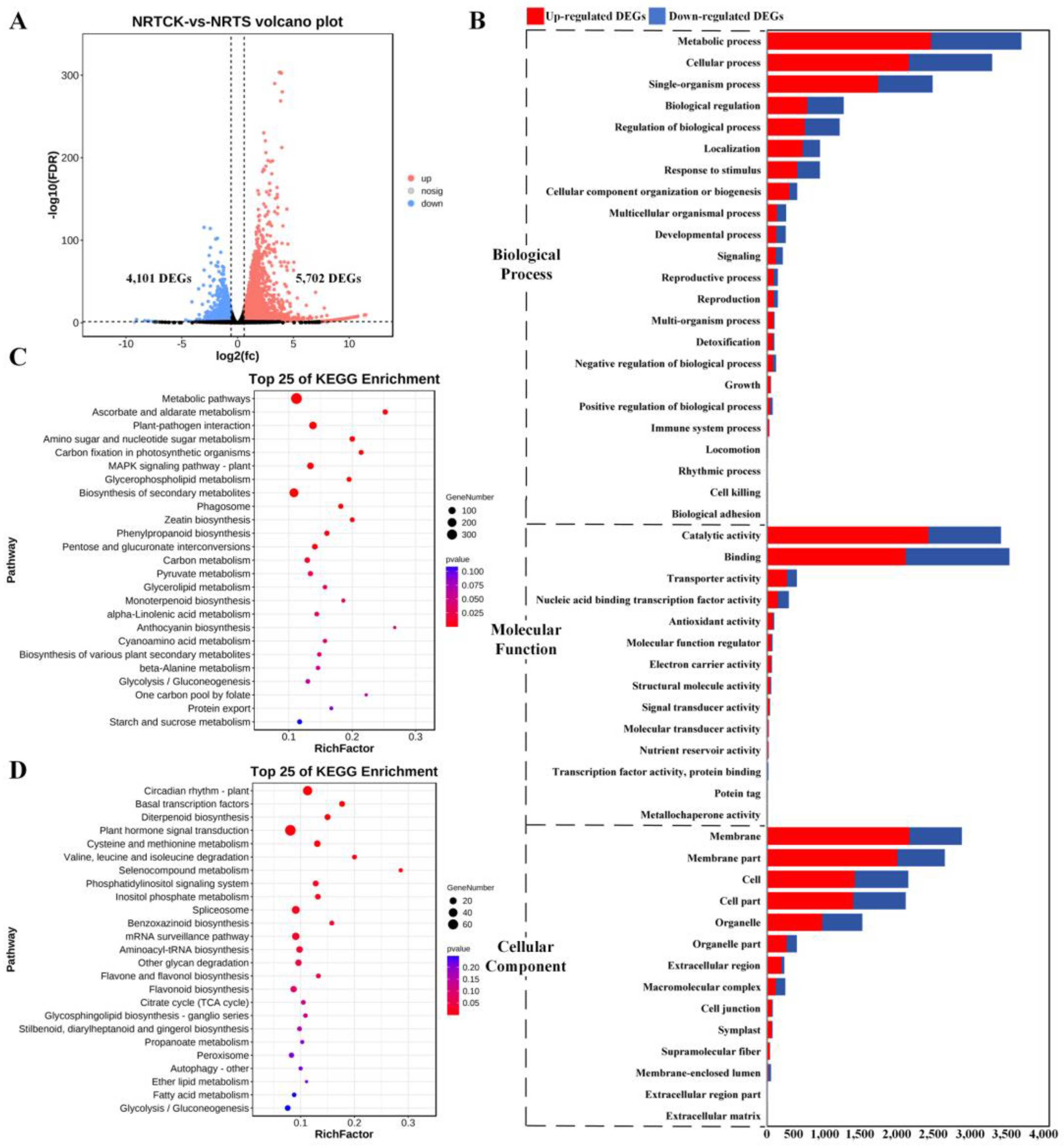
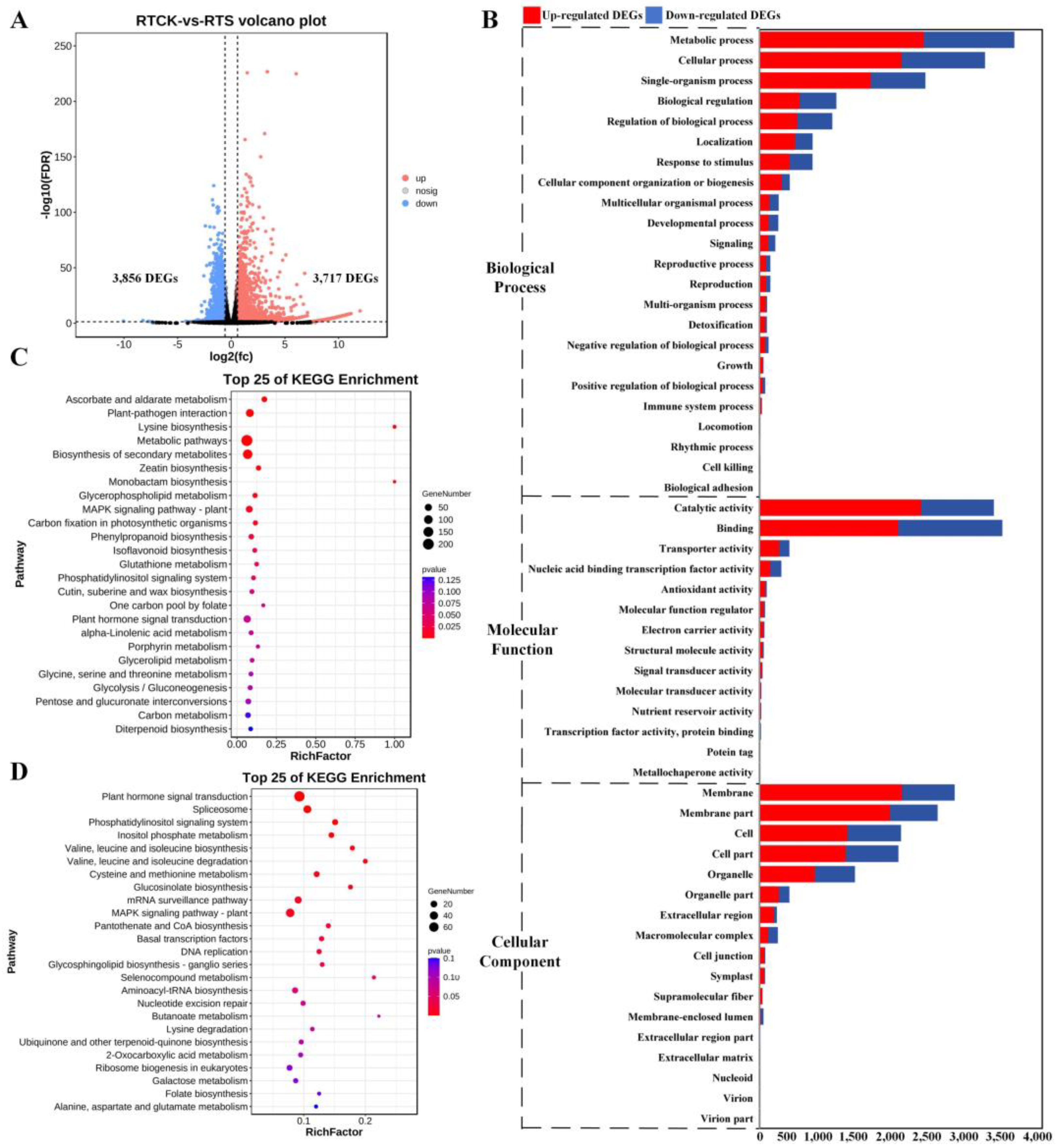
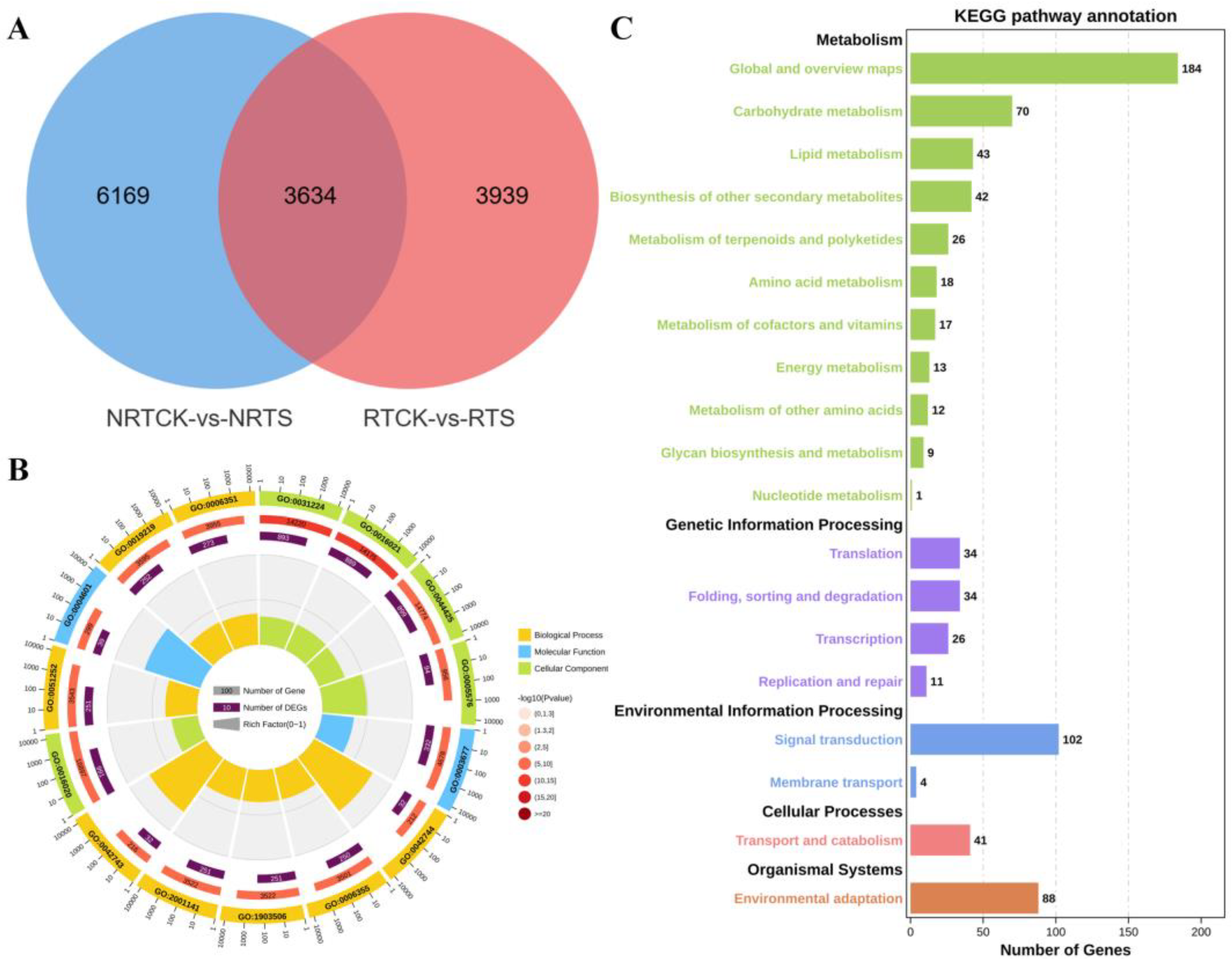
2.2. Identification and Functional Enrichment Analysis of DEGs
2.3. Key DEG Screening and Functional Enrichment Investigation
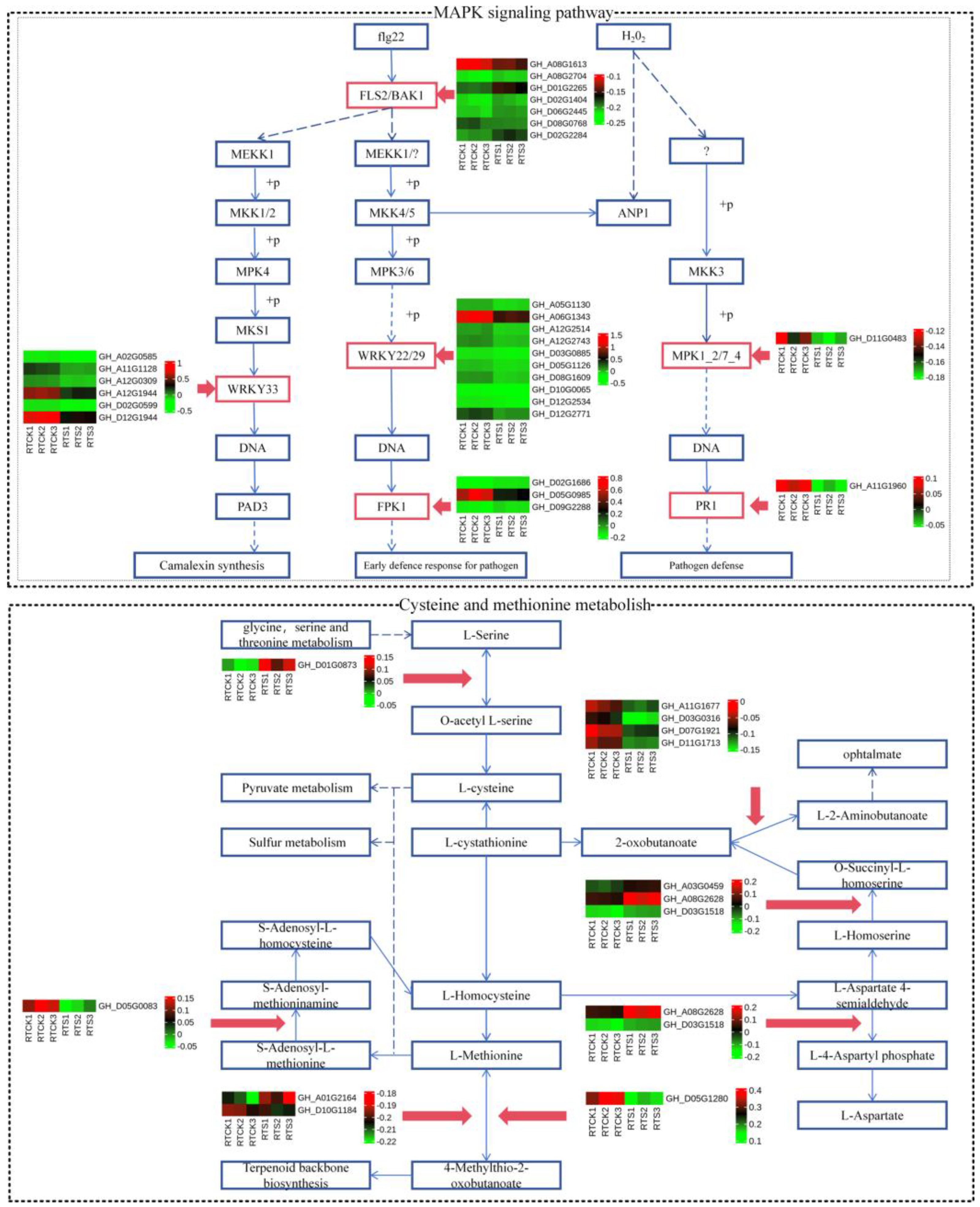
2.4. Analysis of Key Pathways Related to Compound Salinze-Alkali Stress
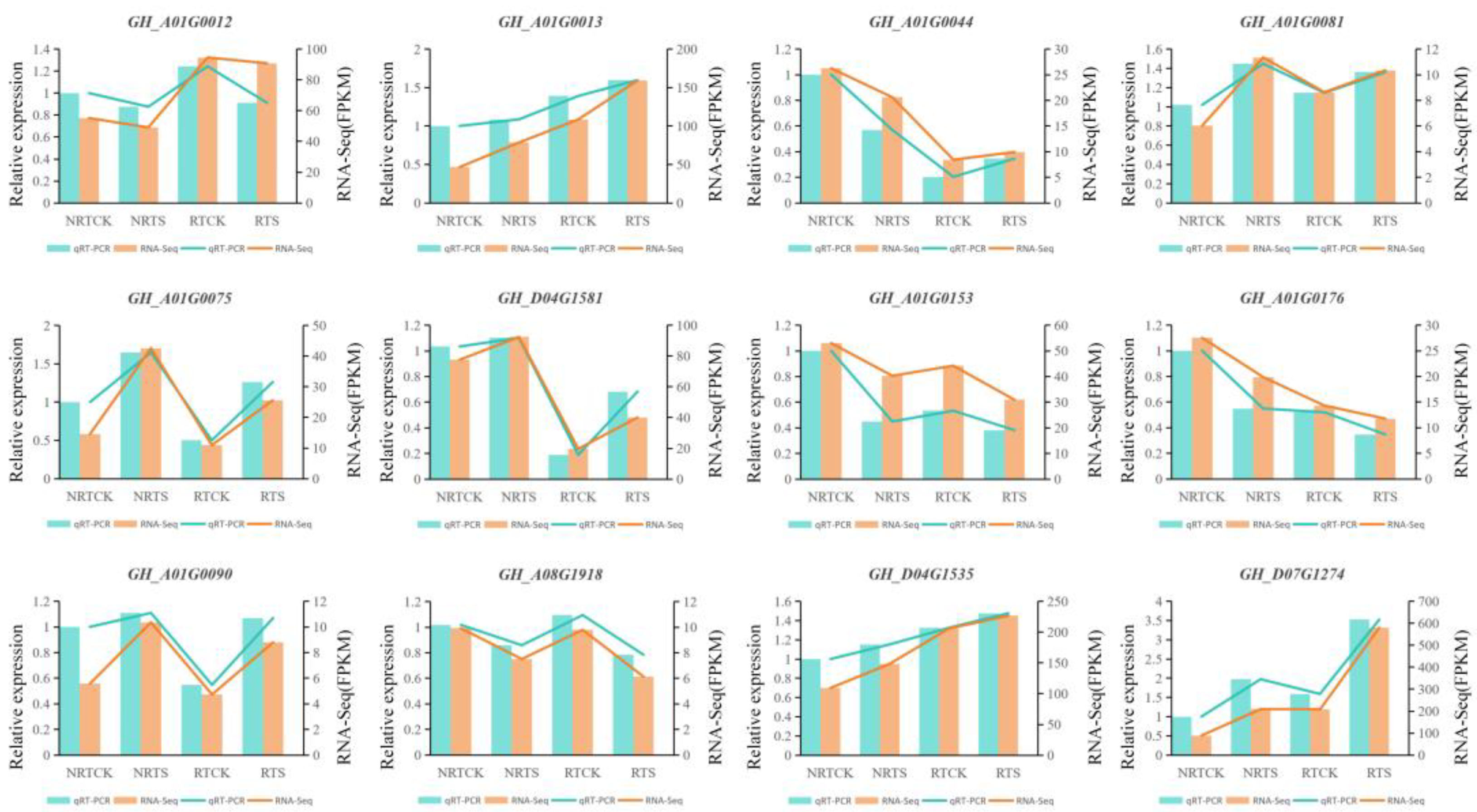
2.5. Validation of RNA-Seq Data Using qRT-PCR Experiment
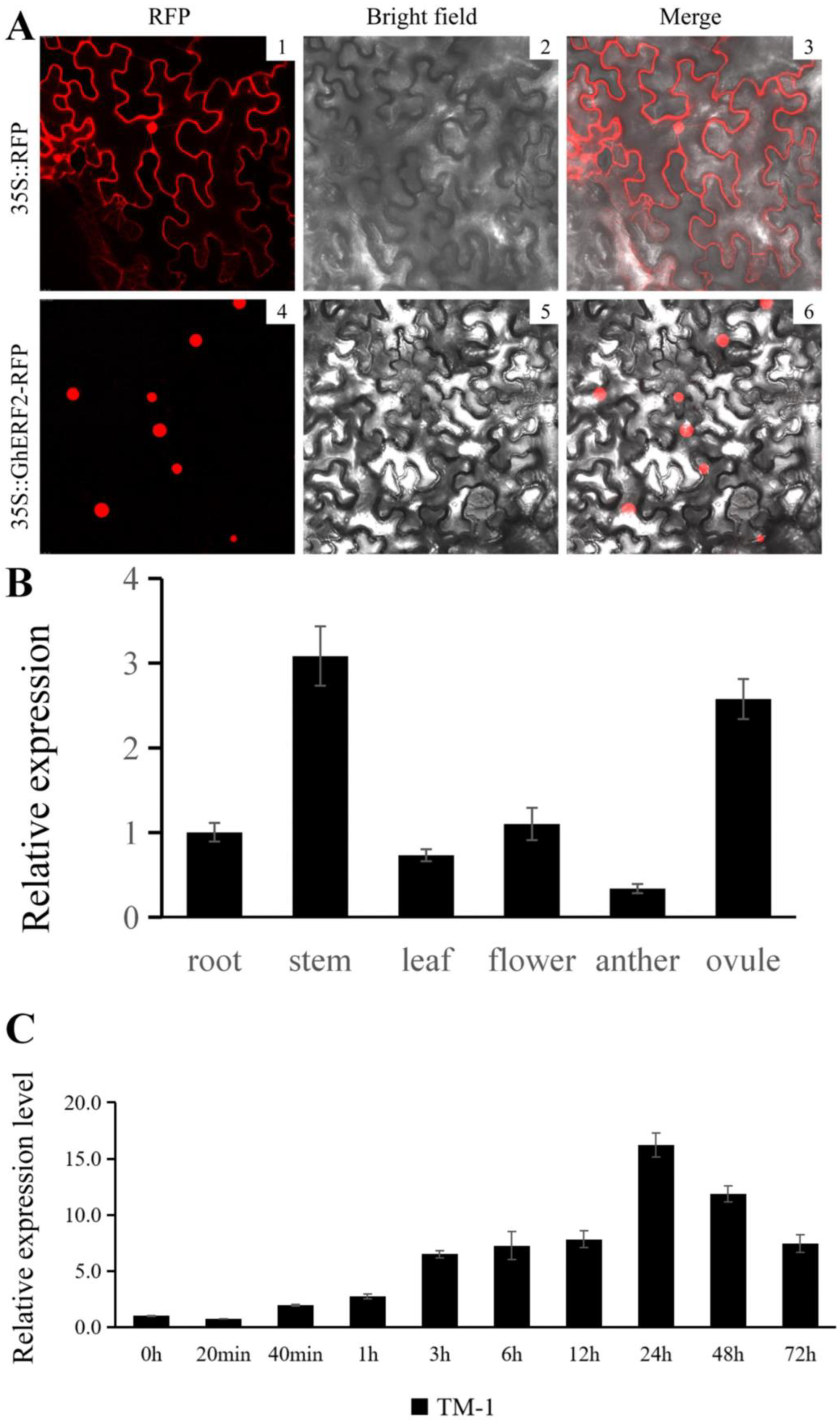
2.6. Expression Pattern Analysis of GhERF2 Under Compound Saline-Alkali Stress
2.7. Silencing of GhERF2 Reduces Cotton Tolerance to Saline-Alkali Stress
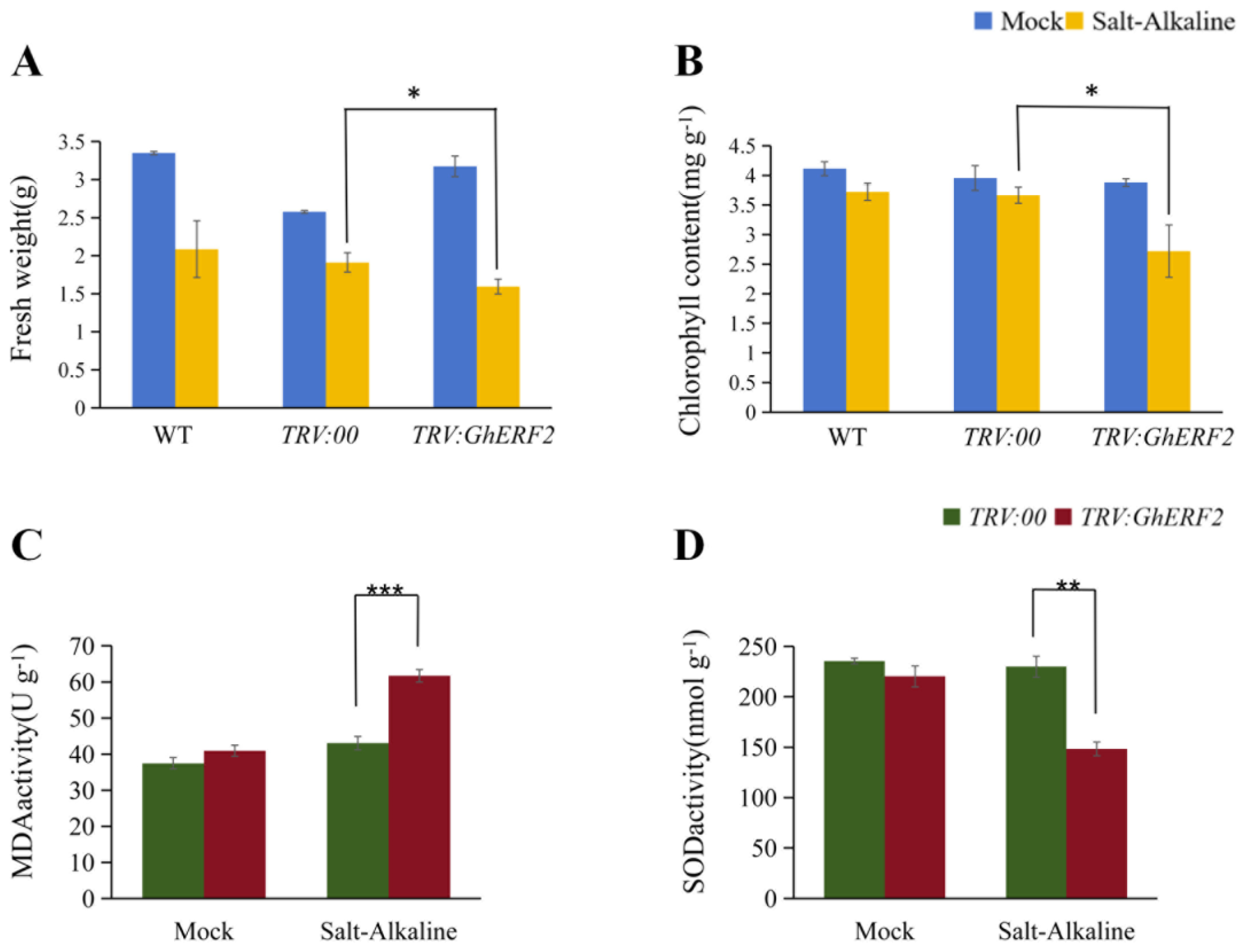
2.8. Physiological Characteristics of GhERF2-Silenced Cotton Plants
3. Discussion
3.1. Transcriptome Sequencing and Analysis
3.2. Role of Candidate Gene GhERF2 in Cotton’s Response to Compound Saline-Alkali Stress
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. RNA Extraction and Library Sequencing
4.3. Transcriptome Sequencing and Data Analysis
4.4. qRT-PCR Analyses of Gene Expression
4.5. Cloning of GhERF2 and Transient Transformation in Tobacco
4.6. Virus-Induced Gene Silencing (VIGS) Experiment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, L.; Wang, B. Protection of Halophytes and Their Uses for Cultivation of Saline-Alkali Soil in China. Biology 2021, 10, 353. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Ren, L.; Wu, L.; Li, J.; Yang, S.; Song, X.; Li, X. Saline-alkali soil amended with biochar derived from maricultural-solid-waste: Ameliorative effect and mechanism. J. Environ. Manag. 2024, 368, 122134. [Google Scholar] [CrossRef] [PubMed]
- Karlova, R.; Boer, D.; Hayes, S.; Testerink, C. Root plasticity under abiotic stress. Plant Physiol. 2021, 187, 1057–1070. [Google Scholar] [CrossRef]
- Gao, L.; Ma, J.B.; Huang, Y.; Muhammad, M.; Lian, H.T.; Shurigin, V.; Egamberdieva, D.; Li, W.J.; Li, L. Insight into endophytic microbial diversity in two halophytes and plant beneficial attributes of Bacillus swezeyi. Front. Microbiol. 2024, 15, 1447755. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Li, Y.; Huang, R. Advances and Challenges in the Breeding of Salt-Tolerant Rice. Int. J. Mol. Sci. 2020, 21, 8385. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Diao, Y.; Jiang, L.; He, Q.; Wang, F.; Hao, W. Exploration of ecological restoration of saline-alkali land based on NbS-Study on the salt resistance and desalination performance of three cash crops. PLoS ONE 2022, 17, e0275828. [Google Scholar] [CrossRef] [PubMed]
- Gulick, S.; Carrico, C.M.; Frey, B.; Baca, D.; Dubey, M.K. Plant versus local soil inorganic ionic composition: The relationship to biomass smoke. Sci. Total Environ. 2023, 895, 164967. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Shi, H.; Yang, Y.; Feng, X.; Chen, X.; Xiao, F.; Lin, H.; Guo, Y. Insights into plant salt stress signaling and tolerance. J. Genet. Genom. 2024, 51, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, X.; You, C.; Wu, X.; Pan, D.; Lv, Z.; Li, T.; Zhang, D.; Shen, Z.; Zhang, X.; et al. Telomere-to-telomere genome of the allotetraploid legume Sesbania cannabina reveals transposon-driven subgenome divergence and mechanisms of alkaline stress tolerance. Sci. China Life Sci. 2024, 67, 149–160. [Google Scholar] [CrossRef]
- Sharma, M.; Tisarum, R.; Kohli, R.K.; Batish, D.R.; Cha-Um, S.; Singh, H.P. Inroads into saline-alkaline stress response in plants: Unravelling morphological, physiological, biochemical, and molecular mechanisms. Planta 2024, 259, 130. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tang, X.; Zhang, N.; Li, S.; Si, H. Role of bZIP Transcription Factors in Plant Salt Stress. Int. J. Mol. Sci. 2023, 24, 7893. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Y.; Jiang, Y.; Li, A.; Cheng, B.; Wu, J. OsASR6 Enhances Salt Stress Tolerance in Rice. Int. J. Mol. Sci. 2022, 23, 9340. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, F.; Xie, P.; Sun, S.; Qiao, X.; Tang, S.; Chen, C.; Yang, S.; Mei, C.; Yang, D.; et al. A Gγ protein regulates alkaline sensitivity in crops. Science 2023, 379, eade8416. [Google Scholar] [CrossRef]
- Sun, Y.; Song, K.; Guo, M.; Wu, H.; Ji, X.; Hou, L.; Liu, X.; Lu, S. A NAC Transcription Factor from ’Sea Rice 86′ Enhances Salt Tolerance by Promoting Hydrogen Sulfide Production in Rice Seedlings. Int. J. Mol. Sci. 2022, 23, 6435. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; An, Y.; Wu, J.; He, S.; Sun, L.; Hao, F. Wheat TaTIP4;1 Confers Enhanced Tolerance to Drought, Salt and Osmotic Stress in Arabidopsis and Rice. Int. J. Mol. Sci. 2022, 23, 2085. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Liu, D.; Wu, P.; Wang, Y.; Gai, Z.; Liu, L.; Yang, F.; Li, C.; Guo, G. Transcriptome analysis of sugar beet (Beta vulgaris L.) in response to alkaline stress. Plant Mol. Biol. 2020, 102, 645–657. [Google Scholar] [CrossRef]
- Wei, L.; Zhang, R.; Zhang, M.; Xia, G.; Liu, S. Functional analysis of long non-coding RNAs involved in alkaline stress responses in wheat. J. Exp. Bot. 2022, 73, 5698–5714. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Si, L.; Zhang, L.; Guo, R.; Wang, R.; Dong, H.; Guo, C. Metabolomics and transcriptomics analysis revealed the response mechanism of alfalfa to combined cold and saline-alkali stress. Plant J. 2024, 119, 1900–1919. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.H.; Chen, X.Y.; Yin, F.X.; Xia, G.M.; Yi, Y.; Zhang, Y.B.; Liu, S.W.; Li, F. Hybridization affects the structure and function of root microbiome by altering gene expression in roots of wheat introgression line under saline-alkali stress. Sci. Total Environ. 2022, 835, 155467. [Google Scholar] [CrossRef] [PubMed]
- Gandullo, J.; Ahmad, S.; Darwish, E.; Karlova, R.; Testerink, C. Phenotyping Tomato Root Developmental Plasticity in Response to Salinity in Soil Rhizotrons. Plant Phenomics 2021, 2021, 2760532. [Google Scholar] [CrossRef]
- Ci, D.; Qin, F.; Tang, Z.; Zhang, G.; Zhang, J.; Si, T.; Yang, J.; Xu, Y.; Yu, T.; Xu, M.; et al. Arbuscular Mycorrhizal Fungi Restored the Saline-Alkali Soil and Promoted the Growth of Peanut Roots. Plants 2023, 12, 3426. [Google Scholar] [CrossRef]
- Nikonorova, N.; Murphy, E.; Fonseca de Lima, C.F.; Zhu, S.; van de Cotte, B.; Vu, L.D.; Balcerowicz, D.; Li, L.; Kong, X.; De Rop, G.; et al. The Arabidopsis Root Tip (Phospho) Proteomes at Growth-Promoting versus Growth-Repressing Conditions Reveal Novel Root Growth Regulators. Cells 2021, 10, 1665. [Google Scholar] [CrossRef] [PubMed]
- Bengough, A.G.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.; Fernandez Garcia, M.N.; Powers, S.J.; Vaughan, S.; Bennett, M.J.; Phillips, A.L.; Thomas, S.G.; Hedden, P. Mapping sites of gibberellin biosynthesis in the Arabidopsis root tip. New Phytol. 2021, 229, 1521–1534. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Zhang, K.; Fang, Y.; Yang, Y.; Cao, X.; Liu, L.; Tian, Y. Systematically and Comprehensively Understanding the Regulation of Cotton Fiber Initiation: A Review. Plants 2023, 12, 3771. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; He, S.; Sun, G.; Pan, Z.; Sun, J.; Geng, X.; Peng, Z.; Gong, W.; Wang, L.; Pang, B.; et al. Favorable pleiotropic loci for fiber yield and quality in upland cotton (Gossypium hirsutum). Sci. Rep. 2021, 11, 15935. [Google Scholar] [CrossRef] [PubMed]
- Shamshina, J.L.; Acharya, S.; Rumi, S.S.; Liyanage, S.; Parajuli, P.; Abidi, N. Cryogenic grinding of cotton fiber cellulose: The effect on physicochemical properties. Carbohydr. Polym. 2022, 289, 119408. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhang, T. MIXTAs and phytohormones orchestrate cotton fiber development. Curr. Opin. Plant Biol. 2021, 59, 101975. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Yang, S.; Dong, S.; Pang, Q.; Smith, P.; Abdalla, M.; Zhang, J.; Wang, G.; Xu, Y. Simulating soil salinity dynamics, cotton yield and evapotranspiration under drip irrigation by ensemble machine learning. Front. Plant Sci. 2023, 14, 1143462. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Liu, X.; Zhang, L.; Zhou, W. Drip irrigation in agricultural saline-alkali land controls soil salinity and improves crop yield: Evidence from a global meta-analysis. Sci. Total Environ. 2023, 880, 163226. [Google Scholar] [CrossRef]
- Sun, S.; Song, H.; Li, J.; Chen, D.; Tu, M.; Jiang, G.; Yu, G.; Zhou, Z. Comparative transcriptome analysis reveals gene expression differences between two peach cultivars under saline-alkaline stress. Hereditas 2020, 157, 9. [Google Scholar] [CrossRef]
- Fan, Y.; Lu, X.; Chen, X.; Wang, J.; Wang, D.; Wang, S.; Guo, L.; Rui, C.; Zhang, Y.; Cui, R.; et al. Cotton transcriptome analysis reveals novel biological pathways that eliminate reactive oxygen species (ROS) under sodium bicarbonate (NaHCO3) alkaline stress. Genomics 2021, 113, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Li, S.; Tian, S.; Wang, B.; Zhao, X. Transcriptome analysis of genes involved in defense against alkaline stress in roots of wild jujube (Ziziphus acidojujuba). PLoS ONE 2017, 12, e0185732. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ye, L.; Zhou, L.; Yu, J.; Pang, B.; Zuo, D.; Gu, L.; Zhu, B.; Du, X.; Wang, H. Co-Expression Network Analysis of the Transcriptome Identified Hub Genes and Pathways Responding to Saline-Alkaline Stress in Sorghum bicolor L. Int. J. Mol. Sci. 2023, 24, 16831. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Li, B.; Xiong, C.; Eneji, A.E.; Zhang, M.; Li, F.; Tian, X.; Li, Z. The Cotton High-Affinity K+ Transporter, GhHAK5a, Is Essential for Shoot Regulation of K+ Uptake in Root under Potassium Deficiency. Plant Cell Physiol. 2019, 60, 888–899. [Google Scholar] [CrossRef]
- Wang, C.; Lu, W.; He, X.; Wang, F.; Zhou, Y.; Guo, X.; Guo, X. The Cotton Mitogen-Activated Protein Kinase Kinase 3 Functions in Drought Tolerance by Regulating Stomatal Responses and Root Growth. Plant Cell Physiol. 2016, 57, 1629–1642. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, Y.; Rui, C.; Zhang, H.; Xu, N.; Wang, J.; Han, M.; Lu, X.; Chen, X.; Wang, D.; et al. Molecular structures and functional exploration of NDA family genes respond tolerant to alkaline stress in Gossypium hirsutum L. Biol. Res. 2022, 55, 4. [Google Scholar] [CrossRef] [PubMed]
- Sui, D.; Wang, B.; El-Kassaby, Y.A.; Wang, L. Integration of Physiological, Transcriptomic, and Metabolomic Analyses Reveal Molecular Mechanisms of Salt Stress in Maclura tricuspidata. Plants 2024, 13, 397. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lu, X.; Chen, X.; Malik, W.A.; Wang, D.; Zhao, L.; Wang, J.; Wang, S.; Guo, L.; Cui, R.; et al. Transcriptome analysis of upland cotton revealed novel pathways to scavenge reactive oxygen species (ROS) responding to Na2SO4 tolerance. Sci. Rep. 2021, 11, 8670. [Google Scholar] [CrossRef]
- Seok, H.Y.; Tran, H.T.; Lee, S.Y.; Moon, Y.H. AtERF71/HRE2, an Arabidopsis AP2/ERF Transcription Factor Gene, Contains Both Positive and Negative Cis-Regulatory Elements in Its Promoter Region Involved in Hypoxia and Salt Stress Responses. Int. J. Mol. Sci. 2022, 3, 5310. [Google Scholar] [CrossRef]
- Hong, Y.; Wang, H.; Gao, Y.; Bi, Y.; Xiong, X.; Yan, Y.; Wang, J.; Li, D.; Song, F. ERF Transcription Factor OsBIERF3 Positively Contributes to Immunity against Fungal and Bacterial Diseases but Negatively Regulates Cold Tolerance in Rice. Int. J. Mol. Sci. 2022, 23, 606. [Google Scholar] [CrossRef]
- Zhu, Z.; Shi, J.; Xu, W.; Li, H.; He, M.; Xu, Y.; Xu, T.; Yang, Y.; Cao, J.; Wang, Y. Three ERF transcription factors from Chinese wild grapevine Vitis pseudoreticulata participate in different biotic and abiotic stress-responsive pathways. J. Plant Physiol. 2013, 170, 923–933. [Google Scholar] [CrossRef]
- Palumbo, F.; Gabelli, G.; Pasquali, E.; Vannozzi, A.; Farinati, S.; Draga, S.; Ravi, S.; Della Lucia, M.C.; Bertoldo, G. Barcaccia RNA-seq analyses on gametogenic tissues of alfalfa (Medicago sativa) revealed plant reproduction- and ploidy-related genes. BMC Plant Biol. 2024, 24, 826. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Zhan, L.; Zhang, J.; Song, W.; Xu, X. Identification of Rapeseed (Brassica napus L.) Plant Height-Associated QTL Using BSA-seq and RNA-seq. Int. J. Mol. Sci. 2024, 25, 9875. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, J.; Wei, S.; Gao, Y.; Pei, H.; Geng, R.; Lu, Z.; Wang, P.; Zhou, W. Maize GOLDEN2-LIKE proteins enhance drought tolerance in rice by promoting stomatal closure. Plant Physiol. 2024, 194, 774–786. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, C.; Xi, J.; Wang, Y.; Guo, J.; Liu, Q.; Liu, Y.; Ma, Y.; Zhang, J.; Ma, F.; et al. The MdHSC70-MdWRKY75 module mediates basal apple thermotolerance by regulating the expression of heat shock factor genes. Plant Cell. 2024, 36, 3631–3653. [Google Scholar] [CrossRef] [PubMed]
- Gomes, G.L.B.; Scortecci, K.C. Auxin and its role in plant development: Structure, signalling, regulation and response mechanisms. Plant Biol. 2021, 23, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yan, X.; Wu, Z.; Hou, L.; Li, M. Transcriptome Sequencing Reveals the Mechanism of Auxin Regulation during Root Expansion in Carrot. Int. J. Mol. Sci. 2024, 25, 3425. [Google Scholar] [CrossRef] [PubMed]
- Shtin, M.; Dello Ioio, R.; Del Bianco, M. It’s Time for a Change: The Role of Gibberellin in Root Meristem Development. Front. Plant Sci. 2022, 13, 882517. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Lian, Y.; Wang, J.; Yong, T.; Gao, H.; Wu, H.; Yang, T.; Wang, C. AtHSPR functions in gibberellin-mediated primary root growth by interacting with KNAT5 and OFP1 in Arabidopsis. Plant Cell Rep. 2023, 2, 1629–1649. [Google Scholar] [CrossRef]
- Lantzouni, O.; Alkofer, A.; Falter-Braun, P.; Schwechheimer, C. GROWTH-REGULATING FACTORS Interact with DELLAs and Regulate Growth in Cold Stress. Plant Cell. 2020, 32, 1018–1034. [Google Scholar] [CrossRef]
- Takatsuka, H.; Sasaki, A.; Takahashi, N.; Shibata, M.; Sugimoto, K.; Tanaka, M.; Seki, M.; Umeda, M. Cytokinin signaling promotes root hair growth by directly regulating RSL4 expression. J. Exp. Bot. 2023, 74, 3579–3594. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.E.; Marzol, E.; Estevez, J.M.; Muday, G.K. Ethylene signaling increases reactive oxygen species accumulation to drive root hair initiation in Arabidopsis. Development 2022, 149, dev200487. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Hartman, S.; van Veen, H.; Zhang, H.; Leeggangers, H.A.C.F.; Martopawiro, S.; Bosman, F.; de Deugd, F.; Su, P.; Hummel, M.; et al. Ethylene augments root hypoxia tolerance via growth cessation and reactive oxygen species amelioration. Plant Physiol. 2022, 190, 1365–1383. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Lee, H.S.; Han, H.K.; Choi, C.I. Saikosaponin A and D Inhibit Adipogenesis via the AMPK and MAPK Signaling Pathways in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2021, 22, 11409. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Meng, X. MAPK Signaling: Emerging Roles in Lateral Root Formation. Trends Plant Sci. 2020, 25, 126–129. [Google Scholar] [CrossRef]
- Bandyopadhyay, P.; Pramanick, I.; Biswas, R.; Ps, S.; Sreedharan, S.; Singh, S.; Rajmani, R.S.; Laxman, S.; Dutta, S.; Singh, A. Adenosylmethionine-responsive cystathionine β-synthase modulates sulfur metabolism and redox balance in Mycobacteriumtuberculosis. Sci. Adv. 2022, 8, eabo0097. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Chiba, Y.; Hirai, M.Y. Metabolism and Regulatory Functions of O-Acetylserine, S-Adenosylmethionine, Homocysteine, and Serine in Plant Development and Environmental Responses. Front. Plant Sci. 2021, 2, 643403. [Google Scholar] [CrossRef]
- Andrés-Barrao, C.; Alzubaidy, H.; Jalal, R.; Mariappan, K.G.; de Zélicourt, A.; Bokhari, A.; Artyukh, O.; Alwutayd, K.; Rawat, A.; Shekhawat, K.; et al. Coordinated bacterial and plant sulfur metabolism in Enterobacter sp. SA187-induced plant salt stress tolerance. Proc. Natl. Acad. Sci. USA 2021, 118, e2107417118. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Becana, M. Molecular responses of legumes to abiotic stress: Post-translational modifications of proteins and redox signaling. J. Exp. Bot. 2021, 72, 5876–5892. [Google Scholar] [CrossRef]
- Chen, K.; Tang, W.; Zhou, Y.; Chen, J.; Xu, Z.; Ma, R.; Dong, Y.; Ma, Y.; Chen, M. AP2/ERF transcription factor GmDREB1 confers drought tolerance in transgenic soybean by interacting with GmERFs. Plant Physiol. Biochem. 2022, 170, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yang, L.; Li, A.; Guo, J.; Wang, H.; Qi, H.; Li, M.; Yang, P.; Song, S. An AP2/ERF transcription factor confers chilling tolerance in rice. Sci. Adv. 2024, 10, eado4788. [Google Scholar] [CrossRef]
- Li, Z.; Tian, Y.; Xu, J.; Fu, X.; Gao, J.; Wang, B.; Han, H.; Wang, L.; Peng, R.; Yao, Q. A tomato ERF transcription factor, SlERF84, confers enhanced tolerance to drought and salt stress but negatively regulates immunity against Pseudomonas syringae pv. tomato DC3000. Plant Physiol. Biochem. 2018, 132, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Yan, X.; Guo, J. Physiological and Transcriptomic Analyses of Yellow Horn (Xanthoceras Sorbifolia) Provide Important Insights into Salt and Saline-Alkali Stress Tolerance. PLoS ONE 2020, 15, e0244365. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Ma, B.; Bu, X.; Dang, Z.; Wang, Y. Transcriptional Profiling Analysis Providing Insights into the Harsh Environments Tolerance Mechanisms of Krascheninnikovia Arborescens. Int. J. Mol. Sci. 2024, 25, 11891. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, A.; Duan, X.; Wang, S.; Sun, X.; Duanmu, H.; Zhu, D.; Chen, C.; Cao, L.; Xiao, J.; et al. GsERF6, an Ethylene-Responsive Factor from Glycine Soja, Mediates the Regulation of Plant Bicarbonate Tolerance in Arabidopsis. Planta 2016, 244, 681–698. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K.A.; Virdee, S.K.; McArthur, A.G. Strandedness during cDNA Synthesis, the Stranded Parameter in Htseq-Count and Analysis of RNA-Seq Data. Brief. Funct. Genom. 2020, 19, 339–342. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Z.; Zhu, R.; Wang, F.; Cheng, Y.; Liu, Y. Three Differential Expression Analysis Methods for RNA Sequencing: Limma, EdgeR, DESeq2. J. Vis. Exp. 2021, 175, e62528. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, J.; Sun, L.; Ma, Y.; Xu, J.; Liang, S.; Deng, J.; Tan, J.; Zhang, Q.; Tu, L.; et al. High efficient multisites genome editing in allotetraploid cotton (Gossypium hirsutum) using CRISPR/Cas9 system. Plant Biotechnol. J. 2018, 16, 137–150. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Clean Reads | Clean Base | Read Length | Q20 (%) | GC (%) |
|---|---|---|---|---|---|
| NRCK1 | 24,013,230 | 7,203,969,000 | PE150 | 96.30 | 43.93 |
| NRCK2 | 24,069,156 | 7,220,746,800 | PE150 | 96.56 | 43.98 |
| NRCK3 | 24,102,895 | 7,230,868,500 | PE150 | 96.01 | 43.94 |
| RCK1 | 23,546,010 | 7,063,803,000 | PE150 | 96.19 | 44.27 |
| RCK2 | 24,006,143 | 7,201,842,900 | PE150 | 96.45 | 44.18 |
| RCK3 | 24,115,835 | 7,234,750,500 | PE150 | 96.30 | 44.07 |
| RS1 | 24,097,237 | 7,229,171,100 | PE150 | 96.53 | 44.43 |
| RS2 | 24,130,131 | 7,239,039,300 | PE150 | 96.54 | 44.41 |
| RS3 | 24,044,474 | 7,213,342,200 | PE150 | 96.59 | 44.49 |
| NRS1 | 24,073,680 | 7,222,104,000 | PE150 | 96.58 | 44.70 |
| NRS2 | 24,138,006 | 7,241,401,800 | PE150 | 96.72 | 44.73 |
| NRS3 | 24,114,157 | 7,234,247,100 | PE150 | 96.62 | 44.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.; Liu, Q.; Du, X.; Xing, B.; Zhang, S.; Li, Y.; Hao, L.; Wei, Y.; Liu, Y.; Li, P.; et al. Tissue-Specific RNA-Seq Analysis of Cotton Roots’ Response to Compound Saline-Alkali Stress and the Functional Validation of the Key Gene GhERF2. Plants 2025, 14, 756. https://doi.org/10.3390/plants14050756
Zhang A, Liu Q, Du X, Xing B, Zhang S, Li Y, Hao L, Wei Y, Liu Y, Li P, et al. Tissue-Specific RNA-Seq Analysis of Cotton Roots’ Response to Compound Saline-Alkali Stress and the Functional Validation of the Key Gene GhERF2. Plants. 2025; 14(5):756. https://doi.org/10.3390/plants14050756
Chicago/Turabian StyleZhang, Aiming, Qiankun Liu, Xue Du, Baoguang Xing, Shaoliang Zhang, Yanfang Li, Liuan Hao, Yangyang Wei, Yuling Liu, Pengtao Li, and et al. 2025. "Tissue-Specific RNA-Seq Analysis of Cotton Roots’ Response to Compound Saline-Alkali Stress and the Functional Validation of the Key Gene GhERF2" Plants 14, no. 5: 756. https://doi.org/10.3390/plants14050756
APA StyleZhang, A., Liu, Q., Du, X., Xing, B., Zhang, S., Li, Y., Hao, L., Wei, Y., Liu, Y., Li, P., Hu, S., & Peng, R. (2025). Tissue-Specific RNA-Seq Analysis of Cotton Roots’ Response to Compound Saline-Alkali Stress and the Functional Validation of the Key Gene GhERF2. Plants, 14(5), 756. https://doi.org/10.3390/plants14050756