
Current Distribution of the Invasive Kelp Undaria pinnatifida (Harvey) Suringar, 1873 Along Artificial and Natural Habitats in North Portugal—Impacts and Mitigation Initiatives

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results
2.1. Environmental Conditions and Associated Macroalgae
2.2. Distribution of Undaria pinnatifida
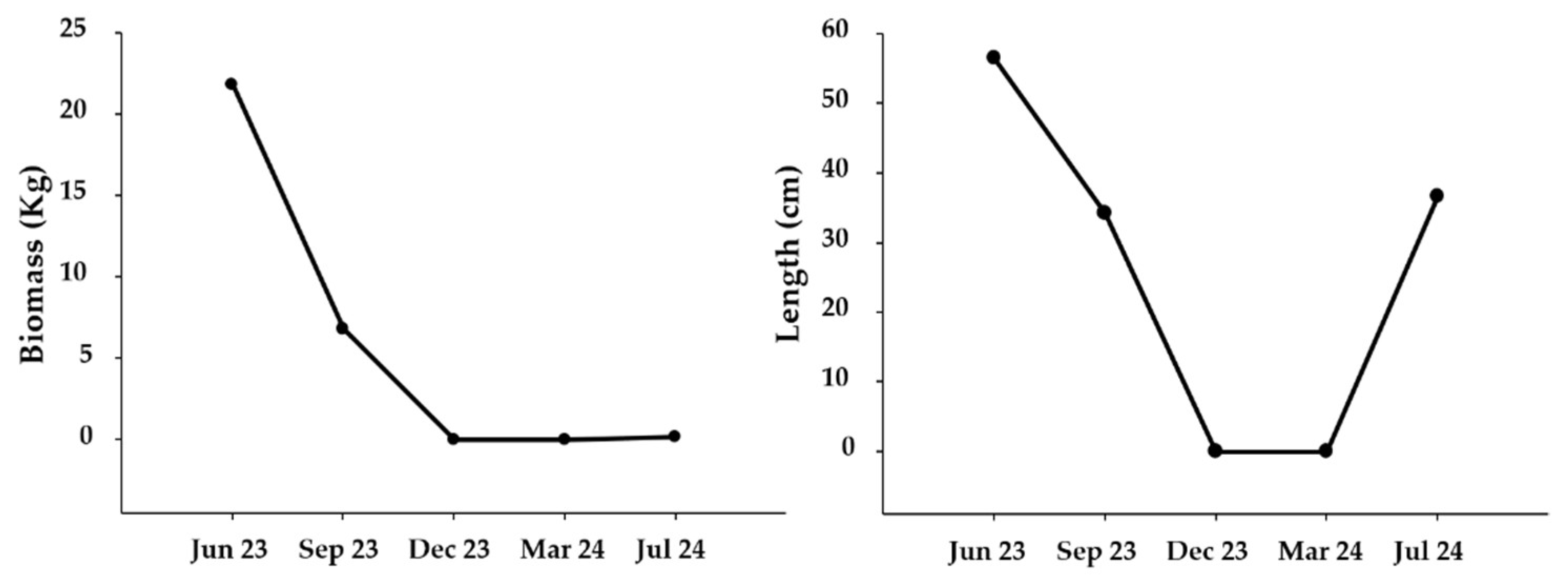
2.3. Mitigation Experiment
3. Discussion
4. Materials and Methods
4.1. Studied Area
4.2. Field and Laboratory Methods
4.3. Mitigation Experiment
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marinas | Salinity (psu) | Temperature (°C) | Dissolved Oxygen (mg L−1) |
|---|---|---|---|
| Viana-m | 34.08 ± 0.15 | 17.72 ± 0.55 | 10.00 ± 0.28 |
| Viana-e | 21.49 ± 2.74 | 21.20 ± 1.00 | 8.56 ± 0.40 |
| Esposende | 12.27 ± 9.55 | 19.00 ± 0.71 | 7.68 ± 0.63 |
| Povoa do Varzim | 31.17 ± 0.79 | 16.50 ± 0.32 | 8.96 ± 0.39 |
| Leça da Palmeira | 33.27 ± 1.25 | 19.87 ± 0.91 | 8.65 ± 0.40 |
| Afurada | 21.55 ± 5.33 | 21.42 ± 0.59 | 8.22 ± 0.18 |
| Macroalgae | Viana-m | Viana-e | Esposende | Póvoa de Varzim | Leça da Palmeira | Afurada |
|---|---|---|---|---|---|---|
| Ulva spp. | + | + | + | + | + | + |
| Grateloupia turuturu Yamada, 1941 | + | − | − | + | + | − |
| Chondrus crispus Stackhouse 1797 | − | − | − | − | + | − |
| Fucus aff spiralis Linnaeus 1753 | − | + | + | − | − | + |
References
- Funahashi, S. Marine algae from Vladivostok and its vicinity. Bull. Jpn. Soc. Phycol. 1966, 14, 127–145. [Google Scholar]
- Akiyama, K.; Kurogi, M. Cultivation of Undaria pinnatifida (Harvey) Suringar, the decrease in crops from natural plants following crops increase from cultivation. Bull. Tohoku Reg. Fish. Res. Lab. 1982, 44, 91–100. [Google Scholar]
- Zhang, D.M.; Miao, G.R.; Pei, L.Q. Studies on Undaria pinnatifida. Hydrobiologia 1984, 116, 263–265. [Google Scholar] [CrossRef]
- Perez, R.; Kaas, R.; Barbaroux, O. Culture experimentale de l’algue Undaria pinnatifida sur les cotes des France. Sci. Pech. 1984, 343, 5–15. [Google Scholar]
- Floc’h, J.Y.; Pajot, R.; Wallentinus, I. The Japanese brown alga Undaria pinnatifida on the coasts of France and the possibilities of its establishment in European waters. J. Cons. Int. Explor. Met. 1991, 47, 379–390. [Google Scholar] [CrossRef]
- Floc’h, J.Y.; Pajot, R.; Mouret, V. Undaria pinnatifida (Laminariales, Phaeophyta) 12 years after its introduction into the Atlantic Ocean. Hydrobiologia 1996, 326, 217–222. [Google Scholar] [CrossRef]
- Pérez, R.; Lee, J.Y.; Juge, C. Observations sur la biologie de l’algue japonaise Undaria pinnatifida (Harvey) Suringar introduite accidentellemant dans l’Étang de Thau. Sci. Pech. 1981, 315, 1–12. [Google Scholar]
- Schiller, J.; Lackschewitz, D.; Buschbaum, C.; Reise, K.; Pang, S.; Bischof, K. Heading northward to Scandinavia: Undaria pinnatifida in the northern Wadden Sea. Bot. Mar. 2018, 61, 365–371. [Google Scholar] [CrossRef]
- Veiga, P.; Torres, A.C.; Rubal, M.; Troncoso, J.; Sousa-Pinto, I. The invasive kelp Undaria pinnatifida (Laminariales, Ochrophyta) along the north coast of Portugal: Distribution model versus field observations. Mar. Pollut. Bull. 2014, 84, 363–365. [Google Scholar] [CrossRef]
- South, P.M.; Floerl, O.; Forrest, B.M.; Thomsen, M.S. A review of three decades of research on the invasive kelp Undaria pinnatifida in Australasia: An assessment of its success, impacts and status as one of the world’s worst invaders. Mar. Environ. Res. 2017, 131, 243–257. [Google Scholar] [CrossRef]
- Piriz, M.; Casas, G. Occurence of Undaria pinnatifida in Golfo Nuevo, Argentina. Appl. Phycol. Forum 1994, 10, 4. [Google Scholar]
- Silva, P.S.; Woodfield, R.A.; Cohen, A.N.; Harris, L.H. First report of the Asian kelp Undaria pinnatifida in the northeastern Pacific Ocean. Biol. Invasions 2002, 4, 333–338. [Google Scholar] [CrossRef]
- Aguilar-Rosas, R.; Aguilar-Rosas, L.E.; Ávila-Serrano, G.; Marcos-Ramírez, R. First record of Undaria pinnatifida (Harvey) Suringar (Laminariales, Phaeophyta) on the Pacific coast of Mexico. Bot. Mar. 2004, 47, 255–258. [Google Scholar] [CrossRef]
- Raffo, M.P.; Eyras, M.C.; Iribarne, O.O. The invasion of Undaria pinnatifida to a Macrocystis pyrifera kelp in Patagonia (Argentina, south-west Atlantic). J. Mar. Biol. Assoc. 2009, 89, 1571–1580. [Google Scholar] [CrossRef]
- Arnold, M.; Teagle, H.; Brown, M.P.; Smale, D.A. The structure and diversity of epibiotic assemblages associated with the invasive kelp Undaria pinnatifida in comparison to native habitat-forming macroalgae on a subtidal temperate reef. Biol. Invasions 2016, 18, 661–676. [Google Scholar] [CrossRef]
- Suárez-Jiménez, R.; Hepburn, C.D.; Hyndes, G.A.; McLeod, R.J.; Taylor, R.B.; Hurd, C.L. The invasive kelp Undaria pinnatifida hosts an epifaunal assemblage similar to native seaweeds with comparable morphologies. Mar. Ecol. Prog. Ser. 2017, 582, 45–55. [Google Scholar] [CrossRef]
- Irigoyen, A.; Trobbiani, G.; Sgarlatta, M.; Raffo, M. Effects of the alien algae Undaria pinnatifida (Phaeophyceae, Laminariales) on the diversity and abundance of benthic macrofauna in Golfo Nuevo (Patagonia, Argentina): Potential implications for local food webs. Biol. Invasions 2011, 13, 1521–1532. [Google Scholar] [CrossRef]
- Casas, G.; Scrosati, R.; Luz Piriz, M. The invasive kelp Undaria pinnatifida (Phaeophyceae, Laminariales) reduces native seaweed diversity in Nuevo Gulf (Patagonia, Argentina). Biol. Invasions 2004, 6, 411–416. [Google Scholar] [CrossRef]
- Forrest, B.; Taylor, M. Assessing invasion impact: Survey design considerations and implications for management of an invasive marine plant. Biol. Invasions 2002, 4, 375–386. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species A Selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000; pp. 1–12. [Google Scholar]
- Gallardo, B. Europe’s top 10 invasive species: Relative importance of climatic, habitat and socio-economic factors. Ethol. Ecol. Evol. 2014, 26, 130–151. [Google Scholar] [CrossRef]
- James, K.; Shears, N.T. Proliferation of the invasive kelp Undaria pinnatifida at aquaculture sites promotes spread to coastal reefs. Mar. Biol. 2016, 163, 34. [Google Scholar] [CrossRef]
- Epstein, G.; Smale, D.A. Environmental and ecological factors influencing the spillover of the non-native kelp, Undaria pinnatifida, from marinas into natural rocky reef communities. Biol. Invasions 2017, 20, 1049–1072. [Google Scholar] [CrossRef] [PubMed]
- Santiago Caamaño, J.; Acuña Castroviejo, R.; Duran Neir, C. Aparición de Undaria pinnatifida en las costas de Galicia (España): Un nuevo caso en la problemática de introducción de especies foráneas. Inf. Tec. Invest. Sub. S.L. (Cis) 1990, 3, 1–45. [Google Scholar]
- Salinas, J.M.; Llera, E.M.; Fuertes, C. Nota sobre la presencia de Undaria pinnatifida (Harvey) Suringar (Laminariales, Phaeophyta) en Asturias (mar Cantábrico). Bol. Inst. Esp. Oceanogr. 1996, 12, 77–79. [Google Scholar]
- Cremades, J.; Freire, O.; Peteiro, C. Biología, distribución e integración del alga alóctona Undaria pinnatifida (Laminariales, Phaeophyta) en las comunidades bentónicas de las costas de Galicia (NW de la Península Ibérica). An. Jard. Bot. Madrid 2006, 63, 169–187. [Google Scholar] [CrossRef]
- Carreira-Flores, D.; Rubal, M.; Moreira, J.; Guerrero-Mesenguer, L.; Gomes, P.T.; Veiga, P. Recent changes on the abundance and distribution of non-indigenous macroalgae along the southwest coast of the Bay of Biscay. Aquat. Bot. 2023, 189, 103685. [Google Scholar] [CrossRef]
- Araújo, R.; Bárbara, I.; Tibaldo, M.; Berecibar, E.; Diaz Tapia, P.; Pereira, R.; Santos, R.; Sousa-Pinto, I. Checklist of benthic marine algae and cyanobacteria of northern Portugal. Bot. Mar. 2009, 52, 24–46. [Google Scholar] [CrossRef]
- Báez, J.C.; Olivero, J.; Peteiro, C.; Ferri-Yáñez, F.; Garcia-Soto, C.; Real, R. Macroenvironmental modelling of the current distribution of Undaria pinnatifida (Laminariales, Ochrophyta) in northern Iberia. Biol. Invasions 2010, 12, 2131–2139. [Google Scholar] [CrossRef]
- Russell, L.K.; Hepburn, C.D.; Hurd, C.L.; Stuart, M.D. The expanding range of Undaria pinnatifida in southern New Zealand: Distribution, dispersal mechanisms and the invasion of wave-exposed environments. Biol. Invasions 2008, 10, 103–115. [Google Scholar] [CrossRef]
- Farrell, P.; Fletcher, R.L. An investigation of dispersal of the introduced brown alga Undaria pinnatifida (Harvey) Suringar and its competition with some species on the manmade structures of Torquay Marina (Devon, UK). J. Exp. Mar. Biol. Ecol. 2006, 334, 236–243. [Google Scholar] [CrossRef]
- Bollen, M.; Pilditch, C.A.; Battershill, C.N.; Bischof, K. Salinity and temperature tolerance of the invasive alga Undaria pinnatifida and native New Zealand kelps: Implications for competition. Mar. Biol. 2016, 163, 194. [Google Scholar] [CrossRef]
- Peteiro, C.; Sánchez, N. Comparing salinity tolerance in early stages of the sporophytes of a non-indigenous kelp (Undaria pinnatifida) and a native kelp (Saccharina latissima). Russ. J. Mar. Biol. 2012, 38, 197–200. [Google Scholar] [CrossRef]
- Nanba, N.; Fujiwara, T.; Kuwano, K.; Ishikawa, Y.; Ogawa, H.; Kado, R. Effect of water flow velocity on growth and morphology of cultured Undaria pinnatifida sporophytes (Laminariales, Phaeophyceae) in Okirai Bay on the Sanriku coast, Northeast Japan. J. Appl. Phycol. 2011, 23, 1023–1030. [Google Scholar] [CrossRef]
- Morelissen, B.; Dudley, B.D.; Phillips, N.E. Recruitment of the invasive kelp Undaria pinnatifida does not always benefit from disturbance to native algal communities in low-intertidal habitats. Mar. Biol. 2016, 163, 241. [Google Scholar] [CrossRef]
- South, P.M.; Thomsen, M.S. The ecological role of invading Undaria pinnatifida: An experimental test of the driver–passenger models. Mar. Biol. 2016, 163, 175. [Google Scholar] [CrossRef]
- De Leij, R.; Epstein, G.; Brown, M.P.; Smale, D.A. The influence of native macroalgal canopies on the distribution and abundance of the non-native kelp Undaria pinnatifida in natural reef habitats. Mar. Biol. 2017, 164, 156. [Google Scholar] [CrossRef]
- Rubal, M.; Torres, C.A.; Veiga, P. Low diversity of intertidal canopy-forming macroalgae at urbanized areas along the North Portuguese Coast. Diversity 2020, 12, 211. [Google Scholar] [CrossRef]
- Hewitt, C.L.; Campbell, M.L.; McEnnulty, F.; Moore, K.M.; Murfet, N.B.; Robertson, B.; Schaffelke, B. Efficacy of physical removal of a marine pest: The introduced kelp Undaria pinnatifida in a Tasmanian marine reserve. Biol. Invasions 2005, 7, 251–263. [Google Scholar] [CrossRef]
- Forrest, B.M.; Hopkins, G.A. Population control to mitigate the spread of marine pests: Insights from management of the Asian kelp Undaria pinnatifida and colonial ascidian Didemnum vexillum. Manag. Biol. Invasions 2013, 4, 317–326. [Google Scholar] [CrossRef]
- Lemos, R.T.; Pires, H.O. The upwelling regime off the west Portuguese coast, 1941–2000. Int. J. Climatol. 2004, 24, 511–524. [Google Scholar] [CrossRef]
- Guzinski, J.; Ruggeri, P.; Ballenghien, M.; Mauger, S.; Jacquemin, B.; Jollivet, C.; Coudret, J.; Jaugeon, L.; Destombe, C.; Valero, M. Seascape Genomics of the Sugar Kelp Saccharina latissima along the North Eastern Atlantic Latitudinal gradient. Genes 2020, 11, 1503. [Google Scholar] [CrossRef] [PubMed]




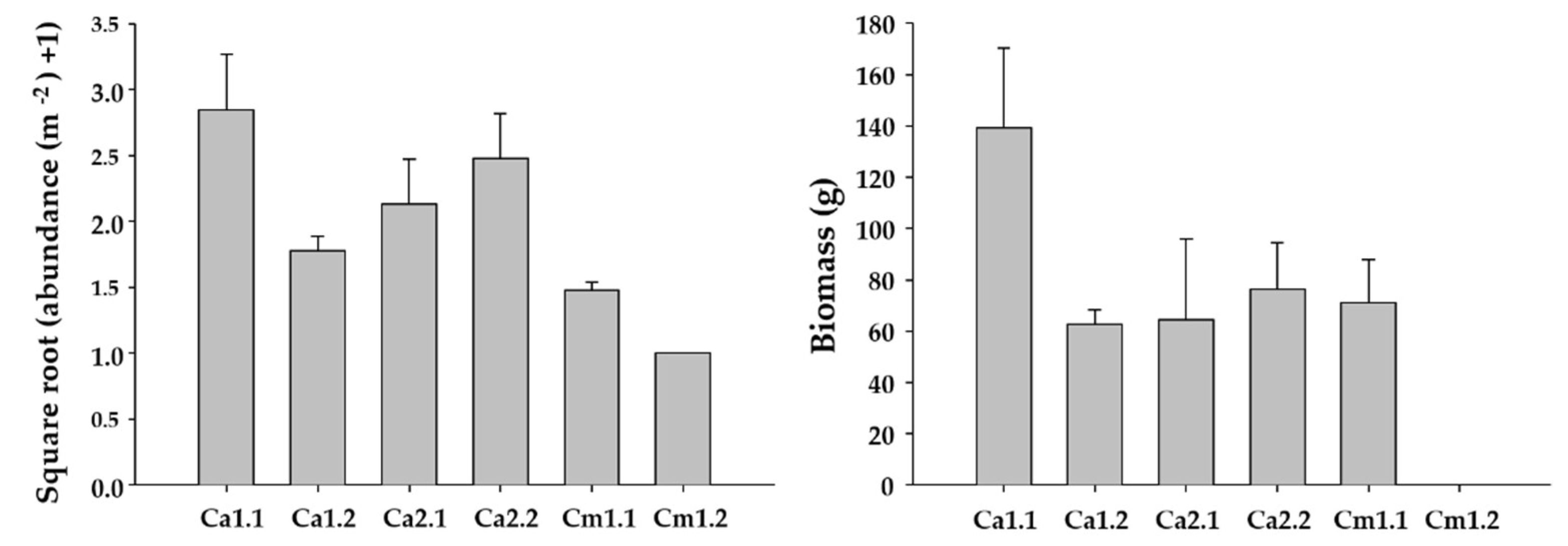

| Source | df | Salinity (psu) | Temperature (°C) | Dissolved Oxygen (mg L−1) | F-Versus | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MS | F | p | MS | F | p | MS | F | p | |||
| Ha | 1 | 3159.75 | 34.01 | 0.004 | 56.75 | 4.03 | 0.11 | 9.84 | 4.70 | 0.09 | Ma (Ha) |
| Ma (Ha) | 4 | 92.9 | 4.31 | 0.007 | 14.08 | 27.31 | 0.0001 | 2.09 | 12.93 | 0.0002 | Residual |
| Residual | 30 | 21.56 | 0.52 | 0.16 | |||||||
| Total | 35 | ||||||||||
| Cochran’s Test | C = 0.41 (n.s.) | C = 0.32 (n.s.) | C = 0.41 (n.s.) | ||||||||
| Transformation | None | None | None | ||||||||
| Source | df | Length (cm) | Biomass (g) | F-Versus | ||||
|---|---|---|---|---|---|---|---|---|
| MS | F | p | MS | F | p | |||
| Ma | 2 | 785.15 | 1.82 | 0.30 | 18.46 | 1.39 | 0.37 | Si (Ha) |
| Si (Ma) | 3 | 431.13 | 4.07 | 0.02 | 13.29 | 4.64 | 0.01 | Residual |
| Residual | 18 | 105.80 | 2.86 | |||||
| Total | 23 | 23 | ||||||
| Cochran’s Test | C = 0.52 (Not Significant) | C = 0.48 (Not Significant) | ||||||
| Source | df | Density (Individuals m−2) | Biomass (g m−2) | F-Versus | ||||
|---|---|---|---|---|---|---|---|---|
| MS | F | p | MS | F | p | |||
| Lo | 2 | 3.81 | 3.05 | 0.19 | 10,708.61 | 1.16 | 0.42 | Si (Ar) |
| Si (Lo) | 3 | 1.25 | 3.55 | 0.03 | 9230.82 | 4.28 | 0.01 | Residual |
| Residual | 24 | 0.35 | 2155.40 | |||||
| Total | 29 | |||||||
| Cochran’s Test | C = 0.41 (Not Significant) | C = 0.38 (Not Significant) | ||||||
| Transformation | Sqrt(X + 1) | None | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubal, M.; Fernández-Gutiérrez, J.; Carreira-Flores, D.; Gomes, P.T.; Veiga, P. Current Distribution of the Invasive Kelp Undaria pinnatifida (Harvey) Suringar, 1873 Along Artificial and Natural Habitats in North Portugal—Impacts and Mitigation Initiatives. Plants 2025, 14, 658. https://doi.org/10.3390/plants14050658
Rubal M, Fernández-Gutiérrez J, Carreira-Flores D, Gomes PT, Veiga P. Current Distribution of the Invasive Kelp Undaria pinnatifida (Harvey) Suringar, 1873 Along Artificial and Natural Habitats in North Portugal—Impacts and Mitigation Initiatives. Plants. 2025; 14(5):658. https://doi.org/10.3390/plants14050658
Chicago/Turabian StyleRubal, Marcos, Jesús Fernández-Gutiérrez, Diego Carreira-Flores, Pedro T. Gomes, and Puri Veiga. 2025. "Current Distribution of the Invasive Kelp Undaria pinnatifida (Harvey) Suringar, 1873 Along Artificial and Natural Habitats in North Portugal—Impacts and Mitigation Initiatives" Plants 14, no. 5: 658. https://doi.org/10.3390/plants14050658
APA StyleRubal, M., Fernández-Gutiérrez, J., Carreira-Flores, D., Gomes, P. T., & Veiga, P. (2025). Current Distribution of the Invasive Kelp Undaria pinnatifida (Harvey) Suringar, 1873 Along Artificial and Natural Habitats in North Portugal—Impacts and Mitigation Initiatives. Plants, 14(5), 658. https://doi.org/10.3390/plants14050658