
Fusarium Head Blight in Barley from Subtropical Southern Brazil: Associated Fusarium Species and Grain Contamination Levels of Deoxynivalenol and Nivalenol

 , , ,
, , ,
Abstract
1. Introduction
2. Material and Methods
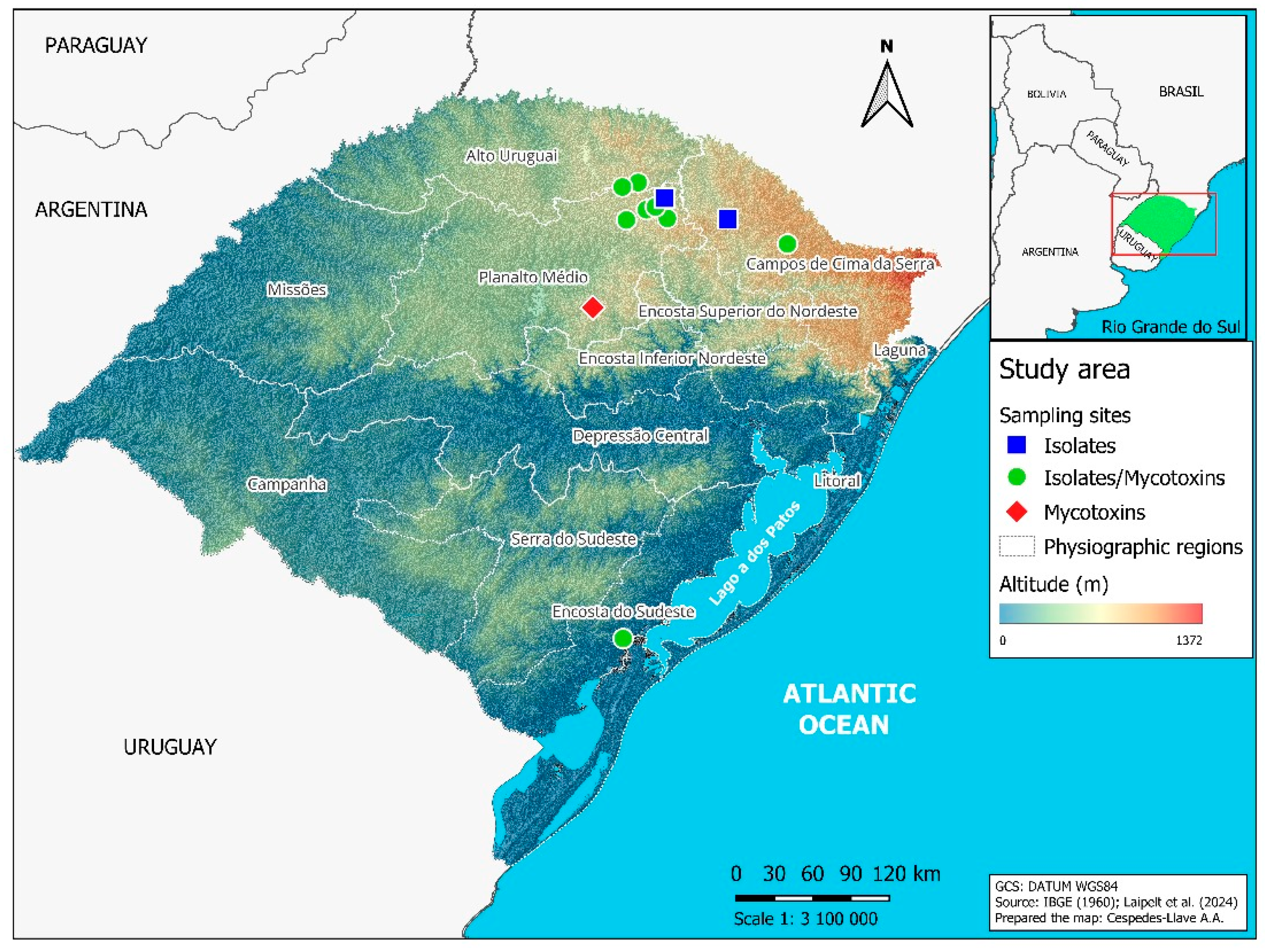
2.1. Barley Sampling
2.2. Pathogen Isolation from Symptomatic Barley Spikes
2.3. DNA Extraction, Polymerase Chain Reaction (PCR) and Sequencing
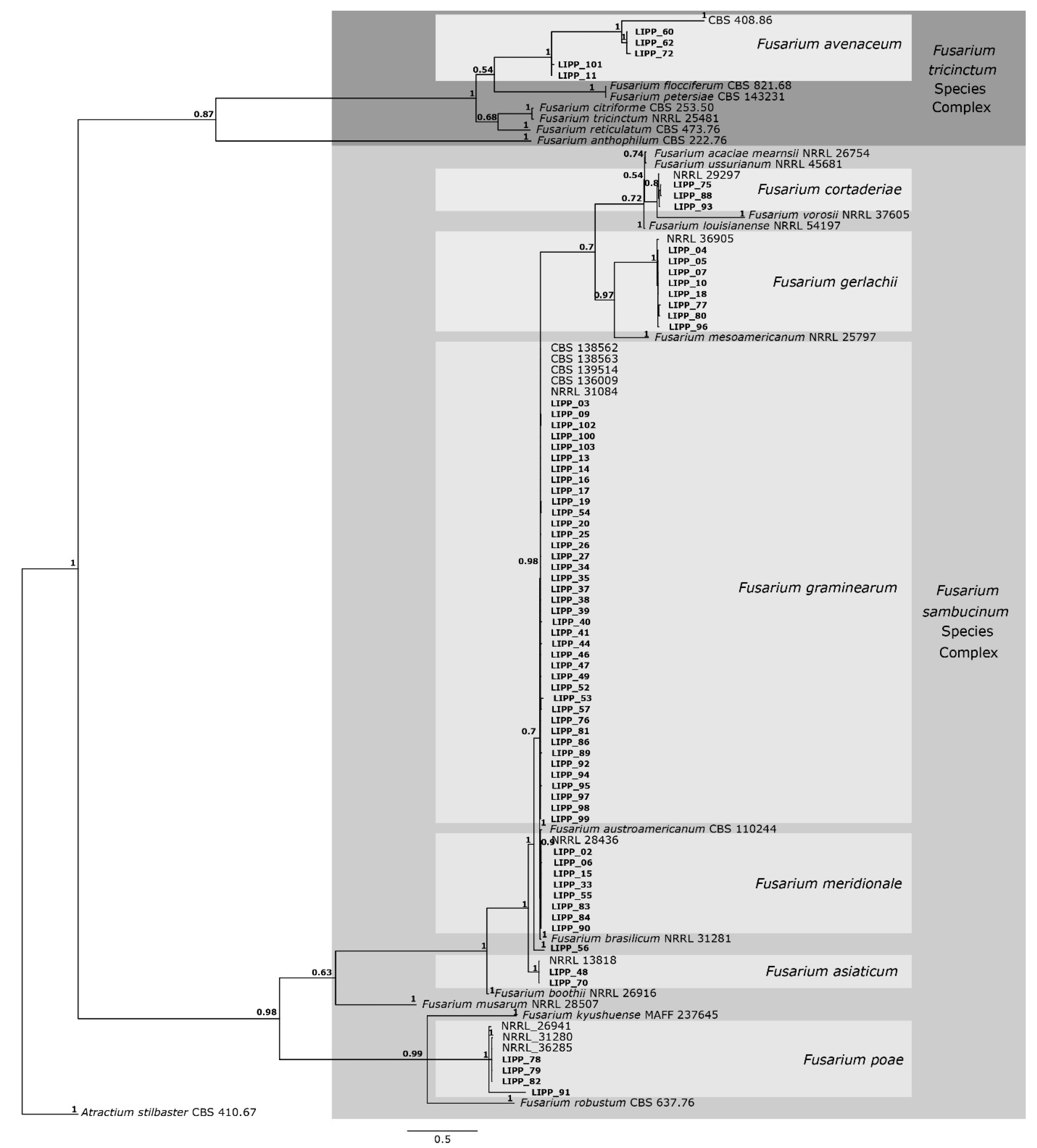
2.4. Phylogenetic Analysis
2.5. Mycotoxin Quantification
2.5.1. Deoxynivalenol Analysis
2.5.2. Nivalenol Analysis
2.5.3. HPLC-MS/MS Analysis Parameters
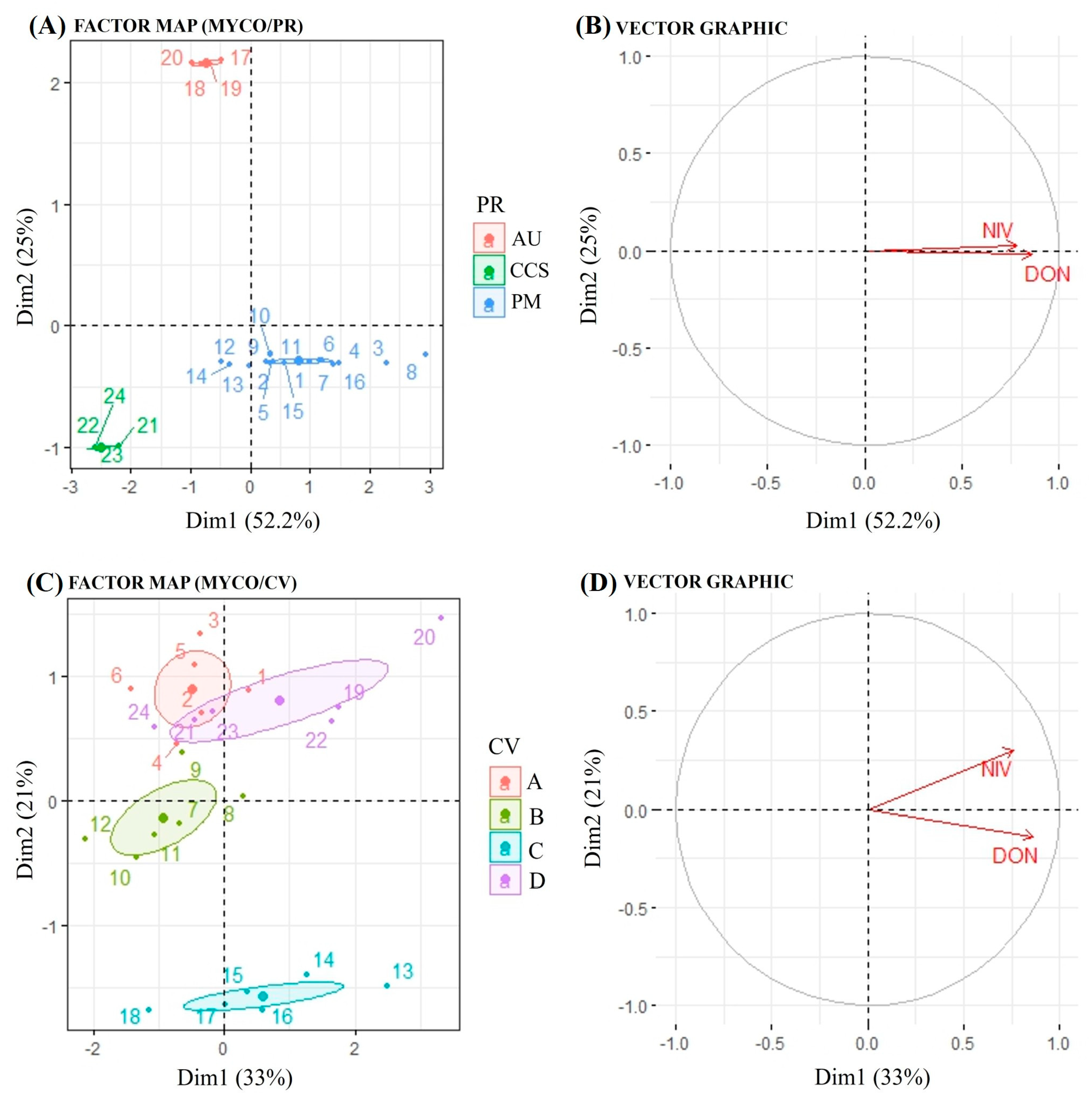
2.6. Data Analysis
3. Results
3.1. Fusarium Species Identification
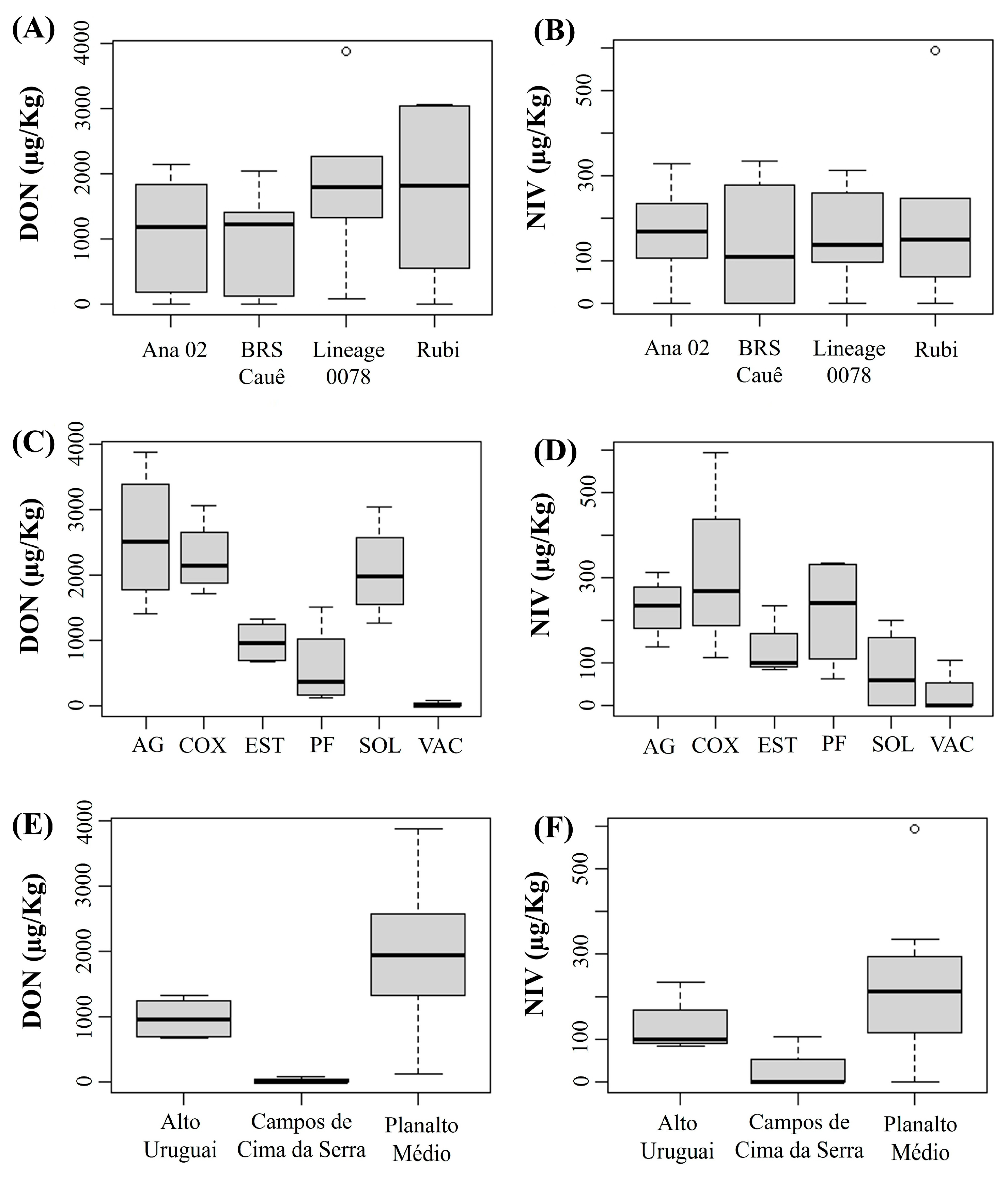
3.2. Detection and Quantification of DON and NIV
Concentrations of DON and NIV by Cultivar and Physiographic Region
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Conab. Companhia Nacional de Abastecimento. Acompanhamento da safra brasileira—Grãos: Décimo Segundo Levantamento, Setembro 2023—Safra 2022/2023: Brasília: Companhia Nacional de Abastecimento. 2023. Available online: https://www.conab.gov.br/info-agro/safras/graos/boletim-da-safra-de-graos (accessed on 3 September 2023).
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef]
- Perincherry, L.; Lalak-Kańczugowska, J.; Stępień, Ł. Fusarium-produced mycotoxins in plant-pathogen interactions. Toxins 2019, 11, 664. [Google Scholar] [CrossRef]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium head blight from a microbiome perspective. Front. Microbiol. 2021, 12, 628373. [Google Scholar] [CrossRef]
- Pestka, J.J. Toxicological mechanisms and potential health effects of deoxynivalenol and nivalenol. World Mycotoxin. J. 2010, 3, 323–347. [Google Scholar] [CrossRef]
- Cheat, S.; Gerez, J.; Cognié, J.; Alassane-Kpembi, I.; Bracarense, A.; Raymond-Letron, I.; Oswald, I.; Kolf-Clauw, M. Nivalenol has a greater impact than deoxynivalenol on pig jejunum mucosa in vitro on explants and in vivo on intestinal loops. Toxins 2015, 7, 1945–1961. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Puel, O.; Pinton, P.; Cossalter, A.; Chou, T.; Oswald, I. Co-exposure to low doses of the food contaminants deoxynivalenol and nivalenol has synergistic inflammatory effect on intestinal explants. Arch. Toxicol. 2017, 91, 2677–2687. [Google Scholar] [CrossRef]
- Brasil Ministério da Saúde. Instrução Normativa n° 160, de 1° de Julho de 2022; Diário Oficial da União: Brasília, Brazil, 2022. [Google Scholar]
- Cheli, F.; Battaglia, D.; Gallo, R.; Dell’orto, V. EU legislation on cereal safety: An update with a focus on mycotoxins. Food Control 2014, 37, 315–325. [Google Scholar] [CrossRef]
- Cowger, C.; Ward, T.J.; Nilsson, K.; Arellano, C.; McCormick, S.P.; Busman, M. Regional and field-specific differences in Fusarium species and mycotoxins associated with blighted North Carolina wheat. Int. J. Food Microbiol. 2020, 323, 108594. [Google Scholar] [CrossRef]
- Johns, L.E.; Bebber, D.P.; Gurr, S.J.; Brown, N.A. Emerging health threat and cost of Fusarium mycotoxins in European wheat. Nat. Food 2022, 3, 1014–1019. [Google Scholar] [CrossRef]
- Infantino, A.; Belocchi, A.; Quaranta, F.; Reverberi, M.; Beccaccioli, M.; Lombardi, D.; Vitale, M. Effects of climate change on the distribution of Fusarium spp. in Italy. Sci. Total Environ. 2023, 882, 163640. [Google Scholar] [CrossRef]
- Boutigny, A.L.; Ward, T.J.; Ballois, N.; Iancu, G.; Ioos, R. Diversity of the Fusarium graminearum species complex on French cereals. Eur. J. Plant Pathol. 2014, 138, 133–148. [Google Scholar] [CrossRef]
- Islam, M.N.; Banik, M.; Sura, S.; Tucker, J.R.; Wang, X. Implications of crop rotation and fungicide on Fusarium and mycotoxin spectra in Manitoba barley, 2017–2019. Toxins 2022, 14, 463. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, T.; Xing, Y.; Dai, C.; Deji, C.; Yang, X.; Wu, X.; Mduduzi, P.M.; Ademola, O.O.; Shi, J.; et al. Characterization of Fusarium species causing head blight of highland barley (qingke) in Tibet, China. Int. J. Food Microbiol. 2024, 416, 110728. [Google Scholar]
- Pereira, C.B.; Ward, T.J.; Del Ponte, E.M.; Mara Moreira, G.; Busman, M.; McCormick, S.; Feksa, H.; Almeida, J.; Tessmann, D.J. Five-year survey uncovers extensive diversity and temporal fluctuations among Fusarium head blight pathogens of wheat and barley in Brazil. Plant Pathol. 2021, 70, 426–435. [Google Scholar] [CrossRef]
- Hoffmann, G.R.; Arend, L.M.; Silveira, J.C.B.D.; Bellomo, H.R.; Nunes, J.L.M. Rio Grande do Sul. Aspectos da Geografia; Martins Livreiro: Porto Alegre, Brazil, 1997. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Reeb, V.; Lutzoni, F.; Roux, C. Contribution of RPB2 to multilocus phylogenetic studies of the euascomycetes (Pezizomycotina, Fungi) with special emphasis on the lichen-forming Acarosporaceae and evolution of polyspory. Mol. Phylogenet Evol. 2004, 32, 1036–1060. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. MBE 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2; Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Simões, C.T.; Vidal, J.K.; Da Silva, C.R.; Sarturi, J.A.; Laber, I.F.; Madalosso, T.; Mallmann, C.A. A two-year study on the occurrence and concentration of mycotoxins in corn varieties with different endosperm textures. J. Sci. Food Agric. 2023, 103, 7199–7206. [Google Scholar] [CrossRef]
- Mallmann, C.A.; Tyska, D.; Almeida, C.A.A.; Oliveira, M.; Gressler, L.T. Mycotoxicological monitoring of breakfast and infant cereals marketed in Brazil. Int. J. Food Microbiol. 2020, 331, 108628. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Gilbert, J.; Fernando, W.G.D. Epidemiology and biological control of Gibberella zeae/Fusarium graminearum. Can. J. Plant Pathol. 2004, 26, 464–472. [Google Scholar] [CrossRef]
- Reverberi, M.; Ricelli, A.; Zjalic, S.; Fabbri, A.A.; Fanelli, C. Natural functions of mycotoxins and control of their biosynthesis in fungi. Appl. Microbiol. Biotechnol. 2010, 87, 899–911. [Google Scholar] [CrossRef]
- Pagès, J. Analyse factorielle de donnees mixtes: Principe et exemple d’application. Rev. Statist. Appl. 2004, 52, 93–111. [Google Scholar]
- Astolfi, P.; Santos, J.; Schneider, L.; Gomes, L.B.; Silva, C.N.; Tessmann, D.J. Molecular survey of trichothecene genotypes of Fusarium graminearum species complex from barley in Southern Brazil. Int. J. Food Microbiol. 2011, 148, 197–201. [Google Scholar] [CrossRef]
- Del Ponte, E.M.; Spolti, P.; Ward, T.J.; Gomes, L.B.; Nicolli, C.P.; Kuhnem, P.R.; Silva, C.N.; Tessmann, D.J. Regional and field-specific factors affect the composition of Fusarium head blight pathogens in subtropical no-till wheat agroecosystem of Brazil. Phytopathology 2015, 105, 246–254. [Google Scholar] [CrossRef]
- Alaniz-Zanon, M.S.; Bossa, M.; Rosales Cavaglieri, L.A.; Palazzini, J.M.; Sulyok, M.; Chulze, S.N.; Chiotta, M.L. Fusarium head blight in Argentina, a profile of produced mycotoxins and a biocontrol strategy in barley during micro-malting process. Toxins 2025, 17, 39. [Google Scholar] [CrossRef]
- Khanal, R.; Hudson, K.; Foster, A.; Wang, X.; Brauer, E.K.; Witte, T.E.; Overy, D.P. Pathogenicity and metabolomic characterization of Fusarium graminearum and Fusarium poae challenge in barley under controlled conditions. J. Fungi. 2024, 10, 670. [Google Scholar] [CrossRef]
- Martínez, M.; Arata, A.F.; Dinolfo, M.I.; Arrigoni, A.; Lázaro, L.; Stenglein, S.A. Effect of warm nights on Fusarium graminearum-bread wheat pathosystem: Impact on grain quality and rheological properties in two contrasting commercial genotypes. Field Crops Res. 2024, 314, 109432. [Google Scholar] [CrossRef]
- Mallmann, C.A.; Dilkin, P.; Mallmann, A.O.; Oliveira, M.S.; Adaniya, Z.N.C.; Tonini, C. Prevalence and levels of deoxynivalenol and zearalenone in commercial barley and wheat grain produced in Southern Brazil: An eight-year (2008 to 2015) summary. Trop. Plant Pathol. 2017, 42, 146–152. [Google Scholar] [CrossRef]
- Ibáñez-Vea, M.; Lizarraga, E.; González-Peñas, E.; de Cerain, A.L. Co-occurrence of type-A and type-B trichothecenes in barley from a northern region of Spain. Food Control 2012, 25, 81–88. [Google Scholar] [CrossRef]
- Iwase, C.H.; Piacentini, K.C.; Giomo, P.P.; Čumová, M.; Wawroszová, S.; Běláková, S.; Minella, E.; Rocha, L.O. Characterization of the Fusarium sambucinum species complex and detection of multiple mycotoxins in Brazilian barley samples. Food Res. Int. 2020, 136, 109336. [Google Scholar] [CrossRef]
- Bolechová, M.; Benesová, K.; Beláková, S.; Cáslavsky, J.; Pospíchalová, M.; Mikulíková, R. Determination of seventeen mycotoxins in barley and mal in the Czech Republic. Food Control 2015, 47, 108–113. [Google Scholar] [CrossRef]
- Llorens, A.; Mateo, R.; Hinojo, M.J.; Valle-Algarra, F.M.; Jiménez, M. Influence of environmental factors on the biosynthesis of type B trichothecenes by isolates of Fusarium spp. from Spanish crops. Int. J. Food Microbiol. 2004, 94, 43–54. [Google Scholar] [CrossRef]
- Hope, R.; Aldred, D.; Magan, N. Comparison of environmental profiles for growth and deoxynivalenol production by Fusarium culmorum and F. graminearum on wheat grain. Lett. Appl. Microbiol. 2005, 40, 295–300. [Google Scholar] [CrossRef]
- Rybecky, A.I.; Chulze, S.N.; Chiotta, M.L. Effect of water activity and temperature on growth and trichothecene production by Fusarium meridionale. Int. J. Food Microbiol. 2018, 285, 69–73. [Google Scholar] [CrossRef]
- Palladino, C.; Puigvert, F.; Muela, A.; Taborda, B.; Pérez, C.A.; Pérez-Parada, A.; Pareja, L. Evaluation of Fusarium mycotoxins and fungicide residues in barley grain produced in Uruguay. J. Agric. Food Res. 2021, 3, 100092. [Google Scholar] [CrossRef]
- Iwase, C.H.T.; Piacentini, K.C.; Silva, N.C.C.; Rebellato, A.P.; Rocha, L.O. Deoxynivalenol and zearalenone in Brazilian barley destined for brewing. Food Addit. Contam. Part B Surveill. 2023, 16, 86–92. [Google Scholar] [CrossRef]
- Munkvold, G.P.; Proctor, R.H.; Moretti, A. Mycotoxin production in Fusarium according to contemporary species concepts. Annu. Rev. Phytopathol. 2021, 59, 373–402. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiographic Regions | Municipality | Field 1 | Geographic Coordinates | Altitude (m) | Barley Genotype 2 | Sample | ||
|---|---|---|---|---|---|---|---|---|
| Longitude | Latitude | Spike | Grains | |||||
| Alto Uruguai | Estação | AMBEV | −52.291794 | −27.898500 | 763 | A/B/C/D | × | × |
| Commercial field | −52.291794 | −27.898500 | 763 | B | × | × | ||
| Ipiranga do Sul | Commercial field I | −52.450942 | −27.945414 | 650 | B | × | × | |
| Commercial field II | −52.450942 | −27.945414 | 650 | B | × | × | ||
| Encosta do Sudeste | Capão do Leão | UFPel Area I | −52.419113 | −31.798330 | 20 | B | × | × |
| UFPel Area II | −52.505765 | −31.800805 | 20 | B/E | × | × | ||
| Planalto Médio | Água Santa | AMBEV | −52.045521 | −28.191851 | 794 | A/B/C/D | × | × |
| Coxilha | AMBEV | −52.215816 | −28.093141 | 627 | A/B/C/D | × | × | |
| Commercial field | −52.24083 | −28.108747 | D | × | ns | |||
| Passo Fundo | AMBEV | −52.392713 | −28.218805 | 661 | A/B/C/D | × | × | |
| Soledade | AMBEV | −52.677361 | −28.969638 | 697 | A/B/C/D | ns | × | |
| Tapejara | Commercial field | −52.065267 | −28.032335 | B | × | ns | ||
| Vila Lângaro | Commercial field | −52.123922 | −28.107910 | 643 | B | × | × | |
| Campos de Cima da Serra | Lagoa Vermelha | Commercial field | −51.5262 | −28.2107 | B | × | ns | |
| Vacaria | AMBEV | −51.016944 | −28.423889 | 962 | A/B/C/D | × | × | |
| Commercial field I | −51.028333 | −28.124722 | 962 | A/D | × | × | ||
| Commercial field II | −50.930311 | −28.510093 | 962 | B | × | × | ||
| Bioclimatic Variables | Descriptions | Unit |
|---|---|---|
| BIO1 | Annual mean temperature | °C |
| BIO2 | Mean diurnal range (mean of monthly (maximum temperature − minimum temperature)) | °C |
| BIO3 | Isothermality (BIO2/BIO7) (∗100) | |
| BIO4 | Temperature seasonality (standard deviation of temperatures ∗ 100) | |
| BIO5 | Maximum temperature of warmest month | °C |
| BIO6 | Minimum temperature of coldest month | °C |
| BIO7 | Annual temperature range (BIO5-BIO6) | °C |
| BIO8 | Mean temperature of wettest quarter | °C |
| BIO9 | Mean temperature of driest quarter | °C |
| BIO10 | Mean temperature of warmest quarter | °C |
| BIO11 | Mean temperature of coldest quarter | °C |
| BIO12 | Annual preciptation | mm |
| BIO13 | Precipitation of wettest month | mm |
| BIO14 | Precipitation of driest month | mm |
| BIO15 | Precipitation seasonality (Coefficient of variation) | mm |
| BIO16 | Precipitation of wettest quarter | mm |
| BIO17 | Precipitation of driest quarter | mm |
| BIO18 | Precipitation of warmest quarter | mm |
| BIO19 | Precipitation of coldest quarter | mm |
| Fusarium Species | Isolates | Municipalities | Physiographic Region | Field |
|---|---|---|---|---|
| Fusarium asiaticum | LIPP 48 | Capão do Leão | Encosta do Sudeste | UFPel I |
| LIPP 70 | Capão do Leão | Encosta do Sudeste | UFPel II | |
| Fusarium avenaceum | LIPP 60, LIPP 62 and LIPP 72 | Capão do Leão | Encosta do Sudeste | UFPel II |
| LIPP 11 | Ipiranga do Sul | Alto Uruguai | Commercial field II | |
| LIPP 101 | Lagoa Vermelha | Campos de Cima da Serra | Commercial field | |
| Fusarium cortaderiae | LIPP 75 | Estação | Alto Uruguai | AMBEV |
| LIPP 88 | Vacaria | Campos de Cima da Serra | AMBEV | |
| LIPP 93 | Coxilha | Planalto Médio | Commercial field | |
| Fusarium gerlachii | LIPP 77 and LIPP 80 | Estação | Alto Uruguai | AMBEV |
| LIPP 96 | Lagoa Vermelha | Campos de Cima da Serra | Commercial field | |
| LIPP 18 | Tapejara | Planalto Médio | Commercial field | |
| LIPP 04, LIPP 05, LIPP 07 and LIPP 10 | Passo Fundo | Planalto Médio | AMBEV | |
| Fusarium graminearum s.str. | LIPP 46, LIPP 47, LIPP 53, LIPP 95, LIPP 99, LIPP 100 and LIPP 102 | Água Santa | Planalto Médio | AMBEV |
| LIPP 25, LIPP 26, LIPP 27, LIPP 49 and LIPP 52 | Capão do Leão | Encosta do Sudeste | UFPel I | |
| LIPP 37, LIPP 38, LIPP 39, LIPP 40, LIPP 86, and LIPP92 | Coxilha | Planalto Médio | AMBEV | |
| LIPP 41, LIPP 44, and LIPP54 | Coxilha | Planalto Médio | Commercial field | |
| LIPP 16, LIPP 34, LIPP 76, LIPP 94 and LIPP 103 | Estação | Alto Uruguai | AMBEV | |
| LIPP 13 and LIPP 14 | Ipiranga do Sul | Alto Uruguai | Commercial field I | |
| LIPP 03 and LIPP 09 | Passo Fundo | Planalto Médio | AMBEV | |
| LIPP 17 and LIPP 35 | Tapejara | Planalto Médio | Commercial field | |
| LIPP 81 and LIPP 89 | Vacaria | Campos de Cima da Serra | AMBEV | |
| LIPP 57, LIPP 97 and LIPP 98 | Vacaria | Campos de Cima da Serra | Commercial field I | |
| LIPP 19 and LIPP 20 | Vila Lângaro | Planalto Médio | Commercial field * | |
| Fusarium meridionale | LIPP 02, LIPP 5 and LIPP 6 | Passo Fundo | Planalto Médio | AMBEV |
| LIPP 33 | Vacaria | Campos de Cima da Serra | Commercial field II | |
| LIPP 83, LIPP 84 and LIPP 90 | Vacaria | Campos de Cima da Serra | AMBEV | |
| LIPP 15 | Estação | Alto Uruguai | AMBEV | |
| Fusarium poae | LIPP 78 and LIPP 79 | Estação | Alto Uruguai | Commercial field |
| LIPP 82 | Vacaria | Campos de Cima da Serra | AMBEV | |
| LIPP 91 | Vacaria | Campos de Cima da Serra | Commercial field I | |
| Fusarium sp. | LIPP 56 | Vacaria | Campos de Cima da Serra | Commercial field II |
| Physiographic Regions | Municipality | Cultivar | Mycotoxins (µg/Kg) | |
|---|---|---|---|---|
| DON | NIV | |||
| Alto Uruguai | Estação (AMBEV 1) | ‘Ana 02’ | 671 | 236 |
| ‘BRS Cauê’ | 1170 | 82.7 | ||
| Lineage 0078 | 1330 | 95.7 | ||
| ‘ABI Rubi’ | 727 | 102 | ||
| Estação (Commercial field 2) | ‘BRS Cauê’ | 381 | 327 | |
| Ipiranga do Sul (Commercial field I) | ‘BRS Cauê’ | 973 | 100 | |
| Ipiranga do Sul (Commercial field II) | ‘BRS Cauê’ | 314 | 166 | |
| Encosta do Sudeste | Capão do Leão (UFPEL 3 Área I) | ‘BRS Cauê’(+F *) | 5930 | 825 |
| ‘BRS Cauê’(−F *) | 7620 | 1540 | ||
| ‘BRS Cauê’(+F) | 4410 | 1560 | ||
| ‘BRS Cauê’(−F) | 10,200 | 1630 | ||
| Capão do Leão (UFPEL Área II) | ‘BRS Cauê’(+F) | 0 | 0 | |
| ‘BRS Cauê’(+F *) | 0 | 135 | ||
| ‘BRS Cauê’(−F *) | 0 | 114 | ||
| ‘BRS Cauê’(−F) | 0 | 0 | ||
| ‘BRS Brau’(+F) | 0 | 0 | ||
| ‘BRS Brau’(+F *) | 0 | 0 | ||
| ‘BRS Brau’(−F *) | 0 | 0 | ||
| ‘BRS Brau’(−F) | 0 | 0 | ||
| Planalto Médio | Água Santa (AMBEV) | ‘Ana 02’ | 2140 | 222 |
| ‘BRS Cauê’ | 1400 | 139 | ||
| Lineage 0078 | 3880 | 313 | ||
| ‘ABI Rubi’ | 2890 | 246 | ||
| Coxilha (AMBEV) | ‘Ana 02’ | 1710 | 113 | |
| ‘BRS Cauê’ | 2040 | 278 | ||
| Lineage 0078 | 2260 | 260 | ||
| ‘ABI Rubi’ | 3060 | 595 | ||
| Passo Fundo (AMBEV) | ‘Ana 02’ | 184 | 327 | |
| ‘BRS Cauê’ | 126 | 335 | ||
| Lineage 0078 | 1500 | 154 | ||
| ‘ABI Rubi’ | 558 | 62.7 | ||
| Soledade (AMBEV) | ‘Ana 02’ | 1840 | 0 | |
| ‘BRS Cauê’ | 1270 | 0 | ||
| Lineage 0078 | 2110 | 119 | ||
| ‘ABI Rubi’ | 3030 | 201 | ||
| Vila Lângaro (Commercial field) | ‘BRS Cauê’ | 2710 | 334 | |
| Campos de Cima da Serra | Vacaria (AMBEV) | ‘Ana 02’ | 0 | 106 |
| ‘BRS Cauê’ | 0 | 0 | ||
| Lineage 0078 | 73.7 | 0 | ||
| ‘ABI Rubi’ | 0 | 0 | ||
| Vacaria (Commercial field I) | ‘Ana 02’ | 1960 | 0 | |
| ‘ABI Rubi’ | 188 | 0 | ||
| Vacaria (Commercial field II) | ‘BRS Cauê’ | 425 | 129 | |
| Mycotoxin | GLM | Model | Family | AIC | VIF |
|---|---|---|---|---|---|
| DON | DON~BIO4 + BIO15 | DON~ + BIO4 * + BIO15 | Gaussian | 591.6724 | 1.08; 1.08 |
| NIV | NIV~BIO4 + BIO15 | NIV~ + BIO4 + BIO15 . | Gaussian | 465.2927 | 1.08; 1.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furtado, E.B.; Guatimosim, E.; de Barros, D.R.; Mallmann, C.A.; Araujo Filho, J.V.d.; Martins, S.d.O.; Tessmann, D.J.; Rombaldi, C.V.; Schlösser, L.M.d.L.; Favaretto, A.; et al. Fusarium Head Blight in Barley from Subtropical Southern Brazil: Associated Fusarium Species and Grain Contamination Levels of Deoxynivalenol and Nivalenol. Plants 2025, 14, 2327. https://doi.org/10.3390/plants14152327
Furtado EB, Guatimosim E, de Barros DR, Mallmann CA, Araujo Filho JVd, Martins SdO, Tessmann DJ, Rombaldi CV, Schlösser LMdL, Favaretto A, et al. Fusarium Head Blight in Barley from Subtropical Southern Brazil: Associated Fusarium Species and Grain Contamination Levels of Deoxynivalenol and Nivalenol. Plants. 2025; 14(15):2327. https://doi.org/10.3390/plants14152327
Chicago/Turabian StyleFurtado, Emanueli Bizarro, Eduardo Guatimosim, Danielle Ribeiro de Barros, Carlos Augusto Mallmann, Jeronimo Vieira de Araujo Filho, Sabrina de Oliveira Martins, Dauri José Tessmann, Cesar Valmor Rombaldi, Luara Medianeira de Lima Schlösser, Adriana Favaretto, and et al. 2025. "Fusarium Head Blight in Barley from Subtropical Southern Brazil: Associated Fusarium Species and Grain Contamination Levels of Deoxynivalenol and Nivalenol" Plants 14, no. 15: 2327. https://doi.org/10.3390/plants14152327
APA StyleFurtado, E. B., Guatimosim, E., de Barros, D. R., Mallmann, C. A., Araujo Filho, J. V. d., Martins, S. d. O., Tessmann, D. J., Rombaldi, C. V., Schlösser, L. M. d. L., Favaretto, A., & Dallagnol, L. J. (2025). Fusarium Head Blight in Barley from Subtropical Southern Brazil: Associated Fusarium Species and Grain Contamination Levels of Deoxynivalenol and Nivalenol. Plants, 14(15), 2327. https://doi.org/10.3390/plants14152327