


Origanum majorana Extracts: A Preliminary Comparative Study on Phytochemical Profiles and Bioactive Properties of Valuable Fraction and By-Product

 ,
,  , , ,
, , ,  ,
,  ,
, 
Abstract
1. Introduction
2. Results
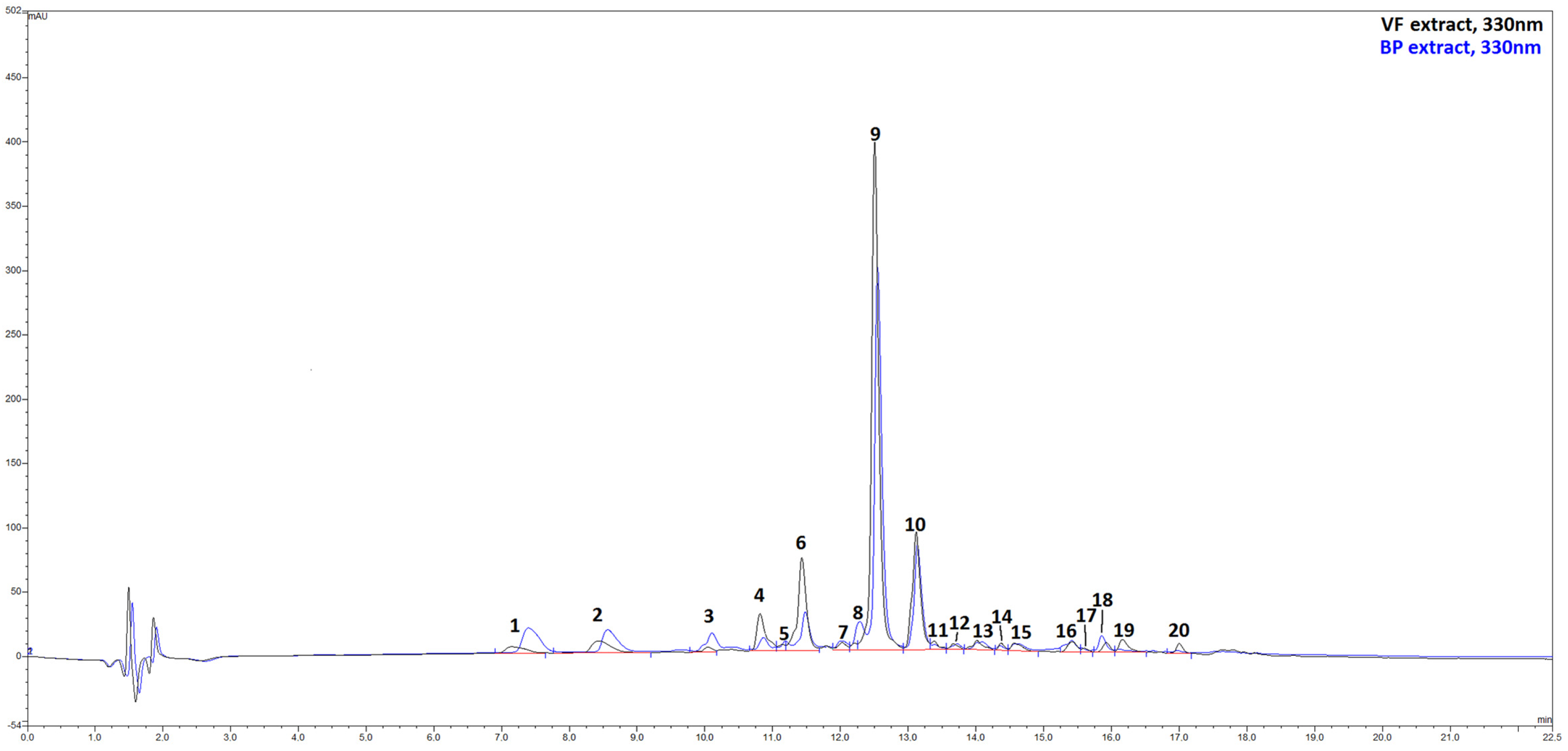
2.1. Extraction Yield and Phytochemical Analyses by HPLC-ESI-MS of VF and BP from O. majorana
2.2. In Vitro Cell-Free Antioxidant Properties of VF and BP from O. majorana
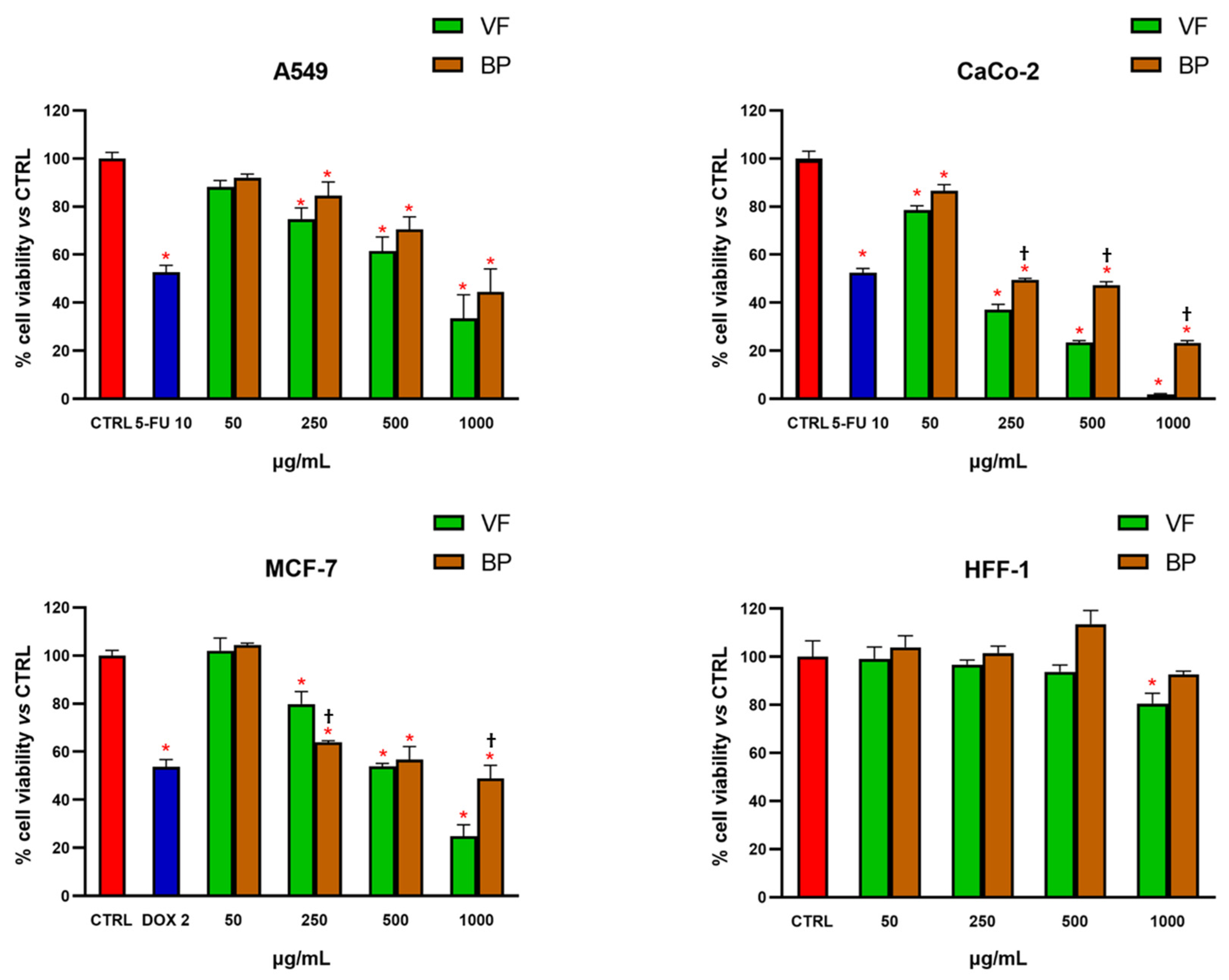
2.3. MTT Test on Normal and Cancer Cells
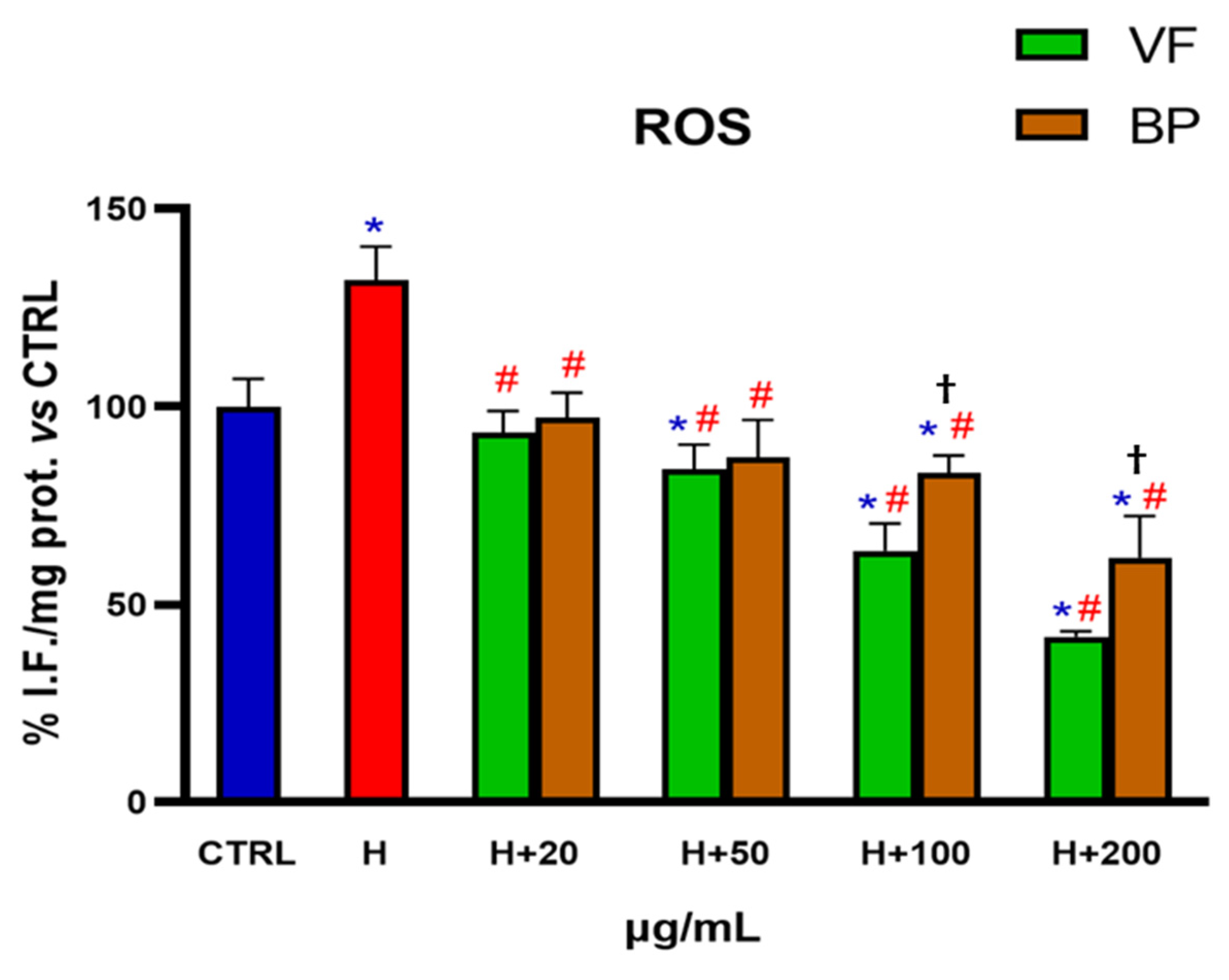
2.4. Antioxidant Activity in Cells
2.4.1. Determination of ROS Levels in Human AC16 Cardiomyocytes
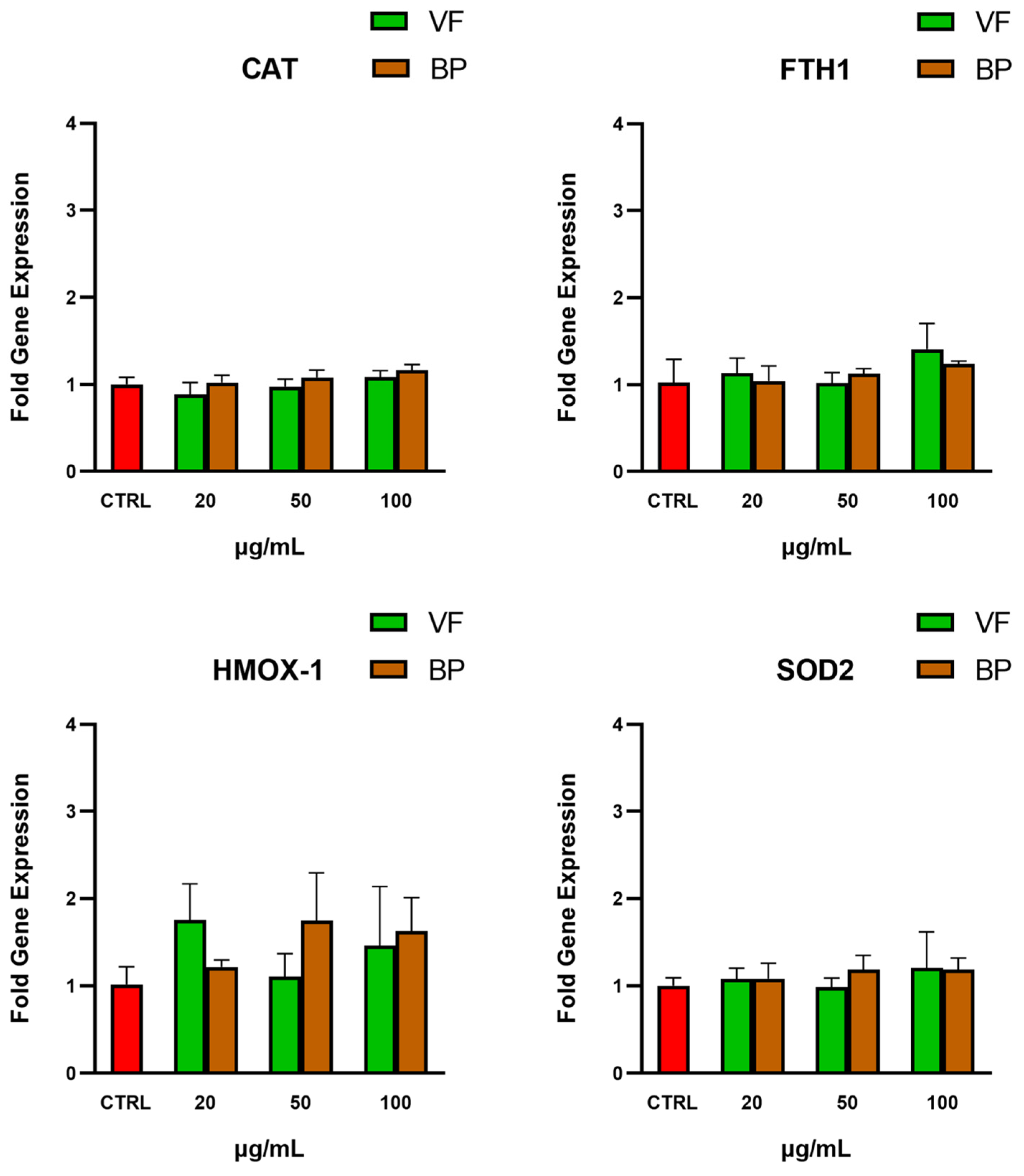
2.4.2. Gene Expression of Antioxidant Enzymes in Human AC16 Cardiomyocytes
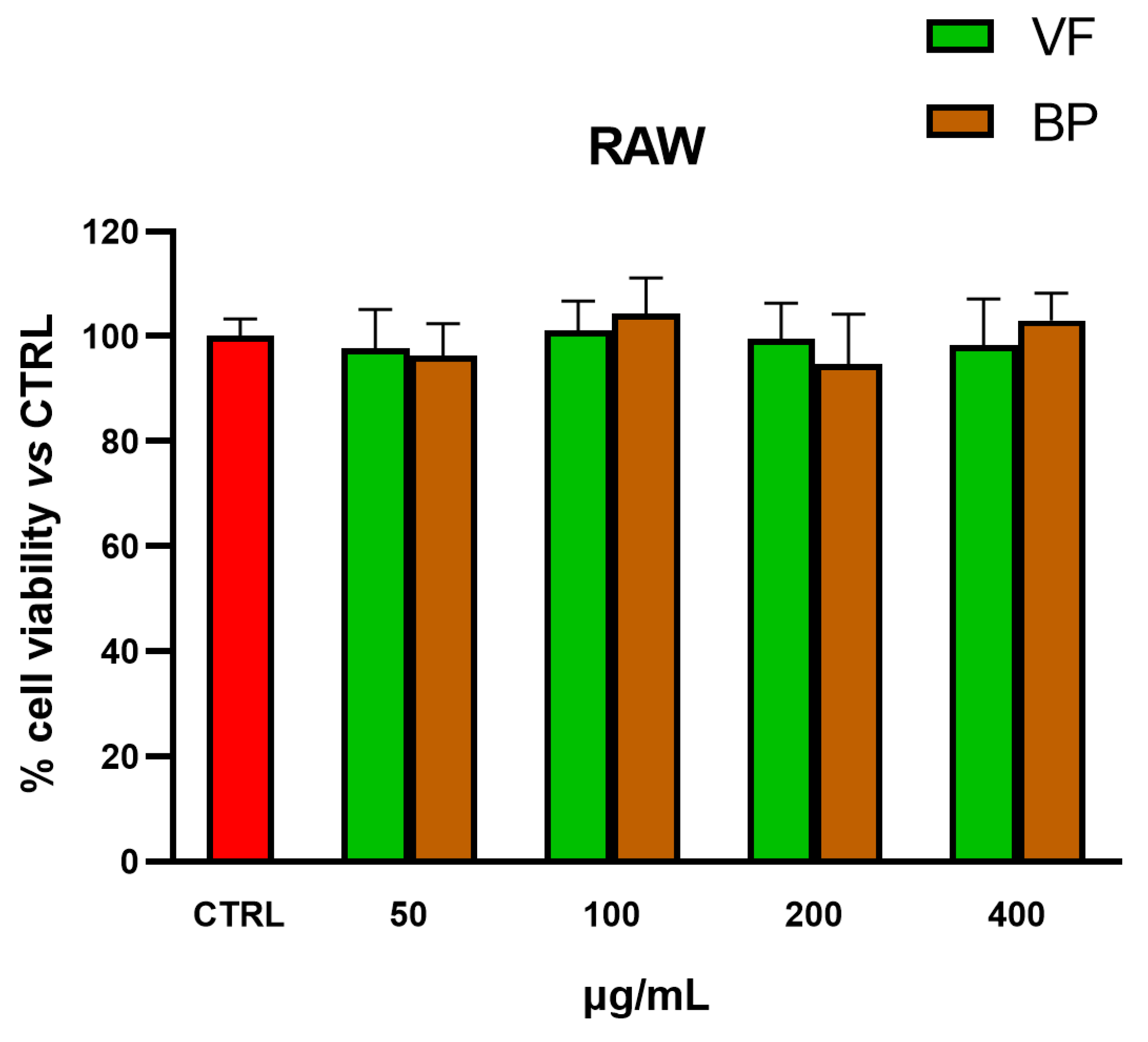
2.5. Anti-Inflammatory Activity in Cells
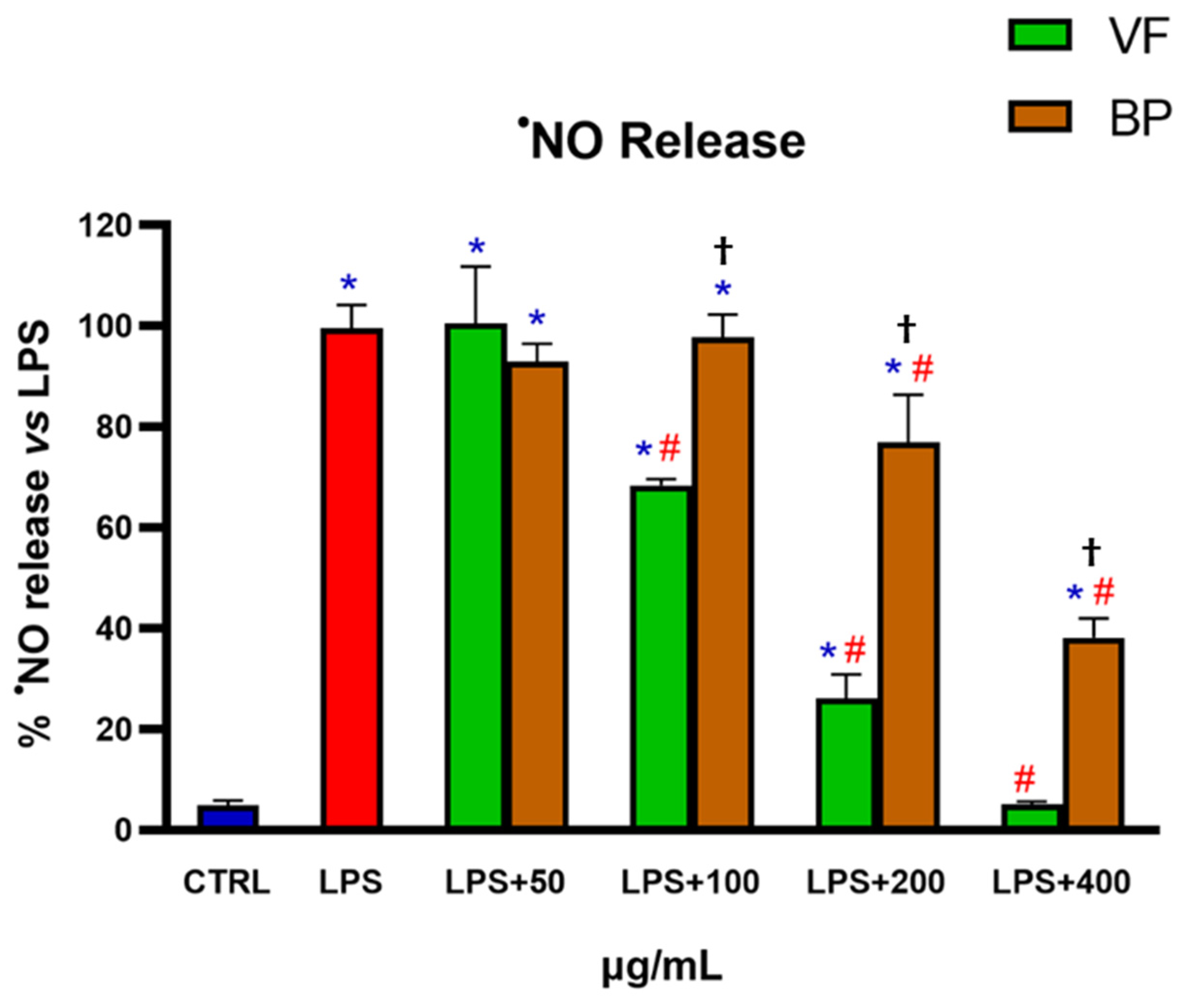
Determination of •NO Release in Murine RAW 264.7 Macrophages
2.6. Inhibition of Cytocrome P450 Isoforms
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material and Extraction Procedure
4.3. HPLC/DAD and HPLC/-ESI-MS Analyses
4.4. Determination of Antioxidant Activity by DPPH Test
4.5. SOD-like Activity Assay
4.6. Catalase-like Activity Assay
4.7. Cell Culture
4.8. Cell Viability by MTT Assay
4.9. Reactive Oxygen Species Assay
4.10. Quantification of •NO Release
4.11. RNA Extraction and Gene Expression Evaluation by RT-qPCR
4.12. Cytochrome P450 Isoforms Activity Assay
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Ascorbic acid |
| BP | By-product |
| CAT | Catalase |
| CYP450 | Cytochrome P450 |
| DPPH | 1,1-Diphenyl-2-picrylhydrazyl |
| FBS | Fetal Bovine Serum |
| FTH1 | ferritin heavy chain 1 |
| H2DCF-DA | dichlorodihydrofluorescein diacetate |
| HMOX1 | heme oxygenase 1 |
| I.F. | Intensity of fluorescence |
| ROS | Reactive oxygen species |
| S.D. | Standard Deviation |
| SOD | Superoxide dismutase |
| VF | Valuable fraction |
References
- Origanum majorana L. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:453303-1 (accessed on 6 May 2025).
- Bouyahya, A.; Chamkhi, I.; Benali, T.; Guaouguaou, F.E.; Balahbib, A.; El Omari, N.; Taha, D.; Belmehdi, O.; Ghokhan, Z.; El Menyiy, N. Traditional use, phytochemistry, toxicology, and pharmacology of Origanum majorana L. J. Ethnopharmacol. 2021, 265, 113318. [Google Scholar] [CrossRef] [PubMed]
- Ietswaart, J.H. A taxonomic revision of the genus Origanum (Labiatae). Leiden Bot. Ser. 1980, 4, 1–153. [Google Scholar]
- Lukas, B.; Novak, J. Origanum majorana L. (Marjoram). In Medicinal, Aromatic and Stimulant Plants. Handbook of Plant Breeding; Novak, J., Blüthner, W.D., Eds.; Springer: Cham, Switzerland, 2020; Volume 12, pp. 407–417. [Google Scholar] [CrossRef]
- Dikr, W. Role of Intercropping some Aromatic and Medicinal Plants with Fruit Vegetables Crops, a Review. Glob. Acad. J. Agri. Biosci. 2022, 4, 22–30. [Google Scholar] [CrossRef]
- Szczech, M.; Kowalska, B.; Wurm, F.R.; Ptaszek, M.; Jarecka-Boncela, A.; Trzciński, P.; Borup Løvschall, K.; Roldan Velasquez, S.T.; Maciorowski, R. The Effects of Tomato Intercropping with Medicinal Aromatic Plants Combined with Trichoderma Applications in Organic Cultivation. Agronomy 2024, 14, 2572. [Google Scholar] [CrossRef]
- Muqaddas, R.A.K.; Nadeem, F.; Jilani, M.I. Essential chemical constituents and medicinal uses of Marjoram (Origanum majorana L.)–A comprehensive review. Int. J. Chem. Biochem. Sci. 2016, 9, 56–62. [Google Scholar]
- Bina, F.; Rahimi, R. Sweet Marjoram: A Review of Ethnopharmacology, Phytochemistry, and Biological Activities. J. Evid. Based Complement. Altern. Med. 2017, 22, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Kakouri, E.; Daferera, D.; Kanakis, C.; Revelou, P.K.; Kaparakou, E.H.; Dervisoglou, S.; Perdikis, D.; Tarantilis, P.A. Origanum majorana Essential Oil—A Review of Its Chemical Profile and Pesticide Activity. Life 2022, 12, 1982. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.R.; Caramão, E.B.; Santos, J.G.; Dariva, C.; Oliveira, J.V. The Effects of Temperature and Pressure on the Characteristics of the Extracts from High-Pressure CO2 Extraction of Majorana hortensis Moench. J. Agric. Food Chem. 2003, 51, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.; Velasco, C.V.; Loureiro, J.M.; Rodrigues, A.E. Effect of cosmetic matrices on the release and odour profiles of the supercritical CO2 extract of Origanum majorana L. Int. J. Cosmet. Sci. 2016, 38, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Ragab, T.I.M.; El Gendy, A.N.G.; Saleh, I.A.; Esawy, M.A. Chemical Composition and Evaluation of Antimicrobial Activity of the Origanum majorana Essential Oil Extracted by Microwave-Assisted Extraction, Conventional Hydro-Distillation and Steam Distillation. J. Essent. Oil Bear. Plants 2019, 22, 563–573. [Google Scholar] [CrossRef]
- Busatta, C.; Barbosa, J.; Cardoso, R.I.; Paroul, N.; Rodrigues, M.; de Oliveira, D.; de Oliveira, J.V.; Cansian, R.L. Chemical Profiles of Essential Oils of Marjoram (Origanum majorana) and Oregano (Origanum vulgare) Obtained by Hydrodistillation and Supercritical CO2. J. Essent. Oil Res. 2017, 29, 367–374. [Google Scholar] [CrossRef]
- Gutiérrez-Grijalva, E.P.; Picos-Salas, M.A.; Leyva-López, N.; Criollo-Mendoza, M.S.; Vazquez-Olivo, G.; Heredia, J.B. Flavonoids and Phenolic Acids from Oregano: Occurrence, Biological Activity and Health Benefits. Plants 2018, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Almaraz-Sánchez, I.; Amaro-Reyes, A.; Acosta-Gallegos, J.A.; Mendoza-Sánchez, M. Processing agroindustry by-products for obtaining value-added products and reducing environmental impact. J. Chem. 2022, 2022, 3656932. [Google Scholar] [CrossRef]
- Ngo, Y.L.; Lau, C.H.; Chua, L.S. Review on rosmarinic acid extraction, fractionation and its anti-diabetic potential. Food Chem. Toxicol. 2018, 121, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Capello, C.; Fischer, U.; Hungerbühler, K. What is a green solvent? A comprehensive framework for the environmental assessment of solvents. Green Chem. 2007, 9, 927–993. [Google Scholar] [CrossRef]
- Tekin, K.; Hao, N.; Karagoz, S.; Ragauskas, A.J. Ethanol: A Promising Green Solvent for the Deconstruction of Lignocellulose. Chem. Sus. Chem. 2018, 11, 3559–3575. [Google Scholar] [CrossRef] [PubMed]
- Tobiszewski, M.; Namieśnik, J.; Pena-Pereira, F. Environmental risk-based ranking of solvents using the combination of a multimedia model and multi-criteria decision analysis. Green Chem. 2017, 19, 1034–1042. [Google Scholar] [CrossRef]
- Gomes, F.; Dias, M.I.; Lima, Â.; Barros, L.; Rodrigues, M.E.; Ferreira, I.C.F.R.; Henriques, M. Satureja montana L. and Origanum majorana L. decoctions: Antimicrobial activity, mode of action and phenolic characterization. Antibiotics 2020, 9, 294. [Google Scholar] [CrossRef] [PubMed]
- Taamalli, A.; Arráez-Román, D.; Abaza, L.; Iswaldi, I.; Fernández-Gutiérrez, A.; Zarrouk, M.; Segura-Carretero, A. LC-MS-based metabolite profiling of methanolic extracts from the medicinal and aromatic species Mentha pulegium and Origanum majorana. Phytochem. Analysis 2015, 26, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Amaghnouje, A.; Mechchate, H.; Es-Safi, I.; Boukhira, S.; Aliqahtani, A.S.; Noman, O.M.; Nasr, F.A.; Conte, R.; Calarco, A.; Bousta, D. Subacute assessment of the toxicity and antidepressant-like effects of Origanum majorana L. Polyphenols in Swiss albino mice. Molecules 2020, 25, 5653. [Google Scholar] [CrossRef] [PubMed]
- Wagdy, R.; Abdel-Kader, R.M.; El-Khatib, A.H.; Linscheid, M.W.; Handoussa, H.; Hamdi, N. Origanum majorana L. Protects against Neuroinflammation-Mediated Cognitive Impairment: A Phyto-Pharmacological Study. BMC Complement. Med. Ther. 2023, 23, 165. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Antioxidant Metabolism Pathways in Vitamins, Polyphenols, and Selenium: Parallels and Divergences. Int. J. Mol. Sci. 2024, 25, 2600. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Azzini, E.; Zucca, P.; Maria Varoni, E.; Anil Kumar, N.V.; Dini, L.; Panzarini, E.; Rajkovic, J.; Valere Tsouh Fokou, P.; Peluso, I.; et al. Plant-Derived Bioactives and Oxidative Stress-Related Disorders: A Key Trend towards Healthy Aging and Longevity Promotion. Appl. Sci. 2020, 10, 947. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef] [PubMed]
- Chiabrando, D.; Vinchi, F.; Fiorito, V.; Mercurio, S.; Tolosano, E. Heme in Pathophysiology: A Matter of Scavenging, Metabolism and Trafficking across Cell Membranes. Front. Pharmacol. 2014, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhao, H.; Lin, Z.; Ye, T.; Xu, D.; Zeng, Q. Heme in Cardiovascular Diseases: A Ubiquitous Dangerous Molecule Worthy of Vigilance. Front. Cell Dev. Biol. 2022, 9, 781839. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Jin, Z.; Xia, C.; Chen, S.; Zeng, L.; Qin, S.; He, Q. Inhibition of Free Heme-Catalyzed Fenton-like Reaction Prevents Non-Alcoholic Fatty Liver Disease by Hepatocyte-Targeted Hydrogen Delivery. Biomaterials 2023, 301, 122230. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.; Elazab, S.T.; Abdelbagi, O.; Saati, A.A.; Babateen, O.; Baokbah, T.A.S.; Qusty, N.F.; Mahmoud, M.E.; Ibrahim, M.M.; Badawy, A.M. Phytochemical Analysis of Origanum majorana L. Extract and Investigation of Its Antioxidant, Anti-Inflammatory and Immunomodulatory Effects against Experimentally Induced Colitis Downregulating Th17 Cells. J. Ethnopharmacol. 2023, 317, 117244. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.; Augustine, D.; Rao, R.S.; Sowmya, S.V.; Haragannavar, V.C.; Nambiar, S.; Prasad, K.; Awan, K.H.; Patil, S. Naturally Available Extracts Inhibiting Cancer Progression: A Systematic Review. J. Evid. Based Complement. Altern. Med. 2017, 22, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Niture, S.K.; Rao, U.S.; Srivenugopal, K.S. Chemopreventative strategies targeting the MGMT repair protein: Augmented expression in human lymphocytes and tumor cells by ethanolic and aqueous extracts of several Indian medicinal plants. Int. J. Oncol. 2006, 29, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Massih, R.M.; Fares, R.; Bazzi, S.; El-Chami, N.; Baydoun, E. The apoptotic and anti-proliferative activity of Origanum majorana extracts on human leukemic cell line. Leuk. Res. 2010, 34, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- El-Ashmawy, I.M.; El-Nahas, A.F.; Salama, O.M. Protective effect of volatile oil, alcoholic and aqueous extracts of Origanum majorana on lead acetate toxicity in mice. Basic Clin. Pharmacol. Toxicol. 2025, 97, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic Inflammation in the Etiology of Disease across the Life Span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Margină, D.; Ungurianu, A.; Purdel, C.; Tsoukalas, D.; Sarandi, E.; Thanasoula, M.; Tekos, F.; Mesnage, R.; Kouretas, D.; Tsatsakis, A. Chronic Inflammation in the Context of Everyday Life: Dietary Changes as Mitigating Factors. Int. J. Environ. Res. Public Health 2020, 17, 4135. [Google Scholar] [CrossRef] [PubMed]
- Bosma-den Boer, M.M.; van Wetten, M.L.; Pruimboom, L. Chronic Inflammatory Diseases Are Stimulated by Current Lifestyle: How Diet, Stress Levels and Medication Prevent Our Body from Recovering. Nutr. Metab. 2012, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Zekrumah, M.; Begua, P.; Razak, A.; Wahab, J.; Moffo, N.; Ivane, A.; Oman, M.; Elrashied, H.; Zou, X.; Zhang, D. Role of Dietary Polyphenols in Non-Communicable Chronic Disease Prevention, and Interactions in Food Systems: An Overview. Nutrition 2023, 112, 112034. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Zhou, M.; Ma, H.L.; Qiao, Y.B.; Li, Q.S. The Role of Chronic Inflammation in Various Diseases and Anti-Inflammatory Therapies Containing Natural Products. ChemMedChem 2021, 16, 1576–1592. [Google Scholar] [CrossRef] [PubMed]
- Gheitasi, I.; Motaghi, N.; Sadeghi, H.; Sadeghi, H.; Moslemi, Z.; Eftekhari, M.; Shakerinasab, N.; Doustimotlagh, A.H. Antioxidant and Anti-Inflammatory Effects of Origanum majorana L. Methanolic Extract on Bile Duct Ligation in Male Rats. Evid. Based Complement. Alternat. Med. 2021, 2021, 9927196. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, S.; Qu, J.; Zhou, H.; Fatma, A.Z.K.K.A.; Kang, W.; Xi, X.; Liu, Z. Anti-Inflammatory Phytoconstituents of Origanum majorana. J. Food Qual. 2022, 2022, 3888075. [Google Scholar] [CrossRef]
- Azhar, M.K.; Anwar, S.; Hasan, G.M.; Shamsi, A.; Islam, A.; Parvez, S.; Hassan, M.I. Comprehensive Insights into Biological Roles of Rosmarinic Acid: Implications in Diabetes, Cancer and Neurodegenerative Diseases. Nutrients 2023, 15, 4297. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Imran, M.; Aslam Gondal, T.; Imran, A.; Shahbaz, M.; Muhammad Amir, R.; Wasim Sajid, M.; Batool Qaisrani, T.; Atif, M.; Hussain, G.; et al. Therapeutic Potential of Rosmarinic Acid: A Comprehensive Review. Appl. Sci. 2019, 9, 3139. [Google Scholar] [CrossRef]
- Qin, T.; Rasul, A.; Sarfraz, A.; Sarfraz, I.; Hussain, G.; Anwar, H.; Riaz, A.; Liu, S.; Wei, W.; Li, J.; et al. Salvianolic Acid A & B: Potential Cytotoxic Polyphenols in Battle against Cancer via Targeting Multiple Signaling Pathways. Int. J. Biol. Sci. 2019, 15, 2256–2264. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Liu, W.; Mu, Y.P.; Zhang, H.; Wang, X.N.; Zhao, C.Q.; Chen, J.M.; Liu, P. Pharmacological Effects of Salvianolic Acid B Against Oxidative Damage. Front. Pharmacol. 2020, 11, 572373. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, M.F.; Ahmad, N.; Ahmed, Z.; Siddique, R.; Zeng, X.A.; Rahaman, A.; Muhammad Aadil, R.; Wahab, A. Novel Extraction Techniques and Pharmaceutical Activities of Luteolin and Its Derivatives. J. Food Biochem. 2019, 43, e12974. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.L.; Huang, H.Y.; Lin, Y.C.; Cai, X.X.; Kong, X.J.; Luo, D.L.; Zhou, Y.H.; Huang, H.D. Enzyme Activity of Natural Products on Cytochrome P450. Molecules 2022, 27, 515. [Google Scholar] [CrossRef] [PubMed]
- Luca, T.; Malfa, G.A.; Siracusa, L.; La Mantia, A.; Bianchi, S.; Napoli, E.; Puleo, S.; Sergi, A.; Acquaviva, R.; Castorina, S. Redox State Modulatory Activity and Cytotoxicity of Olea europaea L. (Oleaceae) Leaves Extract Enriched in Polyphenols Using Macroporous Resin. Antioxidants 2024, 13, 73. [Google Scholar] [CrossRef] [PubMed]
- Genovese, C.; Acquaviva, R.; Ronsisvalle, S.; Tempera, G.; Malfa, G.A.; D’Angeli, F.; Ragusa, S.; Nicolosi, D. In Vitro Evaluation of Biological Activities of Orobanche crenata Forssk. Leaves Extract. Nat. Prod. Res. 2020, 34, 3234–3238. [Google Scholar] [CrossRef] [PubMed]
- Al-Amiery, A.A.; Al-Majedy, Y.K.; Kadhum, A.A.H.; Mohamad, A.B. Hydrogen Peroxide Scavenging Activity of Novel Coumarins Synthesized Using Different Approaches. PLoS ONE 2015, 10, e0132175. [Google Scholar] [CrossRef] [PubMed]
- Tomasello, B.; Malfa, G.A.; La Mantia, A.; Miceli, N.; Sferrazzo, G.; Taviano, M.F.; Di Giacomo, C.; Renis, M.; Acquaviva, R. Anti-Adipogenic and Anti-Oxidant Effects of a Standardised Extract of Moro Blood Oranges (Citrus sinensis (L.) Osbeck) during Adipocyte Differentiation of 3T3-L1 Preadipocytes. Nat. Prod. Res. 2021, 35, 2660–2667. [Google Scholar] [CrossRef] [PubMed]
- Malfa, G.A.; Bianchi, S.; Spadaro, V.; Kozuharova, E.; Marino, P.; Pappalardo, F.; Di Giacomo, C.; Acquaviva, R. Protective Effects of Wild Sulla coronaria (Fabaceae) Flowers Phytocomplex in Human Dermal Fibroblasts Stimulated with Interleukin-1β. Plants 2024, 13, 2748. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt (min) a | Compound Tentative Identification | VF | BP |
|---|---|---|---|---|
| 1 | 7.147 | Caffeoyl-hexose | 103.7 ± 5.9 | 298.9 ± 10.7 |
| 2 | 8.436 | Apigenin di-C-hexoside | 315.0 ± 4.5 | 416.4 ± 5.7 |
| 3 | 10.048 | Luteolin di-O-glucuronide | 63.4 ± 2.3 | 239.9 ± 1.7 |
| 4 | 10.815 | Caffeoylquinic acid | 291.1 ± 5.5 | 67.8 ± 5.2 |
| 5 | 11.146 | Quercetin-O-glucuronide | 45.4 ± 2.8 | 63.6 ± 4.3 |
| 6 | 11.431 | Luteolin -O- glucuronide | 1176.4 ± 9.0 | 409.0 ± 5.6 |
| 7 | 12.024 | p-Coumaroylquinic acid 1 | 90.7 ± 5.6 | 79.1 ± 7.0 |
| 8 | 12.210 | p-Coumaroylquinic acid 2 | 56.1 ± 2.1 | 277.0 ± 5.0 |
| 9 | 12.506 | Rosmarinic acid b | 3321.1 ±45.9 | 2152.9 ± 19.0 |
| 10 | 13.118 | Salvianolic acid B b | 1700.7 ± 45.0 | 1219.5 ± 7.0 |
| 11 | 13.382 | Quercetin-O-hexoside | 63.7 ± 2.0 | 19.7 ± 0.2 |
| 12 | 13.667 | Dicaffeoylquinic acid 1 | 53.9 ± 3.4 | n.d. |
| 13 | 14.015 | di-p-Coumaroylquinic acid 1 | 176.5 ± 1.3 | 138.4 ± 8.7 |
| 14 | 14.370 | Dicaffeoylquinic acid 2 | 50.9 ± 4.8 | 17.3 ± 0.5 |
| 15 | 14.562 | Luteolin b | 52.1 ± 2.5 | 38.2 ± 0.9 |
| 16 | 15.421 | Methyluteolin | 72.6 ± 4.3 | 60.3 ± 5.1 |
| 17 | 15.601 | di-p-Coumaroylquinic acid 2 | 40.7 ± 2.3 | 25.7 ± 0.3 |
| 18 | 15.927 | Salvianolic acid B isomer | 44.0 ± 0.9 | n.d. |
| 19 | 16.162 | Dimethylluteolin | 107.2 ± 2.5 | 29.0 ± 0.8 |
| 20 | 17.001 | Hesperetin b | 100.9 ± 1.2 | 16.3 ± 1.0 |
| Total Polyphenols | 7926.4 ± 69.4 | 5688.8 ± 29.2 |
| O. majorana | VF (IC50 μg/mL ± S.D.) | BP (IC50 μg/mL ± S.D.) | AA (IC50 μg/mL ± S.D.) |
|---|---|---|---|
| DPPH | 30.11 ± 3.46 | 31.72 ± 1.46 | 4.97 ± 0.062 |
| SOD-Like | 0.71 ± 0.062 | 0.79 ± 0.070 | 0.019 ± 0.0055 |
| Catalase-Like | 103.09 ± 4.97 | 119.55 ± 10.58 | 42.55 ± 0.65 |
| Cell Lines | VF (IC50 μg/mL ± S.D.) | BP (IC50 μg/mL ± S.D.) |
|---|---|---|
| CaCo-2 | 154.1 ± 6.22 | 305.2 ± 15.94 † |
| MCF-7 | 624.6 ± 10.27 | 917.9 ± 9.87 † |
| A549 | 720.8 ± 13.66 | 920.2 ± 16.79 † |
| HFF-1 | N.D. a | N.D. a |
| CYP Isoform | VF (IC50 μg/mL ± S.D.) | BP (IC50 μg/mL ± S.D.) | PIC (IC50 μg/mL ± S.D.) |
|---|---|---|---|
| CYP1A2 | 497.45 ± 9.64 | 719.72 ± 11.37 † | 0.0404 ± 0.0021 |
| CYP3A4 | N.D. a | N.D. a | 0.0563 ± 0.0040 |
| CYP2C19 | N.D. a | N.D. a | 0.153 ± 0.0071 |
| CYP2D6 | 637.15 ± 14.78 | 588.70 ± 11.01 † | 0.00630 ± 0.00046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchi, S.; Acquaviva, R.; Di Giacomo, C.; Siracusa, L.; Issop-Merlen, L.; Motterlini, R.; Foresti, R.; Condorelli, D.; Malfa, G.A. Origanum majorana Extracts: A Preliminary Comparative Study on Phytochemical Profiles and Bioactive Properties of Valuable Fraction and By-Product. Plants 2025, 14, 2264. https://doi.org/10.3390/plants14152264
Bianchi S, Acquaviva R, Di Giacomo C, Siracusa L, Issop-Merlen L, Motterlini R, Foresti R, Condorelli D, Malfa GA. Origanum majorana Extracts: A Preliminary Comparative Study on Phytochemical Profiles and Bioactive Properties of Valuable Fraction and By-Product. Plants. 2025; 14(15):2264. https://doi.org/10.3390/plants14152264
Chicago/Turabian StyleBianchi, Simone, Rosaria Acquaviva, Claudia Di Giacomo, Laura Siracusa, Leeyah Issop-Merlen, Roberto Motterlini, Roberta Foresti, Donata Condorelli, and Giuseppe Antonio Malfa. 2025. "Origanum majorana Extracts: A Preliminary Comparative Study on Phytochemical Profiles and Bioactive Properties of Valuable Fraction and By-Product" Plants 14, no. 15: 2264. https://doi.org/10.3390/plants14152264
APA StyleBianchi, S., Acquaviva, R., Di Giacomo, C., Siracusa, L., Issop-Merlen, L., Motterlini, R., Foresti, R., Condorelli, D., & Malfa, G. A. (2025). Origanum majorana Extracts: A Preliminary Comparative Study on Phytochemical Profiles and Bioactive Properties of Valuable Fraction and By-Product. Plants, 14(15), 2264. https://doi.org/10.3390/plants14152264