
Modeling the Future of a Wild Edible Fern Under Climate Change: Distribution and Cultivation Zones of Pteridium aquilinum var. latiusculum in the Dadu–Min River Region

 ,
,
Abstract
1. Introduction
2. Results
2.1. Current Distribution Projections and Model Accuracy Validation
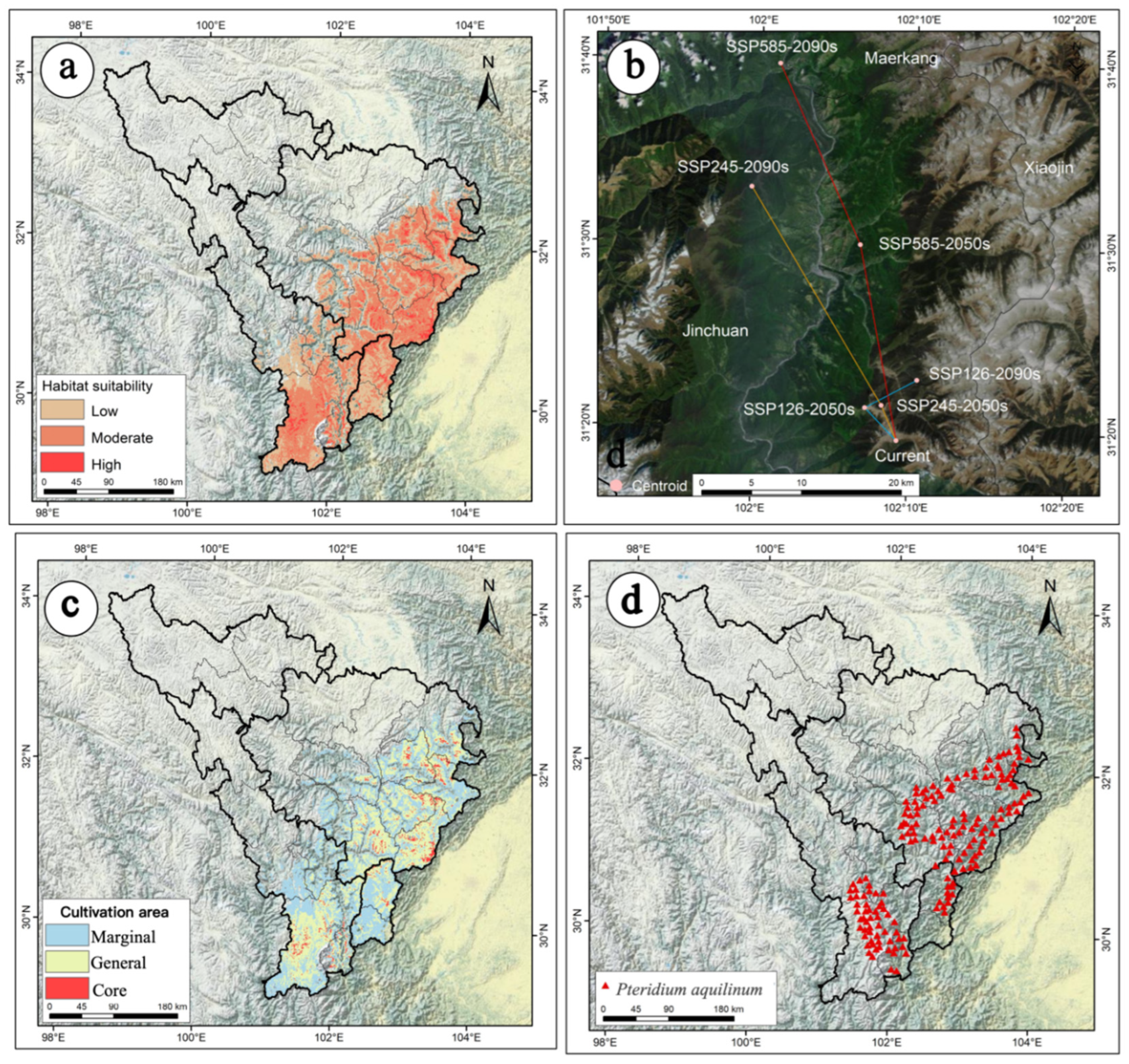
2.2. The Current Potential Distribution of P. aquilinum var. latiusculum in the Upper Dadu River–Min River Basin
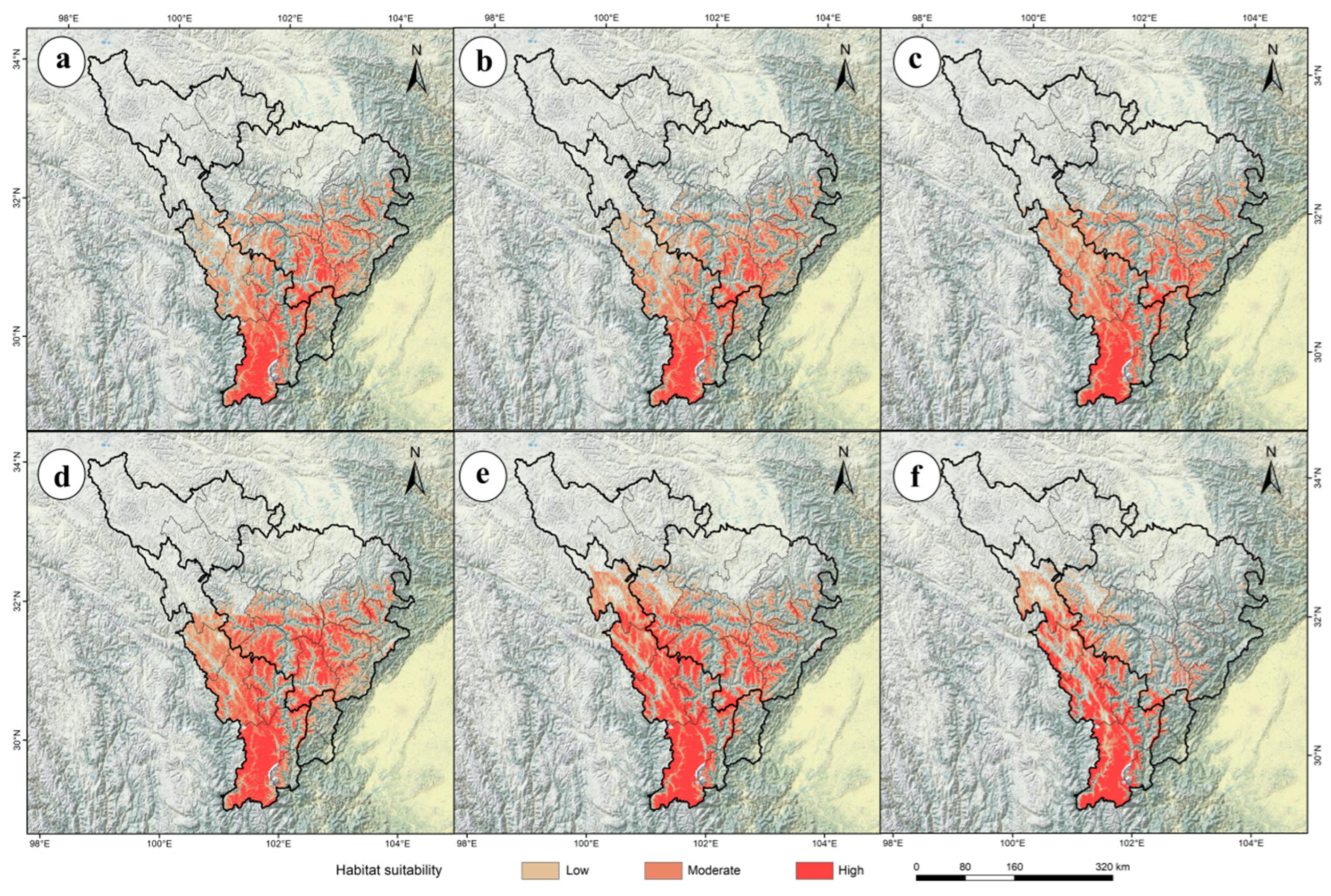
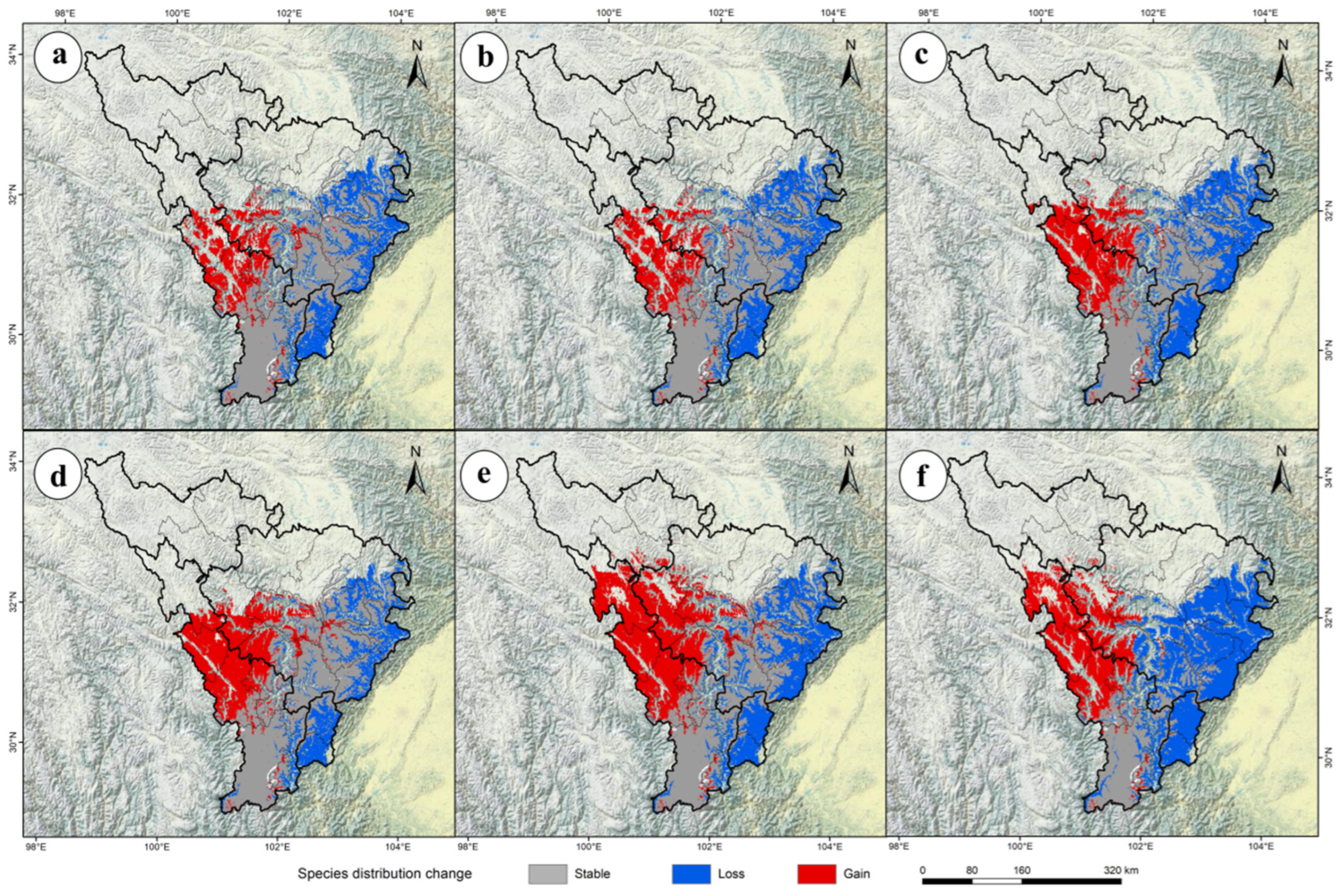
2.3. The Future Potential Distribution of P. aquilinum var. latiusculum in the Upper Dadu River–Min River Basin
2.4. The Movement Trajectory of the Centroid of Suitable Habitats Under Future Climate Change
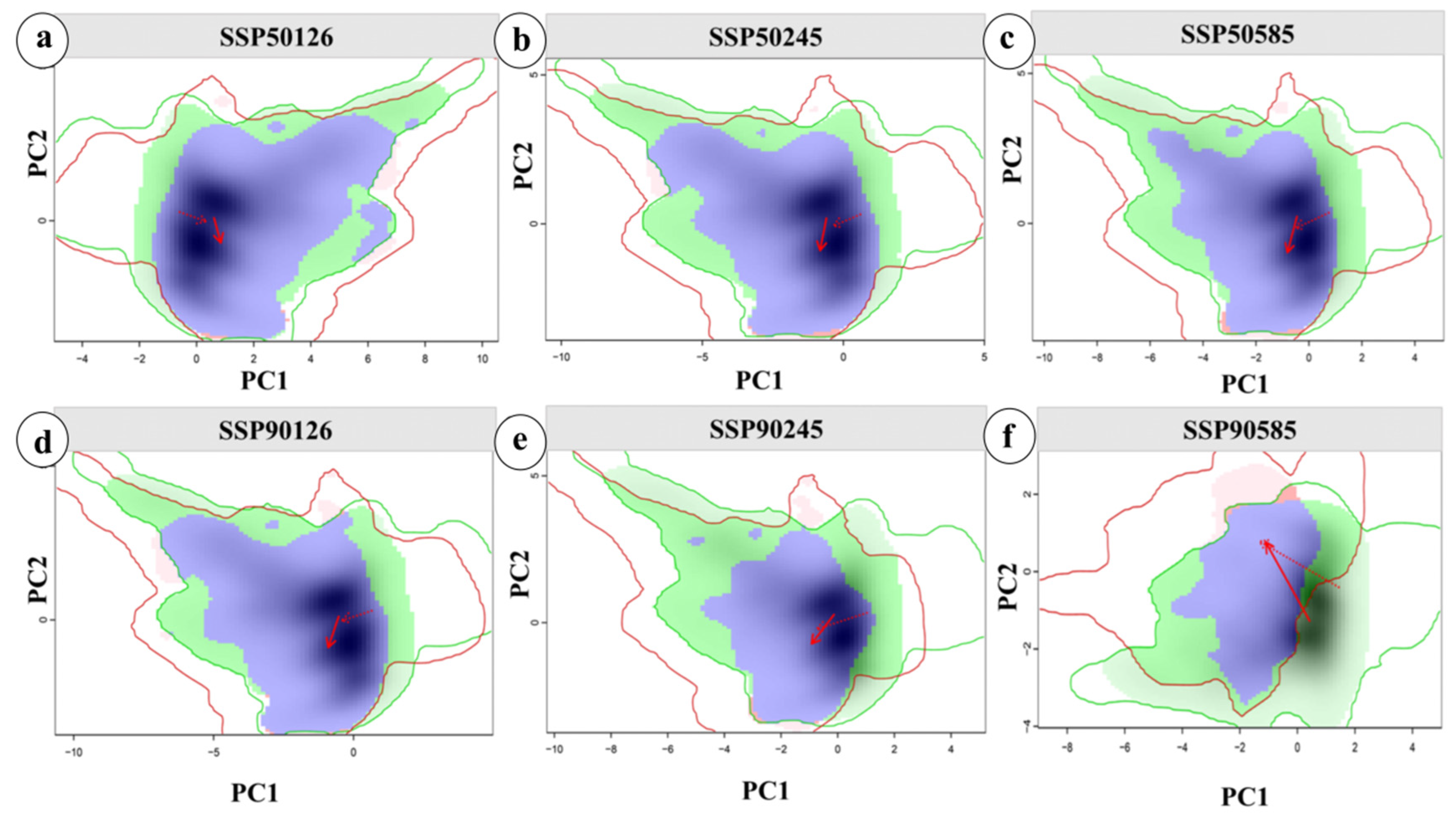
2.5. Analysis of Niche Changes in Future Periods
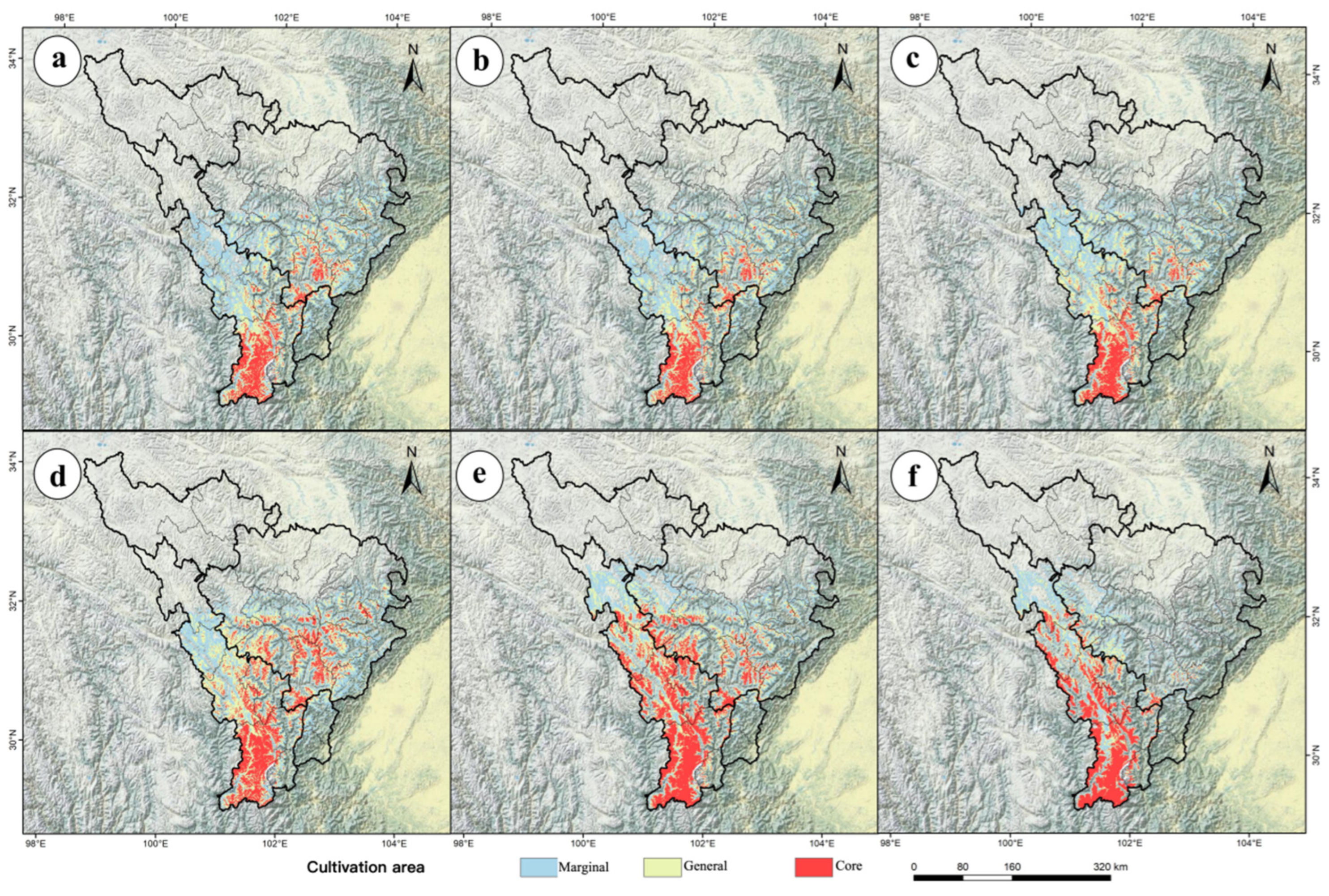
2.6. Delineation of Potential Cultivation Areas for P. aquilinum var. latiusculum in Different Periods
3. Discussion
3.1. The Significance of Conducting Distribution Modeling for P. aquilinum var. latiusculum
3.2. Advantages of Using Ensemble Models in This Study
3.3. Impacts of Climate Change on P. aquilinum var. latiusculum
3.4. Adaptive Management of P. aquilinum var. latiusculum Under Climate Warming
3.5. Research Prospects
4. Materials and Methods
4.1. Sample Collection and Species Distribution Records
4.2. Selection and Processing of Environmental Variables
4.3. Ensemble Model Construction
4.4. Niche Dynamics Analysis
4.5. Centroid Migration Trajectory
4.6. Establishing Cultivation Productivity–Suitability Relationships
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wan, J.N.; Wang, S.W.; Leitch, A.R.; Leitch, I.J.; Jian, J.B.; Wu, Z.Y.; Xin, H.P.; Rakotoarinivo, M.; Onjalalaina, G.E.; Gituru, R.W.; et al. The Rise of Baobab Trees in Madagascar. Nature 2024, 629, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Mi, C.; Ma, L.; Yang, M.; Li, X.; Meiri, S.; Roll, U.; Oskyrko, O.; Pincheira-Donoso, D.; Harvey, L.P.; Jablonski, D.; et al. Global Protected Areas as Refuges for Amphibians and Reptiles under Climate Change. Nat. Commun. 2023, 14, 1389. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction Risk from Climate Change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.H.; Dong, G.H.; Zhang, D.J.; Liu, X.Y.; Jia, X.; An, C.B.; Ma, M.M.; Xie, Y.W.; Barton, L.; Ren, X.Y.; et al. Agriculture Facilitated Permanent Human Occupation of the Tibetan Plateau after 3600 B.P. Science 2015, 347, 248–250. [Google Scholar] [CrossRef]
- Avasthi, A. California Tries to Connect Its Scattered Marine Reserves. Science 2005, 308, 487–488. [Google Scholar] [CrossRef] [PubMed]
- Brooks, T.M.; Mittermeier, R.A.; da Fonseca, G.A.B.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.G.; Pilgrim, J.D.; Rodrigues, A.S.L. Global Biodiversity Conservation Priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Teixidor-Toneu, I.; Jordan, F.M.; Hawkins, J.A. Comparative Phylogenetic Methods and the Cultural Evolution of Medicinal Plant Use. Nat. Plants 2018, 4, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Cantwell-Jones, A.; Ball, J.; Collar, D.; Diazgranados, M.; Douglas, R.; Forest, F.; Hawkins, J.; Howes, M.-J.R.; Ulian, T.; Vaitla, B.; et al. Global Plant Diversity as a Reservoir of Micronutrients for Humanity. Nat. Plants 2022, 8, 225–232. [Google Scholar] [CrossRef]
- Cheng, Z.; Lu, X.; Hu, X.; Zhang, Q.; Ali, M.; Long, C. Dulong People’s Traditional Knowledge of Caryota obtusa (Arecaceae): A Potential Starch Plant with Emphasis on Its Starch Properties and Distribution Prediction. Econ. Bot. 2023, 77, 63–81. [Google Scholar] [CrossRef]
- Cheng, Z.; Lu, X.; Lin, F.; Naeem, A.; Long, C. Ethnobotanical Study on Wild Edible Plants Used by Dulong People in Northwestern Yunnan, China. J. Ethnobiol. Ethnomed. 2022, 18, 3. [Google Scholar] [CrossRef]
- Pan, Y.; Guan, Y.; Lv, S.; Huang, X.; Lin, Y.; Wei, C.; Xu, D. Assessing the Potential Distribution of Lonicera Japonica in China Under Climate Change: A Biomod2 Ensemble Model-Based Study. Agriculture 2025, 15, 393. [Google Scholar] [CrossRef]
- Yu, H.; Zhuo, Z.; He, Z.; Liu, Q.; Deng, X.; Xu, D. Distribution of Trachelospermum Jasminoides Under the Influence of Different Environmental Factors. Agriculture 2025, 15, 285. [Google Scholar] [CrossRef]
- Wen, X.; Fang, G.; Chai, S.; He, C.; Sun, S.; Zhao, G.; Lin, X. Can Ecological Niche Models Be Used to Accurately Predict the Distribution of Invasive Insects? A Case Study of Hyphantria cunea in China. Ecol. Evol. 2024, 14, e11159. [Google Scholar] [CrossRef]
- Zhan, P.; Wang, F.; Xia, P.; Zhao, G.; Wei, M.; Wei, F.; Han, R. Assessment of Suitable Cultivation Region for Panax Notoginseng under Different Climatic Conditions Using MaxEnt Model and High-Performance Liquid Chromatography in China. Ind. Crops Prod. 2022, 176, 114416. [Google Scholar] [CrossRef]
- Chen, C.; Xiao, W.; Chen, H.Y.H. Meta-Analysis Reveals Global Variations in Plant Diversity Effects on Productivity. Nature 2025, 638, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Cui, X.; Sun, J.; Li, T.; Wang, Q.; Ye, X.; Fan, B. Analysis of the Distribution Pattern of Chinese Ziziphus jujuba under Climate Change Based on Optimized Biomod2 and MaxEnt Models. Ecol. Indic. 2021, 132, 108256. [Google Scholar] [CrossRef]
- Yang, J.; Huang, Y.; Jiang, X.; Chen, H.; Liu, M.; Wang, R. Potential Geographical Distribution of the Edangred Plant Isoetes under Human Activities Using MaxEnt and GARP. Glob. Ecol. Conserv. 2022, 38, e02186. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting Species Distribution: Offering More than Simple Habitat Models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD–a Platform for Ensemble Forecasting of Species Distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, R.; Zhou, X.; Zhang, X.; Zhao, G.; Zhang, F. Prediction of Potential Distribution Areas and Priority Protected Areas of Agastache Rugosa Based on Maxent Model and Marxan Model. Front. Plant Sci. 2023, 14, 1200796. [Google Scholar] [CrossRef]
- Huang, Y.; Zeng, Y.; Jiang, P.; Chen, H.; Yang, J. Prediction of Potential Geographic Distribution of Endangered Relict Tree Species Dipteronia Sinensis in China Based on MaxEnt and GIS. Pol. J. Environ. Stud. 2022, 31, 3597–3609. [Google Scholar] [CrossRef]
- Liu, L.; Liang, S.; Xie, C.; Liu, J.; Zheng, Y.; Xue, J. Predicting the Potential Distribution of Aralia chinensis L.(Wild Vegetable) in China Under Different Climate Change Scenarios. Biology 2024, 13, 937. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.; Yang, L.; Chen, K.; Shama, Z.; Jiang, X.; Yang, J.; Zhao, G.; Huang, Y. Prediction of the Potential Distribution and Conservation Strategies of the Endangered Plant Tapiscia sinensis. Forests 2024, 15, 1677. [Google Scholar] [CrossRef]
- Xie, C.; Chen, L.; Li, M.; Jim, C.Y.; Liu, D. BIOCLIM Modeling for Predicting Suitable Habitat for Endangered Tree Tapiscia sinensis (Tapisciaceae) in China. Forests 2023, 14, 2275. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A Review of Evidence about Use and Performance of Species Distribution Modelling Ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Wen, X.; Zhao, G.; Cheng, X.; Chang, G.; Dong, X.; Lin, X. Prediction of the Potential Distribution Pattern of the Great Gerbil (Rhombomys opimus) under Climate Change Based on Ensemble Modelling. Pest Manag. Sci. 2022, 78, 3128–3134. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Extraction, Activity Analysis of Flavonoids from Pteridium aquilinum var. latiusculum and Their Effects on Storage Quality of Pteridium aquilinum var. latiusculum Filling. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2024. [Google Scholar]
- Zimeng, F.; Xiang, H.; Shuangli, L.; Xiaohui, W. Antidepressant-like Effects of Flavonoids from Pteridium aquilinum var. latiusculum. Med. Plant 2023, 14, 55. [Google Scholar] [CrossRef]
- Thepsilvisut, O.; Iad-Ak, R.; Chutimanukul, P. The Effects of Shading and Nutrient Management on Yield Quality of Vegetable Fern. Horticulturae 2023, 9, 259. [Google Scholar] [CrossRef]
- Reddy, M.N.; Adnan, M.; Alreshidi, M.M.; Saeed, M.; Patel, M. Evaluation of Anticancer, Antibacterial and Antioxidant Properties of a Medicinally Treasured Fern Tectaria coadunata with Its Phytoconstituents Analysis by HR-LCMS. Anti-Cancer Agents Med. Chem. 2020, 20, 1845–1856. [Google Scholar] [CrossRef]
- Hao, J.; Fan, X.; Chen, N.; Wang, Y.; Xiao, J.; Zhang, M. A Novel Method for the Simultaneous Determination of Flavonoids in Pteridium aquilinum via SPE Combined with UPLC-DAD via QAMS. J. Food Compos. Anal. 2025, 137, 106881. [Google Scholar] [CrossRef]
- Adebayo, O.T.; Oluremi, B.B.; Ogunlakin, A.D.; Gyebi, G.A.; Sonibare, M.A. Pharmacognostic Evaluation and Antimicrobial Activity of Pteridium aquilinum (L.) Kuhn Leaves (Onocleaceae) via in Vitro and in Silico Perspectives. PLoS ONE 2025, 20, e0318943. [Google Scholar] [CrossRef]
- Li, S.; Chen, N.; Bu, Y.; Hao, J.; Huang, H.; Zheng, S. Network Pharmacology Analysis of the Renal Protective Effects of the Bracken (Pteridium aquilinum) Using Experimental Data Obtained in an Avian (Quail) Hyperuricemia Model. Int. J. Clin. Pharmacol. Ther. 2024, 62, 178–193. [Google Scholar] [PubMed]
- Huang, Y.; Zhao, G.; Yang, J.; Yang, L.; Yang, Y.; Jiaba, W.; Shama, Z.; Yang, J. Evolution of Potential Distribution Areas and Cultivation Zones of Morchella esculenta (L.) Pers. Under Climate Warming: Application of Ensemble Models and Production Dynamics Models. J. Fungi 2025, 11, 475. [Google Scholar]
- Huang, Y.; Yang, J.; Zhao, G.; Yang, Y. Potential Distribution and Cultivation Areas of Argentina anserina (Rosaceae) in the Upper Reaches of the Dadu River and Minjiang River Basin Under Climate Change: Applications of Ensemble and Productivity Dynamic Models. Biology 2025, 14, 668. [Google Scholar] [CrossRef]
- Liu, M.; Su, M.M.; Cai, K.; Qi, P.S.; Gong, W.; Chen, D.-C.; He, L.; Wu, H.W.; Deng, D.Z.; Huang, Y. Distributional Response of Paeonia decomposita to Climate Change and Conservation Strategies. Pol. J. Environ. Stud. 2025, 34, 3657–3669. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Huang, Y.; Zhang, J.; Mou, Q.; Qiu, J.; Wang, R.; Li, Y.; Zhang, D. Simulation of Potential Suitable Distribution of Original Species of Fritillariae Cirrhosae Bulbus in China under Climate Change Scenarios. Environ. Sci. Pollut. Res. 2022, 29, 22237–22250. [Google Scholar] [CrossRef]
- Ghorbani, J.; Cox, E.S.; Alday, J.G.; Santana, V.M.; Lee, H.; McAllister, H.A.; Pakeman, R.J.; Le Duc, M.G.; Marrs, R.H. What Happens When You Turn Weed Management off? A Long-Term Appraisal of the Effectiveness of Pteridium aquilinum (L.) Kuhn Control Treatments and the Role of Sheep Grazing. J. Environ. Manag. 2025, 374, 124111. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the Accuracy of Species Distribution Models: Prevalence, Kappa and the True Skill Statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble Forecasting of Species Distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Linderman, M.; Liu, J.; Qi, J.; An, L.; Ouyang, Z.; Yang, J.; Tan, Y. Using Artificial Neural Networks to Map the Spatial Distribution of Understorey Bamboo from Remote Sensing Data. Int. J. Remote Sens. 2004, 25, 1685–1700. [Google Scholar]
- Thuiller, W. BIOMOD–Optimizing Predictions of Species Distributions and Projecting Potential Future Shifts under Global Change. Glob. Change Biol. 2003, 9, 1353–1362. [Google Scholar] [CrossRef]
- Garcia, K.; Lasco, R.; Ines, A.; Lyon, B.; Pulhin, F. Predicting Geographic Distribution and Habitat Suitability Due to Climate Change of Selected Threatened Forest Tree Species in the Philippines. Appl. Geogr. 2013, 44, 12–22. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Padula, R.; Lorenzoni, M. Population Status and Ecology of the Salmo Trutta Complex in an Italian River Basin under Multiple Anthropogenic Pressures. Ecol. Evol. 2020, 10, 7320–7333. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A Globally Coherent Fingerprint of Climate Change Impacts across Natural Systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Bai, Y.; Han, Z.; Wang, L.; Lian, P. The influence of different cultivation environment conditions on the growth and yield of ferns. Shelter. For. Technol. 2019, 6, 19–20. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Guo, B.; Zang, W.; Yang, F.; Han, B.; Chen, S.; Liu, Y.; Yang, X.; He, T.; Chen, X.; Liu, C.; et al. Spatial and Temporal Change Patterns of Net Primary Productivity and Its Response to Climate Change in the Qinghai-Tibet Plateau of China from 2000 to 2015. J. Arid Land 2020, 12, 1–17. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A Significant Upward Shift in Plant Species Optimum Elevation During the 20th Century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Zhu, K.; Woodall, C.W.; Clark, J.S. Failure to Migrate: Lack of Tree Range Expansion in Response to Climate Change. Glob. Change Biol. 2012, 18, 1042–1052. [Google Scholar]
- Brandt, J.S.; Haynes, M.A.; Kuemmerle, T.; Waller, D.M.; Radeloff, V.C. Regime shift on the roof of the world: Alpine meadows converting to shrublands in the southern Himalayas. Biol. Conserv. 2013, 158, 116–127. [Google Scholar]
- Zizka, A.; Silvestro, D.; Andermann, T.; Azevedo, J.; Duarte Ritter, C.; Edler, D.; Farooq, H.; Herdean, A.; Ariza, M.; Scharn, R. CoordinateCleaner: Standardized Cleaning of Occurrence Records from Biological Collection Databases. Methods Ecol. Evol. 2019, 10, 744–751. [Google Scholar]
- Sun, J.; Jiao, W.; Wang, Q.; Wang, T.; Yang, H.; Jin, J.; Feng, H.; Guo, J.; Feng, L.; Xu, X.; et al. Potential Habitat and Productivity Loss of Populus Deltoides Industrial Forest Plantations Due to Global Warming. For. Ecol. Manag. 2021, 496, 119474. [Google Scholar] [CrossRef]
- Beier, P. Model Selection and Inference: A Practical Information-Theoretic Approach. J. Wildl. Manag. 2001, 65, 606–608. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Climate Scenario | Low Suitability Zone | Moderate Suitability Zone | High Suitability Zone | Total Suitability Zone |
|---|---|---|---|---|---|
| Current | 2.62 | 3.25 | 0.36 | 6.23 | |
| 2050 | SSP126 | 2.29 | 2.18 | 1.19 | 5.66 |
| 2050 | SSP245 | 2.24 | 2.05 | 1.12 | 5.41 |
| 2050 | SSP585 | 2.11 | 2.31 | 1.22 | 5.64 |
| 2090 | SSP126 | 2.09 | 2.89 | 2.06 | 7.04 |
| 2090 | SSP245 | 2.08 | 2.42 | 2.74 | 7.24 |
| 2090 | SSP585 | 1.59 | 1.35 | 1.79 | 4.73 |
| Period | Climate Scenario | Habitat Area | Loss | Stable | Gain |
|---|---|---|---|---|---|
| 2050 | SSP126 | 5.66 | 2.04 | 4.19 | 1.48 |
| 2050 | SSP245 | 5.41 | 2.29 | 3.94 | 1.47 |
| 2050 | SSP585 | 5.64 | 2.62 | 3.61 | 2.03 |
| 2090 | SSP126 | 7.04 | 1.82 | 4.41 | 2.63 |
| 2090 | SSP245 | 7.24 | 2.55 | 3.68 | 3.56 |
| 2090 | SSP585 | 4.73 | 4.08 | 2.15 | 2.58 |
| Period | Climate Scenario | Marginal Cultivation | General Cultivation | Core Cultivation | Total Cultivation |
|---|---|---|---|---|---|
| Current | 3.20 | 2.68 | 0.24 | 6.12 | |
| 2050 | SSP126 | 2.55 | 1.95 | 0.98 | 5.48 |
| 2050 | SSP245 | 2.49 | 1.84 | 0.91 | 5.24 |
| 2050 | SSP585 | 2.46 | 2.05 | 1.01 | 5.52 |
| 2090 | SSP126 | 2.43 | 2.77 | 1.71 | 6.91 |
| 2090 | SSP245 | 2.31 | 2.38 | 2.39 | 7.08 |
| 2090 | SSP585 | 1.68 | 1.28 | 1.63 | 4.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Yang, J.; Zhao, G.; Shama, Z.; Ge, Q.; Yang, Y.; Yang, J. Modeling the Future of a Wild Edible Fern Under Climate Change: Distribution and Cultivation Zones of Pteridium aquilinum var. latiusculum in the Dadu–Min River Region. Plants 2025, 14, 2123. https://doi.org/10.3390/plants14142123
Huang Y, Yang J, Zhao G, Shama Z, Ge Q, Yang Y, Yang J. Modeling the Future of a Wild Edible Fern Under Climate Change: Distribution and Cultivation Zones of Pteridium aquilinum var. latiusculum in the Dadu–Min River Region. Plants. 2025; 14(14):2123. https://doi.org/10.3390/plants14142123
Chicago/Turabian StyleHuang, Yi, Jingtian Yang, Guanghua Zhao, Zixi Shama, Qingsong Ge, Yang Yang, and Jian Yang. 2025. "Modeling the Future of a Wild Edible Fern Under Climate Change: Distribution and Cultivation Zones of Pteridium aquilinum var. latiusculum in the Dadu–Min River Region" Plants 14, no. 14: 2123. https://doi.org/10.3390/plants14142123
APA StyleHuang, Y., Yang, J., Zhao, G., Shama, Z., Ge, Q., Yang, Y., & Yang, J. (2025). Modeling the Future of a Wild Edible Fern Under Climate Change: Distribution and Cultivation Zones of Pteridium aquilinum var. latiusculum in the Dadu–Min River Region. Plants, 14(14), 2123. https://doi.org/10.3390/plants14142123