
Genomic Insights and Plant Growth-Promoting Characterization of Priestia megaterium Strain 53B2, Isolated from Maize-Associated Soil in the Yaqui Valley, Mexico

 , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Results
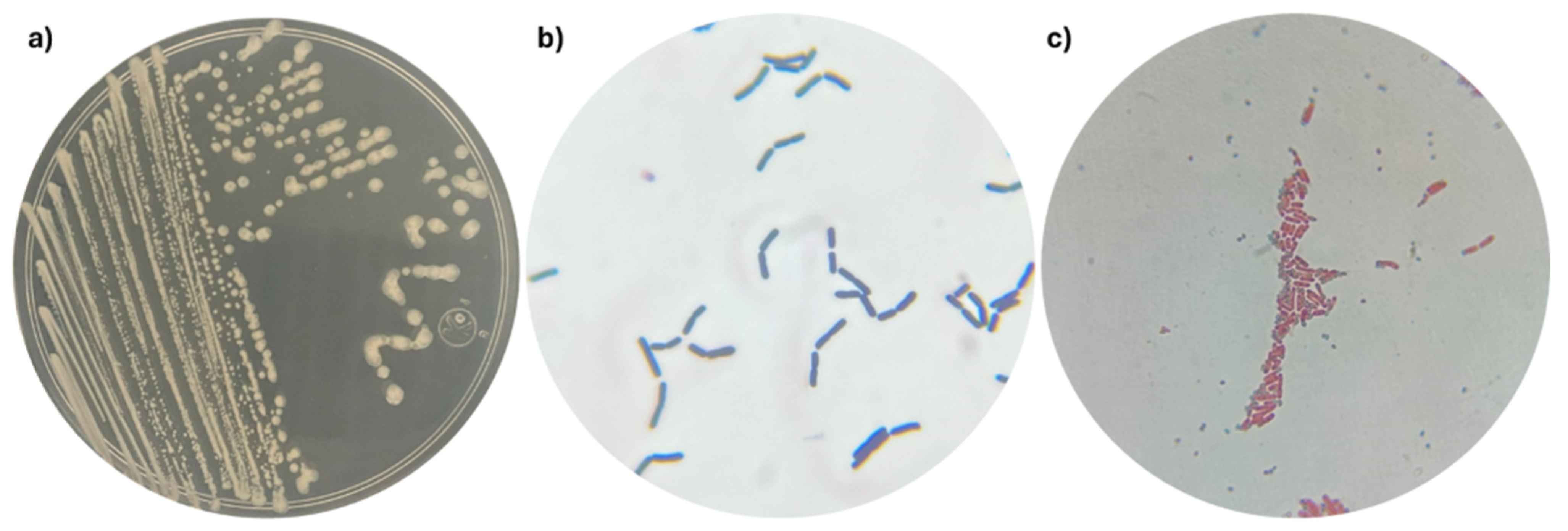
2.1. Morphological and Metabolic Characterization
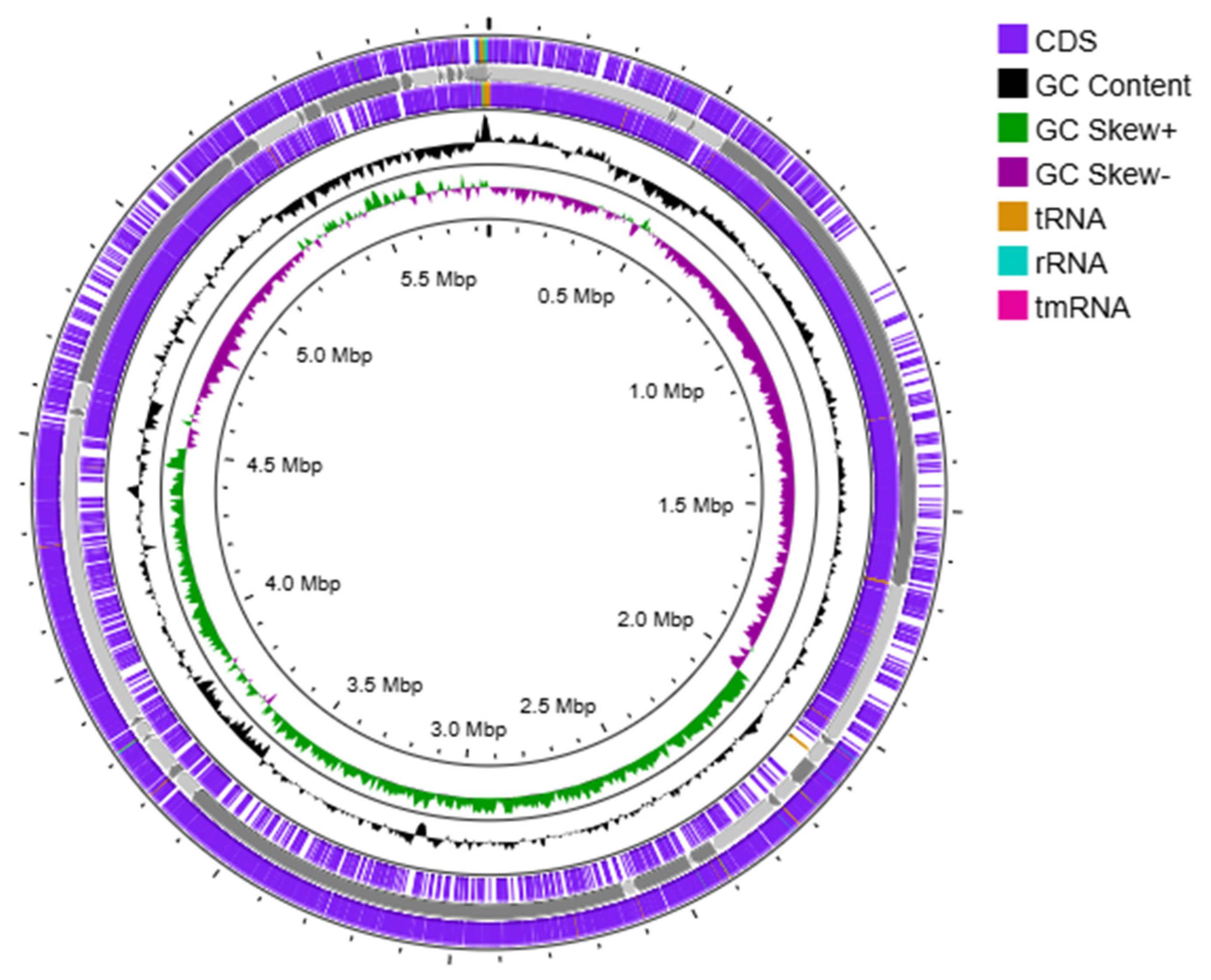
2.2. Genomic Analysis
2.3. Genome Annotation
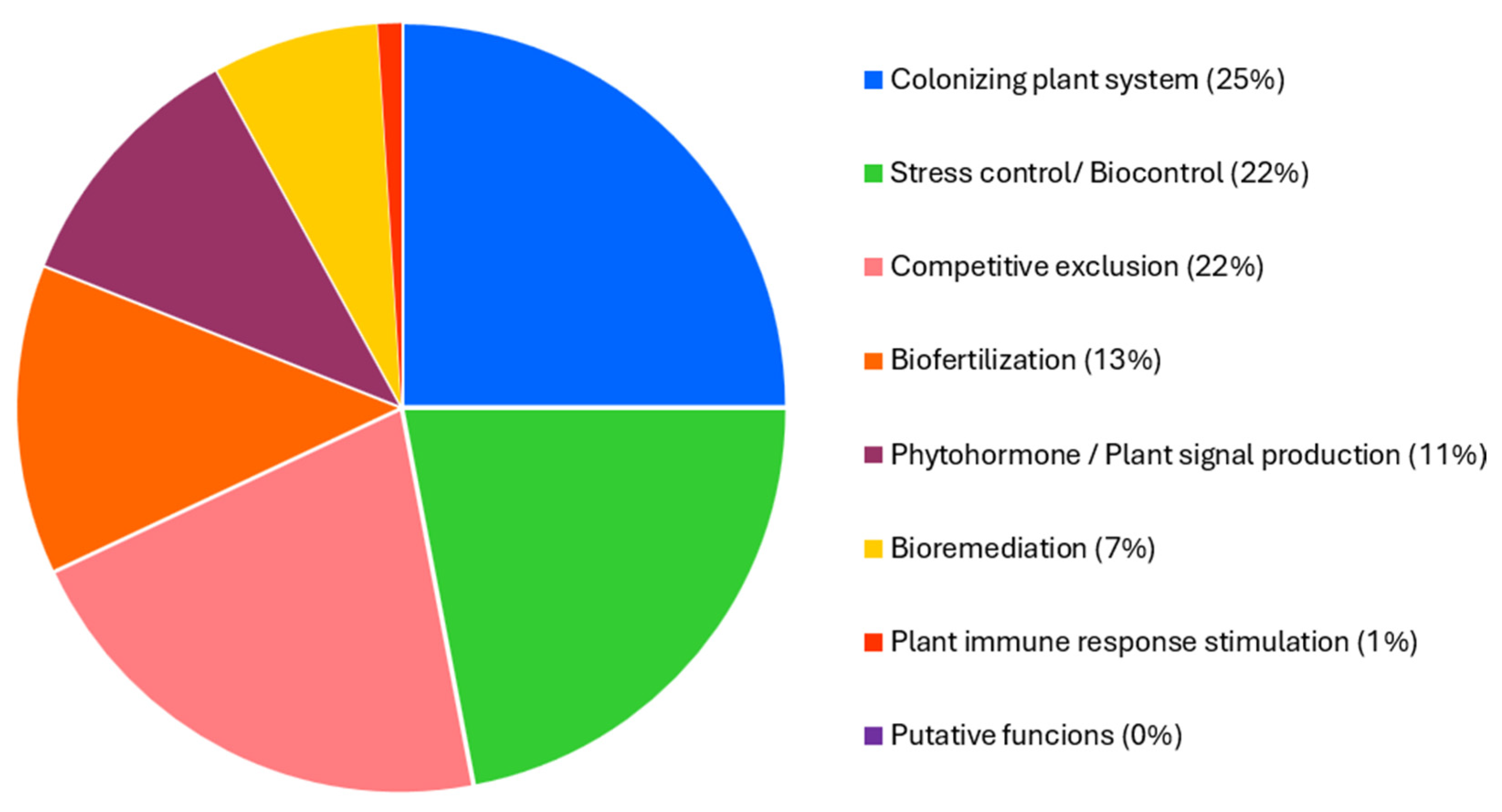
2.4. Genome Mining
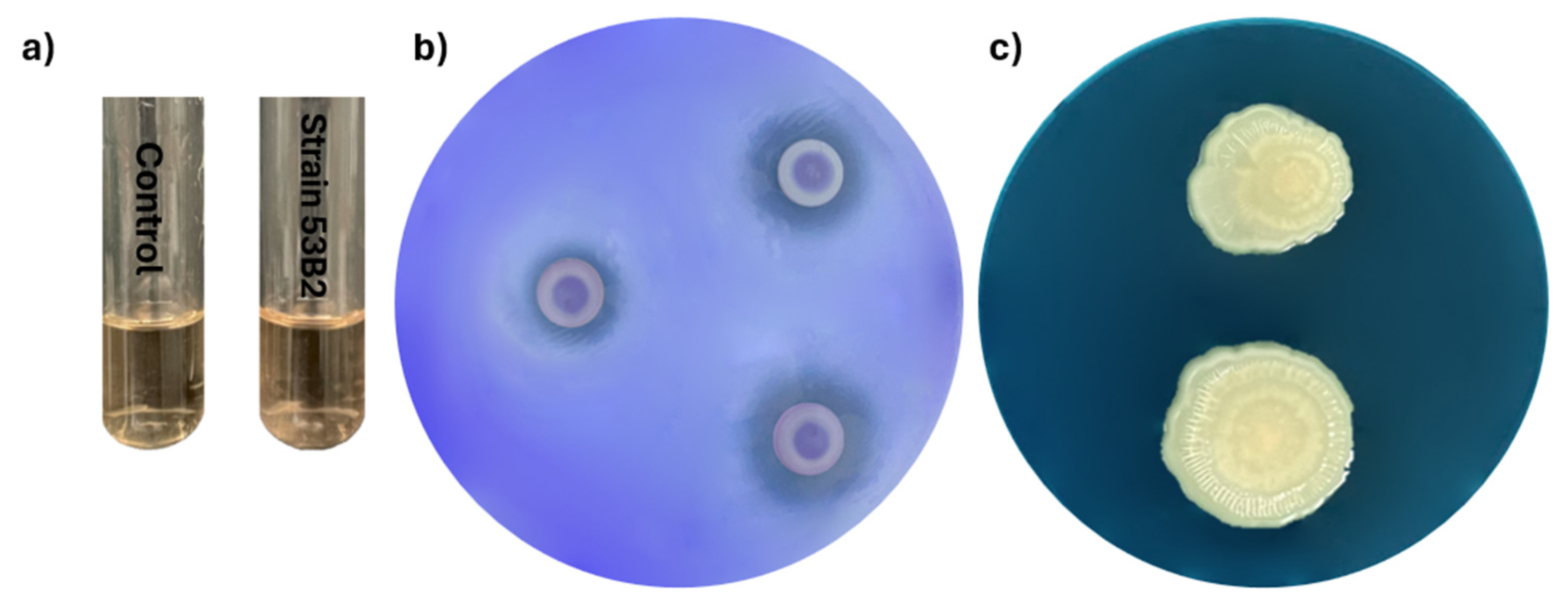
2.5. Metabolic Characterization and Biocontrol Assay
2.6. Maize Growth Promotion Assay
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolation and Cultivation
4.2. Genomic Analysis
4.3. Genome Annotation
4.4. Genome Mining
4.5. Metabolic Characterization
4.6. Biocontrol In Vitro Assay
4.7. Maize Growth Promotion Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). How to Feed the World in 2050. In Proceedings of the Expert Meeting on How to Feed the World in 2050, Rome, Italy, 24–26 June 2009. [Google Scholar]
- Grote, U.; Fasse, A.; Nguyen, T.T.; Erenstein, O. Food Security and the Dynamics of Wheat and Maize Value Chains in Africa and Asia. Front. Sustain. Food Syst. 2021, 4, 617009. [Google Scholar] [CrossRef]
- Byerlee, D.; Fanzo, J. The SDG of Zero Hunger 75 years on: Turning Full Circle on Agriculture and Nutrition. Glob. Food Secur. 2019, 21, 52–59. [Google Scholar] [CrossRef]
- Poole, N.; Donovan, J.; Erenstein, O. Viewpoint: Agri-Nutrition Research: Revisiting the Contribution of Maize and Wheat to Human Nutrition and Health. Food Policy 2021, 100, 101976. [Google Scholar] [CrossRef]
- Yamini, V.; Singh, K.; Antar, M.; El Sabagh, A. Sustainable Cereal Production through Integrated Crop Management: A Global Review of Current Practices and Future Prospects. Front. Sustain. Food Syst. 2025, 9, 1428687. [Google Scholar] [CrossRef]
- SIAP (Servicio de Información Agroalimentaria y Pesquera). Anuario Estadístico de La Producción Agrícola. Available online: https://nube.siap.gob.mx/cierreagricola/ (accessed on 30 January 2025).
- de los Santos-Villalobos, S.; Parra-Cota, F.I.; Ayala-Zepeda, M.; Díaz-Rodríguez, A.M. Tecnologías Para Una Producción Sostenible de Trigo: El Caso Del Valle Del Yaqui, México, 1st ed.; Fontamara: Mexico City, Mexico, 2023. [Google Scholar]
- Parra-Cota, F.I.; Coronel-Acosta, C.-B.; Amézquita-Avilés, C.F.; de los Santos-Villalobos, S.; Escalante-Martínez, D.I. Diversidad Metabólica de Microorganismos Edáficos Asociados al Cultivo de Maíz En El Valle Del Yaqui, Sonora. Rev. Mex. Cienc. Agric. 2018, 9, 431–442. [Google Scholar]
- Ayala-Zepeda, M.; Parra-Cota, F.I.; Chinchilla-Soto, C.; De La Cruz-Torres, E.; Ibba, M.I.; Estrada-Alvarado, M.I.; de los Santos-Villalobos, S. 15N-Nitrogen Use Efficiency, Productivity, and Quality of Durum Wheat Integrating Nitrogen Management and an Indigenous Bacterial Inoculant in a Single Growing Season. Appl. Sci. 2025, 15, 1429. [Google Scholar] [CrossRef]
- Ibarra-Villarreal, A.L.; Gándara-Ledezma, A.; Godoy-Flores, A.D.; Herrera-Sepúlveda, A.; Díaz-Rodríguez, A.M.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Salt-Tolerant Bacillus Species as a Promising Strategy to Mitigate the Salinity Stress in Wheat (Triticum turgidum Subsp. Durum). J. Arid. Environ. 2021, 186, 104399. [Google Scholar] [CrossRef]
- Figueroa-Brambila, K.M.; Escalante-Beltrán, A.; Montoya-Martínez, A.C.; Díaz-Rodríguez, A.M.; López-Montoya, N.D.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Bacillus cabrialesii: Five Years of Research on a Novel Species of Biological Control and Plant Growth-Promoting Bacteria. Plants 2023, 12, 2419. [Google Scholar] [CrossRef]
- Ortega-Urquieta, M.E.; Valenzuela-Ruiz, V.; Mitra, D.; Hyder, S.; Elsheery, N.I.; Kumar Das Mohapatra, P.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Draft Genome Sequence of Priestia Sp. Strain TSO9, a Plant Growth-Promoting Bacterium Associated with Wheat (Triticum turgidum Subsp. Durum) in the Yaqui Valley, Mexico. Plants 2022, 11, 2231. [Google Scholar] [CrossRef]
- de los Santos-Villalobos, S.; Díaz-Rodríguez, A.M.; Ávila-Mascareño, M.F.; Martínez-Vidales, A.D.; Parra-Cota, F.I. COLMENA: A Culture Collection of Native Microorganisms for Harnessing the Agro-Biotechnological Potential in Soils and Contributing to Food Security. Diversity (Basel) 2021, 13, 337. [Google Scholar] [CrossRef]
- García-Montelongo, A.M.; Montoya-Martínez, A.C.; Morales-Sandoval, P.H.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Beneficial Microorganisms as a Sustainable Alternative for Mitigating Biotic Stresses in Crops. Stresses 2023, 3, 210–228. [Google Scholar] [CrossRef]
- Montoya-Martínez, A.C.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Beneficial Microorganisms in Sustainable Agriculture: Harnessing Microbes’ Potential to Help Feed the World. Plants 2022, 11, 372. [Google Scholar] [CrossRef]
- Valenzuela-Ruiz, V.; Gálvez-Gamboa, G.T.; Villa-Rodríguez, E.D.; Parra-Cota, F.I.; Santoyo, G.; de los Santos-Villalobos, S. Lipopeptides Produced by Biological Control Agents of the Genus Bacillus: A Review of Analytical Tools Used for Their Study. Rev. Mex. Cienc. Agric. 2020, 11, 419–432. [Google Scholar]
- Hyder, S.; Rizvi, Z.F.; de los Santos-Villalobos, S.; Santoyo, G.; Gondal, A.S.; Khalid, N.; Fatima, S.N.; Nadeem, M.; Rafique, K.; Rani, A. Applications of Plant Growth-Promoting Rhizobacteria for Increasing Crop Production and Resilience. J. Plant Nutr. 2023, 46, 2551–2580. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Pesticide Residues in Food; WHO: Geneva, Switzerland, 2023. [Google Scholar]
- Biedendieck, R.; Knuuti, T.; Moore, S.J.; Jahn, D. The “Beauty in the Beast”-the Multiple Uses of Priestia Megaterium in Biotechnology. Appl. Microbiol. Biotechnol. 2021, 105, 5719–5737. [Google Scholar] [CrossRef]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust Demarcation of 17 Distinct Bacillus Species Clades, Proposed as Novel Bacillaceae Genera, by Phylogenomics and Comparative Genomic Analyses: Description of Robertmurraya kyonggiensis Sp. Nov. and Proposal for an Emended Genus Bacillus Limiting It Only to the Members of the Subtilis and Cereus Clades of Species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar] [CrossRef] [PubMed]
- Almirón, C.; Petitti, T.D.; Ponso, M.A.; Romero, A.M.; Areco, V.A.; Bianco, M.I.; Espariz, M.; Yaryura, P.M. Functional and Genomic Analyses of Plant Growth Promoting Traits in Priestia aryabhattai and Paenibacillus Sp. Isolates from Tomato Rhizosphere. Sci. Rep. 2025, 15, 3498. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. AntiSMASH 7.0: New and Improved Predictions for Detection, Regulation, Chemical Structures and Visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef] [PubMed]
- Dhar, S.K.; Kaur, J.; Singh, G.B.; Chauhan, A.; Tamang, J.; Lakhara, N.; Asyakina, L.; Atuchin, V.; Mudgal, G.; Abdi, G. Novel Bacillus and Prestia Isolates from Dwarf Century Plant Enhance Crop Yield and Salinity Tolerance. Sci. Rep. 2024, 14, 14645. [Google Scholar] [CrossRef]
- Radjasa, O.K.; Steven, R.; Natanael, Y.; Nugrahapraja, H.; Radjasa, S.K.; Kristianti, T.; Moeis, M.R.; Trinugroho, J.P.; Suharya, H.B.; Rachmatsyah, A.O.; et al. From the Depths of the Java Trench: Genomic Analysis of Priestia Flexa JT4 Reveals Bioprospecting and Lycopene Production Potential. BMC Genom. 2024, 25, 1259. [Google Scholar] [CrossRef]
- Cui, Z.; Hu, L.; Zeng, L.; Meng, W.; Guo, D.; Sun, L. Isolation and Characterization of Priestia Megaterium KD7 for the Biological Control of Pear Fire Blight. Front. Microbiol. 2023, 14, 1099664. [Google Scholar] [CrossRef] [PubMed]
- Wagh, V.S.; Ram, H.; Dastager, S.G. Priestia veravalensis Sp. Nov., Isolated from Coastal Sample. Arch. Microbiol. 2021, 203, 4839–4845. [Google Scholar] [CrossRef] [PubMed]
- Robles-Montoya, R.I.; Valenzuela-Ruiz, V.; Parra-Cota, F.I.; Santoyo, G.; de los Santos-Villalobos, S. Description of a Polyphasic Taxonomic Approach for Plant Growth-Promoting Rhizobacteria (PGPR). In Microbial Services in Restoration Ecology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 259–269. ISBN 9780128199787. [Google Scholar]
- Morales-Sandoval, P.H.; Valenzuela-Ruiz, V.; Ortega-Urquieta, M.E.; Martínez-Vidales, A.D.; Félix-Pablos, C.M.; Chávez-Luzanía, R.A.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Taxonomía Bacteriana Basada En Índices Relacionados al Genoma Completo. La Sociedad Académica 2021, 58, 39–50. [Google Scholar]
- Rode, L.J. Correlation Between Spore Structure and Spore Properties in Bacillus Megaterium. J. Bacteriol. 1968, 95, 1979–1986. [Google Scholar] [CrossRef]
- Freer, J.H.; Levlnson, H.S. Fine Structure of Bacillus Megaterium during Microcycle Sporogenesis. J. Bacteriol. 1967, 94, 441–457. [Google Scholar] [CrossRef]
- Zarazúa-Osorio, B.; Srivastava, P.; Marathe, A.; Zahid, S.H.; Fujita, M. Autoregulation of the Master Regulator Spo0A Controls Cell-Fate Decisions in Bacillus Subtilis. Mol. Microbiol. 2025, 123, 305–329. [Google Scholar] [CrossRef]
- Fujita, M.; Losick, R. Evidence That Entry into Sporulation in Bacillus Subtilis Is Governed by a Gradual Increase in the Level and Activity of the Master Regulator Spo0A. Genes Dev. 2005, 19, 2236–2244. [Google Scholar] [CrossRef]
- Hilbert, D.W.; Piggot, P.J. Compartmentalization of Gene Expression during Bacillus Subtilis Spore Formation. Microbiol. Mol. Biol. Rev. 2004, 68, 234–262. [Google Scholar] [CrossRef]
- Marathe, A.; Zarazúa-Osorio, B.; Srivastava, P.; Fujita, M. The Master Regulator for Entry into Sporulation in Bacillus Subtilis Becomes a Mother Cell-Specific Transcription Factor for Forespore Engulfment. Mol. Microbiol. 2023, 120, 439–461. [Google Scholar] [CrossRef]
- Chastanet, A.; Vitkup, D.; Yuan, G.C.; Norman, T.M.; Liu, J.S.; Losick, R.M. Broadly Heterogeneous Activation of the Master Regulator for Sporulation in Bacillus Subtilis. Proc. Natl. Acad. Sci. USA 2010, 107, 8486–8491. [Google Scholar] [CrossRef]
- Chastanet, A.; Losick, R. Just-in-Time Control of Spo0A Synthesis in Bacillus Subtilis by Multiple Regulatory Mechanisms. J. Bacteriol. 2011, 193, 6366–6374. [Google Scholar] [CrossRef]
- Iber, D. A Quantitative Study of the Benefits of Co-Regulation Using the SpoIIA Operon as an Example. Mol. Syst. Biol. 2006, 2, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Goswami, G.; Panda, D.; Samanta, R.; Boro, R.C.; Modi, M.K.; Bujarbaruah, K.M.; Barooah, M. Bacillus Megaterium Adapts to Acid Stress Condition through a Network of Genes: Insight from a Genome-Wide Transcriptome Analysis. Sci. Rep. 2018, 8, 16105. [Google Scholar] [CrossRef]
- Fawcett, P.; Eichenberger, P.; Losick, R.; Youngman, P. The Transcriptional Profile of Early to Middle Sporulation in Bacillus Subtilis. Proc. Natl. Acad. Sci. USA 2000, 97, 8063–8068. [Google Scholar] [CrossRef]
- Molle, V.; Fujita, M.; Jensen, S.T.; Eichenberger, P.; González-Pastor, J.E.; Liu, J.S.; Losick, R. The Spo0A Regulon of Bacillus Subtilis. Mol. Microbiol. 2003, 50, 1683–1701. [Google Scholar] [CrossRef] [PubMed]
- Reder, A.; Gerth, U.; Hecker, M. Integration of ΣB Activity into the Decision-Making Process of Sporulation Initiation in Bacillus Subtilis. J. Bacteriol. 2012, 194, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Méndez, M.B.; Orsaria, L.M.; Philippe, V.; Pedrido, M.E.; Grau, R.R. Novel Roles of the Master Transcription Factors Spo0A and σ B for Survival and Sporulation of Bacillus Subtilis at Low Growth Temperature. J. Bacteriol. 2004, 186, 989–1000. [Google Scholar] [CrossRef]
- Abhyankar, W.R.; Kamphorst, K.; Swarge, B.N.; van Veen, H.; van der Wel, N.N.; Brul, S.; de Koster, C.G.; de Koning, L.J. The Influence of Sporulation Conditions on the Spore Coat Protein Composition of Bacillus Subtilis Spores. Front. Microbiol. 2016, 7, 1636. [Google Scholar] [CrossRef]
- Saggese, A.; Scamardella, V.; Sirec, T.; Cangiano, G.; Isticato, R.; Pane, F.; Amoresano, A.; Ricca, E.; Baccigalupi, L. Antagonistic Role of CotG and CotH on Spore Germination and Coat Formation in Bacillus Subtilis. PLoS ONE 2014, 9, e104900. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, J.; Du, G.; Zhou, J.; Chen, J. Sporulation and Spore Stability of Bacillus Megaterium Enhance Ketogulonigenium Vulgare Propagation and 2-Keto-l-Gulonic Acid Biosynthesis. Bioresour. Technol. 2012, 107, 399–404. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Dahar, G.Y.; Wang, H.W.; Rajer, F.U.; Jin, P.; Xu, P.; Abro, M.A.; Qureshi, A.S.; Karim, A.; Miao, W. Comparative Genomic Analysis of Bacillus Atrophaeus HAB-5 Reveals Genes Associated with Antimicrobial and Plant Growth-Promoting Activities. Front. Microbiol. 2024, 15, 1384691. [Google Scholar] [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial Iron Homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Miethke, M.; Marahiel, M.A. Siderophore-Based Iron Acquisition and Pathogen Control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef]
- Miethke, M.; Monteferrante, C.G.; Marahiel, M.A.; van Dijl, J.M. The Bacillus Subtilis EfeUOB Transporter Is Essential for High-Affinity Acquisition of Ferrous and Ferric Iron. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 2267–2278. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Benaki, D.; Koukounia, M.; Zervakou, A.; Tzima, A.; Diallinas, G.; Hatzinikolaou, D.G.; Skandalis, N. Direct Antibiotic Activity of Bacillibactin Broadens the Biocontrol Range of Bacillus Amyloliquefaciens MBI600. mSphere 2021, 6, e0037621. [Google Scholar] [CrossRef]
- Nakatsuji, S.; Okumura, K.; Takase, R.; Watanabe, D.; Mikami, B.; Hashimoto, W. Crystal Structures of EfeB and EfeO in a Bacterial Siderophore-Independent Iron Transport System. Biochem. Biophys. Res. Commun. 2022, 594, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S.C. Phosphate-Solubilizing Microorganisms: Mechanism and Their Role in Phosphate Solubilization and Uptake. J. Soil Sci. Plant Nutr. 2021, 21, 49–68. [Google Scholar] [CrossRef]
- Feng, Y.; He, J.; Zhang, H.; Jia, X.; Hu, Y.; Ye, J.; Gu, X.; Zhang, X.; Chen, H. Phosphate Solubilizing Microorganisms: A Sustainability Strategy to Improve Urban Ecosystems. Front. Microbiol. 2023, 14, 1320853. [Google Scholar] [CrossRef]
- Guo, L.; Wang, C.; Shen, R.F. Stronger Effects of Maize Rhizosphere than Phosphorus Fertilization on Phosphatase Activity and Phosphorus-Mineralizing-Related Bacteria in Acidic Soils. Rhizosphere 2022, 23, 1320853. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, G.; Chen, H.; Chen, C.; Wang, J.; Ai, S.; Wei, D.; Li, D.; Ma, B.; Tang, C.; et al. Long-Term Nutrient Inputs Shift Soil Microbial Functional Profiles of Phosphorus Cycling in Diverse Agroecosystems. ISME J. 2020, 14, 757–770. [Google Scholar] [CrossRef]
- Billah, M.; Khan, M.; Bano, A.; Hassan, T.U.; Munir, A.; Gurmani, A.R. Phosphorus and Phosphate Solubilizing Bacteria: Keys for Sustainable Agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Tang, J.; Li, Y.; Zhang, L.; Mu, J.; Jiang, Y.; Fu, H.; Zhang, Y.; Cui, H.; Yu, X.; Ye, Z. Biosynthetic Pathways and Functions of Indole-3-Acetic Acid in Microorganisms. Microorganisms 2023, 11, 2077. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, K. Auxin Herbicides: Current Status of Mechanism and Mode of Action. Pest. Manag. Sci. 2010, 66, 113–120. [Google Scholar] [CrossRef]
- Ibarra-Villarreal, A.L.; Isela Parra-Cota, F.; Yépez, E.A.; Gutiérrez-Coronado, M.A.; Valdez-Torres, L.C.; de los Santos-Villalobos, S. Impacto Del Cambio En El Manejo Del Cultivo de Trigo de convencional a Orgánico Sobre Las Comunidades Fúngicas Del Suelo En El Valle Del Yaqui, México. Agrociencia 2020, 54, 643–659. [Google Scholar] [CrossRef]
- Córdova-Albores, L.C.; Zelaya-Molina, L.X.; Ávila-Alistac, N.; Valenzuela-Ruiz, V.; Cortés-Martínez, N.E.; Parra-Cota, F.I.; Burgos-Canul, Y.Y.; Chávez-Díaz, I.F.; Fajardo-Franco, M.L.; de los Santos-Villalobos, S. Omics Sciences Potential on Bioprospecting of Biological Control Microbial Agents: The Case of the Mexican Agro-Biotechnology. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2020, 39, 147–184. [Google Scholar] [CrossRef]
- Hoffmann, T.; Wensing, A.; Brosius, M.; Steil, L.; Völker, U.; Bremer, E. Osmotic Control of OpuA Expression in Bacillus Subtilis and Its Modulation in Response to Intracellular Glycine Betaine and Proline Pools. J. Bacteriol. 2013, 195, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Gregory, G.J.; Boyd, E.F. Stressed out: Bacterial Response to High Salinity Using Compatible Solute Biosynthesis and Uptake Systems, Lessons from Vibrionaceae. Comput. Struct. Biotechnol. J. 2021, 19, 1014–1027. [Google Scholar] [CrossRef]
- Gazzaniga, F.; Stebbins, R.; Chang, S.Z.; McPeek, M.A.; Brenner, C. Microbial NAD Metabolism: Lessons from Comparative Genomics. Microbiol. Mol. Biol. Rev. 2009, 73, 529–541. [Google Scholar] [CrossRef]
- Ito, M.; Morino, M.; Krulwich, T.A. Mrp Antiporters Have Important Roles in Diverse Bacteria and Archaea. Front. Microbiol. 2017, 8, 2325. [Google Scholar] [CrossRef]
- Kosono, S.; Haga, K.; Tomizawa, R.; Kajiyama, Y.; Hatano, K.; Takeda, S.; Wakai, Y.; Hino, M.; Kudo, T. Characterization of a Multigene-Encoded Sodium/Hydrogen Antiporter (Sha) from Pseudomonas Aeruginosa: Its Involvement in Pathogenesis. J. Bacteriol. 2005, 187, 5242–5248. [Google Scholar] [CrossRef] [PubMed]
- Kredich, N.M. Biosynthesis of Cysteine. EcoSal Plus 2008, 3, 10–1128. [Google Scholar] [CrossRef]
- Kirstein, J.; Molière, N.; Dougan, D.A.; Turgay, K. Adapting the Machine: Adaptor Proteins for Hsp100/Clp and AAA+ Proteases. Nat. Rev. Microbiol. 2009, 7, 589–599. [Google Scholar] [CrossRef]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Wieland Brown, L.C.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into Secondary Metabolism from a Global Analysis of Prokaryotic Biosynthetic Gene Clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Kottmann, R.; Yilmaz, P.; Cummings, M.; Biggins, J.B.; Blin, K.; De Bruijn, I.; Chooi, Y.H.; Claesen, J.; Coates, R.C.; et al. Minimum Information about a Biosynthetic Gene Cluster. Nat. Chem. Biol. 2015, 11, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Semenzato, G.; Bernacchi, A.; Amata, S.; Bechini, A.; Berti, F.; Calonico, C.; Catania, V.; Esposito, A.; Puglia, A.M.; Piccionello, A.P.; et al. Antibacterial Properties of Bacterial Endophytes Isolated from the Medicinal Plant Origanum heracleoticum L. Front. Biosci.- Landmark 2024, 29, 111. [Google Scholar] [CrossRef]
- Lam, V.B.; Ibrahim, H.M.M.; Oni, F.E.; Argüelles-Arias, A.; Marahatta, B.; Zhou, L.; Ferrarini, E.; De Coninck, B.; Cottyn, B.; Ongena, M.; et al. Diversity of Bacillaceae on Rice Grown in Acid Sulfate Soils in Vietnam: Taxonomy, Specialized Metabolites, and Inhibitory Effects on Fungal Pathogens. Phytobiomes J. 2024, 8, 469–483. [Google Scholar] [CrossRef]
- Khalifa, A.; Alsowayeh, N. Whole-Genome Sequence Insight into the Plant-Growth-Promoting Bacterium Priestia Filamentosa Strain AZC66 Obtained from Zygophyllum Coccineum Rhizosphere. Plants 2023, 12, 1944. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhu, X.; Qian, T.; Du, J.; Du, Y.; Ye, J. Mechanism of Salt Tolerance and Plant Growth Promotion in Priestia Megaterium ZS-3 Revealed by Cellular Metabolism and Whole-Genome Studies. Int. J. Mol. Sci. 2023, 24, 15751. [Google Scholar] [CrossRef]
- Jo, H.W.; Lim, K.; Ibal, J.C.; Kim, M.C.; Kim, H.B.; Baek, C.; Heo, Y.M.; Lee, H.; Kang, S.; Lee, D.G.; et al. Growth Increase in the Herbaceous Plant Centella Asiatica by the Plant Growth-Promoting Rhizobacteria Priestia Megaterium HyangYak-01. Plants 2023, 12, 2398. [Google Scholar] [CrossRef]
- Bhat, M.A.; Mishra, A.K.; Jan, S.; Bhat, M.A.; Kamal, M.A.; Rahman, S.; Shah, A.A.; Jan, A.T. Plant Growth Promoting Rhizobacteria in Plant Health: A Perspective Study of the Underground Interaction. Plants 2023, 12, 629. [Google Scholar] [CrossRef] [PubMed]
- Agunbiade, V.F.; Fadiji, A.E.; Agbodjato, N.A.; Babalola, O.O. Isolation and Characterization of Plant-Growth-Promoting, Drought-Tolerant Rhizobacteria for Improved Maize Productivity. Plants 2024, 13, 1298. [Google Scholar] [CrossRef]
- Hwang, H.H.; Chien, P.R.; Huang, F.C.; Yeh, P.H.; Hung, S.H.W.; Deng, W.L.; Huang, C.C. A Plant Endophytic Bacterium Priestia Megaterium StrainBP-R2 Isolated from the Halophyte Bolboschoenus Planiculmis Enhances Plant Growth under Salt and Drought Stresses. Microorganisms 2022, 10, 2047. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, A.B.; Fulton, M.D. A Simplified Method of Staining Endospores. Science (1979) 1933, 77, 194. [Google Scholar] [CrossRef]
- Moirangthem Bidyaswori, D.; Dhar, R.; Bhattacharjee, A.; Dibyendu, P. Screening of Different Media and Heat Shock Treatment Regimens for Enhancing Sporulation in Bacillus Licheniformis. Int. J. Curr. Sci. Res. Rev. 2023, 6, 2666–2674. [Google Scholar] [CrossRef]
- Valenzuela-Aragón, B.; Montoya-Martínez, A.C.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Genomic Insight into a Potential Biological Control Agent for Fusarium-Related Diseases in Potatoes: Bacillus Cabrialesii Subsp. Cabrialesii Strain PE1. Horticulturae 2024, 10, 357. [Google Scholar] [CrossRef]
- Campos-Avelar, I.; Montoya-Martínez, A.C.; Escalante-Beltrán, A.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Do Organic Amendments Foster Only Beneficial Bacteria in Agroecosystems?: The Case of Bacillus Paranthracis TSO55. Plants 2025, 14, 1019. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple Alignment of Conserved Genomic Sequence with Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S RRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Lee, I.; Kim, Y.O.; Park, S.C.; Chun, J. OrthoANI: An Improved Algorithm and Software for Calculating Average Nucleotide Identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.-M.; Lim, J.; Kwon, S.; Chun, J. A Large-Scale Evaluation of Algorithms to Calculate Average Nucleotide Identity. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M.G. Genome Sequence-Based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A Database Tandem for Fast and Reliable Genome-Based Classification and Nomenclature of Prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Phillippy, A.; Delcher, A.L.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S.L. Versatile and Open Software for Comparing Large Genomes. Genome Biol. 2004, 5, 12. [Google Scholar] [CrossRef]
- Carattoli, A.; Hasman, H. Identification and Typing of Plasmid Replicons in Whole-Genome Sequencing (WGS). In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2020; Volume 2075, pp. 285–294. [Google Scholar]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.Y.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-Depth Characterization and Visualization of Bacterial Genomes. Nucleic Acids Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Patz, S.; Gautam, A.; Becker, M.; Ruppel, S.; Rodríguez-Palenzuela, P.; Huson, D.H. PLaBAse: A Comprehensive Web Resource for Analyzing the Plant Growth-Promoting Potential of Plant-Associated Bacteria. bioRxiv 2021. [Google Scholar] [CrossRef]
- de los Santos-Villalobos, S.; de Folter, S.; Délano-Frier, J.P.; Gómez-Lim, M.A.; Guzmán-Ortiz, D.A.; Peña-Cabriales, J.J. Growth Promotion and Flowering Induction in Mango (Mangifera Indica L. Cv “Ataulfo”) Trees by Burkholderia and Rhizobium Inoculation: Morphometric, Biochemical, and Molecular Events. J. Plant Growth Regul. 2013, 32, 615–627. [Google Scholar] [CrossRef]
- Onyia, C.E.; Anyanwu, C.U. Comparative Study on Solubilization of Tri-Calcium Phosphate (TCP) by Phosphate Solubilizing Fungi (PSF) Isolated from Nsukka Pepper Plant Rhizosphere and Root Free Soil. J. Yeast Fungal Res. 2013, 4, 52–57. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of Chrome Azurol S Reagents to Evaluate Siderophore Production by Rhizosphere Bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Meléndez-García, M.; Zárate-Camargo, G.; Meza-Contreras, J.J.; Herrera-Sepúlveda, A.; de los Santos-Villalobos, S.; Parra-Cota, F.I. Abiotic Stress Tolerance of Microorganisms Associated with Oregano (Origanum vulgare L.) in the Yaqui Valley, Sonora. Open Agric. 2017, 2, 260–265. [Google Scholar] [CrossRef]
- Valenzuela-Aragón, B.; Parra-Cota, F.I.; Santoyo, G.; Arellano-Wattenbarger, G.L.; de los Santos-Villalobos, S. Plant-Assisted Selection: A Promising Alternative for in Vivo Identification of Wheat (Triticum turgidum L. Subsp. Durum) Growth Promoting Bacteria. Plant Soil. 2019, 435, 367–384. [Google Scholar] [CrossRef]
- Morales-Sandoval, P.H.; Valenzuela-Ruiz, V.; Santoyo, G.; Hyder, S.; Mitra, D.; Zelaya-Molina, L.X.; Ávila-Alistac, N.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Draft Genome of a Biological Control Agent against Bipolaris Sorokiniana, the Causal Phytopathogen of Spot Blotch in Wheat (Triticum turgidum L. Subsp. Durum): Bacillus Inaquosorum TSO22. Open Agric. 2024, 9, 20220309. [Google Scholar] [CrossRef]
- Villa-Rodríguez, E.; Parra-Cota, F.; Castro-Longoria, E.; López-Cervantes, J.; de los Santos-Villalobos, S. Bacillus Subtilis TE3: A Promising Biological Control Agent against Bipolaris Sorokiniana, the Causal Agent of Spot Blotch in Wheat (Triticum turgidum L. Subsp. Durum). Biol. Control 2019, 132, 135–143. [Google Scholar] [CrossRef]
- Rojas-Padilla, J.; Chaparro-Encinas, L.A.; Robles-Montoya, R.I.; de los Santos-Villalobos, S. Growth Promotion on Wheat (Triticum turgidum L. Subsp. Durum) by Co-Inoculation of Native Bacillus Strains Isolated from the Yaqui Valley, Mexico. Nova Scientia 2020, 12. Available online: https://www.scielo.org.mx/scielo.php?pid=S2007-07052020000100006&script=sci_abstract&tlng=en (accessed on 13 February 2025). [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Name | Strain | GenBank Accession Number | 16S Similarity (%) | Strain | ANIb | ANIm | OrthoANI | GGDC (Formula 2) |
|---|---|---|---|---|---|---|---|---|
| Priestia megaterium | NBRC 15308T | JJMH01000057 | 99.92 | ATCC 14581T | 95.49 | 96.7 | 96.51 | 76.58 |
| Priestia aryabhattai | B8W22T | EF114313 | 99.76 | B8W22T | 94.78 | 95.7 | 95.67 | 76.01 |
| Priestia flexa | NBRC 15715T | BCVD01000224 | 99.04 | NBRC 15715T | 73.43 | 84.62 | 74.43 | 0 |
| Uninoculated Maize (Control) | Inoculated Maize (Priestia megaterium 53B2) | Increment vs. Control (%) | |
|---|---|---|---|
| Stem height (cm) | 10.55 ± 1.1 | 12.96 ± 1.27 * | 22.8% |
| Root length (cm) | 9.07 ± 1.17 | 12.31 ± 2.25 * | 35.2% |
| Stem weight (mg) | 418.8 ± 80 | 584.60 ± 70 * | 39.6% |
| Root weight (mg) | 313.6 ± 140 | 520.80 ± 80 * | 66.1% |
| Stem dry weight (mg) | 52.91 ± 14 | 70.72 ± 9 * | 33.7% |
| Root dry weight (mg) | 54.52 ± 0.15 | 78.91 ± 0.09 * | 44.7% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escalante-Beltrán, A.; Morales-Sandoval, P.H.; González-Astorga, C.B.; Montoya-Martínez, A.C.; Cubedo-Ruiz, E.A.; Santoyo, G.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Genomic Insights and Plant Growth-Promoting Characterization of Priestia megaterium Strain 53B2, Isolated from Maize-Associated Soil in the Yaqui Valley, Mexico. Plants 2025, 14, 2081. https://doi.org/10.3390/plants14132081
Escalante-Beltrán A, Morales-Sandoval PH, González-Astorga CB, Montoya-Martínez AC, Cubedo-Ruiz EA, Santoyo G, Parra-Cota FI, de los Santos-Villalobos S. Genomic Insights and Plant Growth-Promoting Characterization of Priestia megaterium Strain 53B2, Isolated from Maize-Associated Soil in the Yaqui Valley, Mexico. Plants. 2025; 14(13):2081. https://doi.org/10.3390/plants14132081
Chicago/Turabian StyleEscalante-Beltrán, Alina, Pamela Helué Morales-Sandoval, Claudia Berenice González-Astorga, Amelia C. Montoya-Martínez, Edgar A. Cubedo-Ruiz, Gustavo Santoyo, Fannie Isela Parra-Cota, and Sergio de los Santos-Villalobos. 2025. "Genomic Insights and Plant Growth-Promoting Characterization of Priestia megaterium Strain 53B2, Isolated from Maize-Associated Soil in the Yaqui Valley, Mexico" Plants 14, no. 13: 2081. https://doi.org/10.3390/plants14132081
APA StyleEscalante-Beltrán, A., Morales-Sandoval, P. H., González-Astorga, C. B., Montoya-Martínez, A. C., Cubedo-Ruiz, E. A., Santoyo, G., Parra-Cota, F. I., & de los Santos-Villalobos, S. (2025). Genomic Insights and Plant Growth-Promoting Characterization of Priestia megaterium Strain 53B2, Isolated from Maize-Associated Soil in the Yaqui Valley, Mexico. Plants, 14(13), 2081. https://doi.org/10.3390/plants14132081