
Physiological and Transcriptomic Responses of Bok Choy to Heat Stress
 , ,
, ,
Abstract
1. Introduction
2. Results
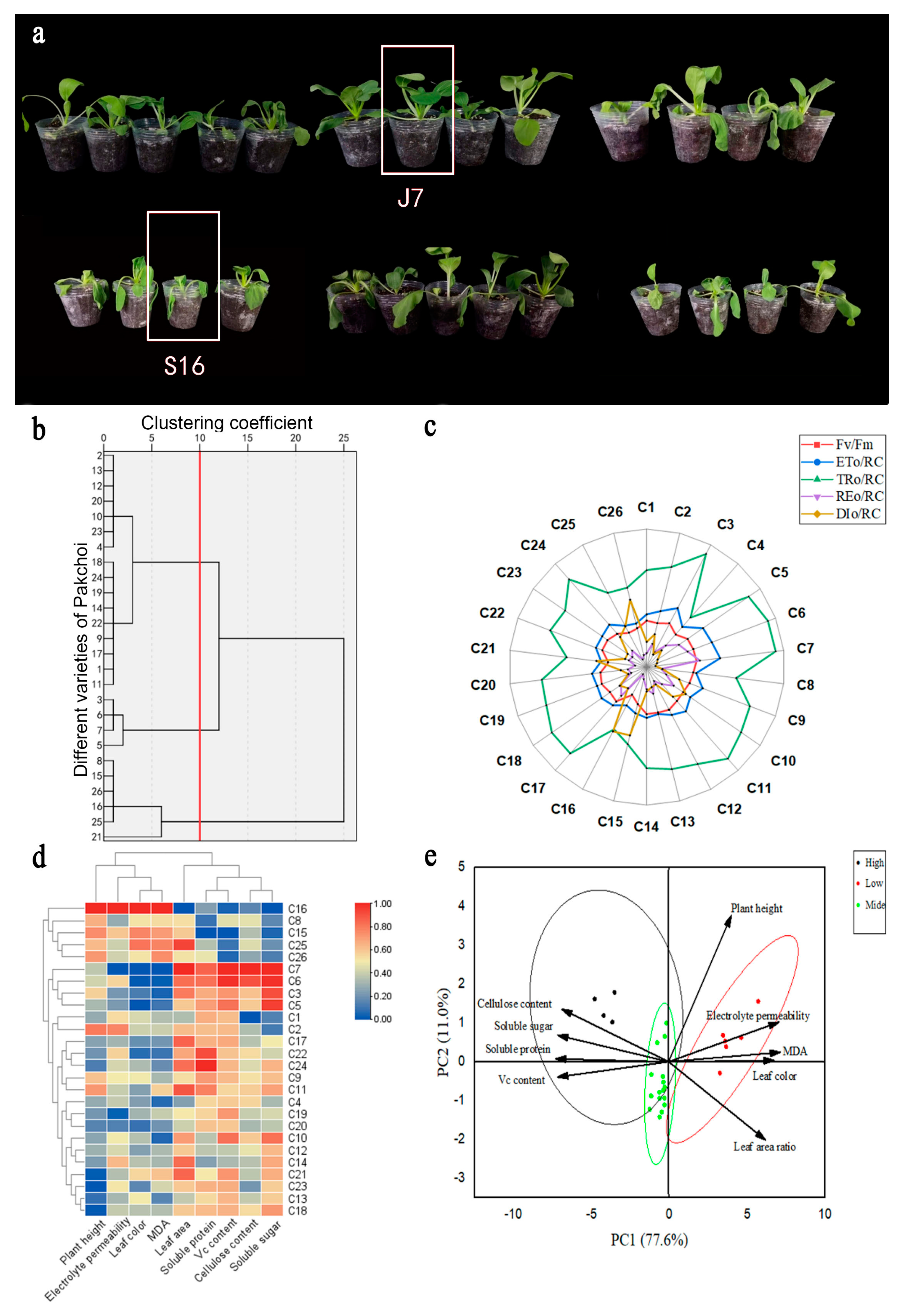
2.1. Evaluation of Heat Tolerance of Different Bok choy Varieties
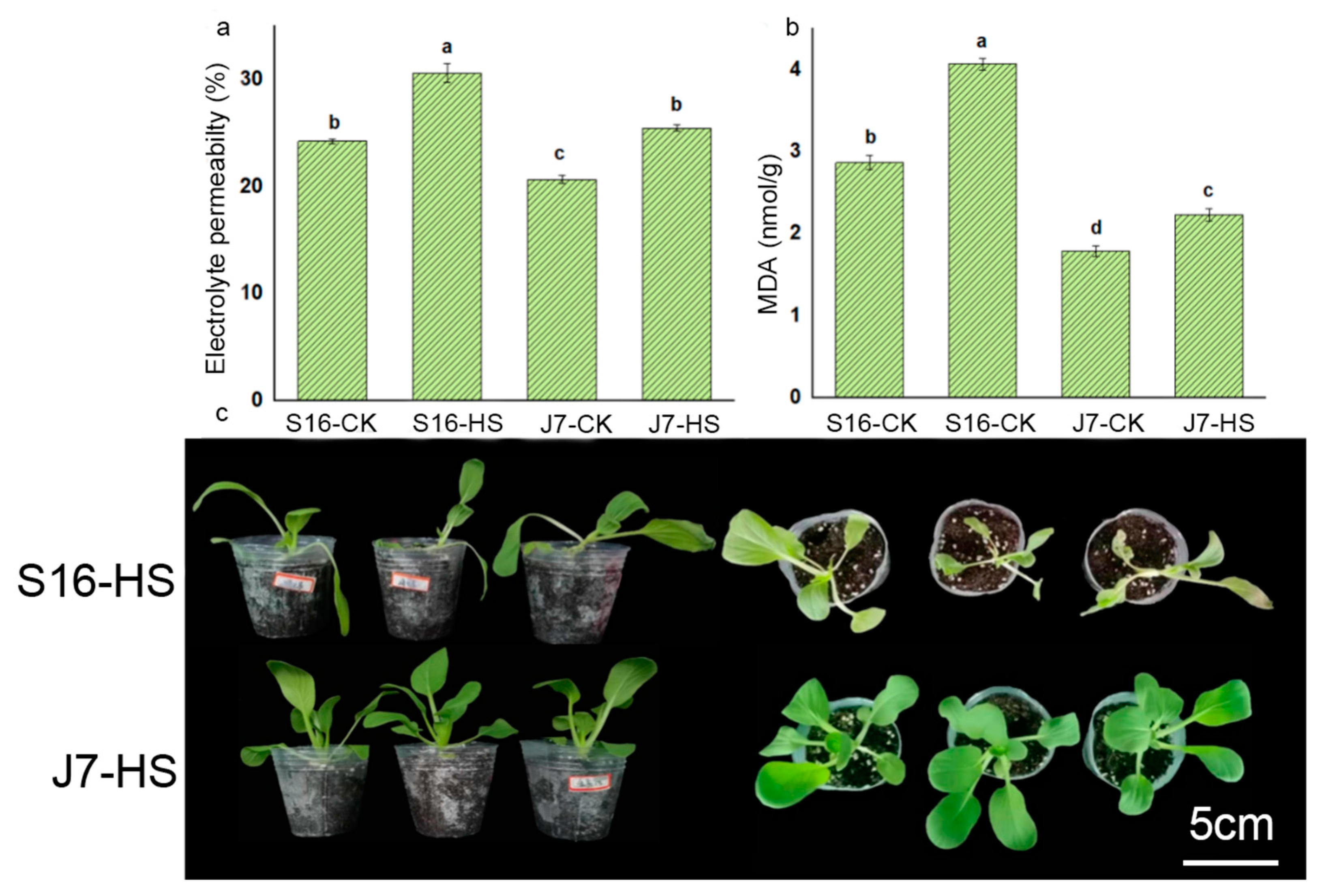
2.2. Agronomic Characteristics under Heat Stress
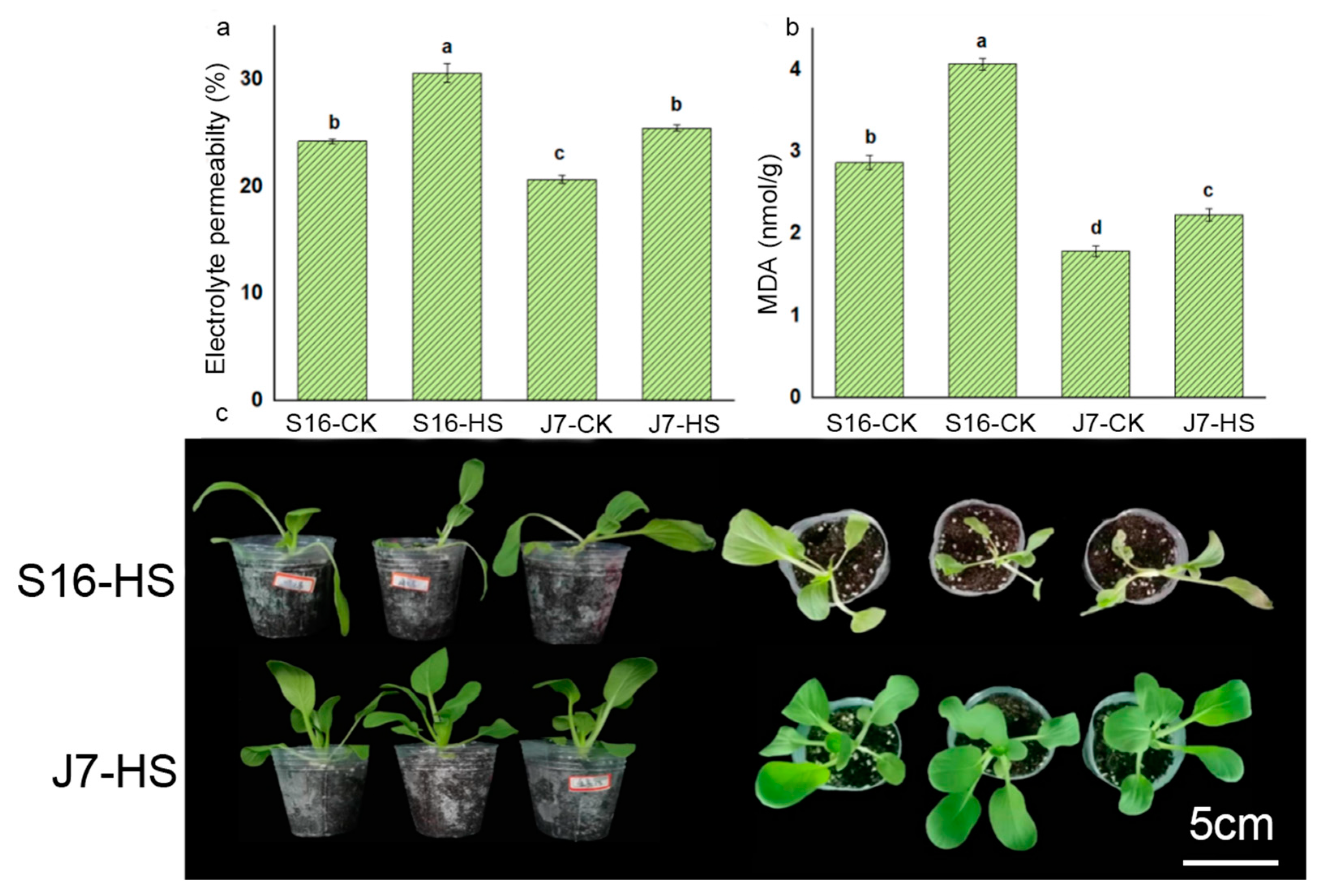
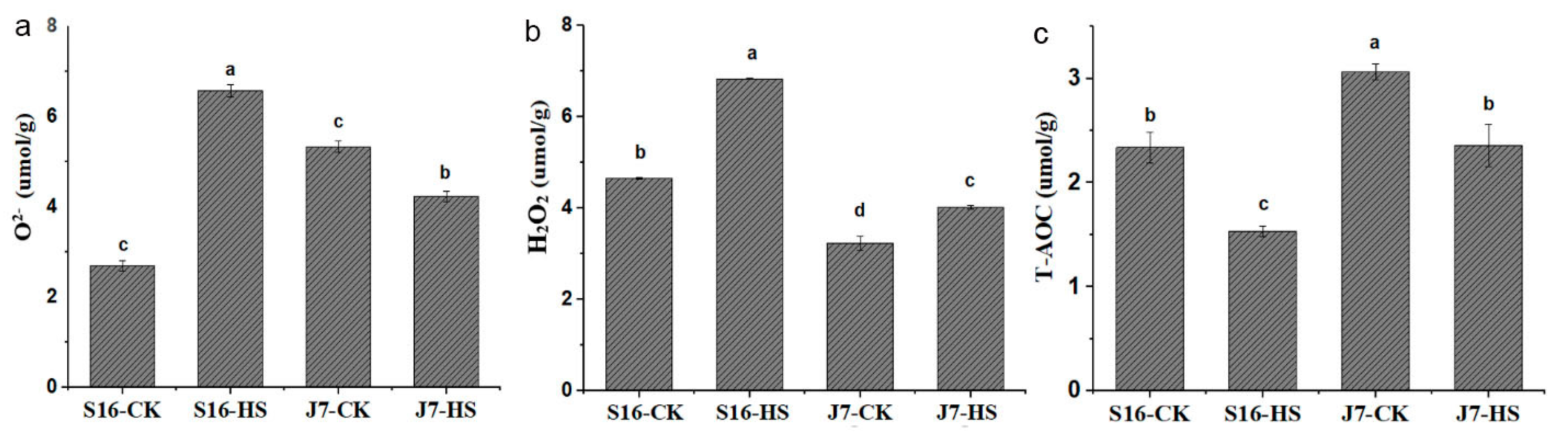
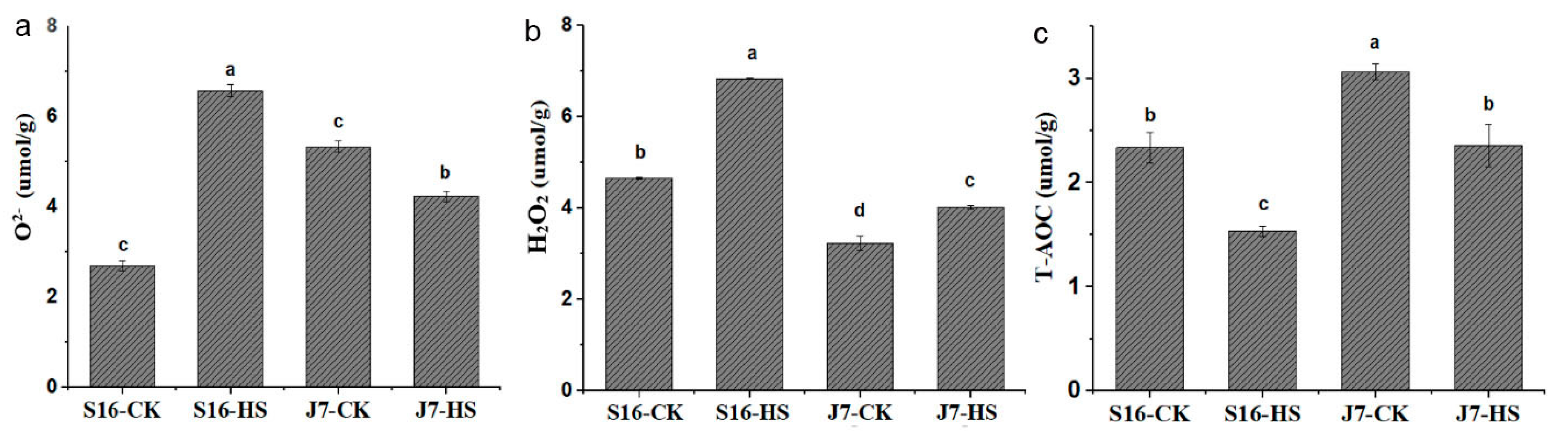
2.3. Physiological and Biochemical Indices under Heat Stress
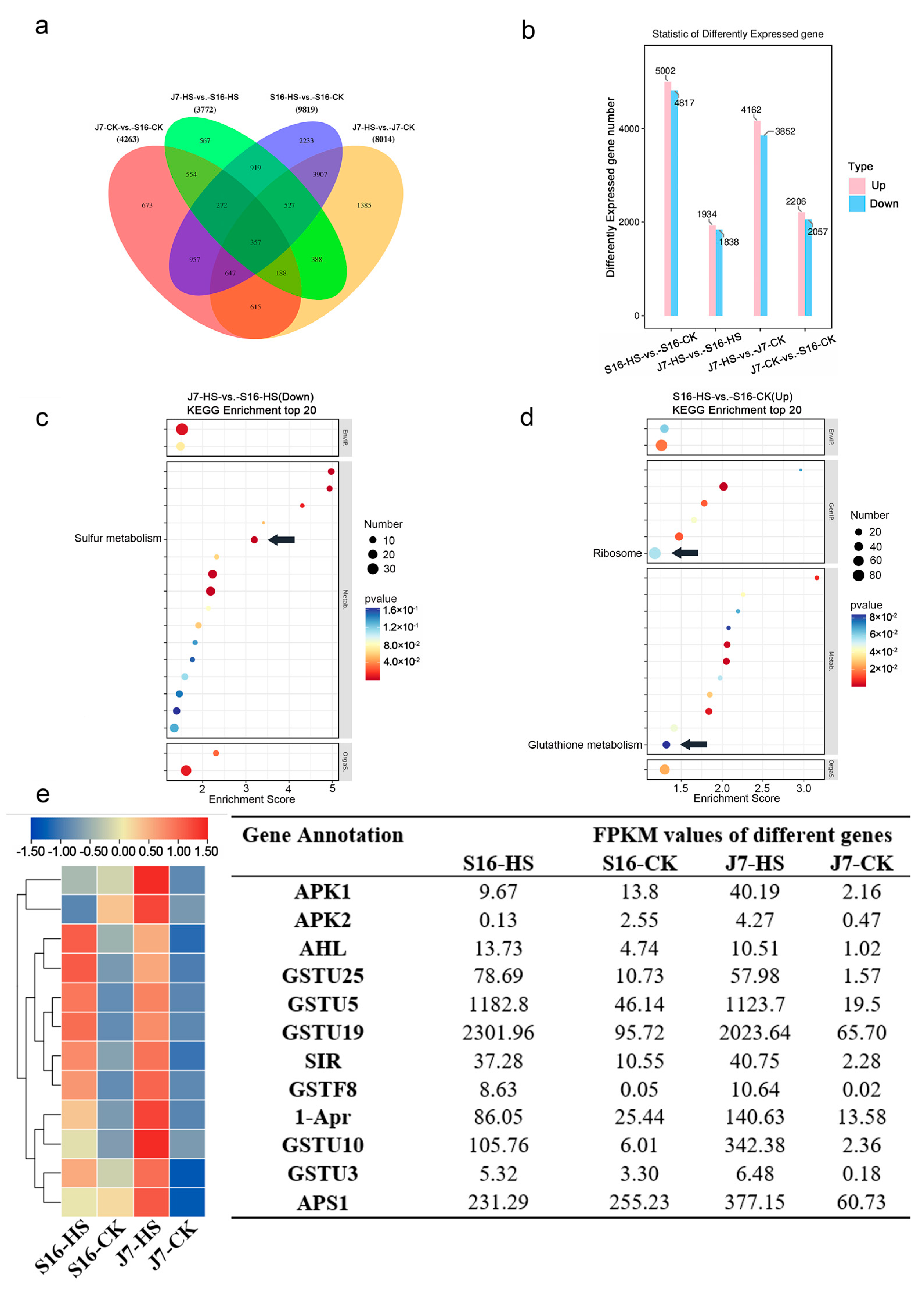
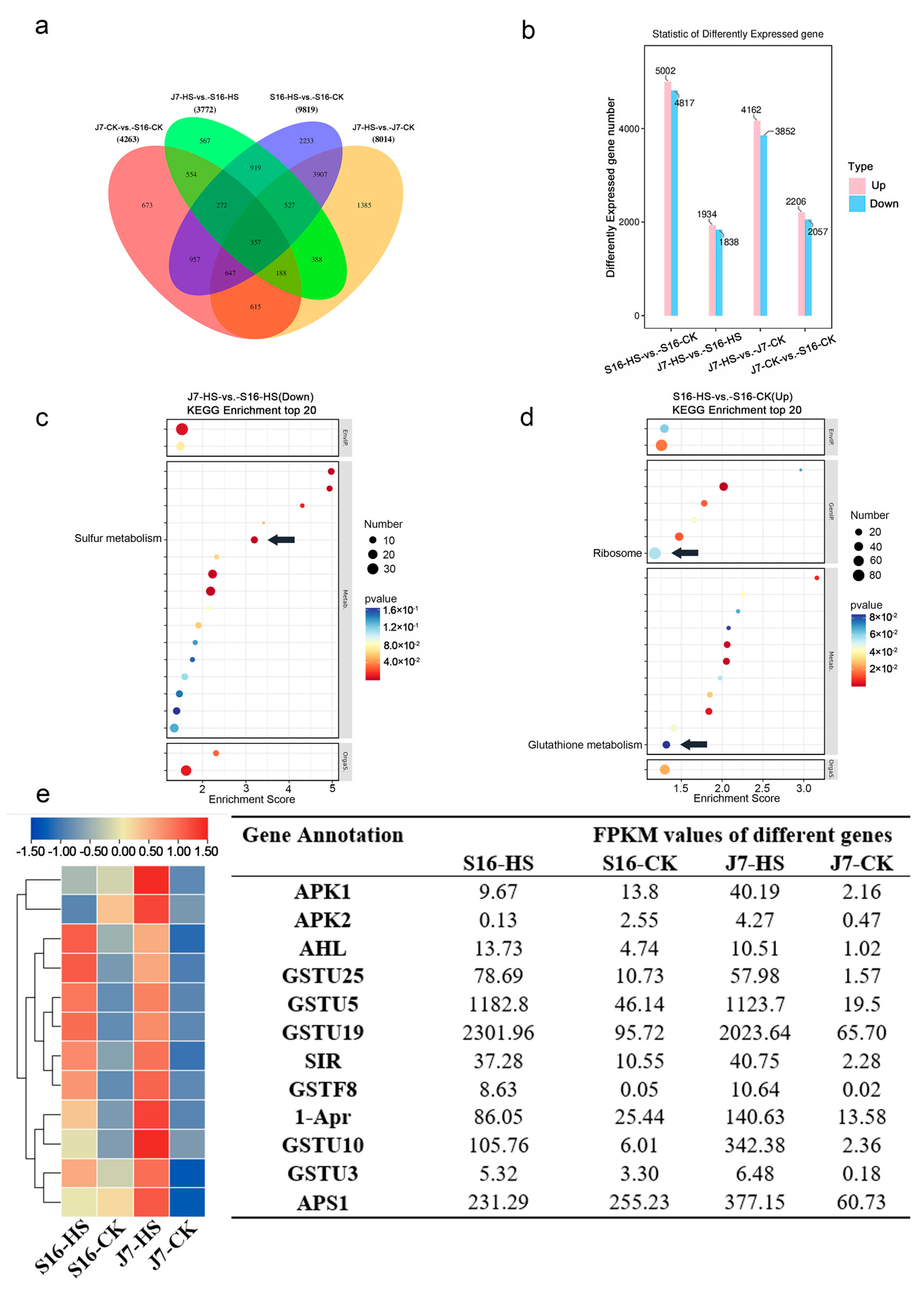
2.4. Identification of Differentially Expressed Genes by Transcriptome Sequencing
2.5. Identification and Enrichment Analysis of DEGs
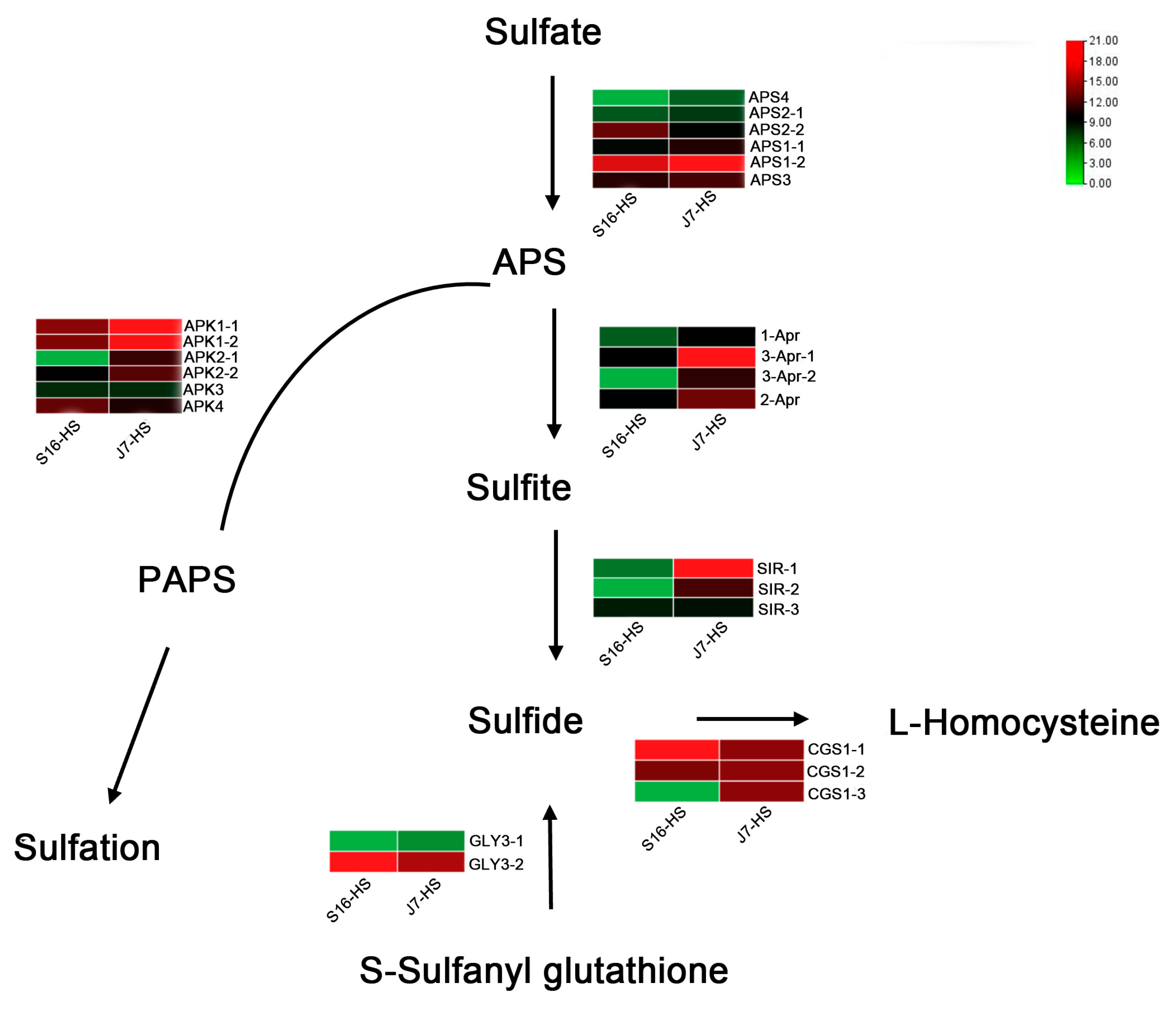
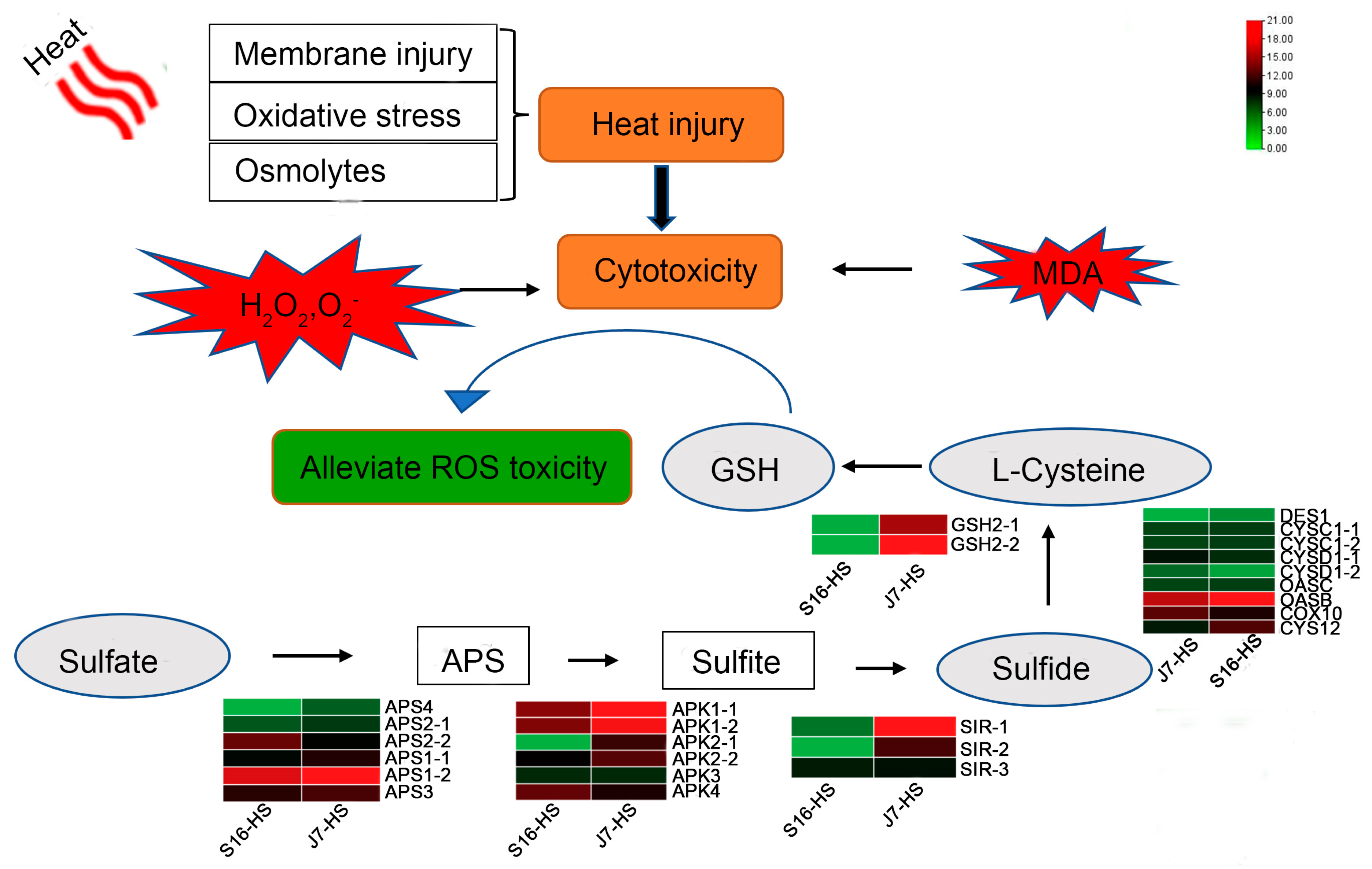
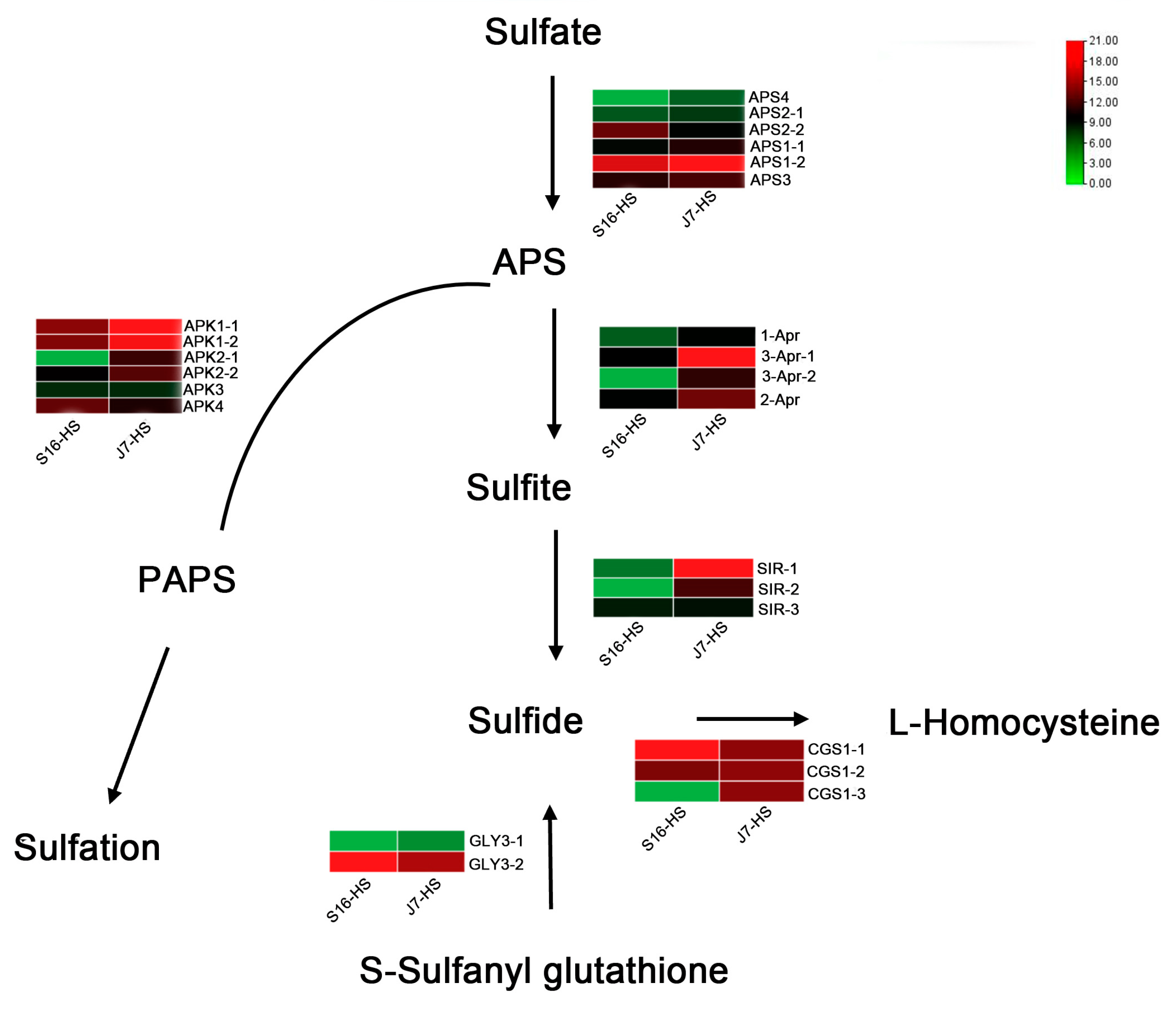
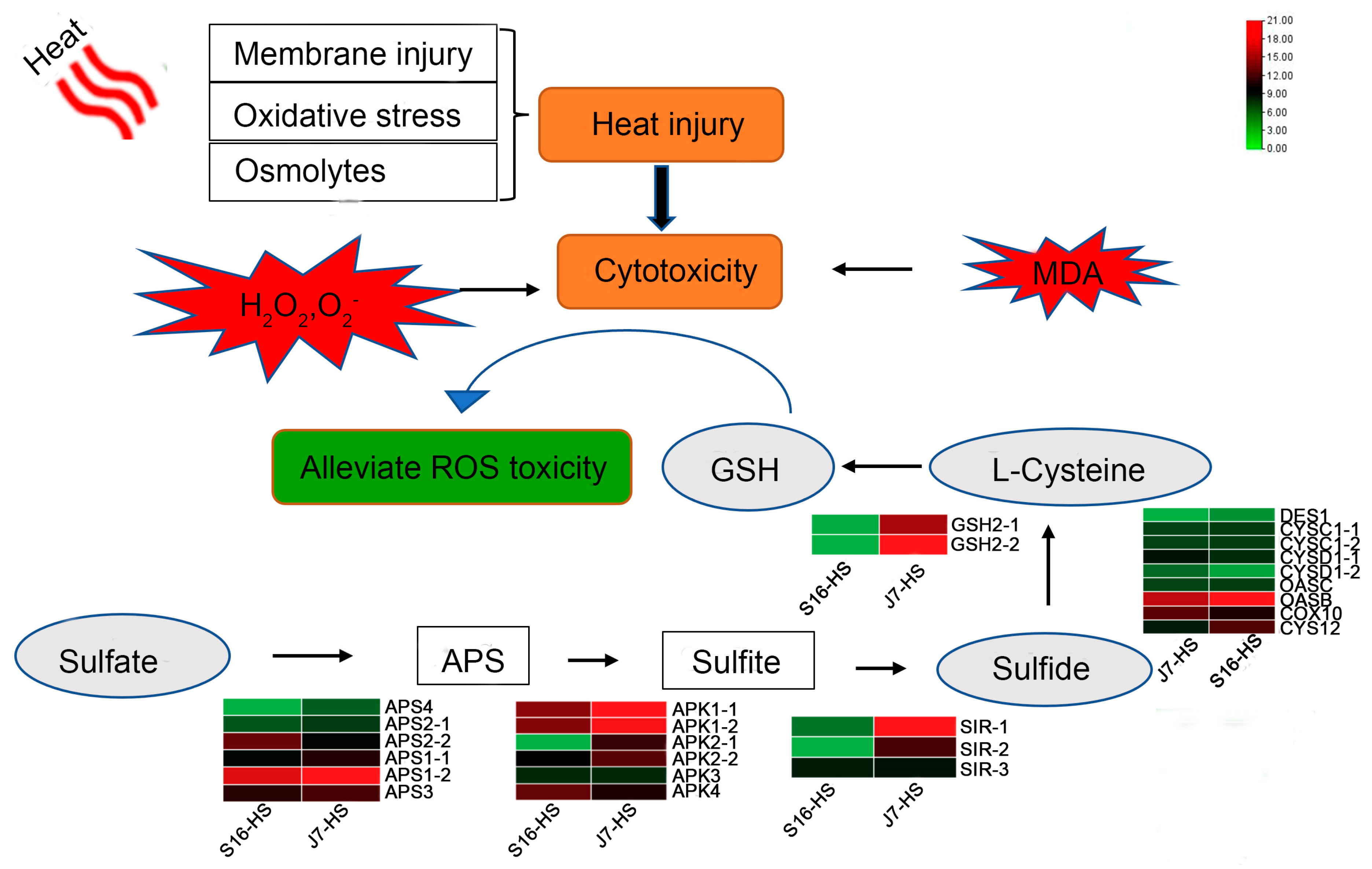
2.6. DEGs of Sulfur Metabolism Pathway
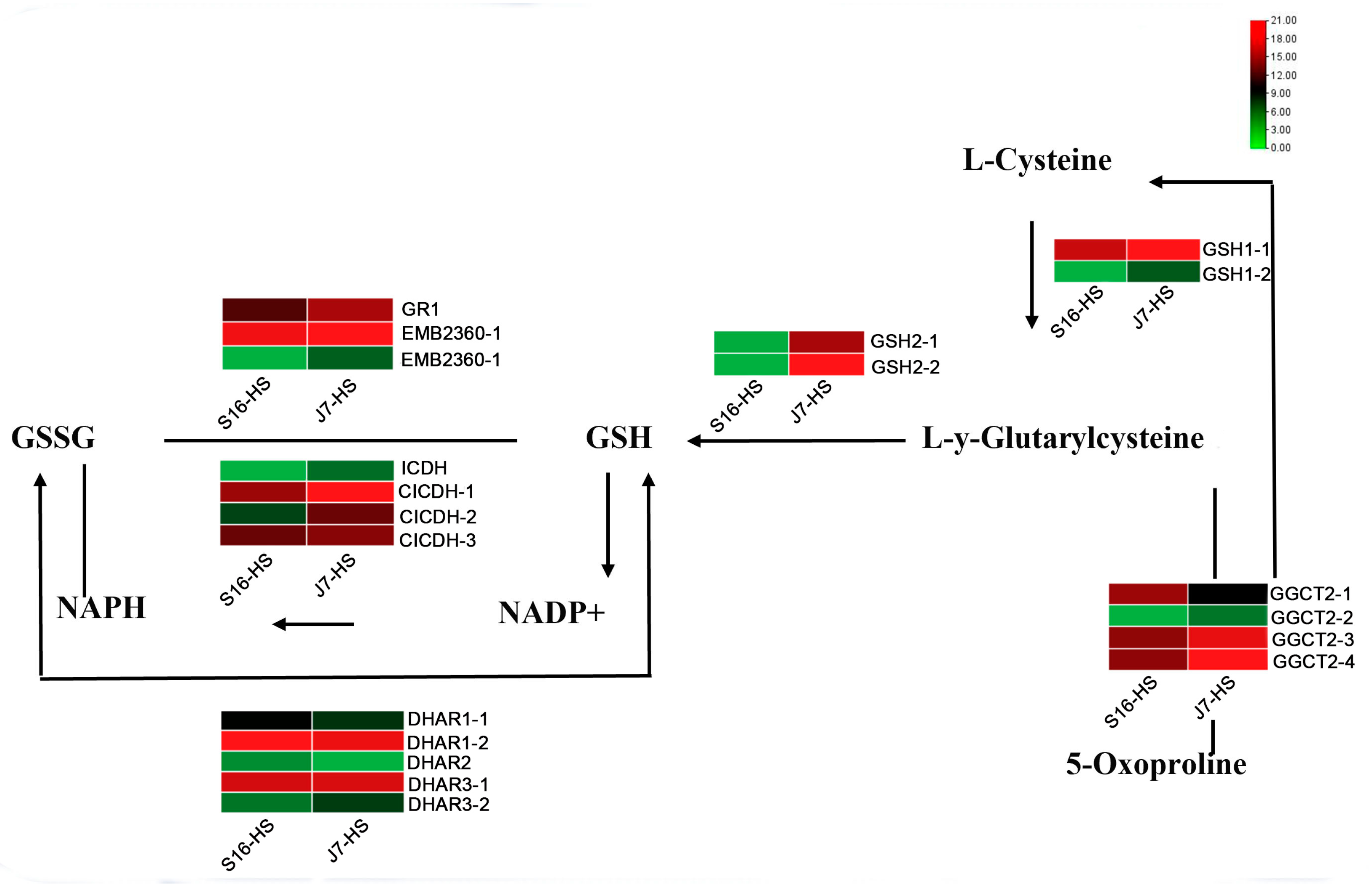
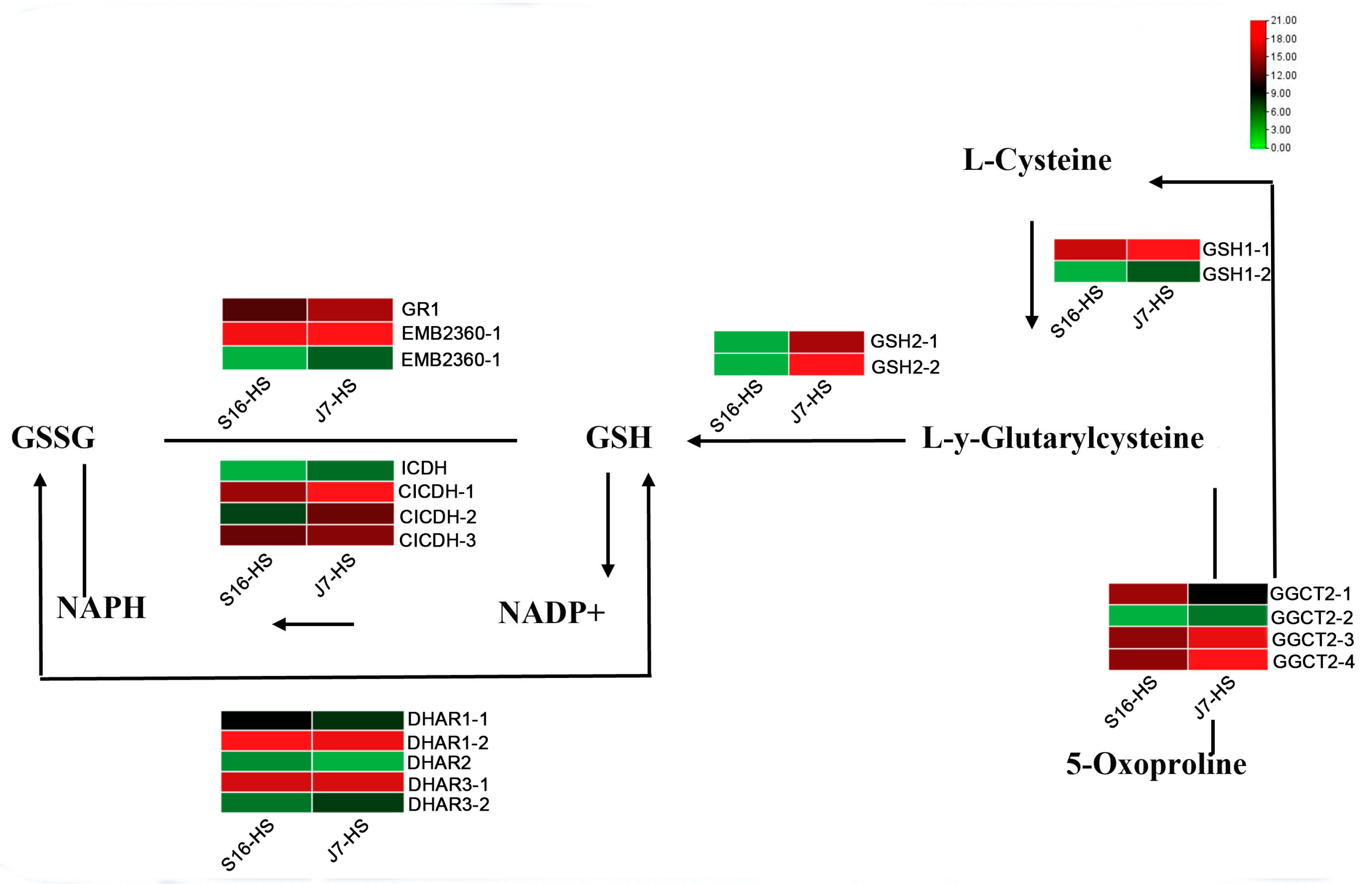
2.7. DEGs of Glutathione Metabolism Pathway
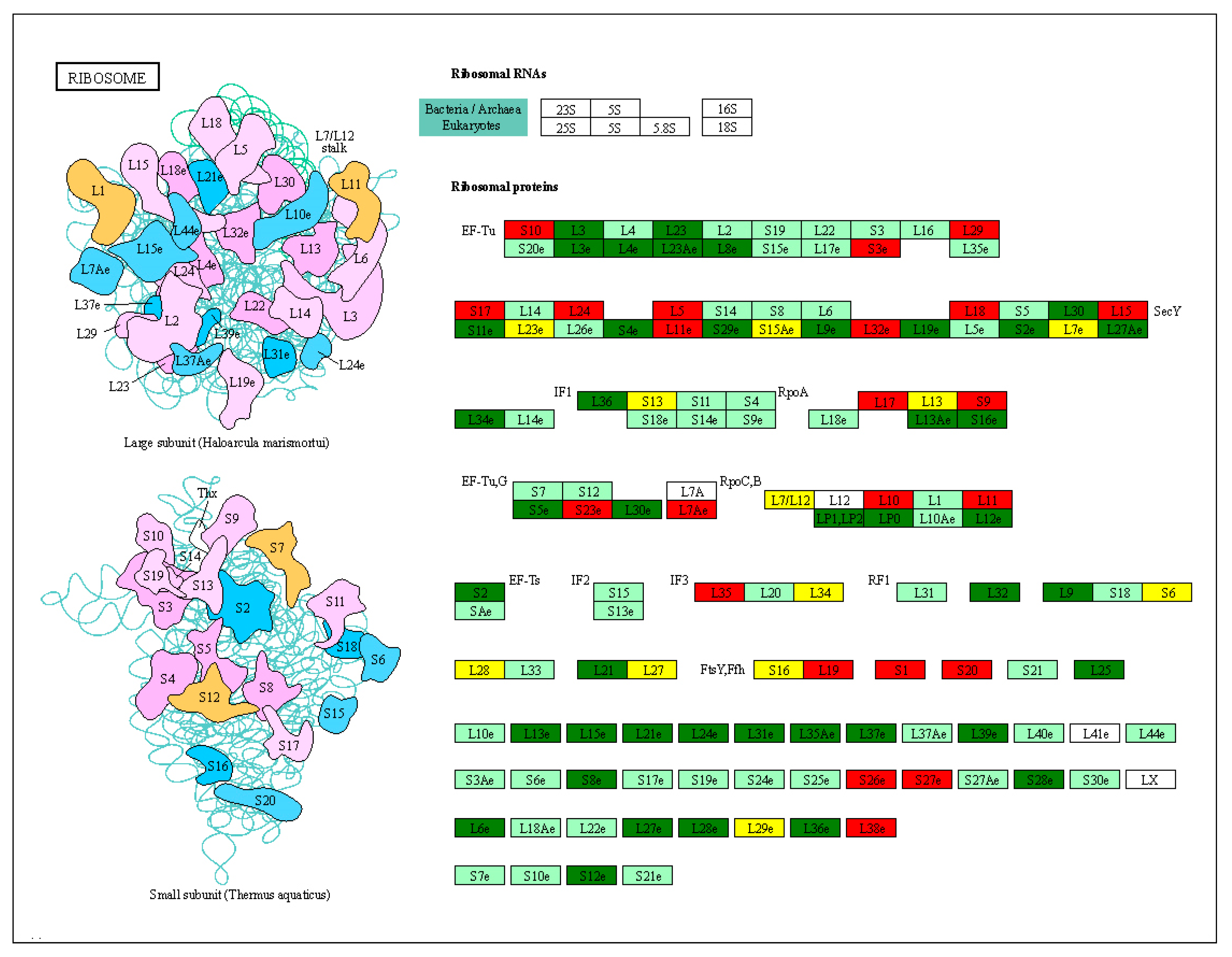
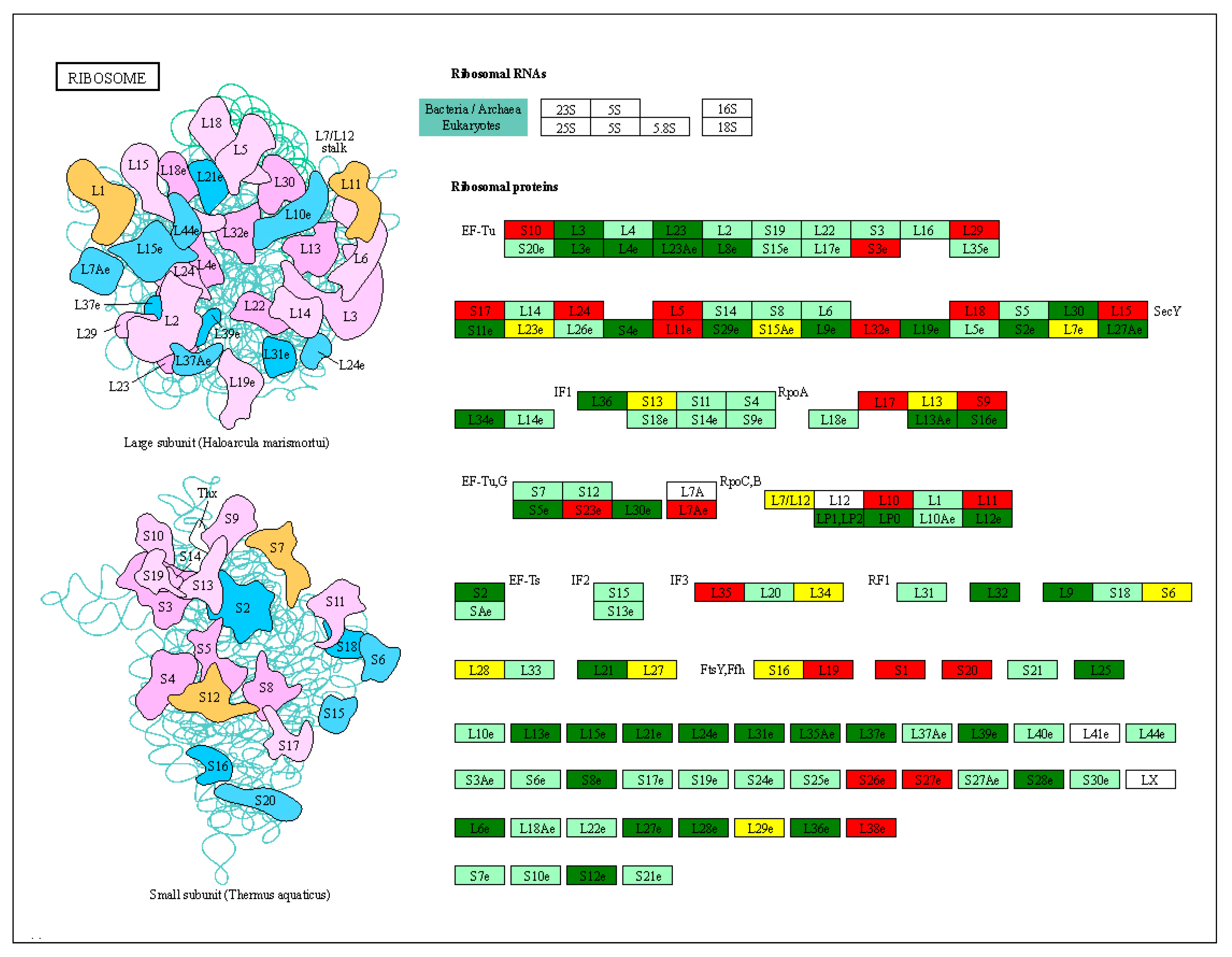
2.8. DEGs of Ribosome Pathway
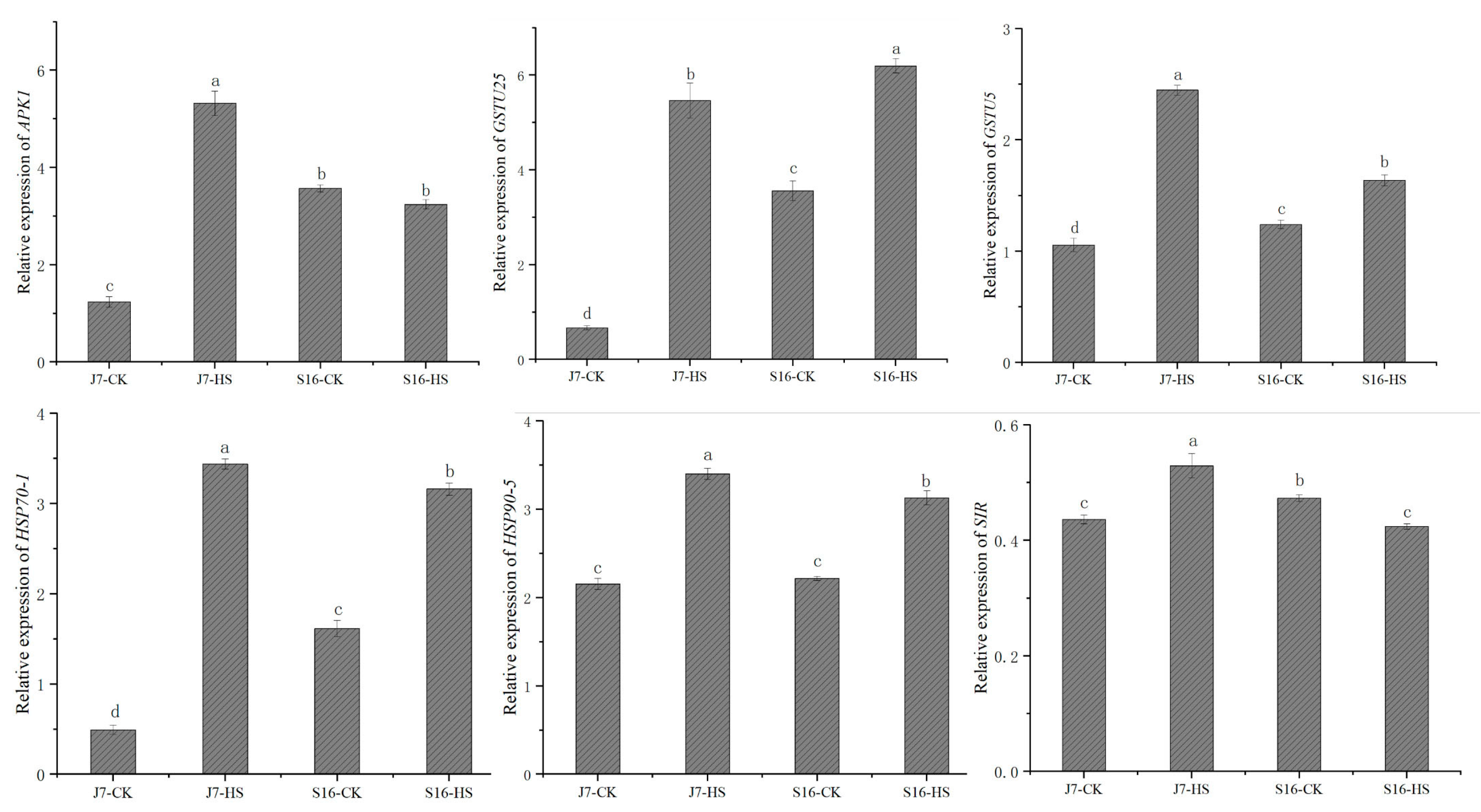
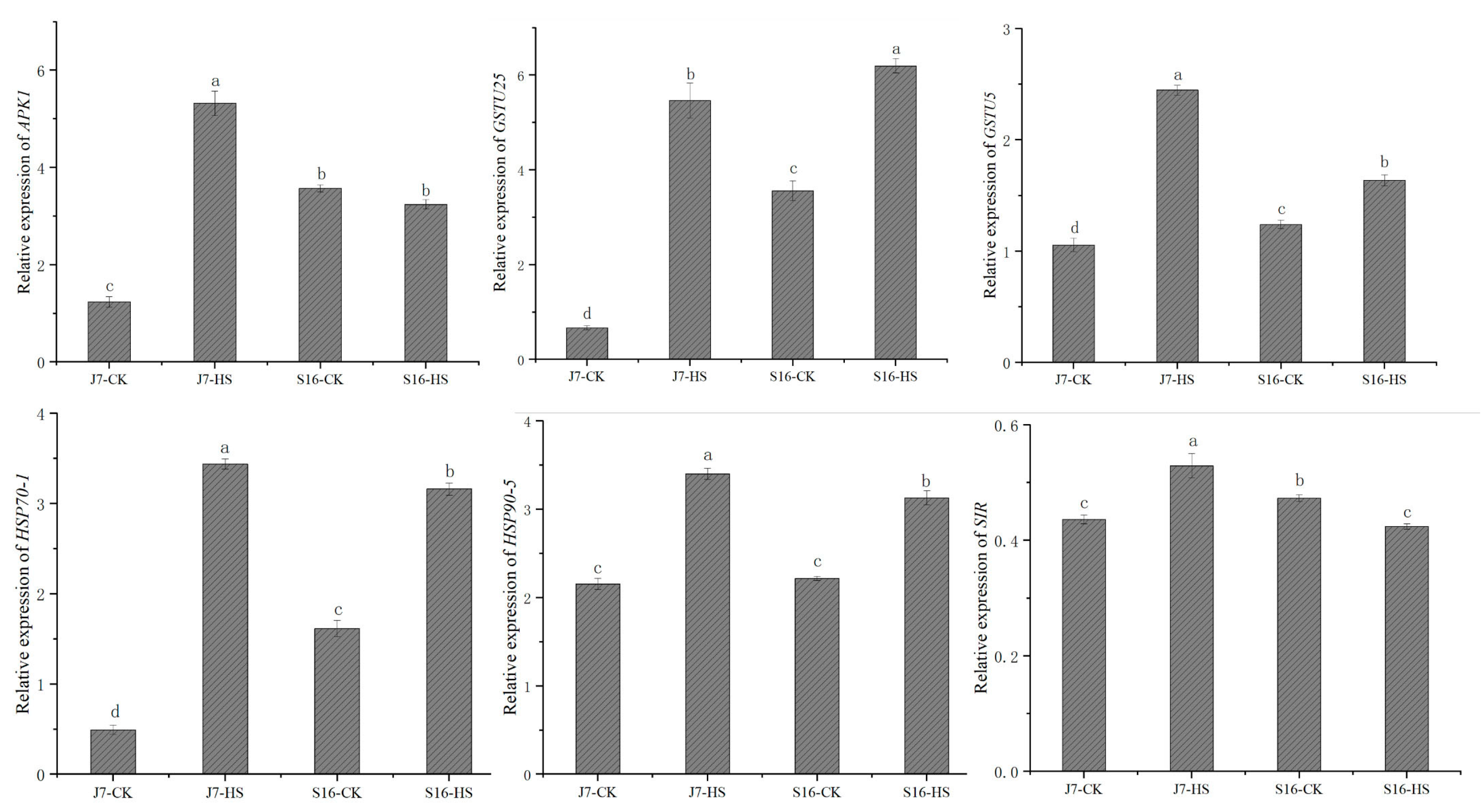
2.9. Experimental Validation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. Growth Parameters
4.3. Heat Injury Index (HII)
4.4. Chl Fluorescence
4.5. Relative Conductivity (REC)
4.6. Malondialdehyde (MDA)
4.7. Soluble Protein, Soluble Sugar, and Vitamin C Content
4.8. Cellulose
4.9. H2O2, O2−, and T-AOC
4.10. Transcriptomic Data Analysis
4.11. Validation of RNA-Seq by qRT-PCR
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bita, C.E.; Gerats, T. Plant Tolerance to High Temperature in a Changing Environment: Scientific Fundamentals and Production of Heat Stress-Tolerant Crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Ort, D.R. More than Taking the Heat: Crops and Global Change. Curr. Opin. Plant Biol. 2010, 13, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Caers, M.; Rudelsheim, P.; Van Onckelen, H.; Horemans, S. Effect of Heat Stress on Photosynthetic Activity and Chloroplast Ultrastructure in Correlation with Endogenous Cytokinin Concentration in Maize Seedlings. Plant Cell Physiol. 1985, 26, 47–52. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The Effect of Drought and Heat Stress on Reproductive Processes in Cereals. Plant Cell Environ. 2007, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Qi, T.; Wasternack, C.; Xie, D. Jasmonate Signaling and Crosstalk with Gibberellin and Ethylene. Curr. Opin. Plant Biol. 2014, 21, 112–119. [Google Scholar] [CrossRef]
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing Climate Change Adaptation Needs for Food Security in 2030. Science 2008, 319, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Scheerer, U.; Haensch, R.; Mendel, R.R.; Kopriva, S.; Rennenberg, H.; Herschbach, C. Sulphur Flux through the Sulphate Assimilation Pathway Is Differently Controlled by Adenosine 5′-Phosphosulphate Reductase under Stress and in Transgenic Poplar Plants Overexpressing γ-ECS, SO, or APR. J. Exp. Bot. 2010, 61, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Choi, S.R.; Wang, Y.; Chhapekar, S.S.; Zhang, X.; Wang, Y.; Zhang, X.; Zhu, M.; Liu, D.; Zuo, Z.; et al. Starch Content Changes and Metabolism-Related Gene Regulation of Chinese Cabbage Synergistically Induced by Plasmodiophora brassicae Infection. Hortic. Res. 2022, 9, uhab071. [Google Scholar] [CrossRef]
- Dong, W.; Stockwell, V.O.; Goyer, A. Enhancement of Thiamin Content in Arabidopsis Thaliana by Metabolic Engineering. Plant Cell Physiol. 2015, 56, 2285–2296. [Google Scholar] [CrossRef]
- Larkindale, J.; Vierling, E. Core Genome Responses Involved in Acclimation to High Temperature. Plant Physiol. 2008, 146, 323–324. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Chen, Q.; Yi, M. Effects of Short-Term Heat Stress on Oxidative Damage and Responses of Antioxidant System in Lilium Longiflorum. Plant Growth Regul. 2007, 54, 45–54. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.; Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Jiang, Y.; Zhao, H.; Hou, M. Acquired Thermotolerance in Plants. Plant Cell Tissue Organ Cult. PCTOC 2012, 111, 265–276. [Google Scholar] [CrossRef]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive Oxygen Species as Signals That Modulate Plant Stress Responses and Programmed Cell Death. BioEssays 2006, 28, 1091–1101. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as Key Players in Plant Stress Signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front. Plant Sci. 2015, 6, 165102. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M. Heat Tolerance in Plants: An Overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Iba, K. A CCLIMATIVE R ESPONSE TO T EMPERATURE S TRESS IN H IGHER P LANTS: Approaches of Gene Engineering for Temperature Tolerance. Annu. Rev. Plant Biol. 2002, 53, 225–245. [Google Scholar] [CrossRef]
- ElBasyoni, I.; Saadalla, M.; Baenziger, S.; Bockelman, H.; Morsy, S. Cell Membrane Stability and Association Mapping for Drought and Heat Tolerance in a Worldwide Wheat Collection. Sustainability 2017, 9, 1606. [Google Scholar] [CrossRef]
- Xu, S.; Li, J.; Zhang, X.; Wei, H.; Cui, L. Effects of Heat Acclimation Pretreatment on Changes of Membrane Lipid Peroxidation, Antioxidant Metabolites, and Ultrastructure of Chloroplasts in Two Cool-Season Turfgrass Species under Heat Stress. Environ. Exp. Bot. 2006, 56, 274–285. [Google Scholar] [CrossRef]
- Li, Z.; Howell, S.H. Heat Stress Responses and Thermotolerance in Maize. Int. J. Mol. Sci. 2021, 22, 948. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Jha, A.B.; Dubey, R.S. Arsenite Treatment Induces Oxidative Stress, Upregulates Antioxidant System, and Causes Phytochelatin Synthesis in Rice Seedlings. Protoplasma 2011, 248, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Furutera, A.; Seki, K.; Toyoda, Y.; Tanaka, K.; Sugimoto, Y. Malondialdehyde Generated from Peroxidized Linolenic Acid Causes Protein Modification in Heat-Stressed Plants. Plant Physiol. Biochem. 2008, 46, 786–793. [Google Scholar] [CrossRef]
- Cloonan, N.; Forrest, A.R.R.; Kolle, G.; Gardiner, B.B.A.; Faulkner, G.J.; Brown, M.K.; Taylor, D.F.; Steptoe, A.L.; Wani, S.; Bethel, G.; et al. Stem Cell Transcriptome Profiling via Massive-Scale MRNA Sequencing. Nat. Methods 2008, 5, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Kumazaki, A.; Suzuki, N. Enhanced Tolerance to a Combination of Heat Stress and Drought in Arabidopsis Plants Deficient in ICS1 Is Associated with Modulation of Photosynthetic Reaction Center Proteins. Physiol. Plant. 2019, 165, 232–246. [Google Scholar] [CrossRef]
- Oyiga, B.C.; Sharma, R.C.; Shen, J.; Baum, M.; Ogbonnaya, F.C.; Léon, J.; Ballvora, A. Identification and Characterization of Salt Tolerance of Wheat Germplasm Using a Multivariable Screening Approach. J. Agron. Crop Sci. 2016, 202, 472–485. [Google Scholar] [CrossRef]
- Nyarko, G.; Alderson, P.G.; Craigon, J.; Murchie, E.; Sparkes, D.L. Comparison of Cell Membrane Thermostability and Chlorophyll Fluorescence Parameters for the Determination of Heat Tolerance in Ten Cabbage Lines. J. Hortic. Sci. Biotechnol. 2008, 83, 678–682. [Google Scholar] [CrossRef]
- Mishra, D.; Shekhar, S.; Agrawal, L.; Chakraborty, S.; Chakraborty, N. Cultivar-Specific High Temperature Stress Responses in Bread Wheat (Triticum aestivum L.) Associated with Physicochemical Traits and Defense Pathways. Food Chem. 2017, 221, 1077–1087. [Google Scholar] [CrossRef]
- Goraya, G.K.; Kaur, B.; Asthir, B.; Bala, S.; Kaur, G.; Farooq, M. Rapid Injuries of High Temperature in Plants. J. Plant Biol. 2017, 60, 298–305. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A. Physiological Implications of Metabolite Biosynthesis for Net Assimilation and Heat-Stress Tolerance of Sugarcane (Saccharum officinarum) Sprouts. J. Plant Res. 2007, 120, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Close, T.J. Expression of Dehydrins under Heat Stress and Their Relationship with Water Relations of Sugarcane Leaves. Biol. Plant. 2007, 51, 104–109. [Google Scholar] [CrossRef]
- Franklin, K.A. Light and Temperature Signal Crosstalk in Plant Development. Curr. Opin. Plant Biol. 2009, 12, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, J.; Manzano, S.; Palma, F.; Carvajal, F.; Garrido, D.; Jamilena, M. Oxidative Stress Associated with Chilling Injury in Immature Fruit: Postharvest Technological and Biotechnological Solutions. Int. J. Mol. Sci. 2017, 18, 1467. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, B.C.; Oelmüller, R. Reactive Oxygen Species Generation and Signaling in Plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Maulana, F.; Ayalew, H.; Anderson, J.D.; Kumssa, T.T.; Huang, W.; Ma, X.-F. Genome-Wide Association Mapping of Seedling Heat Tolerance in Winter Wheat. Front. Plant Sci. 2018, 9, 1272. [Google Scholar] [CrossRef]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen Peroxide Sensing and Signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef]
- Mhamdi, A.; Hager, J.; Chaouch, S.; Queval, G.; Han, Y.; Taconnat, L.; Saindrenan, P.; Gouia, H.; Issakidis-Bourguet, E.; Renou, J.-P.; et al. Arabidopsis GLUTATHIONE REDUCTASE1 Plays a Crucial Role in Leaf Responses to Intracellular Hydrogen Peroxide and in Ensuring Appropriate Gene Expression through Both Salicylic Acid and Jasmonic Acid Signaling Pathways. Plant Physiol. 2010, 153, 1144–1160. [Google Scholar] [CrossRef]
- Min, K.; Son, H.; Lee, J.; Choi, G.J.; Kim, J.-C.; Lee, Y.-W. Peroxisome Function Is Required for Virulence and Survival of Fusarium graminearum. Mol. Plant-Microbe Interact. 2012, 25, 1617–1627. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, Y.; Hu, Y.; Jia, X.; Zhao, T.; Cheng, L.; Wang, Y. Tolerance of Two Apple Rootstocks to Short-Term Salt Stress: Focus on Chlorophyll Degradation, Photosynthesis, Hormone and Leaf Ultrastructures. Acta Physiol. Plant. 2019, 41, 87. [Google Scholar] [CrossRef]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic Stress: Interplay between ROS, Hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Gracey, A.Y.; Fraser, E.J.; Li, W.; Fang, Y.; Taylor, R.R.; Rogers, J.; Brass, A.; Cossins, A.R. Coping with Cold: An Integrative, Multitissue Analysis of the Transcriptome of a Poikilothermic Vertebrate. Proc. Natl. Acad. Sci. USA 2004, 101, 16970–16975. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.R.; Johnston, I.A. Temperature during Embryonic Development Has Persistent Effects on Thermal Acclimation Capacity in Zebrafish. Proc. Natl. Acad. Sci. USA 2012, 109, 14247–14252. [Google Scholar] [CrossRef]
- Somero, G.N. Proteins and Temperature. Annu. Rev. Physiol. 1995, 57, 43–68. [Google Scholar] [CrossRef]
- Simon, S.A.; Advani, S.; McIntosh, T.J. Temperature Dependence of the Repulsive Pressure between Phosphatidylcholine Bilayers. Biophys. J. 1995, 69, 1473–1483. [Google Scholar] [CrossRef]
- Pan, J.; Tristram-Nagle, S.; Kučerka, N.; Nagle, J.F. Temperature Dependence of Structure, Bending Rigidity, and Bilayer Interactions of Dioleoylphosphatidylcholine Bilayers. Biophys. J. 2008, 94, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.J.; Ito, T.; Okada, T.S.; Ohnishi, S.I. A Correlation between Membrane Fluidity and the Critical Temperature for Cell Adhesion. J. Cell Biol. 1976, 71, 670–674. [Google Scholar] [CrossRef]
- Swindell, W.R.; Huebner, M.; Weber, A.P. Plastic and Adaptive Gene Expression Patterns Associated with Temperature Stress in Arabidopsis Thaliana. Heredity 2007, 99, 143–150. [Google Scholar] [CrossRef]
- Zhang, G.; Guo, G.; Hu, X.; Zhang, Y.; Li, Q.; Li, R.; Zhuang, R.; Lu, Z.; He, Z.; Fang, X.; et al. Deep RNA Sequencing at Single Base-Pair Resolution Reveals High Complexity of the Rice Transcriptome. Genome Res. 2010, 20, 646–654. [Google Scholar] [CrossRef]
- Yi, X.; Hou, L. Genetic Expression of Heat Tolerance in Eggplant. J. Hortic. 2002, 529–532. Available online: https://d.wanfangdata.com.cn/periodical/yyxb200206006 (accessed on 30 March 2024).
- Stewart, R.R.C.; Bewley, J.D. Lipid Peroxidation Associated with Accelerated Aging of Soybean Axes. Plant Physiol. 1980, 65, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Yemm, E.W.; Willis, A.J. The Estimation of Carbohydrates in Plant Extracts by Anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.; Cha, M.; Shin, S.; Kim, K. Enzymatic Production of a Soluble-Fibre Hydrolyzate from Carrot Pomace and Its Sugar Composition. Food Chem. 2005, 92, 151–157. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript Assembly and Quantification by RNA-Seq Reveals Unannotated Transcripts and Isoform Switching during Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq Expression Estimates by Correcting for Fragment Bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Dry Shoot wt (g) | Fresh Shoot wt (g) | Leaf Area (cm2) | Leaf Color (ΔE) | Plant ht (cm) |
|---|---|---|---|---|---|
| S16-CK | 0.22 ± 0.13 b | 2.34 ± 0.15 b | 6.81 ± 0.15 b | 5.803 ± 0.17 b | 7.27 ± 0.39 b |
| S16-HS | 0.14 ± 0.02 c | 1.53 ± 0.05 c | 4.87 ± 1.01 c | 8.06 ± 0.45 a | 5.53 ± 0.45 c |
| J7-CK | 0.29 ± 0.05 a | 3.06 ± 0.08 a | 10.09 ± 0.76 a | 3.54 ± 0.11 c | 9.43 ± 0.53 a |
| J7-HS | 0.22 ± 0.15 b | 2.36 ± 0.20 b | 8.93 ± 0.46 a | 5.27 ± 0.02 b | 8.17 ± 0.21 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, C.; Peng, X.; Yang, X.; Wang, C.; Yuan, L.; Chen, G.; Tang, X.; Wang, W.; Wu, J.; Zhu, S.; et al. Physiological and Transcriptomic Responses of Bok Choy to Heat Stress. Plants 2024, 13, 1093. https://doi.org/10.3390/plants13081093
Dong C, Peng X, Yang X, Wang C, Yuan L, Chen G, Tang X, Wang W, Wu J, Zhu S, et al. Physiological and Transcriptomic Responses of Bok Choy to Heat Stress. Plants. 2024; 13(8):1093. https://doi.org/10.3390/plants13081093
Chicago/Turabian StyleDong, Cuina, Xixuan Peng, Xiaona Yang, Chenggang Wang, Lingyun Yuan, Guohu Chen, Xiaoyan Tang, Wenjie Wang, Jianqiang Wu, Shidong Zhu, and et al. 2024. "Physiological and Transcriptomic Responses of Bok Choy to Heat Stress" Plants 13, no. 8: 1093. https://doi.org/10.3390/plants13081093
APA StyleDong, C., Peng, X., Yang, X., Wang, C., Yuan, L., Chen, G., Tang, X., Wang, W., Wu, J., Zhu, S., Huang, X., Zhang, J., & Hou, J. (2024). Physiological and Transcriptomic Responses of Bok Choy to Heat Stress. Plants, 13(8), 1093. https://doi.org/10.3390/plants13081093