
Quantifying the Individual and Combined Effects of Short-Term Heat Stress at Booting and Flowering Stages on Nonstructural Carbohydrates Remobilization in Rice
 , , ,
, , ,  , ,
, ,
Abstract
1. Introduction
2. Results
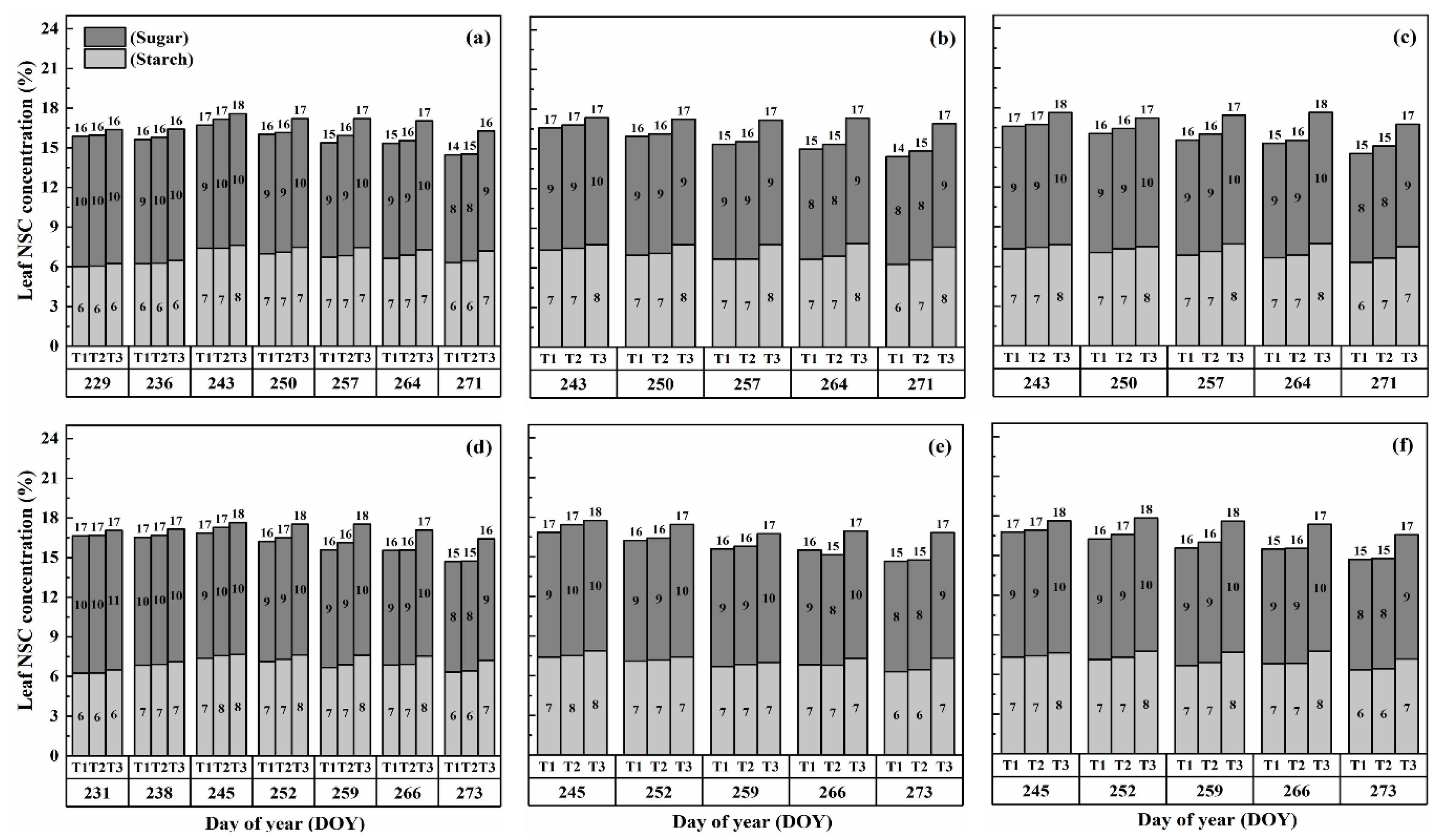
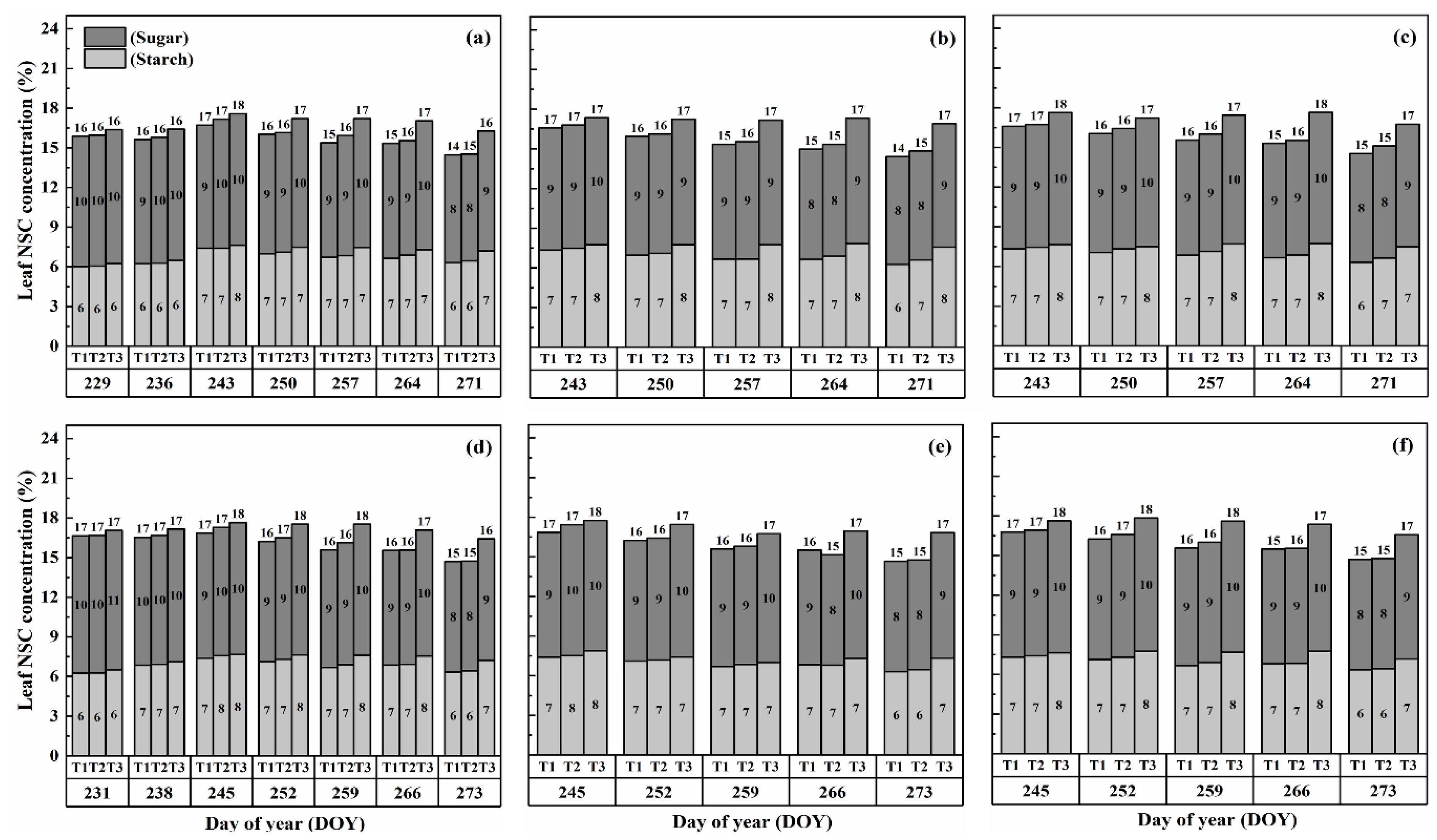
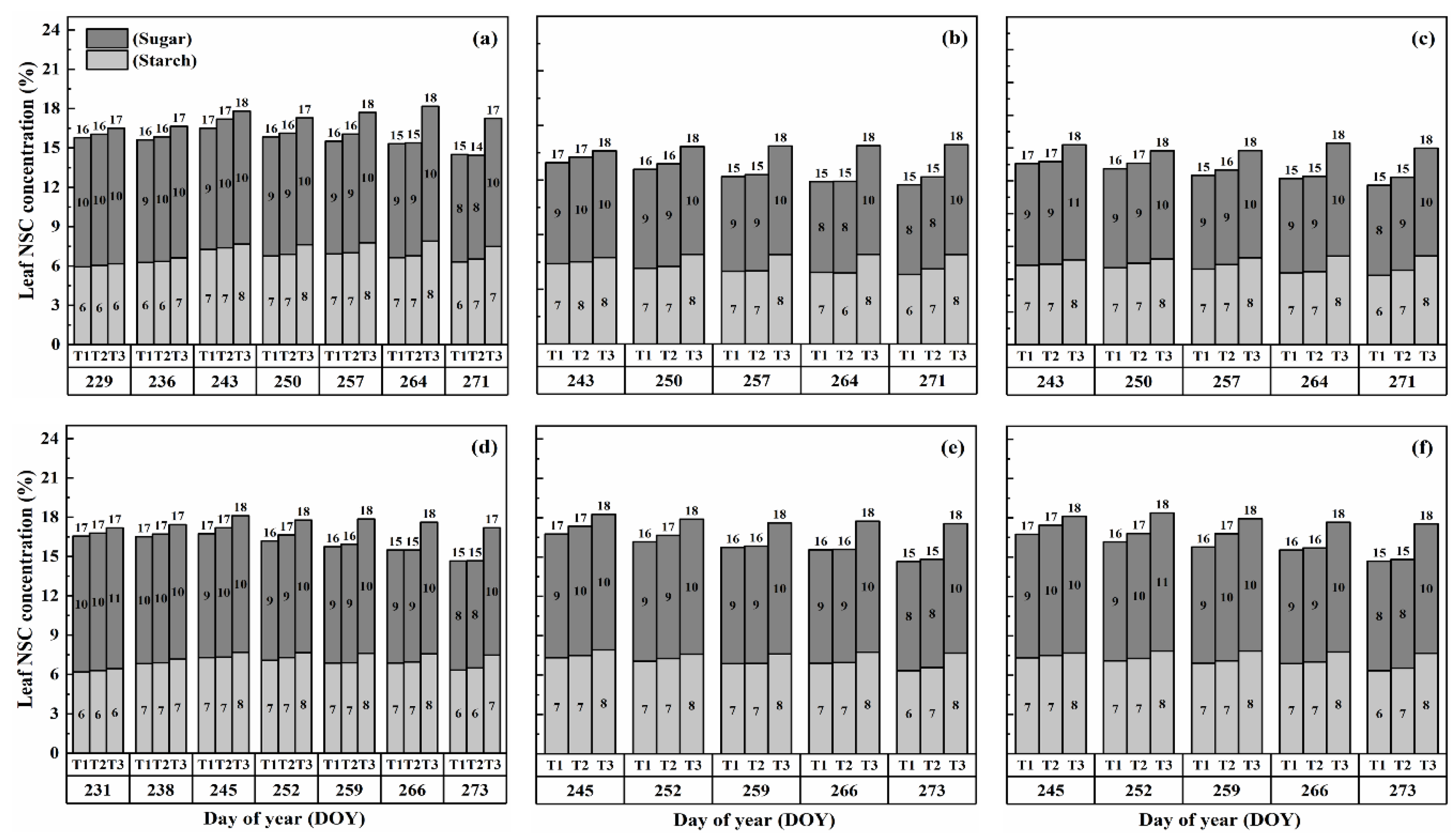
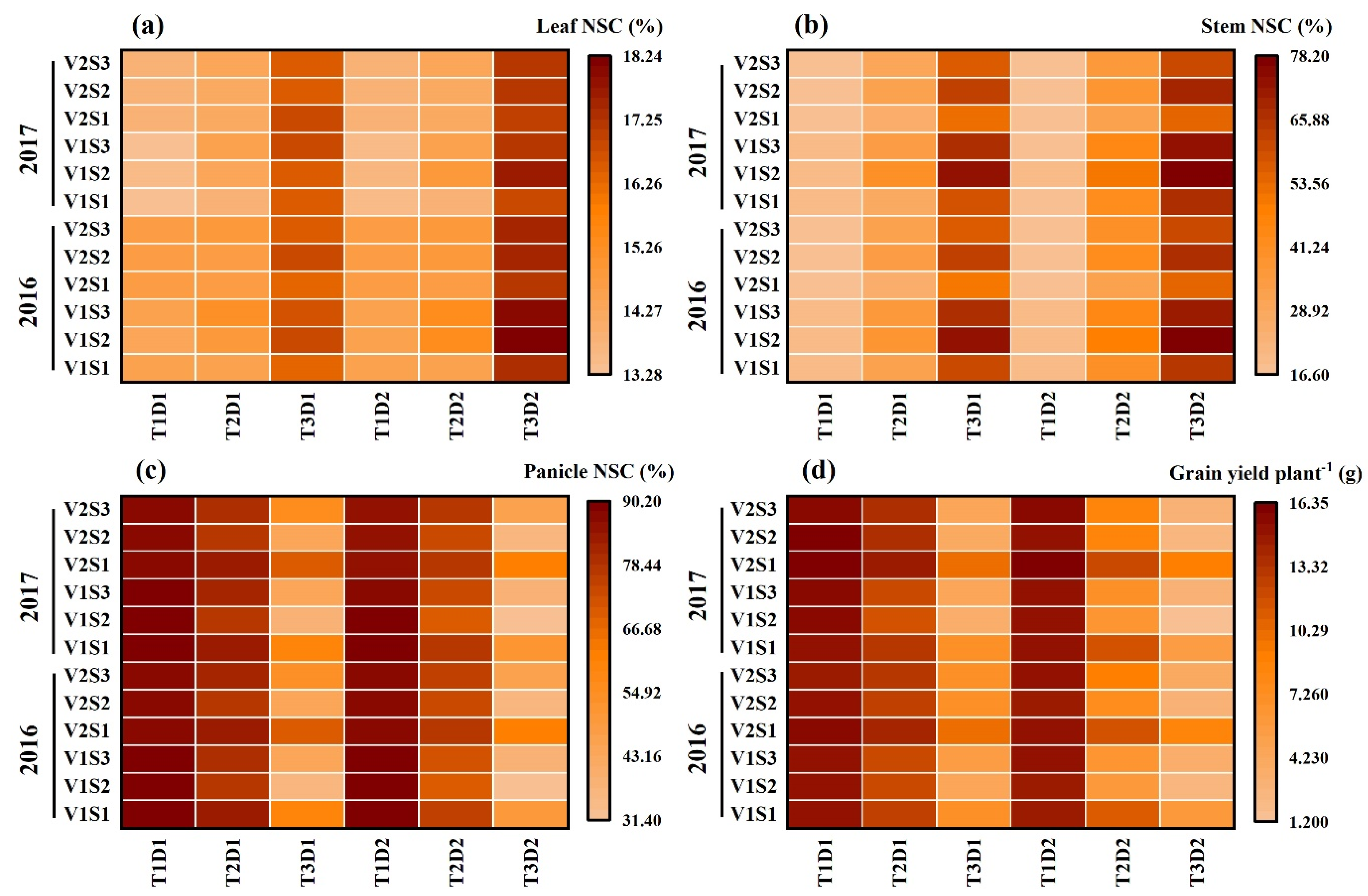
2.1. Dynamics of NSCs Concentration in Leaves
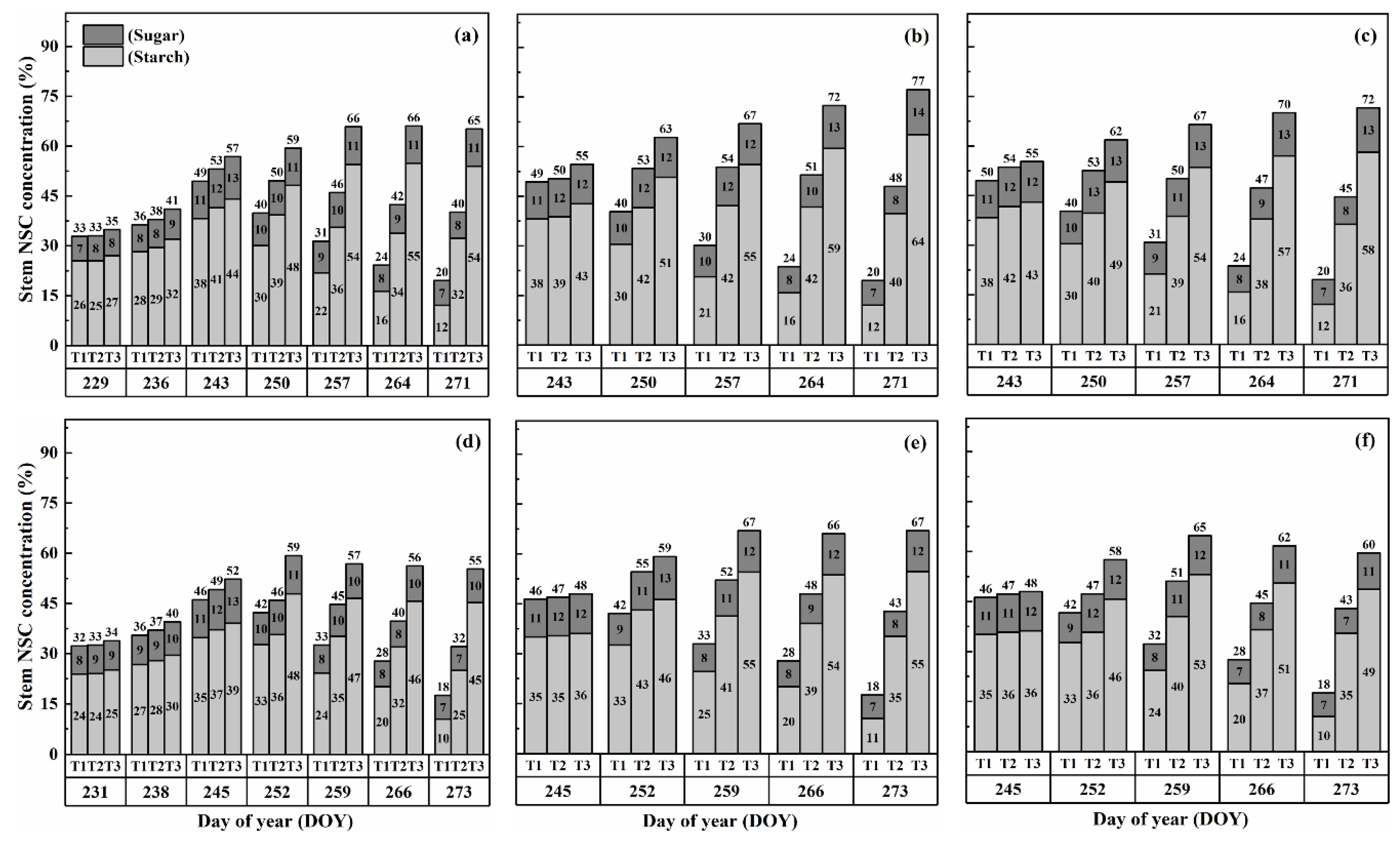
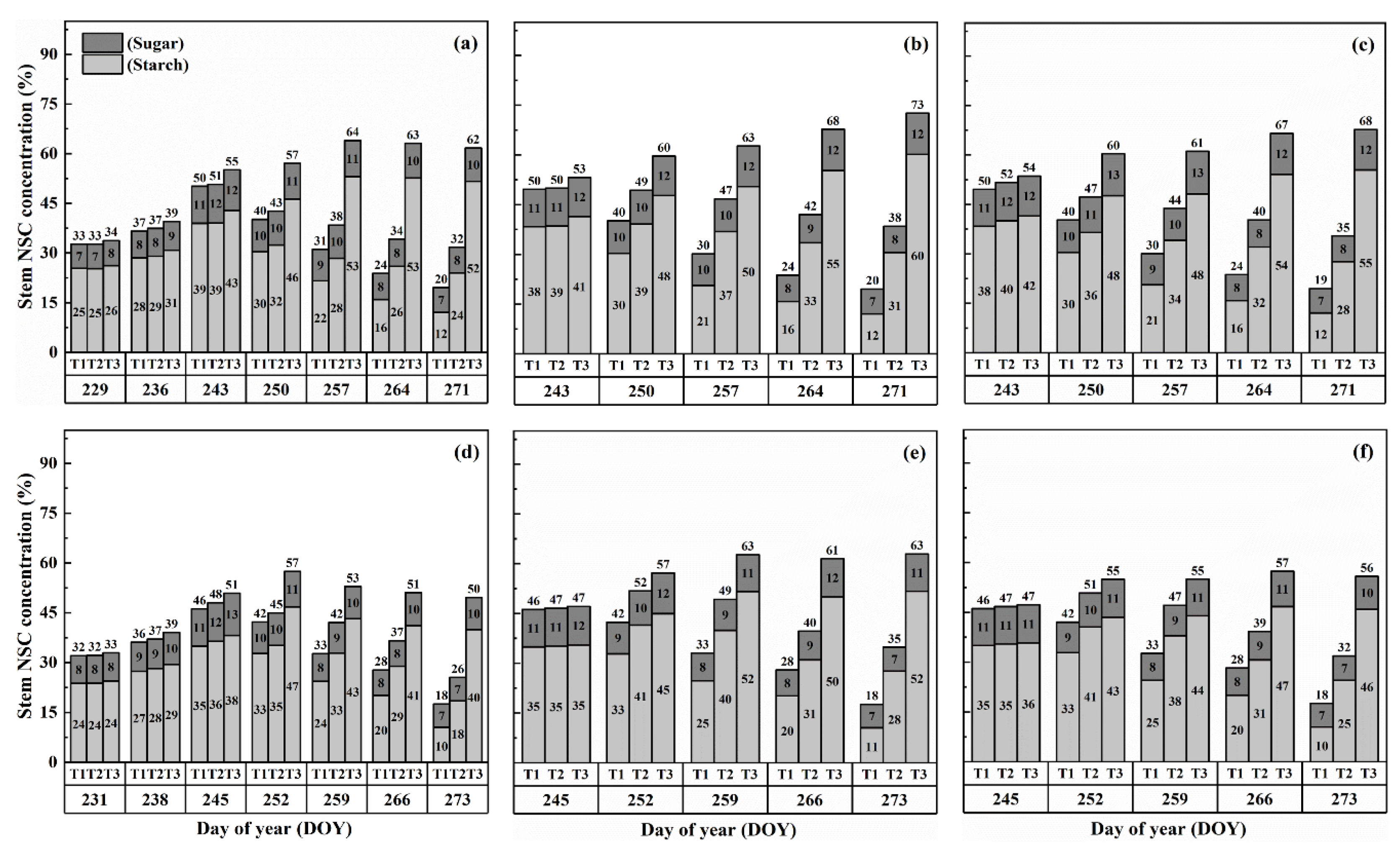
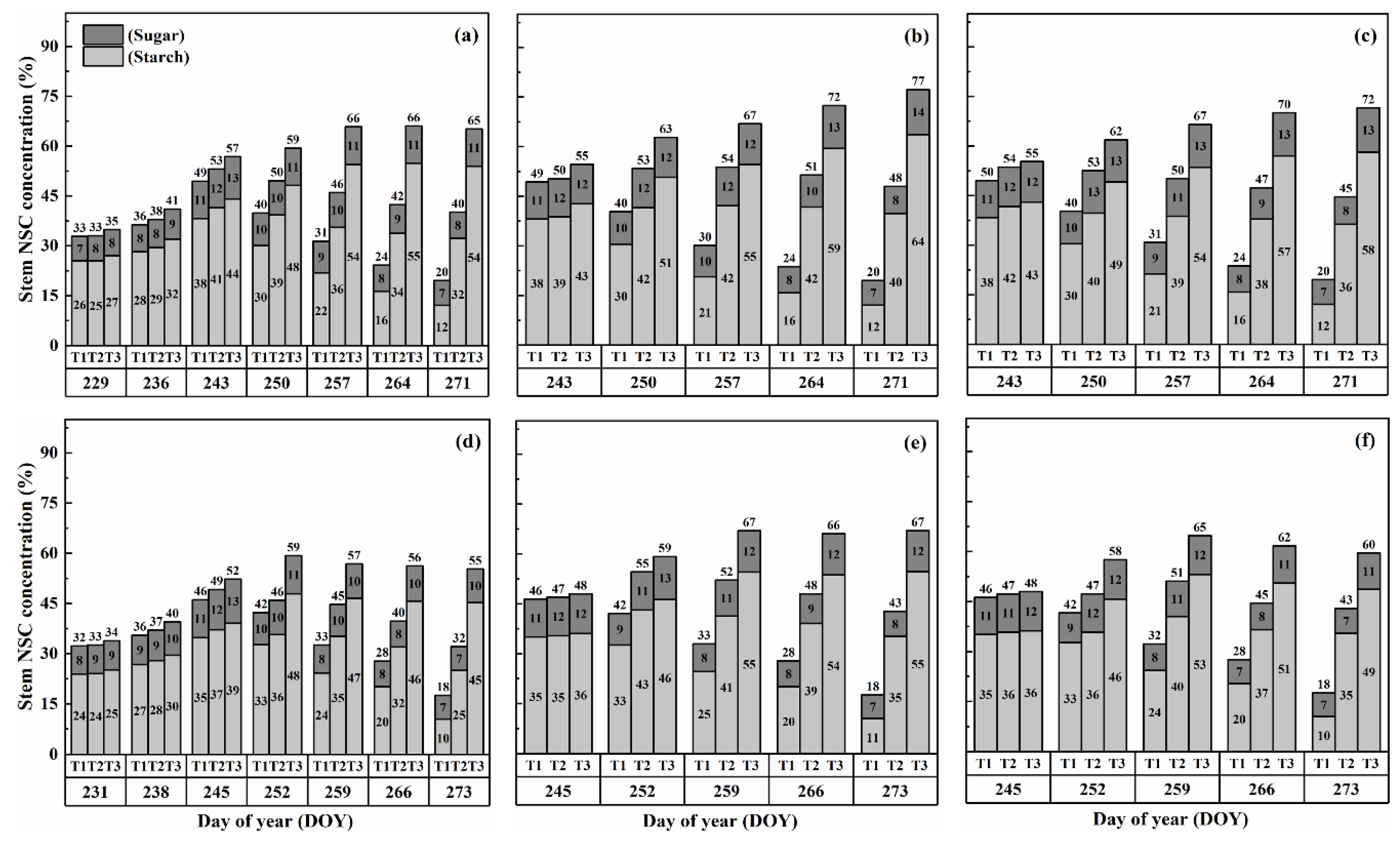
2.2. Dynamics of NSC Concentration in Stems
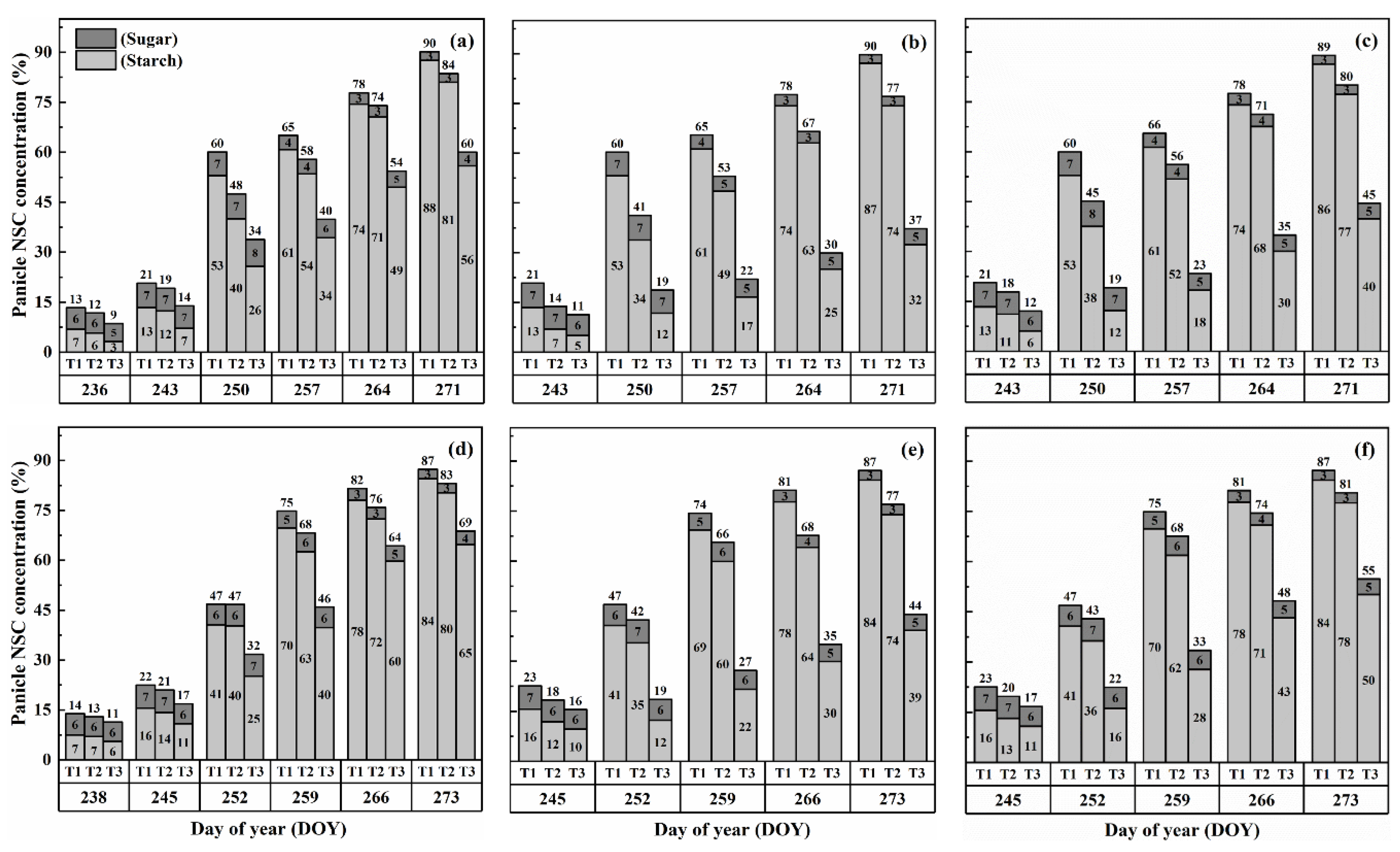
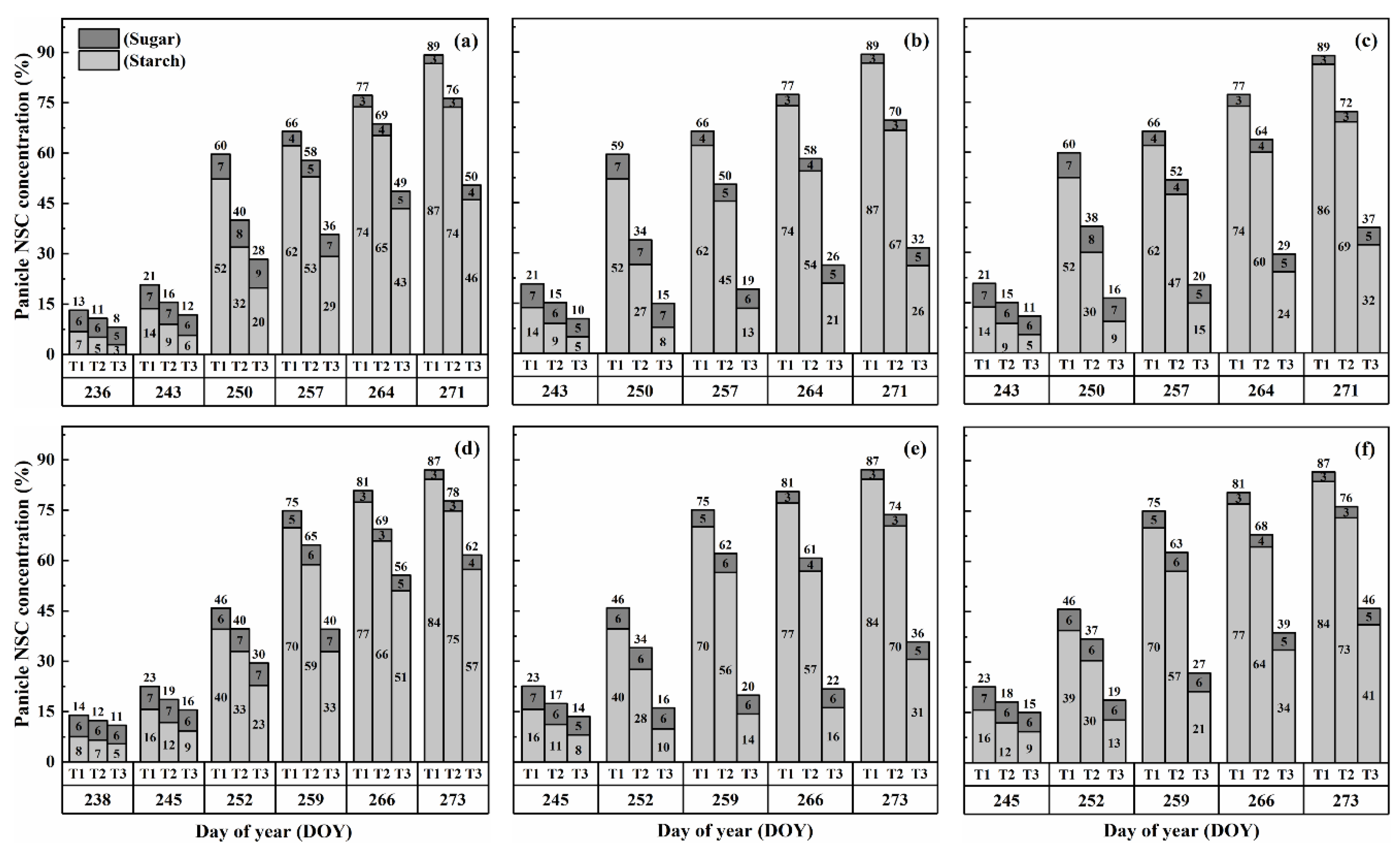
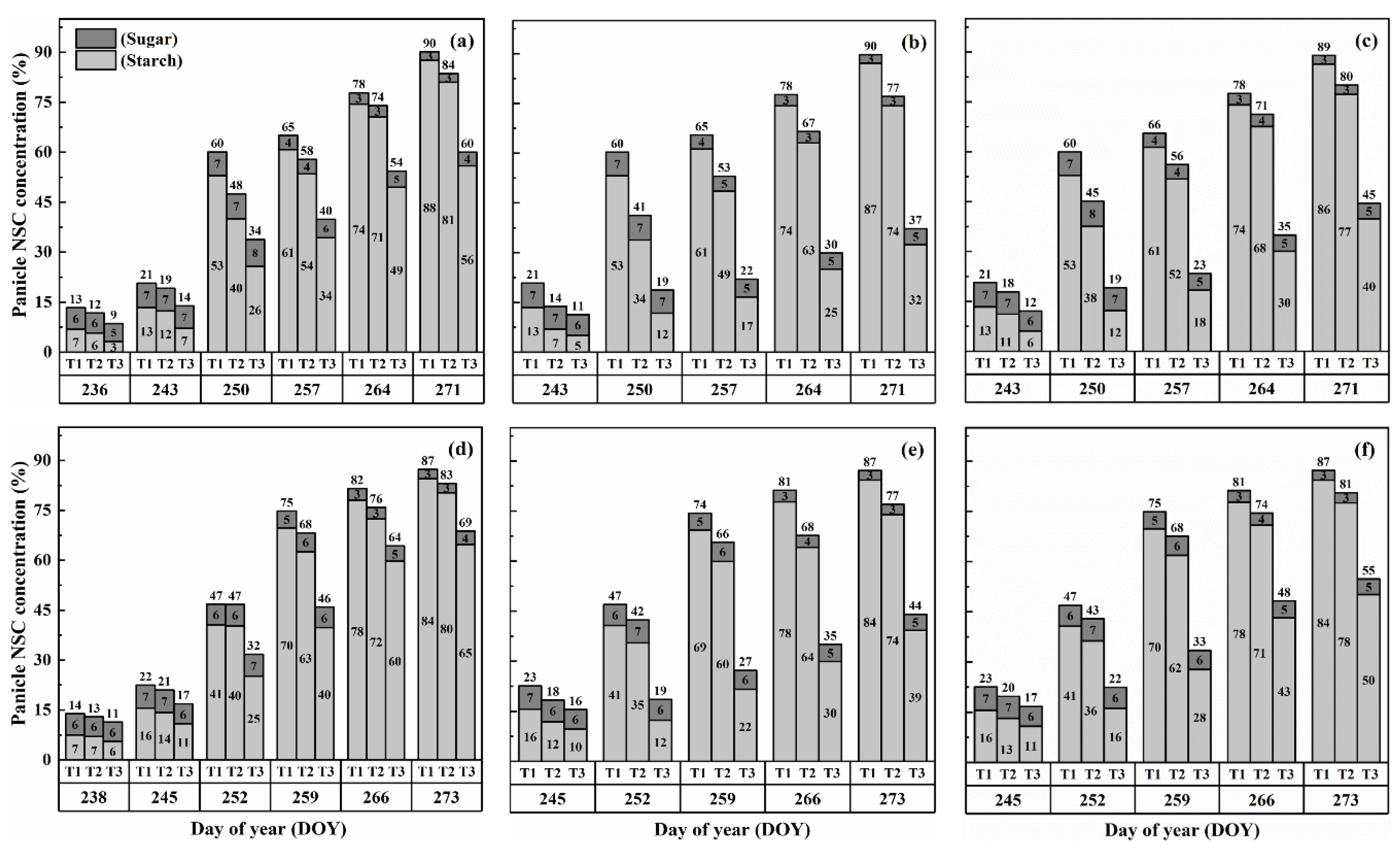
2.3. Dynamics of NSC Concentration in Panicles
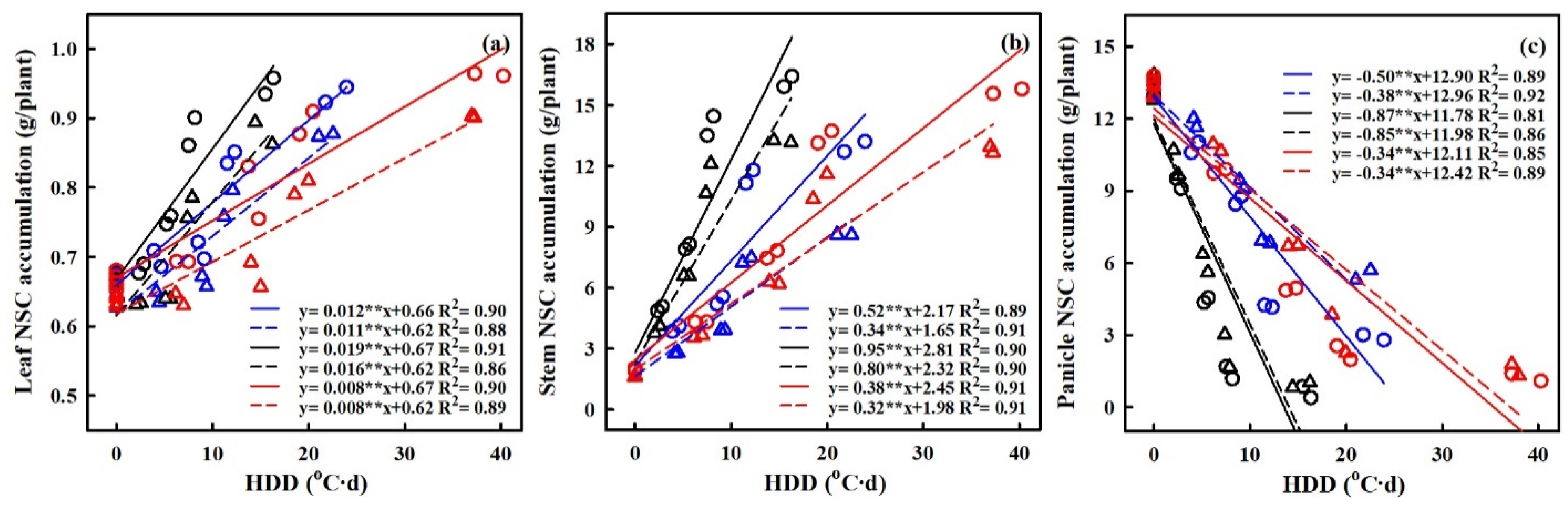
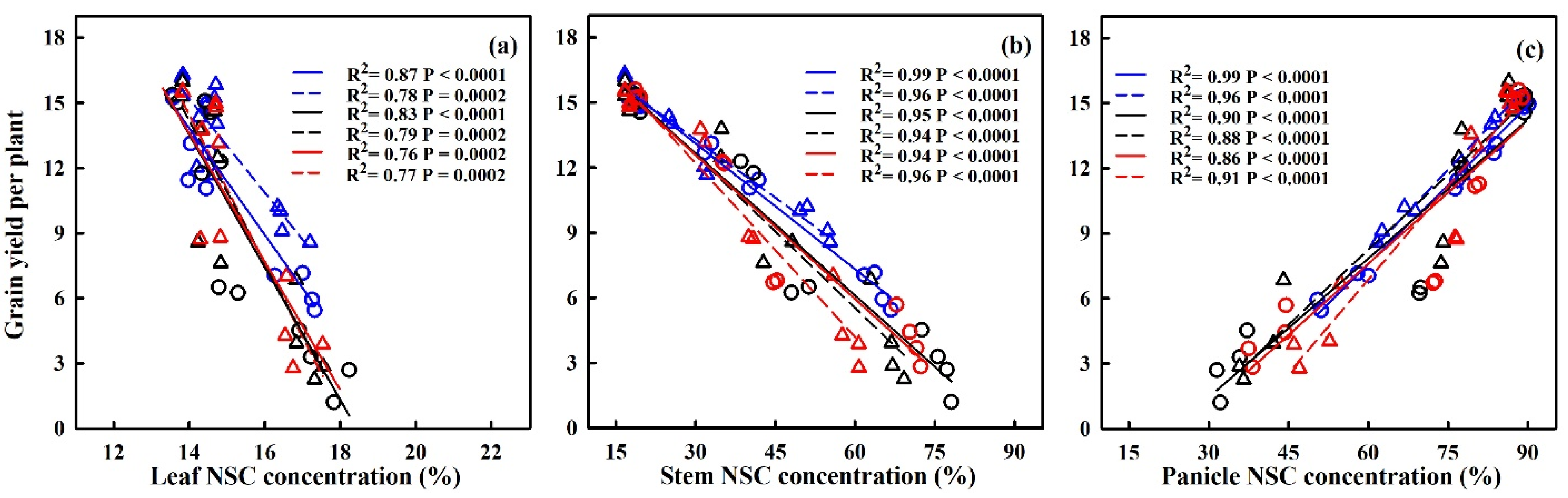
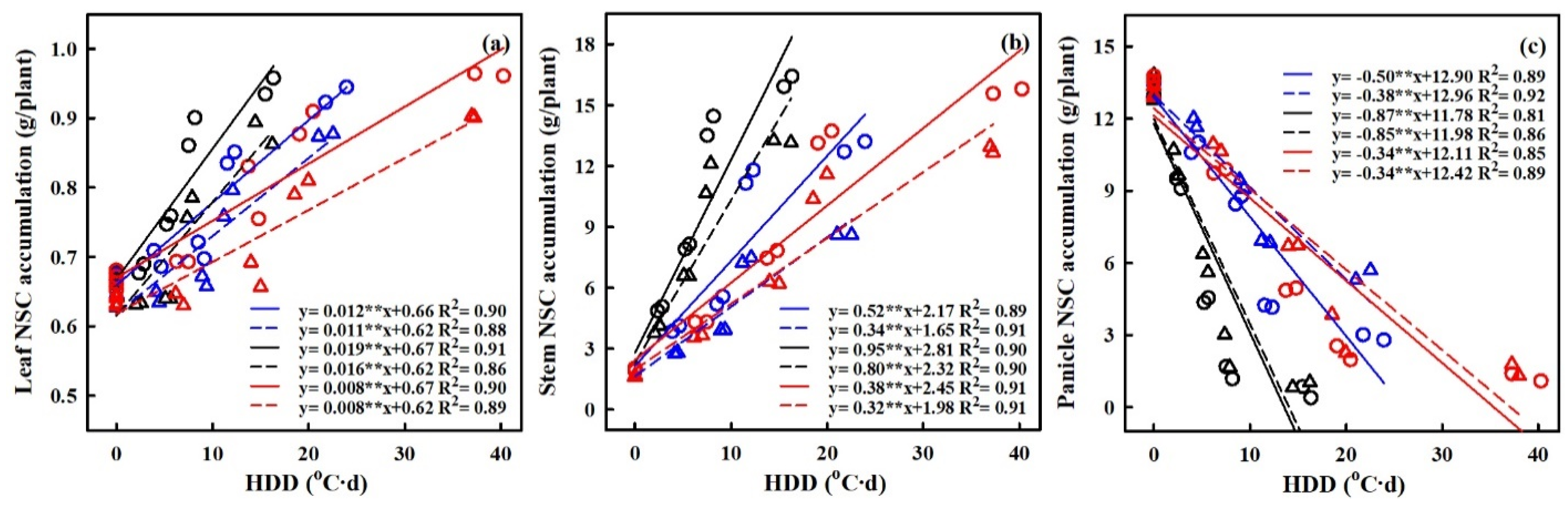
2.4. Effect of Heat Stress on NSC Accumulation at Maturity
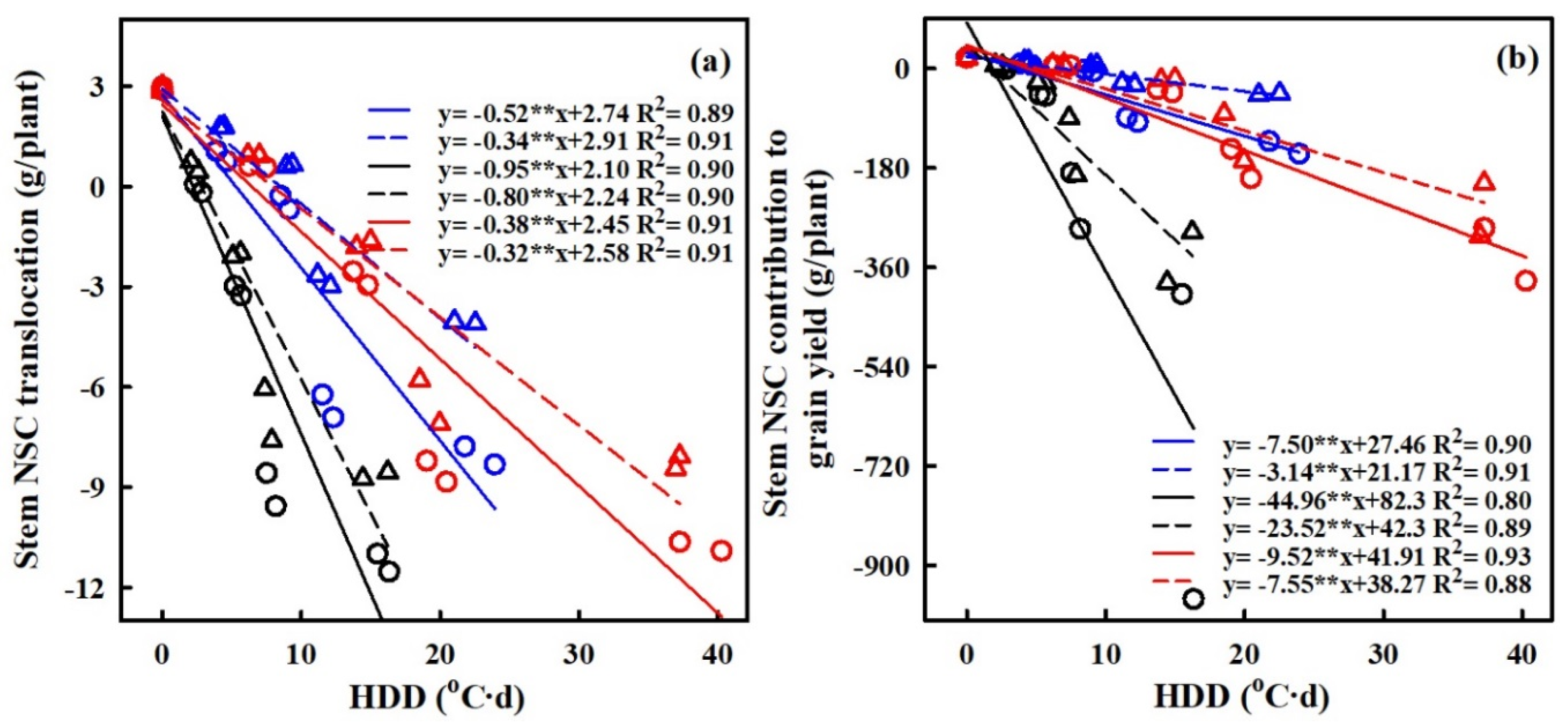
2.5. Effect of Heat Stress on NSC Translocation at Maturity
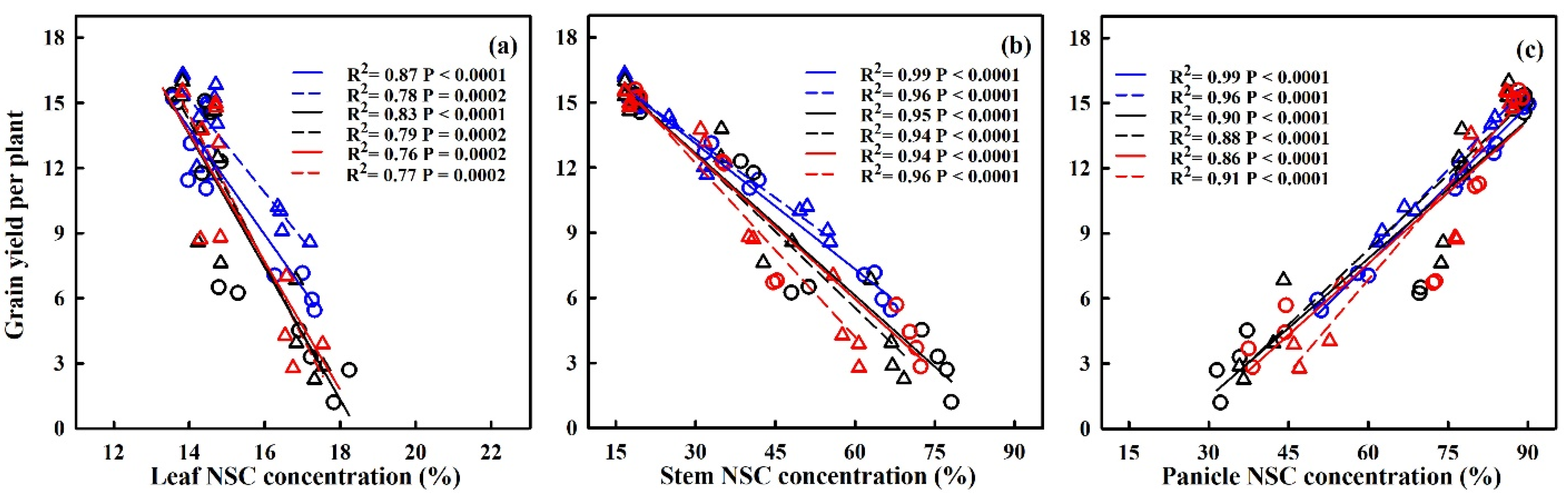
2.6. Quantification of the Effects of Heat Stress on NSC Accumulation
3. Discussion
3.1. Effect of Heat Stress on NonStructural Carbohydrate Dynamics and Remobilization in Plant Parts
3.2. Quantifying the Impact of Heat Stress
4. Materials and Methods
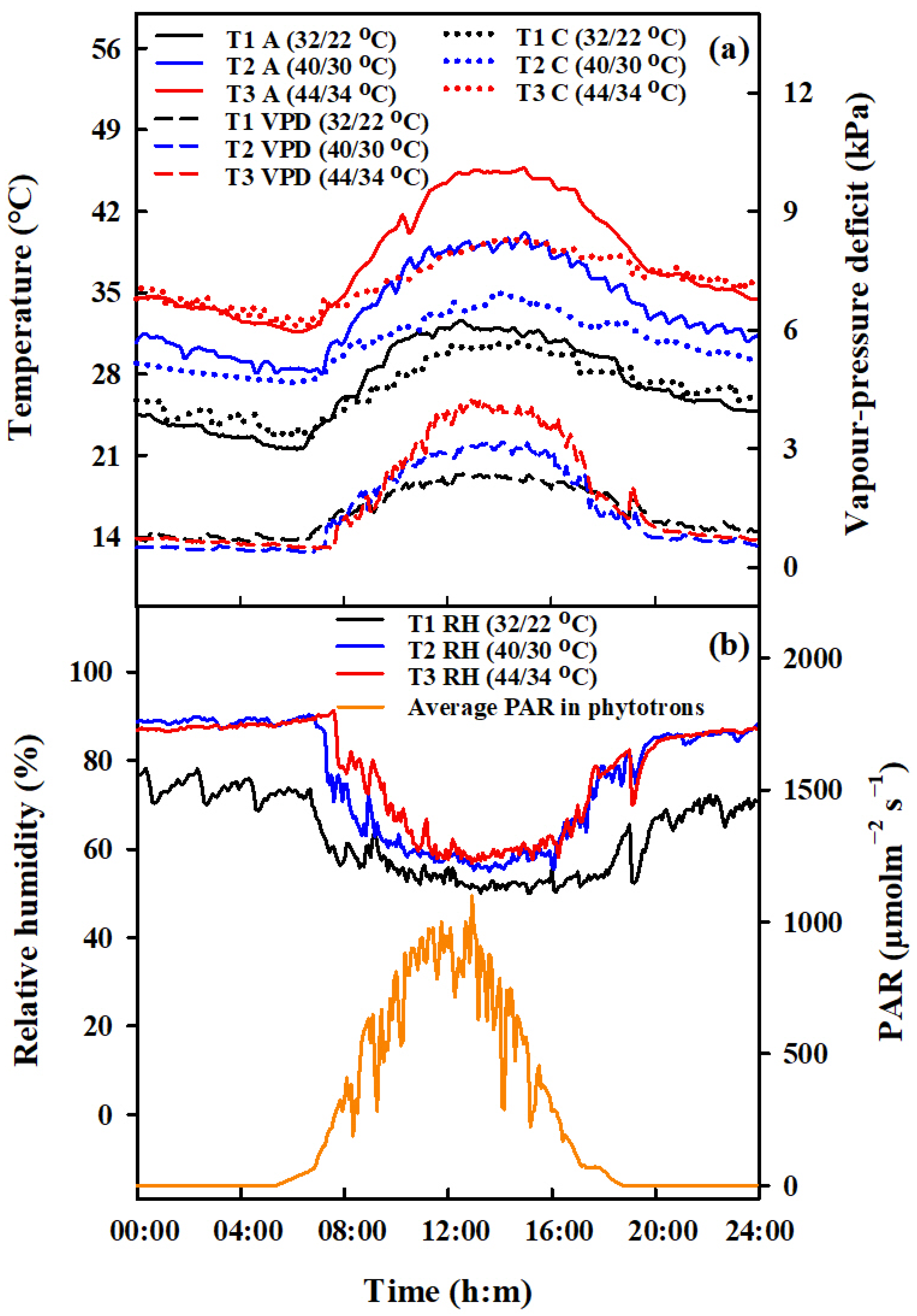
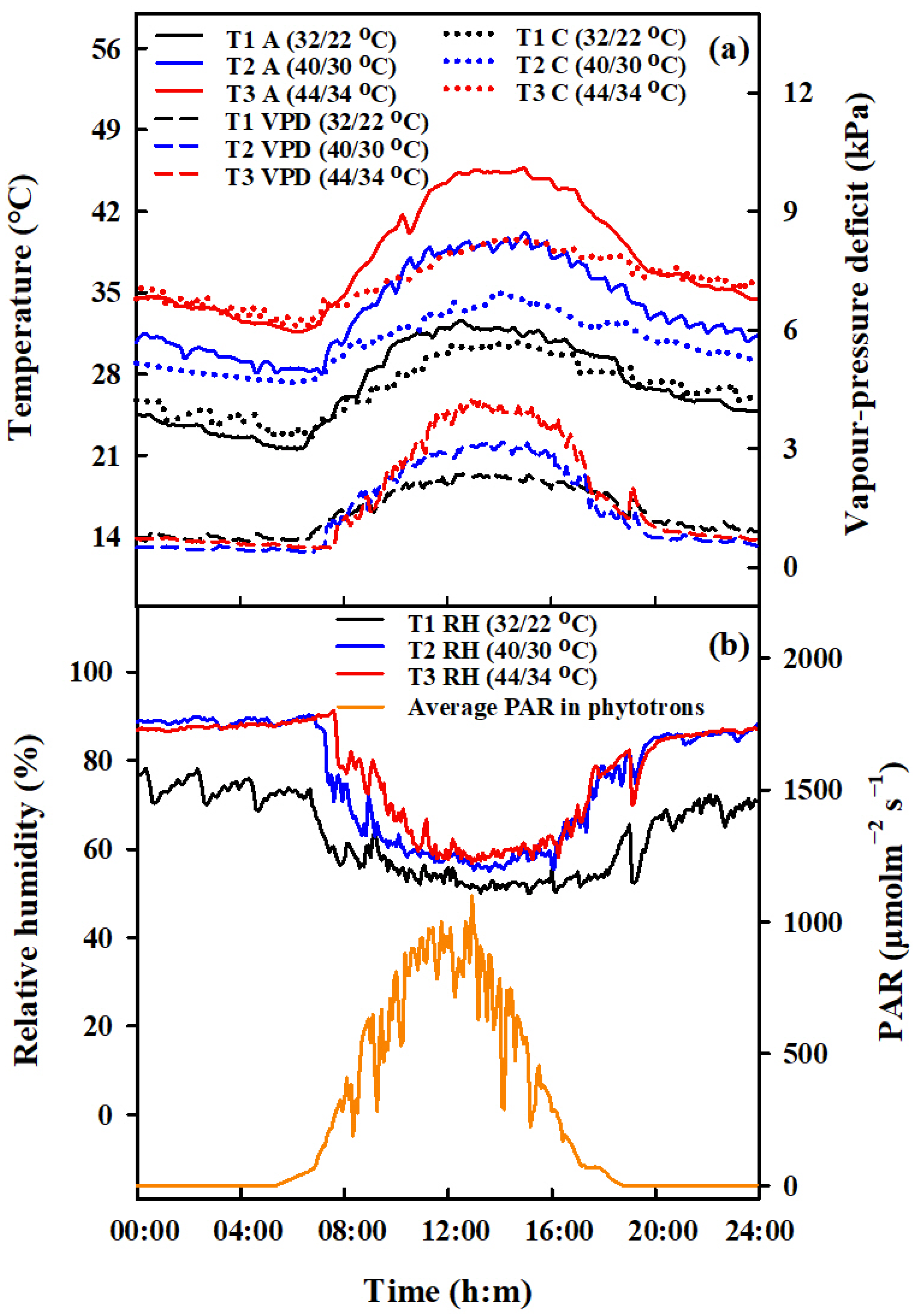
4.1. Experimental Details
4.2. Measurement of Grain Yield Parameters
4.3. Calculation of NSC Accumulation and Translocation Parameters
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- He, L.; Cleverly, J.; Wang, B.; Jin, N.; Mi, C.; Liu, D.L.; Yu, Q. Multi-model ensemble projections of future extreme heat stress on rice across southern China. Theor. Appl. Climatol. 2018, 133, 1107–1118. [Google Scholar] [CrossRef]
- Hussain, S.; Huang, J.; Huang, J.; Ahmad, S.; Nanda, S.; Anwar, S.; Shakoor, A.; Zhu, C.; Zhu, L.; Cao, X.; et al. Rice Production under Climate Change: Adaptations and Mitigating Strategies. In Environment, Climate, Plant and Vegetation Growth; Fahad, S., Hasanuzzaman, M., Alam, M., Ullah, H., Saeed, M., Ali Khan, I., Adnan, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 659–686. [Google Scholar]
- Buckley, L.B.; Huey, R.B. Temperature extremes: Geographic patterns, recent changes, and implications for organismal vulnerabilities. Glob. Chang. Biol. 2016, 22, 3829–3842. [Google Scholar] [CrossRef]
- Krishnan, P.; Ramakrishnan, B.; Reddy, K.R.; Reddy, V. High-temperature effects on rice growth, yield, and grain quality. In Advances in agronomy; Elsevier: Amsterdam, The Netherlands, 2011; Volume 111, pp. 87–206. [Google Scholar]
- Ferguson, J.N.; Tidy, A.C.; Murchie, E.H.; Wilson, Z.A. The potential of resilient carbon dynamics for stabilizing crop reproductive development and productivity during heat stress. Plant Cell Environ. 2021, 44, 2066–2089. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Glob. Chang. Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Zhen, F.; Zhou, J.; Mahmood, A.; Wang, W.; Chang, X.; Liu, B.; Liu, L.; Cao, W.; Zhu, Y.; Tang, L. Quantifying the effects of short-term heat stress at booting stage on nonstructural carbohydrates remobilization in rice. Crop J. 2020, 8, 194–212. [Google Scholar] [CrossRef]
- Mahmood, A.; Ali, I.; Wang, W.; Ata-Ul-Karim, S.T.; Liu, B.; Liu, L.; Zhu, Y.; Cao, W.; Tang, L. Individual and Combined Effects of High-Temperature Stress at Booting and Flowering Stages on Rice Grain Yield. Agronomy 2022, 12, 3092. [Google Scholar] [CrossRef]
- Siddik, M.A.; Zhang, J.; Chen, J.; Qian, H.; Jiang, Y.; kareem Raheem, A.; Deng, A.; Song, Z.; Zheng, C.; Zhang, W. Responses of indica rice yield and quality to extreme high and low temperatures during the reproductive period. Eur. J. Agron. 2019, 106, 30–38. [Google Scholar] [CrossRef]
- Arshad, M.S.; Farooq, M.; Asch, F.; Krishna, J.S.; Prasad, P.V.; Siddique, K.H. Thermal stress impacts reproductive development and grain yield in rice. Plant Physiol. Biochem. 2017, 115, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Tang, L.; Lin, C.; Liu, L.; Wang, H.; Cao, W.; Zhu, Y. Modeling the effects of post-anthesis heat stress on rice phenology. Field Crops Res. 2015, 177, 26–36. [Google Scholar] [CrossRef]
- Wardlaw, I.F. Tansley Review No. 27 The control of carbon partitioning in plants. New Phytol. 1990, 116, 341–381. [Google Scholar] [CrossRef]
- Arai-Sanoh, Y.; Ida, M.; Zhao, R.; Yoshinaga, S.; Takai, T.; Ishimaru, T.; Maeda, H.; Nishitani, K.; Terashima, Y.; Gau, M. Genotypic variations in non-structural carbohydrate and cell-wall components of the stem in rice, sorghum, and sugar vane. Biosci. Biotechnol. Biochem. 2011, 75, 1105072478. [Google Scholar] [CrossRef]
- Zhang, J.; Li, G.; Huang, Q.; Liu, Z.; Ding, C.; Tang, S.; Chen, L.; Wang, S.; Ding, Y.; Zhang, W. Effects of culm carbohydrate partitioning on basal stem strength in a high-yielding rice population. Crop J. 2017, 5, 478–487. [Google Scholar] [CrossRef]
- Sasaki, H.; Hara, T.; Ito, S.; Uehara, N.; Kim, H.-Y.; Lieffering, M.; Okada, M.; Kobayashi, K. Effect of free-air CO2 enrichment on the storage of carbohydrate fixed at different stages in rice (Oryza sativa L.). Field Crops Res. 2007, 100, 24–31. [Google Scholar] [CrossRef]
- Shi, W.; Xiao, G.; Struik, P.C.; Jagadish, K.S.; Yin, X. Quantifying source-sink relationships of rice under high night-time temperature combined with two nitrogen levels. Field Crops Res. 2017, 202, 36–46. [Google Scholar] [CrossRef]
- Nagata, K.; Yoshinaga, S.; Takanashi, J.-I.; Terao, T. Effects of dry matter production, translocation of nonstructural carbohydrates and nitrogen application on grain filling in rice cultivar Takanari, a cultivar bearing a large number of spikelets. Plant Prod. Sci. 2001, 4, 173–183. [Google Scholar] [CrossRef]
- Julius, B.T.; Leach, K.A.; Tran, T.M.; Mertz, R.A.; Braun, D.M. Sugar transporters in plants: New insights and discoveries. Plant Cell Physiol. 2017, 58, 1442–1460. [Google Scholar] [CrossRef]
- Ntanos, D.; Koutroubas, S. Dry matter and N accumulation and translocation for Indica and Japonica rice under Mediterranean conditions. Field Crops Res. 2002, 74, 93–101. [Google Scholar] [CrossRef]
- Okawa, S.; Makino, A.; Mae, T. Effect of irradiance on the partitioning of assimilated carbon during the early phase of grain filling in rice. Ann. Bot. 2003, 92, 357–364. [Google Scholar] [CrossRef]
- Xiong, D.; Yu, T.; Ling, X.; Fahad, S.; Peng, S.; Li, Y.; Huang, J. Sufficient leaf transpiration and nonstructural carbohydrates are beneficial for high-temperature tolerance in three rice (Oryza sativa) cultivars and two nitrogen treatments. Funct. Plant Biol. 2015, 42, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-X.; Feng, B.-H.; Chen, T.-T.; Fu, W.-M.; Li, H.-B.; Li, G.-Y.; Jin, Q.-Y.; Tao, L.-X.; Fu, G.-F. Heat stress-reduced kernel weight in rice at anthesis is associated with impaired source-sink relationship and sugars allocation. Environ. Exp. Bot. 2018, 155, 718–733. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Liu, F.; Jin, M.; Yu, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Pre-anthesis high temperature acclimation alleviates the negative effects of post-anthesis heat stress on stem stored carbohydrates remobilization and grain starch accumulation in wheat. J. Cereal Sci. 2012, 55, 331–336. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Jiang, D.; Liu, F.; Dai, T.; Cao, W. Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat. J. Plant Physiol. 2011, 168, 585–593. [Google Scholar] [CrossRef]
- Fan, Y.; Ma, C.; Huang, Z.; Abid, M.; Jiang, S.; Dai, T.; Zhang, W.; Ma, S.; Jiang, D.; Han, X. Heat priming during early reproductive stages enhances thermo-tolerance to post-anthesis heat stress via improving photosynthesis and plant productivity in winter wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 805. [Google Scholar] [CrossRef]
- Morita, S.; Nakano, H. Nonstructural carbohydrate content in the stem at full heading contributes to high performance of ripening in heat-tolerant rice cultivar Nikomaru. Crop Sci. 2011, 51, 818–828. [Google Scholar] [CrossRef]
- Moura, D.S.; Brito, G.G.; Campos, Â.D.; Moraes, Í.L.; Porto, F.G.; Teixeira, S.B.; Deuner, S. Non-structural carbohydrates accumulation in contrasting rice genotypes subjected to high night temperatures. J. Agric. Sci. 2017, 9, 302–315. [Google Scholar] [CrossRef]
- Sun, T.; Liu, B.; Hasegawa, T.; Liao, Z.; Tang, L.; Liu, L.; Cao, W.; Zhu, Y. Sink-source unbalance leads to abnormal partitioning of biomass and nitrogen in rice under extreme heat stress: An experimental and modeling study. Eur. J. Agron. 2023, 142, 126678. [Google Scholar] [CrossRef]
- Phan, T.; Ishibashi, Y.; Miyazaki, M.; Tran, H.; Okamura, K.; Tanaka, S.; Nakamura, J.; Yuasa, T.; Iwaya-Inoue, M. High Temperature-Induced Repression of the Rice Sucrose Transporter (Os SUT 1) and Starch Synthesis-Related Genes in Sink and Source Organs at Milky Ripening Stage Causes Chalky Grains. J. Agron. Crop Sci. 2013, 199, 178–188. [Google Scholar] [CrossRef]
- Fu, G.; Feng, B.; Zhang, C.; Yang, Y.; Yang, X.; Chen, T.; Zhao, X.; Zhang, X.; Jin, Q.; Tao, L. Heat stress is more damaging to superior spikelets than inferiors of rice (Oryza sativa L.) due to their different organ temperatures. Front. Plant Sci. 2016, 7, 1637. [Google Scholar] [CrossRef] [PubMed]
- Hermann, R.-D.; Gabriel, G.-V. Response of rice plants to heat stress during initiation of panicle primordia or grain-filling phases. J. Stress Physiol. Biochem. 2013, 9, 318–325. [Google Scholar]
- Chen, J.; Tang, L.; Shi, P.; Yang, B.; Sun, T.; Cao, W.; Zhu, Y. Effects of short-term high temperature on grain quality and starch granules of rice (Oryza sativa L.) at post-anthesis stage. Protoplasma 2017, 254, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Kjær, K.H.; Rosenqvist, E.; Yu, X.; Wu, Z.; Ottosen, C.O. Physiological response to heat stress during seedling and anthesis stage in tomato genotypes differing in heat tolerance. J. Agron. Crop Sci. 2017, 203, 68–80. [Google Scholar] [CrossRef]
- Kaushal, N.; Awasthi, R.; Gupta, K.; Gaur, P.; Siddique, K.H.; Nayyar, H. Heat-stress-induced reproductive failures in chickpea (Cicer arietinum) are associated with impaired sucrose metabolism in leaves and anthers. Funct. Plant Biol. 2013, 40, 1334–1349. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Huang, Z.; Wang, Z.; Yang, J.; Zhang, J. Pre-anthesis non-structural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice. Field Crops Res. 2011, 123, 170–182. [Google Scholar] [CrossRef]
- Li, G.; Pan, J.; Cui, K.; Yuan, M.; Hu, Q.; Wang, W.; Mohapatra, P.K.; Nie, L.; Huang, J.; Peng, S. Limitation of unloading in the developing grains is a possible cause responsible for low stem non-structural carbohydrate translocation and poor grain yield formation in rice through verification of recombinant inbred lines. Front. Plant Sci. 2017, 8, 1369. [Google Scholar] [CrossRef]
- Fan, Y.; Lv, Z.; Zhang, Y.; Ma, L.; Qin, B.; Liu, Q.; Zhang, W.; Ma, S.; Ma, C.; Huang, Z. Pre-anthesis night warming improves post-anthesis physiological activity and plant productivity to post-anthesis heat stress in winter wheat (Triticum aestivum L.). Environ. Exp. Bot. 2022, 197, 104819. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Lal, M.K.; Tiwari, R.K.; Gahlaut, V.; Mangal, V.; Kumar, A.; Singh, M.P.; Paul, V.; Kumar, S.; Singh, B.; Zinta, G. Physiological and molecular insights on wheat responses to heat stress. Plant Cell Rep. 2021, 41, 501–518. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, Q.; Wang, X.; Cai, J.; Dai, T.; Cao, W.; Jiang, D. Physiological and transcriptional analyses of induced post-anthesis thermo-tolerance by heat-shock pretreatment on germinating seeds of winter wheat. Environ. Exp. Bot. 2016, 131, 181–189. [Google Scholar] [CrossRef]
- Prasad, P.; Boote, K.; Allen Jr, L.; Sheehy, J.; Thomas, J. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res. 2006, 95, 398–411. [Google Scholar] [CrossRef]
- Sun, T.; Hasegawa, T.; Tang, L.; Wang, W.; Zhou, J.; Liu, L.; Liu, B.; Cao, W.; Zhu, Y. Stage-dependent temperature sensitivity function predicts seed-setting rates under short-term extreme heat stress in rice. Agric. For. Meteorol. 2018, 256, 196–206. [Google Scholar] [CrossRef]
- Shi, W.; Lawas, L.; Raju, B.; Jagadish, S. Acquired thermo-tolerance and trans-generational heat stress response at flowering in rice. J. Agron. Crop Sci. 2016, 202, 309–319. [Google Scholar] [CrossRef]
- Bheemanahalli, R.; Sathishraj, R.; Tack, J.; Nalley, L.L.; Muthurajan, R.; Jagadish, K.S. Temperature thresholds for spikelet sterility and associated warming impacts for sub-tropical rice. Agric. For. Meteorol. 2016, 221, 122–130. [Google Scholar] [CrossRef]
- Jagadish, S.K.; Craufurd, P.Q.; Wheeler, T. High temperature stress and spikelet fertility in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Nath, I. Climate change, the food problem, and the challenge of adaptation through sectoral reallocation. In Proceedings of the 25th Annual Conference on Global Economic Analysis (Virtual Conference), Virtual, 8–10 June 2022. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998; Volume 300, p. D05109. [Google Scholar]
- Yoshida, S. Laboratory Manual for Physiological Studies of Rice; International Rice Research Institute: Los Baños, Philippines, 1976; Volume 23, pp. 61–66. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Cultivar | Treatment | NSCs Accumulation at Maturity (g/plant) | NSCs Translocation (g/plant) | NSCs Translocation Efficiency (%) | Grain Yield per Plant (g) | Contribution of Stem NSCs to Grain Yield (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| Leaf | Stem | Panicle | |||||||
| Booting | HD-5 | T1D2 | 0.67 b | 2.02 e | 13.45 a | 2.90 a | 58.93 a | 14.88 a | 19.49 a |
| T2D1 | 0.71 b | 3.85 d | 10.59 b | 1.07 b | 21.73 b | 12.69 b | 8.56 ab | ||
| T2D2 | 0.72 b | 5.20 c | 8.44 c | −0.28 c | −5.65 c | 11.06 c | −2.38 b | ||
| T3D1 | 0.83 ab | 11.15 b | 4.23 d | −6.23 d | −126.57 d | 7.05 d | −88.66 c | ||
| T3D2 | 0.92 a | 12.70 a | 3.01 e | −7.78 e | −158.16 e | 5.94 e | −131.79 d | ||
| WYJ-24 | T1D2 | 0.63 b | 1.61 d | 13.06 a | 2.99 a | 64.99 a | 15.53 a | 19.33 a | |
| T2D1 | 0.64 b | 2.81 cd | 11.64 b | 1.79 ab | 38.94 ab | 14.03 b | 12.82 a | ||
| T2D2 | 0.66 b | 3.94 c | 9.09 c | 0.66 b | 14.38 b | 11.69 c | 5.59 a | ||
| T3D1 | 0.76 ab | 7.25 b | 6.91 d | −2.65 c | −57.52 c | 10.03 d | −26.89 b | ||
| T3D2 | 0.87 a | 8.64 a | 5.30 e | −4.04 d | −87.75 d | 8.58 e | −47.81 c | ||
| Flowering | HD-5 | T1D2 | 0.67 b | 2.02 e | 13.62 a | 2.90 a | 58.96 a | 14.82 a | 19.62 a |
| T2D1 | 0.68 b | 4.86 d | 9.51 b | 0.06 b | 1.21 b | 12.30 b | 0.53 b | ||
| T2D2 | 0.75 b | 7.90 c | 4.35 c | −2.98 c | −60.5 7c | 6.25 c | −47.54 c | ||
| T3D1 | 0.86 a | 13.50 b | 1.68 d | −8.58 d | −174.40 d | 4.53 d | −189.31 d | ||
| T3D2 | 0.93 a | 15.91 a | 0.85 d | −10.99 e | −223.33 e | 2.70 e | −408.54 e | ||
| WYJ-24 | T1D2 | 0.63 c | 1.62 e | 12.90 a | 2.98 a | 64.83 a | 14.78 a | 20.19 a | |
| T2D1 | 0.64 c | 4.15 d | 9.64 b | 0.45 b | 9.81 b | 12.47 b | 3.54 a | ||
| T2D2 | 0.64 c | 6.58 c | 5.61 c | −1.98 c | −43.15 c | 7.62 c | −26.04 a | ||
| T3D1 | 0.76 b | 10.66 b | 3.01 d | −6.06 d | −131.68 d | 6.85 c | −89.52 b | ||
| T3D2 | 0.86 a | 13.15 a | 1.03 e | −8.55 e | −185.97 e | 2.88 d | −296.55 c | ||
| Combined | HD-5 | T1D2 | 0.68 b | 2.02 e | 13.55 a | 2.90 a | 58.97 a | 15.25 a | 19.09 a |
| T2D1 | 0.69 b | 4.31 d | 9.73 b | 0.61 b | 12.45 b | 12.18 b | 5.04 a | ||
| T2D2 | 0.83 ab | 7.46 c | 4.86 c | −2.54 c | −51.65 c | 6.71 c | −38.13 b | ||
| T3D1 | 0.88 ab | 13.13 b | 2.54 d | −8.21 d | −166.87 d | 5.69 c | −145.88 c | ||
| T3D2 | 0.96 a | 15.56 a | 1.38 e | −10.64 e | −216.27 e | 3.70 d | −288.91 d | ||
| WYJ-24 | T1D2 | 0.63 c | 1.61 e | 12.94 a | 2.99 a | 64.96 a | 14.89 a | 20.04 a | |
| T2D1 | 0.64 c | 3.67 d | 10.64 b | 0.93 b | 20.23 b | 13.16 b | 7.01 a | ||
| T2D2 | 0.66 c | 6.22 c | 6.75 c | −1.62 c | −35.16 c | 8.81 c | −18.69 b | ||
| T3D1 | 0.79 b | 10.39 b | 3.85 d | −5.79 d | −125.81 d | 7.03 d | −82.27 c | ||
| T3D2 | 0.90 a | 12.67 a | 1.79 e | −8.07 e | −175.55 e | 3.89 e | −207.83 d | ||
| Stage | Cultivar | Treatment | NSCs Accumulation at Maturity (g/plant) | NSCs Translocation (g/plant) | NSCs Translocation Efficiency (%) | Grain Yield per Plant (g) | Contribution of Stem NSCs to Grain Yield (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| Leaf | Stem | Panicle | |||||||
| Booting | HD-5 | T1D2 | 0.67 b | 1.94 c | 13.56 a | 2.95 a | 60.33 a | 15.25 a | 19.33 a |
| T2D1 | 0.69 b | 4.12 b | 11.01 b | 0.76 b | 15.57 b | 13.13 b | 6.00 a | ||
| T2D2 | 0.70 b | 5.57 b | 8.78 c | −0.68 b | −13.95 b | 11.44 c | −5.84 a | ||
| T3D1 | 0.85 ab | 11.79 a | 4.15 d | −6.91 c | −141.42 c | 7.16 d | −97.25 b | ||
| T3D2 | 0.94 a | 13.21 a | 2.79 e | −8.32 c | −170.30 c | 5.45 e | −154.40 c | ||
| WYJ-24 | T1D2 | 0.64 b | 1.63 c | 13.37 a | 2.89 a | 63.95 a | 16.23 a | 17.85 a | |
| T2D1 | 0.65 b | 2.76 bc | 12.00 b | 1.76 ab | 38.98 ab | 14.35 b | 12.34 a | ||
| T2D2 | 0.67 b | 3.90 b | 9.44 c | 0.62 b | 13.64 b | 12.04 c | 5.08 a | ||
| T3D1 | 0.80ab | 7.49 a | 6.84 d | −2.97 c | −65.71 c | 10.22 d | −29.52 b | ||
| T3D2 | 0.88 a | 8.61 a | 5.70 d | −4.09 c | −90.61 c | 9.10 d | −45.58 c | ||
| Flowering | HD-5 | T1D2 | 0.67 b | 1.94 e | 13.64 a | 2.95 a | 60.34 a | 15.19 a | 19.42 a |
| T2D1 | 0.69 b | 5.07 d | 9.09 b | −0.18 b | −3.68 b | 11.77 b | −1.46 ab | ||
| T2D2 | 0.76 b | 8.15 c | 4.54 c | −3.26 c | −66.76 c | 6.51 c | −50.02 b | ||
| T3D1 | 0.90 a | 14.45 b | 1.18 d | −9.57 d | −195.85 d | 3.30 d | −290.83 c | ||
| T3D2 | 0.96 a | 16.42 a | 0.39 d | −11.53 e | −236.08 e | 1.21 e | −961.25 d | ||
| WYJ-24 | T1D2 | 0.63 c | 1.65 e | 13.50 a | 2.87 a | 63.42 a | 15.65 a | 18.28 a | |
| T2D1 | 0.63 c | 3.77 d | 10.70 b | 0.75 b | 16.59 b | 13.79 b | 5.44 a | ||
| T2D2 | 0.64 c | 6.60 c | 6.36 c | −2.09 c | −46.16 c | 8.59 c | −24.27 a | ||
| T3D1 | 0.79 b | 12.12 b | 1.66 d | −7.60 d | −168.27 d | 3.94 d | −193.02 b | ||
| T3D2 | 0.89 a | 13.26 a | 0.83 d | −8.74 e | −193.43 e | 2.27 e | −387.29 c | ||
| Combined | HD-5 | T1D2 | 0.66 c | 1.94 e | 13.64 a | 2.95 a | 60.35 a | 15.41 a | 19.20 a |
| T2D1 | 0.69 bc | 4.31 d | 9.89 b | 0.57 b | 11.73 b | 12.28 b | 4.64 a | ||
| T2D2 | 0.75 abc | 7.84 c | 4.95 c | −2.95 c | −60.40 c | 6.80 c | −43.42 b | ||
| T3D1 | 0.91 ab | 13.72 b | 1.97 d | −8.84 d | −180.91 d | 4.44 d | −199.00 c | ||
| T3D2 | 0.96 a | 15.79 a | 1.09 e | −10.91 e | −223.26e | 2.84 e | −384.96 d | ||
| WYJ-24 | T1D2 | 0.64 b | 1.66 e | 13.35 a | 2.86 a | 63.36 a | 15.53 a | 18.39 a | |
| T2D1 | 0.65 b | 3.57 d | 10.94 b | 0.95 b | 20.93 b | 13.77 b | 6.81 a | ||
| T2D2 | 0.69 b | 6.31 c | 6.71 c | −1.80 c | −39.76 c | 8.73 c | −20.93 a | ||
| T3D1 | 0.81 a | 11.61 b | 2.26 d | −7.09 d | −156.93 d | 4.28 d | −167.14 b | ||
| T3D2 | 0.90 a | 12.96 a | 1.31 d | −8.44 e | −186.80 e | 2.79 e | −302.90 c | ||
| Year | Site | Cultivar | Timing of Treatment | Temperature Level & Duration |
|---|---|---|---|---|
| 2016 | Rugao | HD-5 | Booting (BT) (11 August 2016; 12 August 2017) S1 Flowering (FL) (23 August 2016; 26 August 2017) S2 Combined (BT + FL) S3 | [T1, T2, T3] × [D1, D2] |
| WYJ-24 | ||||
| 2017 | Rugao | HD-5 | booting (18 August 2016; 24 August 2017) S1 flowering (1 September 2016; 5 September 2017) S2 Combined (BT + FL) S3 | |
| WYJ-24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmood, A.; Wang, W.; Raza, M.A.; Ali, I.; Liu, B.; Liu, L.; Zhu, Y.; Tang, L.; Cao, W. Quantifying the Individual and Combined Effects of Short-Term Heat Stress at Booting and Flowering Stages on Nonstructural Carbohydrates Remobilization in Rice. Plants 2024, 13, 810. https://doi.org/10.3390/plants13060810
Mahmood A, Wang W, Raza MA, Ali I, Liu B, Liu L, Zhu Y, Tang L, Cao W. Quantifying the Individual and Combined Effects of Short-Term Heat Stress at Booting and Flowering Stages on Nonstructural Carbohydrates Remobilization in Rice. Plants. 2024; 13(6):810. https://doi.org/10.3390/plants13060810
Chicago/Turabian StyleMahmood, Aqib, Wei Wang, Muhammad Ali Raza, Iftikhar Ali, Bing Liu, Leilei Liu, Yan Zhu, Liang Tang, and Weixing Cao. 2024. "Quantifying the Individual and Combined Effects of Short-Term Heat Stress at Booting and Flowering Stages on Nonstructural Carbohydrates Remobilization in Rice" Plants 13, no. 6: 810. https://doi.org/10.3390/plants13060810
APA StyleMahmood, A., Wang, W., Raza, M. A., Ali, I., Liu, B., Liu, L., Zhu, Y., Tang, L., & Cao, W. (2024). Quantifying the Individual and Combined Effects of Short-Term Heat Stress at Booting and Flowering Stages on Nonstructural Carbohydrates Remobilization in Rice. Plants, 13(6), 810. https://doi.org/10.3390/plants13060810