
Physiological and Molecular Response of Liriodendron chinense to Varying Stand Density

Abstract
1. Introduction
2. Results
2.1. Impact of FSD on the Leaf Anatomy of L. chinense
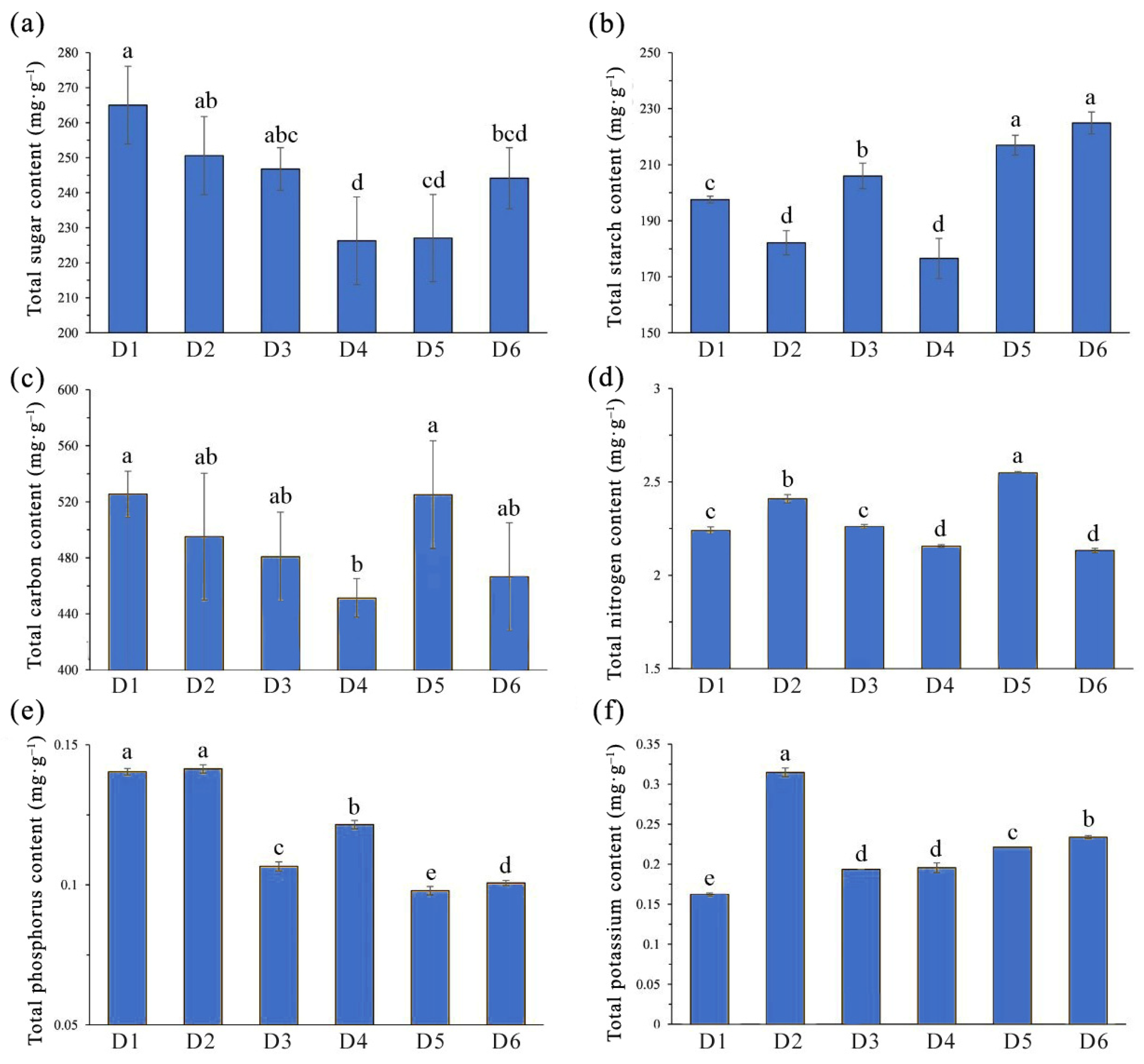
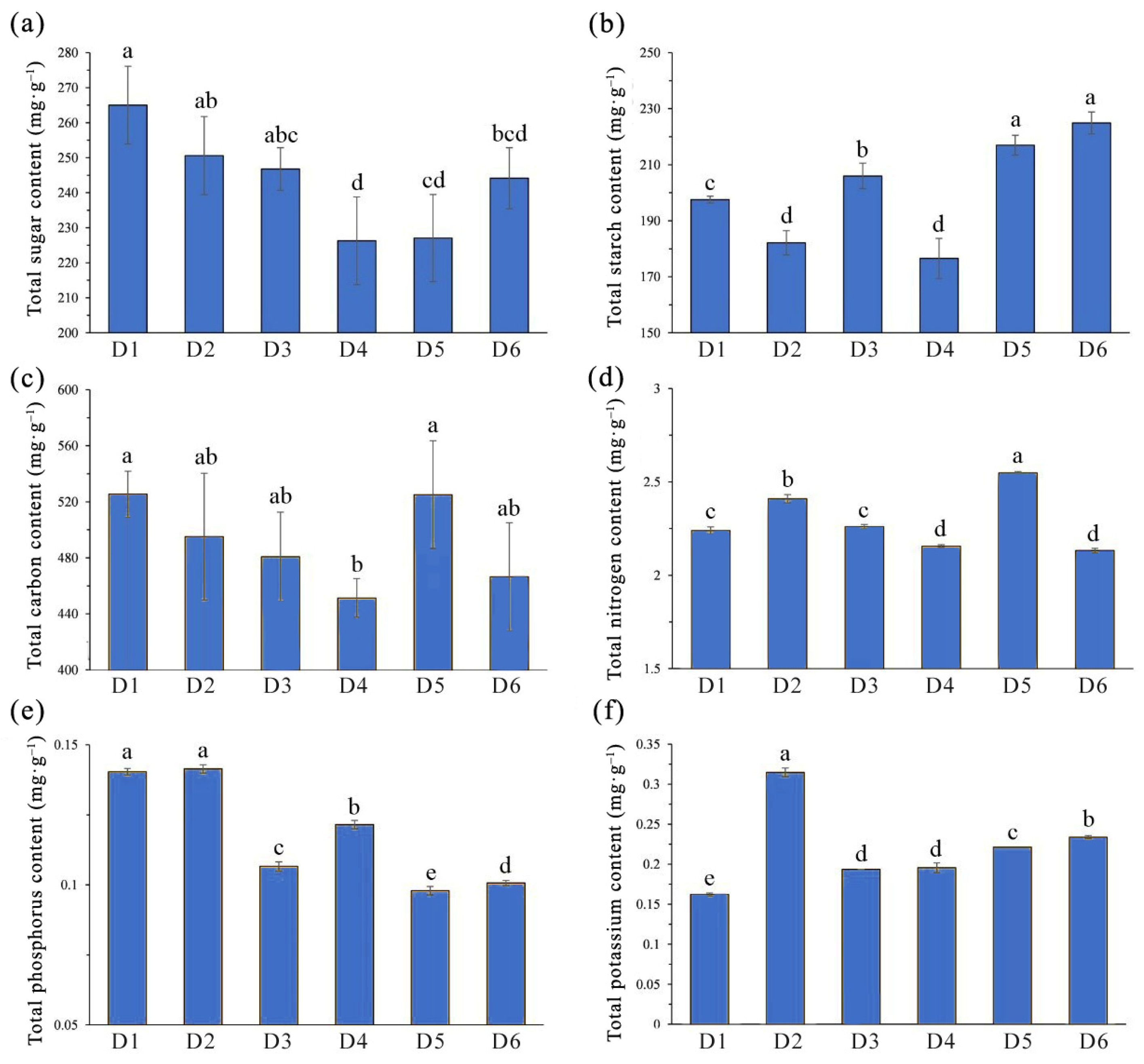
2.2. Influence of FSD on Biomass and Nutrient Accumulation in L. chinense
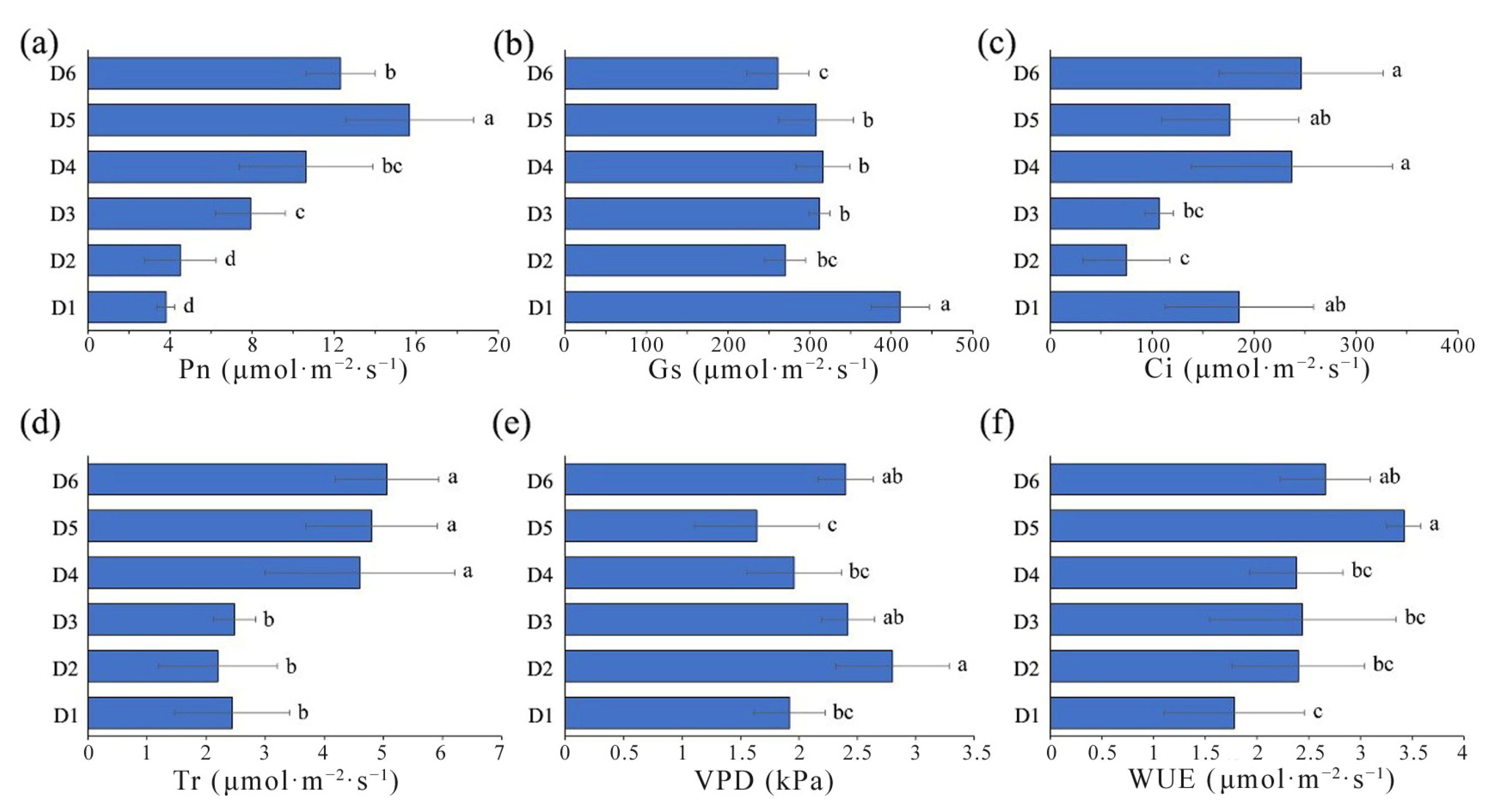
2.3. Comparison of Photosynthetic Characteristics in L. chinense under Different FSDs
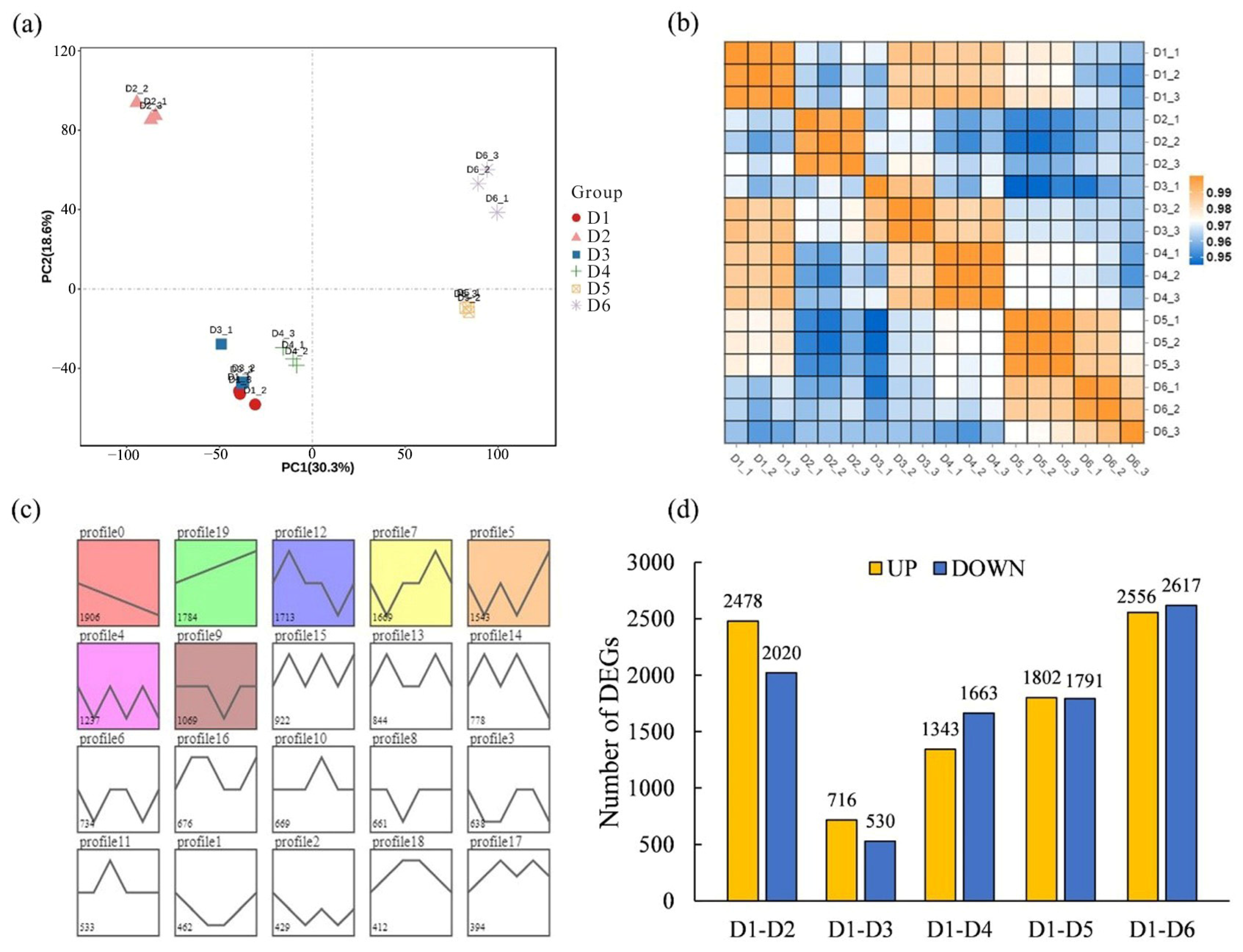
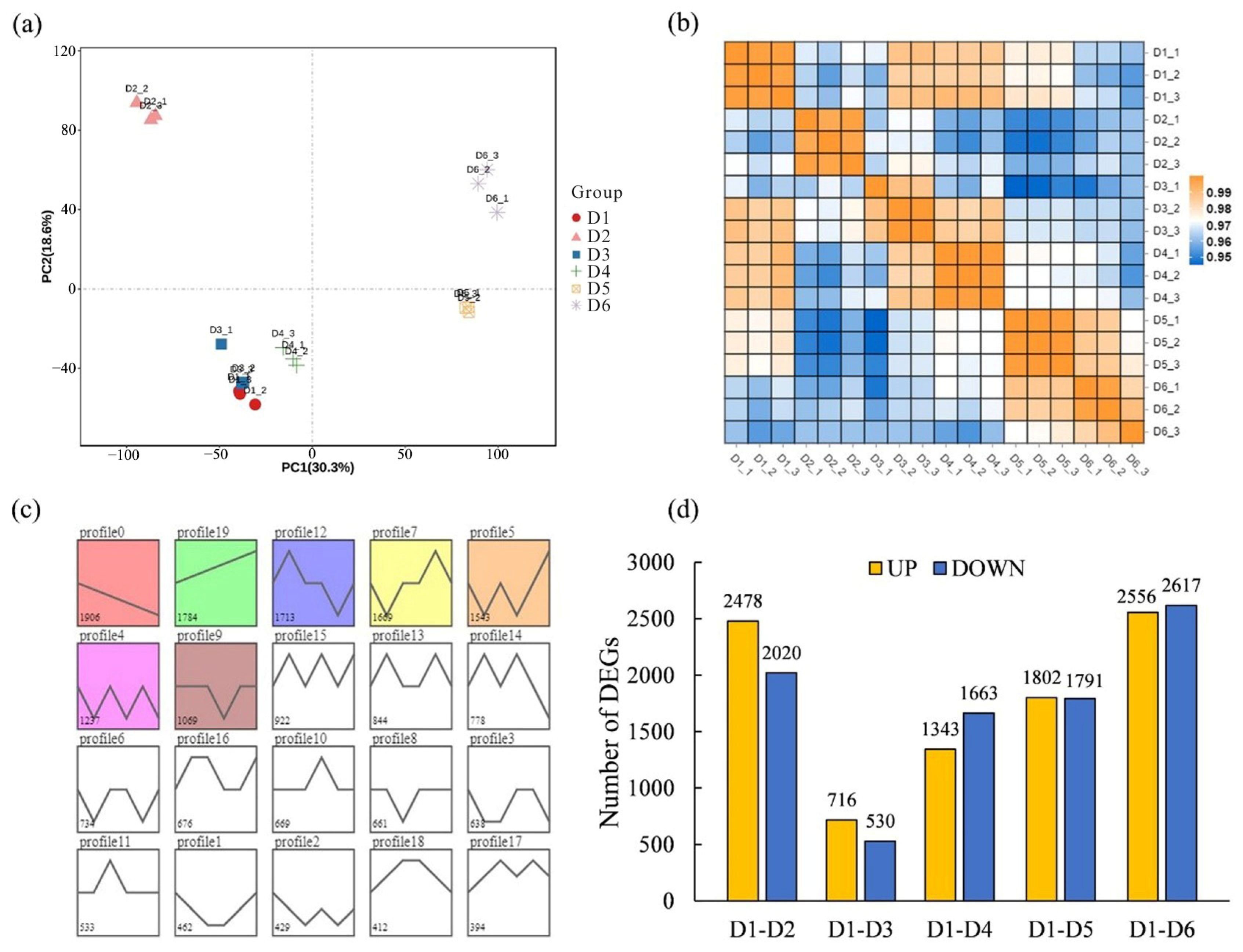
2.4. Differential Transcriptome Analyses of Different FSD Samples
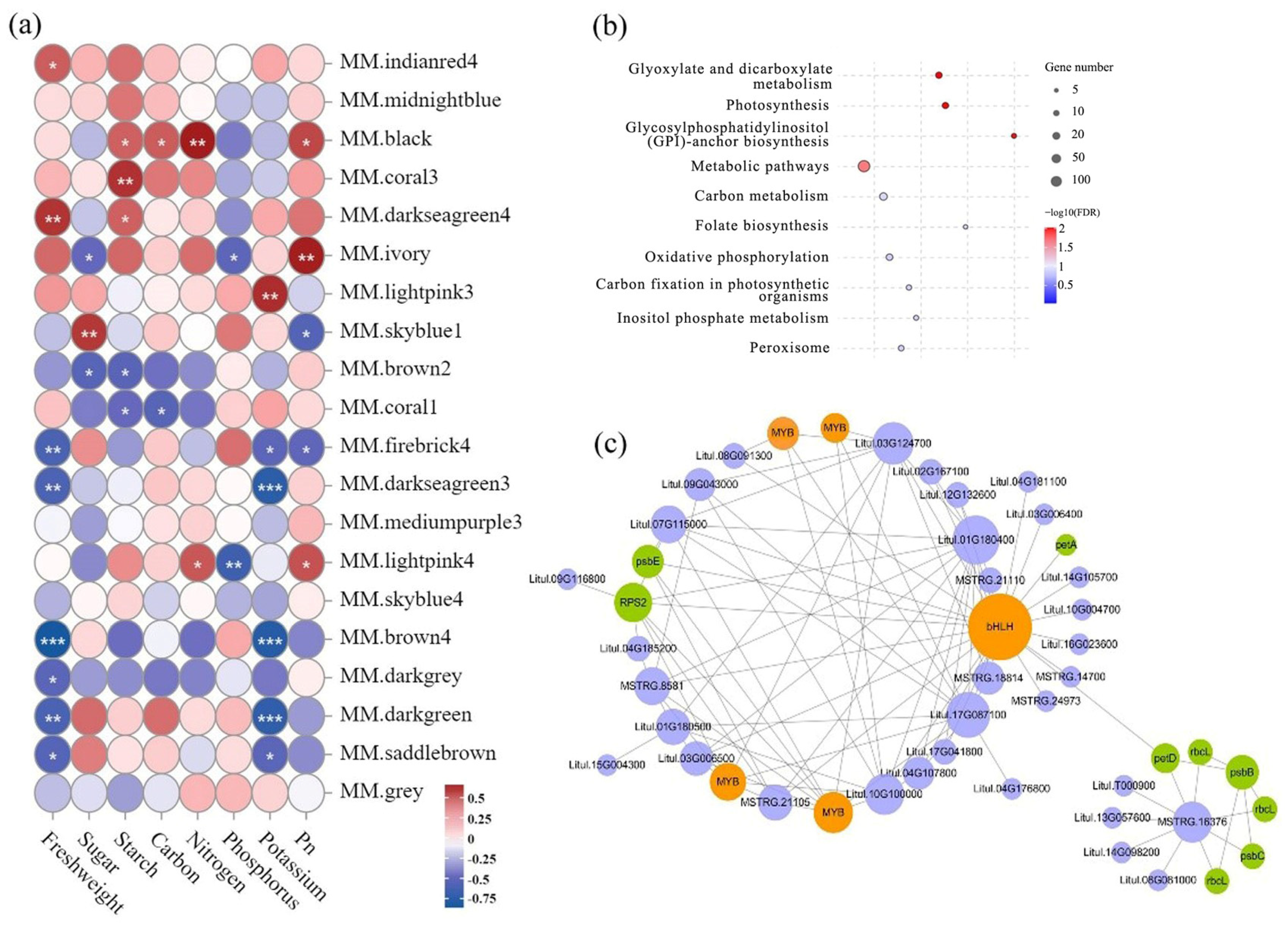
2.5. Analyses of the Co-Expression Network of Photosynthesis-Related Genes
3. Discussion
4. Materials and Methods
4.1. Overview of the Experimental Site
4.2. Experimental Materials and Design
4.3. Observation of Microscopic Structure
4.4. Measurement of Leaf Traits
4.5. Determination of the Nutritional Elements in Leaves
4.6. Measurement of Photosynthetic Parameters
4.7. Sequencing and Analyses of the Transcriptome
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Niinemets, U. Photosynthesis and resource distribution through plant canopies. Plant Cell Environ. 2007, 30, 1052–1071. [Google Scholar] [CrossRef]
- Evans, J.R. Improving photosynthesis. Plant Physiol. 2013, 162, 1780–1793. [Google Scholar] [CrossRef]
- Gower, S.T.; Kucharik, C.J.; Norman, J.M. Direct and Indirect Estimation of Leaf Area Index, fAPAR, and Net Primary Production of Terrestrial Ecosystems. Remote Sens. Environ. 1999, 70, 29–51. [Google Scholar] [CrossRef]
- Will, R.E.; Barron, G.A.; Burkes, E.C.; Shiver, B.; Teskey, R.O. Relationship between intercepted radiation, net photosynthesis, respiration, and rate of stem volume growth of Pinus taeda and Pinus elliottii stands of different densities. For. Ecol. Manag. 2001, 154, 155–163. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Wang, X.; Xiao, W.; Chen, L.; Li, S.; Sun, H.; Deng, X.; Forrester, D.I.; Zeng, L.; et al. Effects of stand age, richness and density on productivity in subtropical forests in China. J. Ecol. 2019, 107, 2266–2277. [Google Scholar] [CrossRef]
- Gong, C.M.; Bai, J.; Deng, J.M.; Wang, G.X.; Liu, X.P. Leaf anatomy and photosynthetic carbon metabolic characteristics in Phragmites communis in different soil water availability. Plant Ecol. 2011, 212, 675–687. [Google Scholar] [CrossRef]
- Ouyang, W.; Struik, P.C.; Yin, X.; Yang, J. Stomatal conductance, mesophyll conductance, and transpiration efficiency in relation to leaf anatomy in rice and wheat genotypes under drought. J. Exp. Bot. 2017, 68, 5191–5205. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Hussain, M.; Ul-Allah, S.; Siddique, K.H. Physiological and agronomic approaches for improving water-use efficiency in crop plants. Agric. Water Manag. 2019, 219, 95–108. [Google Scholar] [CrossRef]
- Chen, J.; Hao, Z.; Guang, X.; Zhao, C.; Wang, P.; Xue, L.; Zhu, Q.; Yang, L.; Sheng, Y.; Zhou, Y.; et al. Liriodendron genome sheds light on angiosperm phylogeny and species–pair differentiation. Nat. Plants 2019, 5, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.B.W. Principles of transcriptome analysis and gene expression quantification: An RNA-seq tutorial. Mol. Ecol. Resour. 2013, 13, 559–572. [Google Scholar] [CrossRef]
- Janiak, A.; Kwasniewski, M.; Sowa, M.; Kuczyńska, A.; Mikołajczak, K.; Ogrodowicz, P.; Szarejko, I. Insights into Barley Root Transcriptome under Mild Drought Stress with an Emphasis on Gene Expression Regulatory Mechanisms. Int. J. Mol. Sci. 2019, 20, 6139. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Hong, W.J.; Kim, Y.-J.; Chandran, A.K.N.; Gho, Y.S.; Yoo, Y.H.; Nguyen, V.N.T.; An, G.; Park, S.K.; Jung, K.-H. Comparative Transcriptome Analysis Reveals Gene Regulatory Mechanism of UDT1 on Anther Development. J. Plant Biol. 2020, 63, 289–296. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y.; Zhang, Z.; Fang, S. Physiological response and molecular regulatory mechanism reveal a positive role of nitric oxide and hydrogen sulfide applications in salt tolerance of Cyclocarya paliurus. Front. Plant Sci. 2023, 14, 1211162. [Google Scholar] [CrossRef] [PubMed]
- Spicer, R.; Groover, A. Evolution of development of vascular cambia and secondary growth. New Phytol. 2010, 186, 577–592. [Google Scholar] [CrossRef]
- Bräutigam, K.; Vining, K.J.; Lafon-Placette, C.; Fossdal, C.G.; Mirouze, M.; Marcos, J.G.; Fluch, S.; Fraga, M.F.; Guevara, M.Á.; Abarca, D.; et al. Epigenetic regulation of adaptive responses of forest tree species to the environment. Ecol. Evol. 2013, 3, 399–415. [Google Scholar] [CrossRef]
- Hao, Z.; Wang, X.; Zong, Y.; Wen, S.; Cheng, Y.; Li, H. Enzymatic activity and functional analysis under multiple abiotic stress conditions of a dehydroascorbate reductase gene derived from Liriodendron Chinense. Environ. Exp. Bot. 2019, 167, 103850. [Google Scholar] [CrossRef]
- Hao, Z.; Liu, S.; Hu, L.; Shi, J.; Chen, J. Transcriptome analysis and metabolic profiling reveal the key role of carotenoids in the petal coloration of Liriodendron tulipifera. Hortic. Res. 2020, 7, 70. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, J.; Chen, X.; Zhang, J.; Lu, L.; Hao, Z.; Shi, J.; Chen, J. PYL Family Genes from Liriodendron chinense Positively Respond to Multiple Stresses. Plants 2023, 12, 2609. [Google Scholar] [CrossRef] [PubMed]
- Rötzer, T.; Grote, R.; Pretzsch, H. Effects of environmental changes on the vitality of forest stands. Eur. J. For. Res. 2005, 124, 349–362. [Google Scholar] [CrossRef]
- Fu, B.; Li, S.; Yu, X.; Yang, P.; Yu, G.; Feng, R.; Zhuang, X. Chinese ecosystem research network: Progress and perspectives. Ecol. Complex. 2010, 7, 225–233. [Google Scholar] [CrossRef]
- Sadeghi, S.M.M.; Attarod, P.; Van Stan, J.T.; Pypker, T.G. The importance of considering rainfall partitioning in afforestation initiatives in semiarid climates: A comparison of common planted tree species in Tehran, Iran. Sci. Total. Environ. 2016, 568, 845–855. [Google Scholar] [CrossRef]
- Mohler, C.L.; Marks, P.L.; Sprugel, D.G. Stand Structure and Allometry of Trees During Self-Thinning of Pure Stands. J. Ecol. 1978, 66, 599–614. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G. Effect of tree species mixing on the size structure, density, and yield of forest stands. Eur. J. For. Res. 2016, 135, 1–22. [Google Scholar] [CrossRef]
- Ashton, M.S.; Kelty, M.J. The Practice of Silviculture: Applied Forest Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Kholdaenko, Y.A.; Belokopytova, L.V.; Zhirnova, D.F.; Upadhyay, K.K.; Tripathi, S.K.; Koshurnikova, N.N.; Sobachkin, R.S.; Babushkina, E.A.; Vaganov, E.A. Stand density effects on tree growth and climatic response in Picea obovata Ledeb. plantations. For. Ecol. Manag. 2022, 519, 120349. [Google Scholar] [CrossRef]
- McDowell, N.G.; Adams, H.D.; Bailey, J.D.; Kolb, T.E. The role of stand density on growth efficiency, leaf area index, and resin flow in southwestern ponderosa pine forests. Can. J. For. Res. 2007, 37, 343–355. [Google Scholar] [CrossRef]
- Falster, D.S.; Brännström, Å.; Dieckmann, U.; Westoby, M. Influence of four major plant traits on average height, leaf-area cover, net primary productivity, and biomass density in single-species forests: A theoretical investigation. J. Ecol. 2011, 99, 148–164. [Google Scholar] [CrossRef]
- Evans, J.R.; Poorter, H. Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 2001, 24, 755–767. [Google Scholar] [CrossRef]
- Hakamada, R.E.; Hubbard, R.M.; Moreira, G.G.; Stape, J.L.; Campoe, O.; Ferraz, S.F.d.B. Influence of stand density on growth and water use efficiency in Eucalyptus clones. For. Ecol. Manag. 2020, 466, 118125. [Google Scholar] [CrossRef]
- Andrews, C.M.; D’Amato, A.W.; Fraver, S.; Palik, B.; Battaglia, M.A.; Bradford, J.B. Low stand density moderates growth declines during hot droughts in semi-arid forests. J. Appl. Ecol. 2020, 57, 1089–1102. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Condés, S.; Del Río, M.; Pretzsch, H.; Ducey, M.J. Maximum stand density strongly depends on species-specific wood stability, shade and drought tolerance. For. Int. J. For. Res. 2018, 91, 459–469. [Google Scholar] [CrossRef]
- Thevathasan, N.; Coleman, B.; Zabek, L.; Ward, T.; Gordon, A. Agroforestry in Canada and its role in farming systems. In Temperate Agroforestry Systems; CAB International: Wallingford, UK, 2018; pp. 7–49. [Google Scholar]
- Lambers, H.; Oliveira, R.S.; Lambers, H.; Oliveira, R.S. Photosynthesis, respiration, and long-distance transport: Photosynthesis. In Plant Physiological Ecology; Springer: Cham, Switzerland, 2019; pp. 11–114. [Google Scholar]
- Pantazopoulou, C.K.; Bongers, F.J.; Pierik, R. Reducing shade avoidance can improve Arabidopsis canopy performance against competitors. Plant Cell Environ. 2021, 44, 1130–1141. [Google Scholar] [CrossRef] [PubMed]
- Li, X.B.; Liu, C.C.; Chen, J.X.; Zhang, M.M.; Zhang, J.H.; Tong, Z.K.; Yang, Q. Leaf Plasticity of the Subtropical Evergreen Tree Phoebe bournei Increases with Ontogeny in Response to Sun and Shade. Forests 2023, 14, 1683. [Google Scholar] [CrossRef]
- Uhl, E.; Biber, P.; Ulbricht, M.; Heym, M.; Horváth, T.; Lakatos, F.; Gál, J.; Steinacker, L.; Tonon, G.; Ventura, M.; et al. Analysing the effect of stand density and site conditions on structure and growth of oak species using Nelder trials along an environmental gradient: Experimental design, evaluation methods, and results. For. Ecosyst. 2015, 2, 17. [Google Scholar] [CrossRef]
- Hechter, U.; Little, K.; Chan, J.; Crous, J.; da Costa, D. Factors affecting eucalypt survival in South African plantation forestry. South. For. J. For. Sci. 2022, 84, 253–270. [Google Scholar] [CrossRef]
- Ning, K.; Ding, C.; Huang, Q.; Zhang, W.; Yang, C.; Liang, D.; Fan, R.; Su, X. Transcriptome profiling revealed diverse gene expression patterns in poplar (Populus × euramericana) under different planting densities. PLoS ONE 2019, 14, e0217066. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Pei, G.; Chen, L.; Zhang, W. WGCNA application to proteomic and metabolomic data analysis. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 585, pp. 135–158. [Google Scholar]
- Du, H.; Zhang, L.; Liu, L.; Tang, X.F.; Yang, W.J.; Wu, Y.M.; Huang, Y.B.; Tang, Y.X. Biochemical and molecular characterization of plant MYB transcription factor family. Biochemistry 2009, 74, 1–11. [Google Scholar] [CrossRef]
- Cao, Y.; Li, K.; Li, Y.; Zhao, X.; Wang, L. MYB Transcription Factors as Regulators of Secondary Metabolism in Plants. Biology 2020, 9, 61. [Google Scholar] [CrossRef]
- Cai, Y.; Liu, Y.; Fan, Y.; Li, X.; Yang, M.; Xu, D.; Wang, H.; Deng, X.W.; Li, J. MYB112 connects light and circadian clock signals to promote hypocotyl elongation in Arabidopsis. Plant Cell 2023, 35, 3485–3503. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Cai, K.; Zhang, Q.; Jiang, L.; Li, H.; Lv, Y.; Qu, G.; Zhao, X. Comparative Transcriptomic and Metabolic Analyses Reveal the Coordinated Mechanisms in Pinus koraiensis under Different Light Stress Conditions. Int. J. Mol. Sci. 2022, 23, 9556. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parts | Indicator (μm) | Samples | Mean | |||||
|---|---|---|---|---|---|---|---|---|
| D1 | D2 | D3 | D4 | D5 | D6 | |||
| Upper epidermis | Stratum corneum thickness | 3.70 a | 3.80 a | 3.50 a | 3.73 a | 2.97 b | 2.93 b | 3.44 |
| Cell length | 36.53 a | 36.17 a | 31.73 ab | 31.70 ab | 28.73 b | 28.77 b | 32.27 | |
| Cell width | 14.33 a | 13.53 b | 13.67 ab | 14.60 a | 10.33 c | 9.93 c | 12.73 | |
| Cell length/width | 2.54 bc | 2.67 ab | 2.33 cd | 2.17 d | 2.78 ab | 2.90 a | 2.57 | |
| Lower epidermis | Stratum corneum thickness | 2.93 c | 2.27 d | 3.27 c | 4.27 a | 3.93 ab | 3.50 bc | 3.36 |
| Cell length | 30.67 a | 27.63 a | 27.57 a | 29.63 a | 27.60 a | 29.57 a | 28.78 | |
| Cell width | 13.27 ab | 12.17 b | 13.20 ab | 13.97 a | 9.57 c | 10.47 c | 12.11 | |
| Cell length/width | 2.30 b | 2.28 b | 2.09 b | 2.12 b | 2.88 a | 2.83 a | 2.42 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Li, T.; Cai, J.; Yu, P.; Guo, Y. Physiological and Molecular Response of Liriodendron chinense to Varying Stand Density. Plants 2024, 13, 508. https://doi.org/10.3390/plants13040508
Chen J, Li T, Cai J, Yu P, Guo Y. Physiological and Molecular Response of Liriodendron chinense to Varying Stand Density. Plants. 2024; 13(4):508. https://doi.org/10.3390/plants13040508
Chicago/Turabian StyleChen, Jun, Ting Li, Jinfeng Cai, Pengfei Yu, and Ying Guo. 2024. "Physiological and Molecular Response of Liriodendron chinense to Varying Stand Density" Plants 13, no. 4: 508. https://doi.org/10.3390/plants13040508
APA StyleChen, J., Li, T., Cai, J., Yu, P., & Guo, Y. (2024). Physiological and Molecular Response of Liriodendron chinense to Varying Stand Density. Plants, 13(4), 508. https://doi.org/10.3390/plants13040508