

Comparative Analysis of Ca2+/Cation Antiporter Gene Family in Rosa roxburghii and Enhanced Calcium Stress Tolerance via Heterologous Expression of RrCAX1a in Tobacco

 ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Result
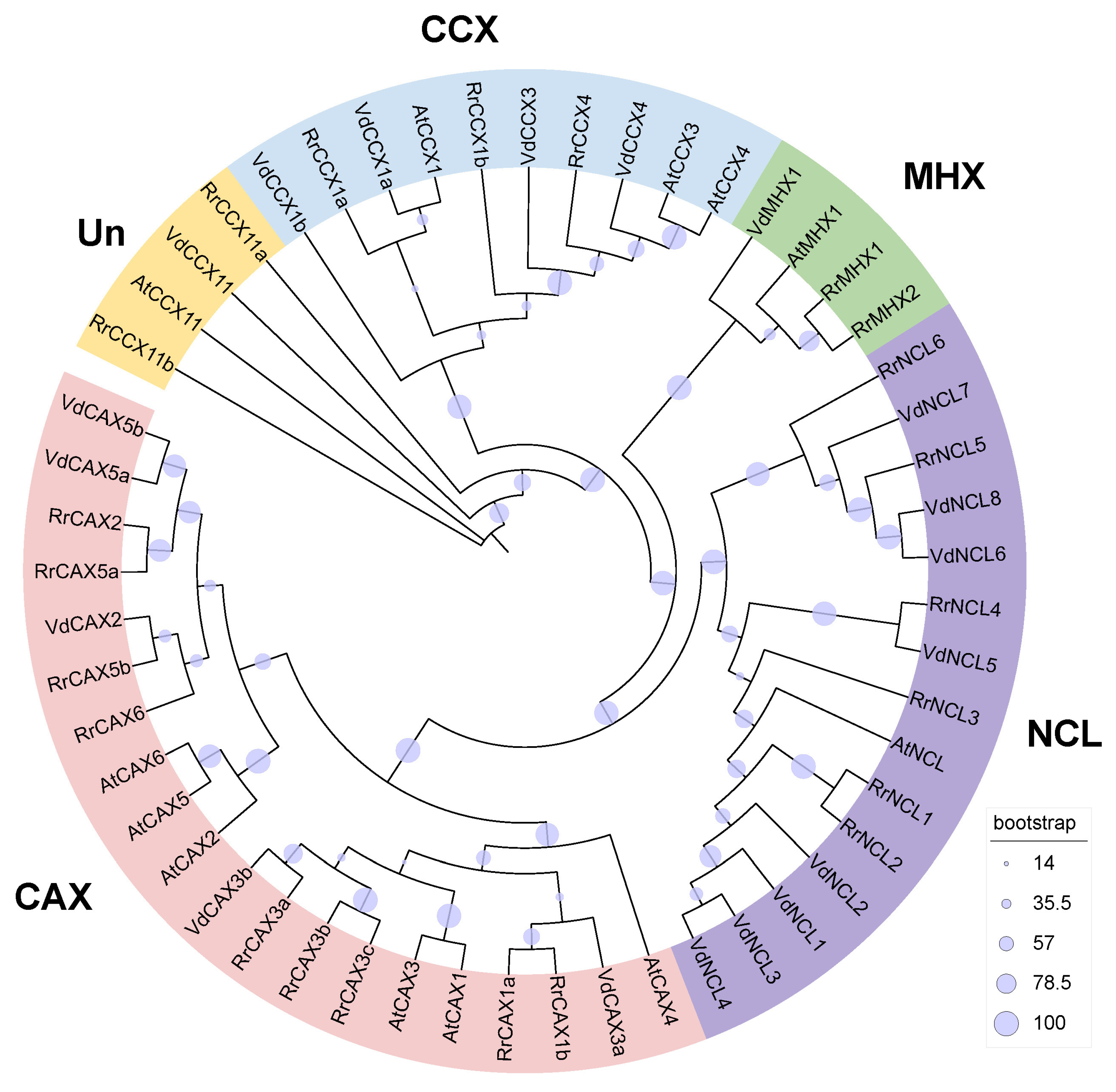
2.1. Phylogenetic Analysis of RrCaCAs
2.2. Chromosomal Location and Collinearity Analysis of RrCaCAs
2.3. Duplication, Synthesis, and Evolutionary Analysis of RrCaCA Gene Family
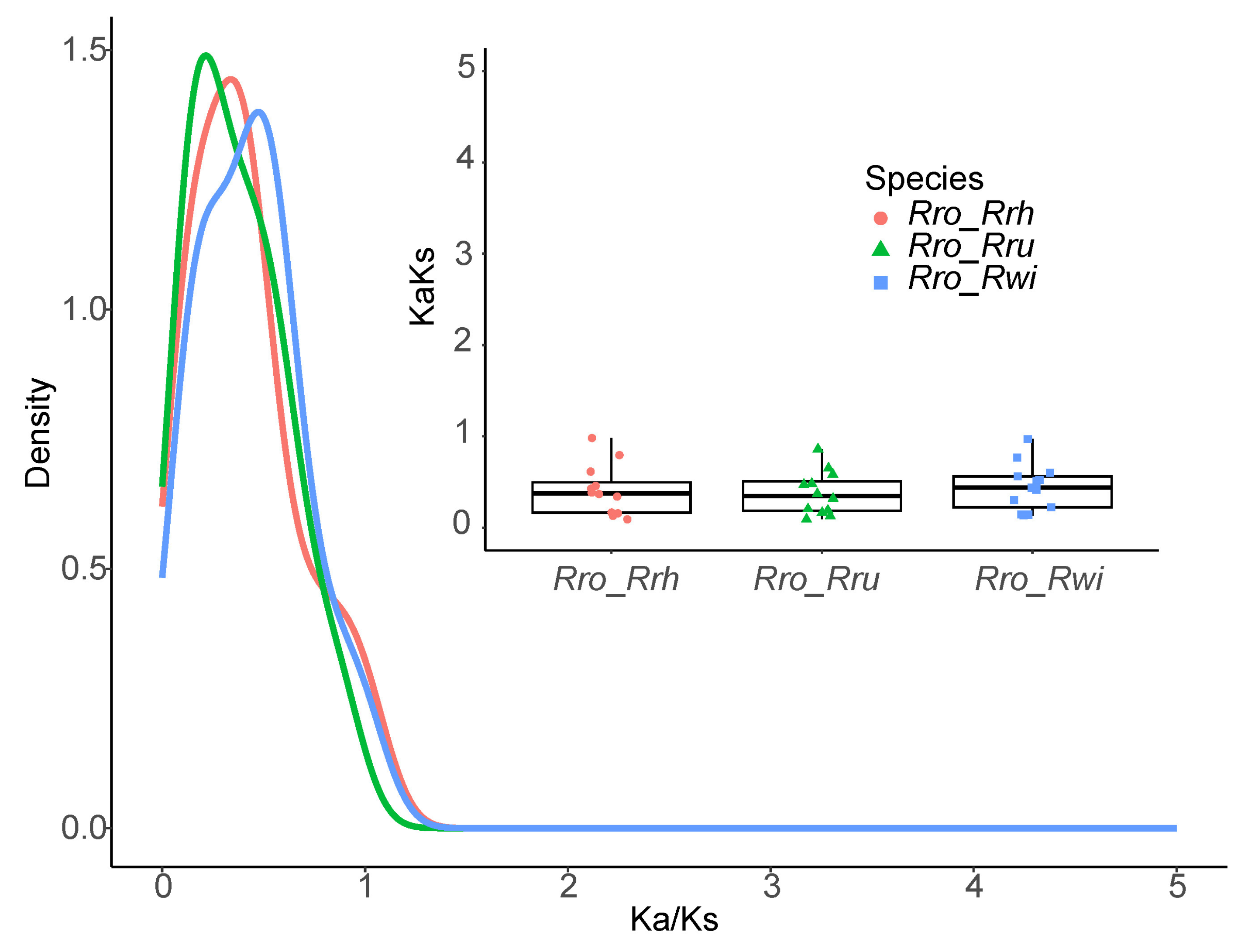
2.4. Analysis of Ka and Ks Substitution Patterns
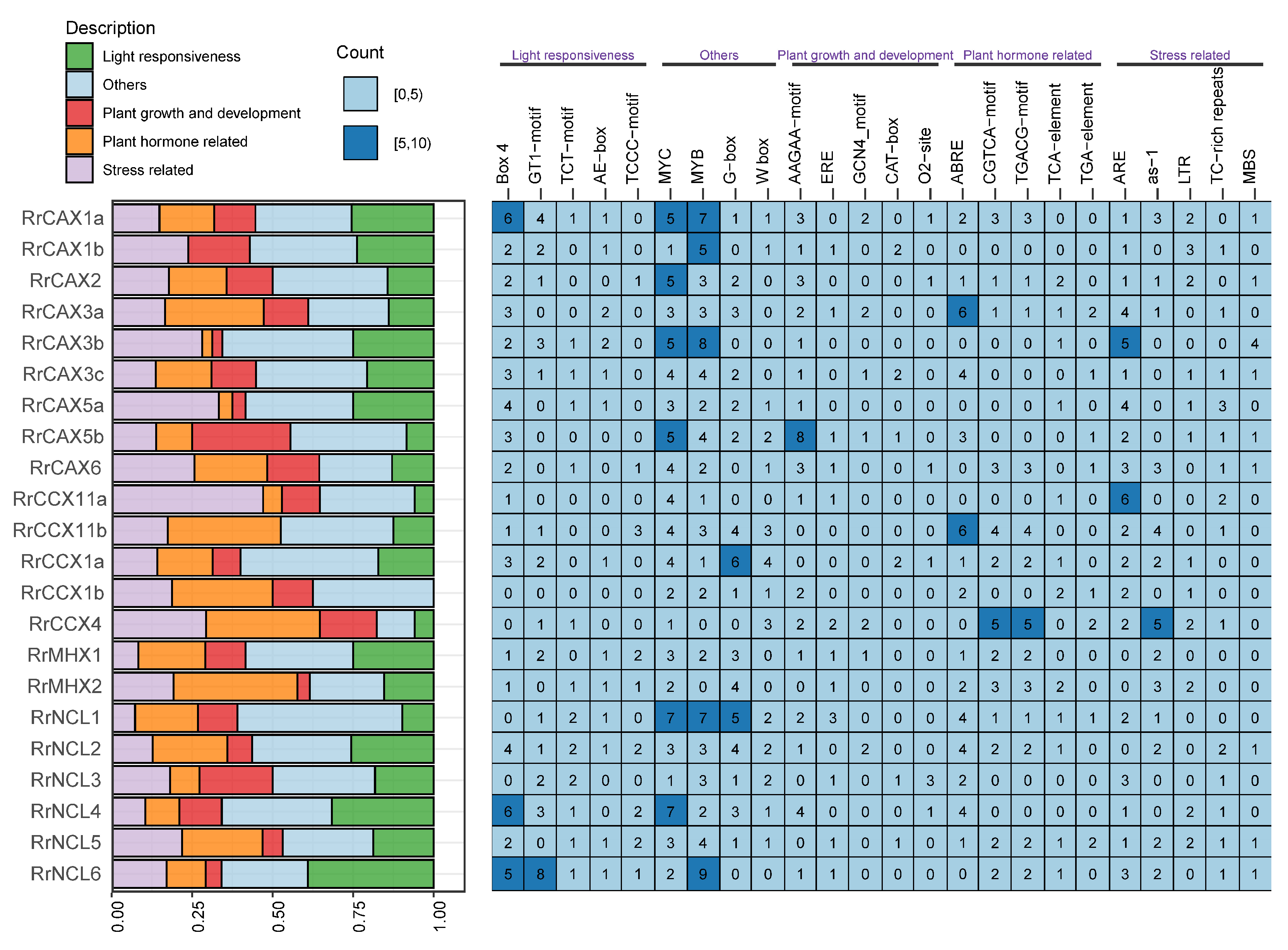
2.5. Identification of Cis-Acting Elements in Promoters of RrCaCAs
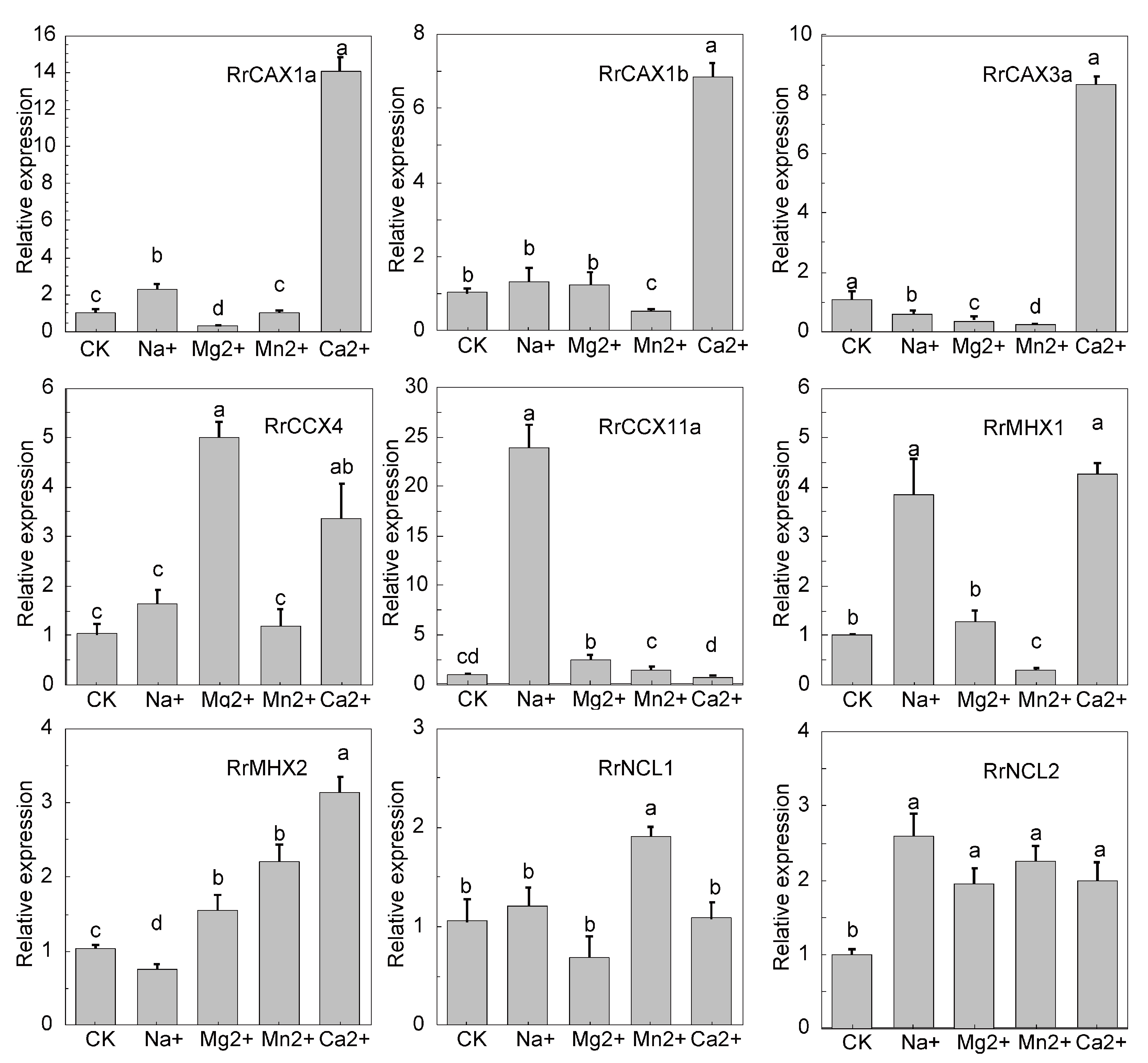
2.6. qRT-PCR Analysis of RrCaCAs Response to Heavy Metal Stress
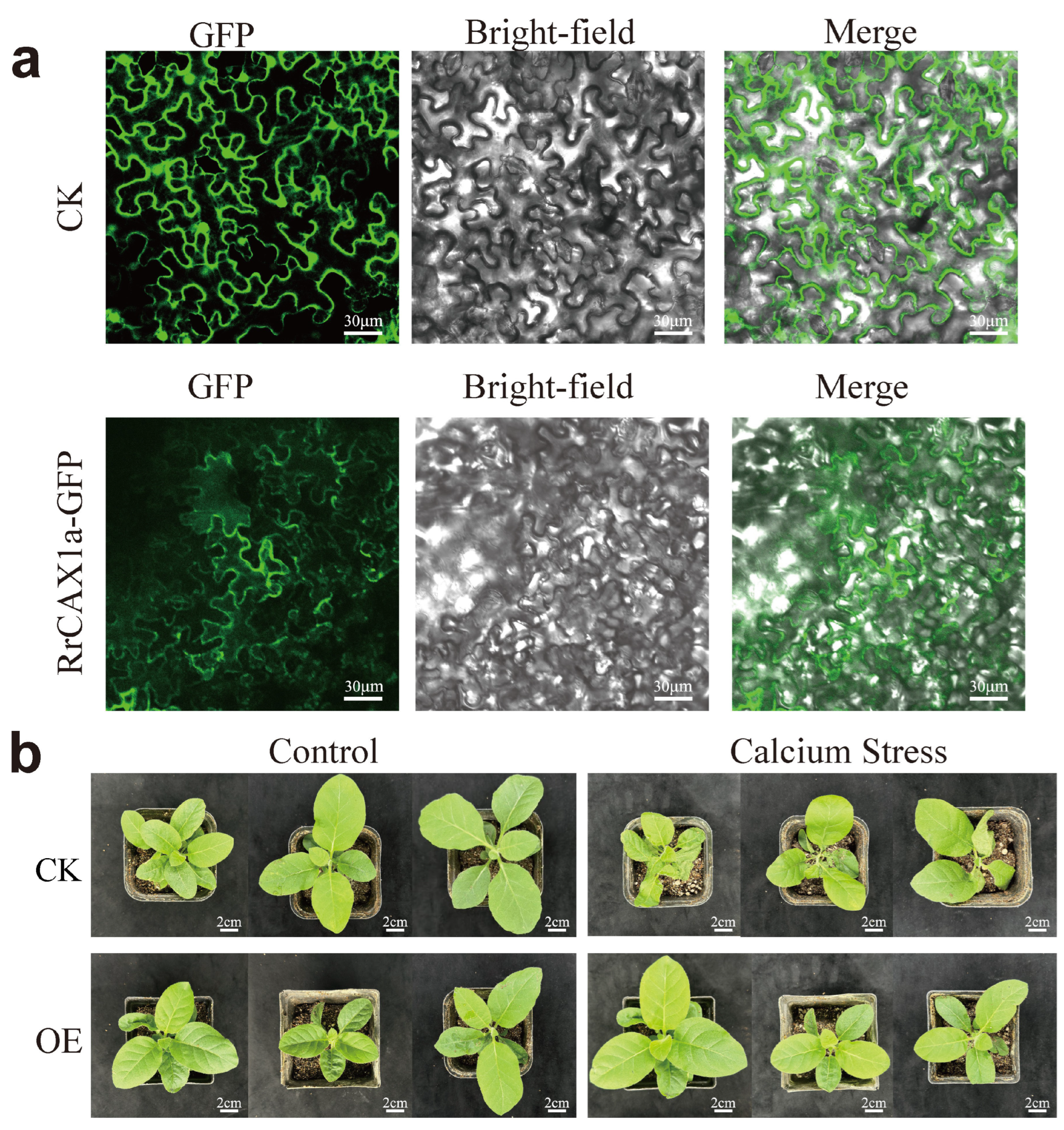
2.7. Subcellular Localization of RrCAX1a and Its Heterologous Expression Enhanced Tobacco Resistance to Calcium Stress
3. Discussion
3.1. RrCaCAs Structural Diversity and Evolutionary Insights
3.2. RrCaCA Regulatory Elements and Stress Responses
3.3. RrCaCA and RrCAX1a Mechanisms in R. roxburghii
3.4. Future Directions
4. Materials and Methods
4.1. Plant Materials
4.2. Identification and Phylogenetic Analysis of RrCaCA Gene Family in R. roxburghii
4.3. Chromosomal Localization, Gene Structure Analysis, and Motif Identification of RrCaCAs
4.4. Comparative Genomic Study of R. roxburghii and Other Rosa Species
4.5. Analysis of Cis-Regulatory Elements in Promoters of RrCaCA Genes
4.6. Stress Treatment of R. roxburghii
4.7. Validation of RrCaCAs with qRT-PCR
4.8. Subcellular Localization Analysis of RrCaCAX1a
4.9. The Genetic Transformation of Tobacco
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hirschi, K.D.; Zhen, R.G.; Cunningham, K.W.; Rea, P.A.; Fink, G.R. CAX1, an H+/Ca2+ antiporter from Arabidopsis. Proc. Natl. Acad. Sci. USA 1996, 93, 8782–8786. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.; Tian, H.; Park, S.; Sreevidya, C.S.; Ward, J.M.; Hirschi, K.D. AtCCX3 is an Arabidopsis endomembrane H+-dependent K+ transporter. Plant Physiol. 2008, 148, 1474–1486. [Google Scholar] [CrossRef]
- Wang, P.; Li, Z.; Wei, J.; Zhao, Z.; Sun, D.; Cui, S. A Na+/Ca2+ exchanger-like protein (AtNCL) involved in salt stress in Arabidopsis. J. Biol. Chem. 2012, 287, 44062–44070. [Google Scholar] [CrossRef]
- Pittman, J.K.; Hirschi, K.D. Phylogenetic analysis and protein structure modelling identifies distinct Ca2+/Cation antiporters and conservation of gene family structure within Arabidopsis and rice species. Rice 2016, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Taneja, M.; Tyagi, S.; Sharma, S.; Upadhyay, S. Ca2+/Cation antiporters (CaCA): Identification, characterization and expression profiling in bread wheat (Triticum aestivum L). Front. Plant Sci. 2016, 7, 1775. [Google Scholar] [CrossRef]
- Amagaya, K.; Shibuya, T.; Nishiyama, M.; Kato, K.; Kanayama, Y. Characterization and expression analysis of the Ca2+/cation antiporter gene family in tomatoes. Plants 2019, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Theerawitaya, C.; Chaum, S.; Kirdmanee, C.; Takabe, T. Expression and functional analysis of putative vacuolar Ca2+-transporters (CAXs and ACAs) in roots of salt tolerant and sensitive rice cultivars. Protoplasma 2014, 251, 1067–1075. [Google Scholar] [CrossRef]
- Cho, D.; Villieres, F.; Kroniewicz, L.; Lee, S.; Seo, Y.J.; Hirschi, K.D.; Leonhardt, N.; Kwak, J.M. Vacuolar CAX1 and CAX3 influence auxin transport in guard cells via regulation of apoplastic pH. Plant Physiol. 2012, 160, 1293–1302. [Google Scholar] [CrossRef]
- Li, P.; Zhang, G.; Gonzales, N.; Guo, Y.; Hu, H.; Park, S.; Zhao, J. Ca2+-regulated and diurnal rhythm-regulated Na+/Ca2+ exchanger AtNCL affects flowering time and auxin signalling in Arabidopsis. Plant Cell Environ. 2016, 39, 377–392. [Google Scholar] [CrossRef]
- Hocking, B.; Conn, S.J.; Manohar, M.; Xu, B.; Athman, A.; Stancombe, M.A.; Webb, A.R.; Hirschi, K.D.; Gilliham, M. Heterodimerization of Arabidopsis calcium/proton exchangers contributes to regulation of guard cell dynamics and plant defense responses. J. Exp. Bot. 2017, 68, 4171–4183. [Google Scholar] [CrossRef]
- Morris, J.; Hawthorne, K.; Hotze, T.; Abrams, S.; Hirschi, K.D. Nutritional impact of elevated calcium transport activity in carrots. Proc. Natl. Acad. Sci. USA 2008, 105, 431–1435. [Google Scholar] [CrossRef] [PubMed]
- Navarro-León, E.; Ruiz, J.; Graham, N.; Blasco, B. Physiological profile of CAX1a TILLING mutants of Brassica rapa exposed to different calcium doses. Plant Sci. 2018, 272, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Elless, M.P.; Park, J.; Jenkins, A.; Lim, W.; Edgar, C.I.V.; Hirschi, K.D. Sensory analysis of calcium-biofortified lettuce. Plant Biotechnol. J. 2009, 7, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Navarro-León, E.; Ruiz, J.M.; Albacete, A.; Blasco, B. Effect of CAX1a TILLING mutations and calcium concentration on some primary metabolism processes in Brassica rapa plants. J. Plant Physiol. 2019, 237, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Qiao, K.; Wang, F.; Liang, S.; Hu, Z.; Chai, T. Heterologous expression of TuCAX1a and TuCAX1b enhances Ca2+ and Zn2+ translocation in Arabidopsis. Plant Cell Rep. 2019, 38, 597–607. [Google Scholar] [CrossRef]
- Ahmadi, H.; Corso, M.; Weber, M.; Verbruggen, N.; Clemens, S. CAX1 suppresses Cd-induced generation of reactive oxygen species in Arabidopsis halleri. Plant Cell Environ. 2018, 41, 2435–2448. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhang, B.; Fu, X.; Huang, Q.; Li, C.; Liu, G.; Hai Liu, R. Recent advances in poly- saccharides from Rose roxburghii Tratt fruits: Isolation, structural character- ization, and bioactivities. Food Funct. 2022, 13, 12561–12571. [Google Scholar] [CrossRef]
- Wang, L.; Wei, T.; Zheng, L.; Jiang, F.; Ma, W.; Lu, M.; Wu, X.; An, H. Recent advances on main active ingredients, pharmacological activities of Rosa roxbughii and its development and utilization. Foods 2023, 12, 1051. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Lu, M.; Rao, T.; Liu, Z.; Wu, X.; An, H. Comparative analysis of Fruit Metabo- lome using widely targeted Metabolomics reveals nutritional characteristics of different Rosa roxburghii genotypes. Foods 2022, 11, 850. [Google Scholar] [CrossRef]
- Van der Westhuizen, F.H.; van Rensburg, C.S.J.; Rautenbach, G.S.; Marnewick, J.L.; Loots, D.T.; Huysamen, C.; Louw, R.; Pretorius, P.J.; Erasmus, E. In vitro antioxidant, antimutagenic and genoprotective activity of Rosa roxburghii fruit extract. Phytother. Res. 2008, 22, 376–383. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, B.; Xiao, J.; Huang, Q.; Li, C.; Fu, X. Physicochemical, functional, and biological properties of water-soluble polysaccharides from Rosa roxburghii Tratt fruit. Food Chem. 2018, 249, 127–135. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, P.; Li, C.; Xu, F.; Chen, J. A polysaccharide from Rosa roxburghii Tratt fruit attenuates high-fat diet-induced intestinal barrier dysfunction and inflammation in mice by modulating the gut microbiota. Food Funct. 2022, 13, 530–547. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.T.; Lv, M.J.; An, J.Y.; Fan, X.H.; Dong, M.Z.; Zhang, S.D.; Wang, J.D.; Wang, Y.Q.; Cai, Z.H.; Fu, Y.J. Botanical characteristics, phytochemistry and related biological activities of Rosa roxburghii Tratt fruit, and its potential use in functional foods: A review. Food Funct. 2021, 12, 1432–1451. [Google Scholar] [CrossRef]
- Meng, Q.-J.; Fan, W.-G. Calcium-tolerance type and adaptability to high-calcium habitats of Rosa roxburghii. Chin. J. Plant Ecol. 2022, 46, 1562. [Google Scholar] [CrossRef]
- Li, L.L.; An, H.M. Effects of Ca2+ and Cu2+ on the expression of genes related to AsA metabolism in Rosa roxburghii. Fruits. J. Hortic. 2016, 43, 1377–1382. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, M.; An, H.M.; Huang, W.; Liu, W. Effects of exogenous divalent cations Ca2+, Mg2+, and Cu2+ and acriflavine on ascorbate biosynthesis in Rosa roxburghii fruits. Chin. Agric. Sci. 2012, 45, 1144–1149. [Google Scholar] [CrossRef]
- Wang, Z.; Lu, M.; An, H. Transcriptome analysis reveals candidate genes involved in calcium absorption of Rosa roxburghii plants and their effects on the bioactive substance accumulation in fruit. J. Soil. Sci. Plant Nutr. 2024, 24, 732–748. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon–intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. TRENDS Genet. 2002, 18, 486–487. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Tang, R.-J.; Kou, S.; Xu, X.; Lu, Y.; Rauscher, K.; Voelker, A.; Luan, S. Mechanisms of calcium homeostasis orchestrate plant growth and immunity. Nature 2024, 627, 382–388. [Google Scholar] [CrossRef]
- Lv, W.; Zhu, L.; Tan, L.; Gu, L.; Wang, H.; Du, X.; Zhu, B.; Zeng, T.; Wang, C. Genome-wide identification and analysis of the GST gene family in wild blueberry (Vaccinium duclouxii) and their impact on anthocyanin accumulation. Plants 2024, 13, 1497. [Google Scholar] [CrossRef] [PubMed]
- Magadum, S.; Banerjee, U.; Murugan, P.; Gangapur, D.; Ravikesavan, R. Gene duplication as a major force in evolution. J. Genet. 2013, 92, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Flagel, L.E.; Wendel, J.F. Gene duplication and evolutionary novelty in plants. New Phytol. 2009, 183, 557–564. [Google Scholar] [CrossRef]
- Taneja, M.; Upadhyay, S.K. An introduction to the calcium transport elements in plants. In Calcium Transport Elements in Plants; Academic Press: Cambridge, MA, USA, 2021; pp. 1–18. [Google Scholar]
- Frei dit Frey, N.; Mbengue, M.; Kwaaitaal, M.; Nitsch, L.; Altenbach, D.; Häweker, H.; Lozano-Duran, R.; Njo, M.F.; Beeckman, T.; Huettel, B. Plasma membrane calcium ATPases are important components of receptor-mediated signaling in plant immune responses and development. Plant Physiol. 2012, 159, 798–809. [Google Scholar] [CrossRef]
- Pittman, J.; Hirschi, K. CAX control: Multiple roles of vacuolar cation/H+ exchangers in metal tolerance, mineral nutrition and environmental signalling. Plant Biol. 2024, 26, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Conn, S.J.; Gilliham, M.; Athman, A.; Schreiber, A.W.; Baumann, U.; Moller, I.; Cheng, N.-H.; Stancombe, M.A.; Hirschi, K.D.; Webb, A.A. Cell-specific vacuolar calcium storage mediated by CAX1 regulates apoplastic calcium concentration, gas exchange, and plant productivity in Arabidopsis. The Plant Cell 2011, 23, 240–257. [Google Scholar] [CrossRef]
- Wdowiak, A.; Kryzheuskaya, K.; Podgórska, A.; Paterczyk, B.; Zebrowski, J.; Archacki, R.; Szal, B. Ammonium nutrition modifies cellular calcium distribution influencing ammonium-induced growth inhibition. J. Plant Physiol. 2024, 298, 154264. [Google Scholar] [CrossRef]
- Han, B.; Tai, Y.; Li, S.; Shi, J.; Wu, X.; Kakeshpour, T.; Weng, J.; Cheng, X.; Park, S.; Wu, Q. Redefining the N-terminal regulatory region of the Ca2+/H+ antiporter CAX1 in tomato. Front. Plant Sci. 2022, 13, 938839. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Yan, H.; Yi, X.; Pan, Q.; Liu, Y.; Zhang, M.; Li, J.; Xiao, Q. The chromosome-level genome and functional database accelerate research about biosynthesis of secondary metabolites in Rosa roxburghii. BMC Plant Biol. 2024, 24, 410. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; He, Z.J.; He, J.F.; Lv, W.; Huang, S.X.; Li, J.W.; Zhu, L.Y.; Wan, S.; Zhou, W.F.; Yang, Z.S.; et al. The Telomere-to-telomere gap-free reference genome of wild blueberry (Vaccinium duclouxii) provides its high soluble sugar and anthocyanin accumulation. Hortic. Res. 2023, 10, uhad209. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res 2018, 47, D427–D432. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Hibrand Saint-Oyant, L.; Ruttink, T.; Hamama, L.; Kirov, I.; Lakhwani, D.; Zhou, N.-N.; Bourke, P.; Daccord, N.; Leus, L.; Schulz, D. A high-quality genome sequence of Rosa chinensis to elucidate ornamental traits. Nat. Plants 2018, 4, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Su, L.; Hu, S.; Xue, J.-Y.; Liu, H.; Liu, G.; Jiang, Y.; Du, J.; Qiao, Y.; Fan, Y. A chromosome-level genome assembly of rugged rose (Rosa rugosa) provides insights into its evolution, ecology, and floral characteristics. Hortic. Res. 2021, 8, 141. [Google Scholar] [CrossRef]
- Zhong, M.-C.; Jiang, X.-D.; Yang, G.-Q.; Cui, W.-H.; Suo, Z.-Q.; Wang, W.-J.; Sun, Y.-B.; Wang, D.; Cheng, X.-C.; Li, X.-M. Rose without prickle: Genomic insights linked to moisture adaptation. Natl. Sci. Rev. 2021, 8, nwab092. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Zeng, T.; Li, J.W.; Xu, Z.Z.; Zhou, L.; Li, J.J.; Yu, Q.; Luo, J.; Chan, Z.L.; Jongsma, M.A.; Hu, H.; et al. TcMYC2 regulates pyrethrin biosynthesis in Tanacetum cinerariifolium. Hortic. Res. 2022, 9, uhac178. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 408, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Sun, T.; Pan, Q.; Anupol, N.; Chen, H.; Shi, J.; Liu, F.; Deqiang, D.; Wang, C.; Zhao, J. Rr MYB 5-and Rr MYB 10-regulated flavonoid biosynthesis plays a pivotal role in feedback loop responding to wounding and oxidation in Rosa rugosa. Plant Biotechnol. J. 2019, 17, 2078–2095. [Google Scholar] [CrossRef]
- Zeng, T.; Yu, Q.; Shang, J.; Xu, Z.; Zhou, L.; Li, W.; Li, J.; Hu, H.; Zhu, L.; Li, J.; et al. TcbHLH14 a jasmonate associated MYC2-like transcription factor positively regulates pyrethrin biosynthesis in Tanacetum cinerariifolium. Int. J. Mol. Sci. 2023, 24, 7379. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W.; Xu, Z.Z.; Zeng, T.; Zhou, L.; Li, J.J.; Hu, H.; Luo, J.; Wang, C.Y. Overexpression of TcCHS increases pyrethrin content when using a genotype-independent transformation system in pyrethrum (Tanacetum cinerariifolium). Plants 2022, 11, 1575. [Google Scholar] [CrossRef]
- Li, J.W.; Zeng, T.; Xu, Z.Z.; Zhou, L.; Shi, A.Q.; Luo, Y.Y.; Zhu, L.Y.; Wang, Y.Y.; Luo, J.; Wang, C.Y. TcWRKY75 participates in pyrethrin biosynthesis by positively regulating the expression of TcCHS, TcAOC, and TcGLIP in Tanacetum cinerariifolium. Ind. Crops Prod. 2023, 202, 117062. [Google Scholar] [CrossRef]
- Ma, L.; Jia, W.; Duan, Q.; Du, W.; Li, X.; Cui, G.; Wang, X.; Wang, J. Heterologous expression of platycodon grandiflorus PgF3′5′H. modifies flower color pigmentation in tobacco. Genes. 2023, 14, 1920. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, T.; Zhu, L.; Su, W.; Gu, L.; Wang, H.; Du, X.; Zhu, B.; Wang, C.; Wu, D. Comparative Analysis of Ca2+/Cation Antiporter Gene Family in Rosa roxburghii and Enhanced Calcium Stress Tolerance via Heterologous Expression of RrCAX1a in Tobacco. Plants 2024, 13, 3582. https://doi.org/10.3390/plants13243582
Zeng T, Zhu L, Su W, Gu L, Wang H, Du X, Zhu B, Wang C, Wu D. Comparative Analysis of Ca2+/Cation Antiporter Gene Family in Rosa roxburghii and Enhanced Calcium Stress Tolerance via Heterologous Expression of RrCAX1a in Tobacco. Plants. 2024; 13(24):3582. https://doi.org/10.3390/plants13243582
Chicago/Turabian StyleZeng, Tuo, Liyong Zhu, Wenwen Su, Lei Gu, Hongcheng Wang, Xuye Du, Bin Zhu, Caiyun Wang, and Di Wu. 2024. "Comparative Analysis of Ca2+/Cation Antiporter Gene Family in Rosa roxburghii and Enhanced Calcium Stress Tolerance via Heterologous Expression of RrCAX1a in Tobacco" Plants 13, no. 24: 3582. https://doi.org/10.3390/plants13243582
APA StyleZeng, T., Zhu, L., Su, W., Gu, L., Wang, H., Du, X., Zhu, B., Wang, C., & Wu, D. (2024). Comparative Analysis of Ca2+/Cation Antiporter Gene Family in Rosa roxburghii and Enhanced Calcium Stress Tolerance via Heterologous Expression of RrCAX1a in Tobacco. Plants, 13(24), 3582. https://doi.org/10.3390/plants13243582