
Floristic Richness in a Mediterranean Hotspot: A Journey across Italy

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
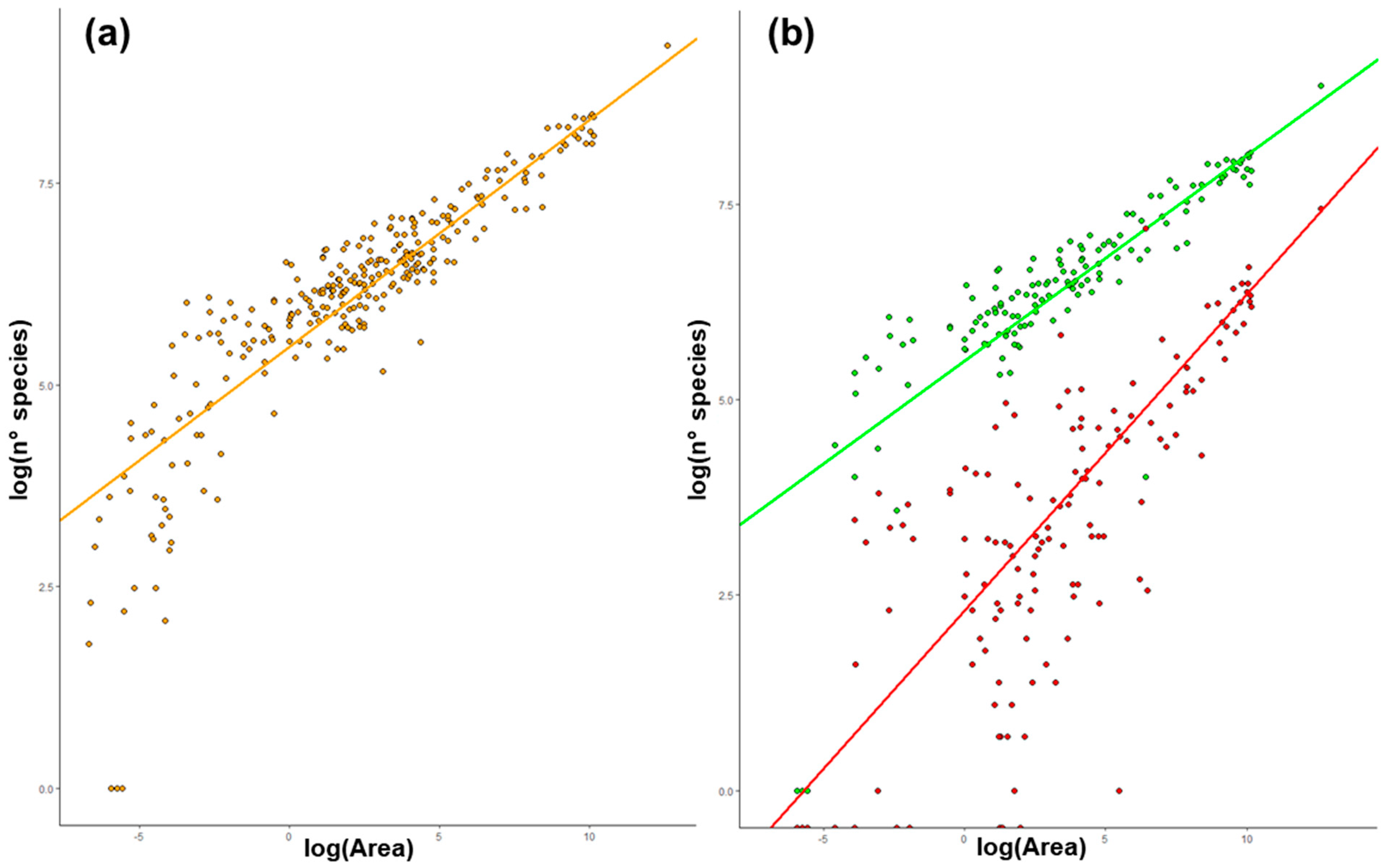
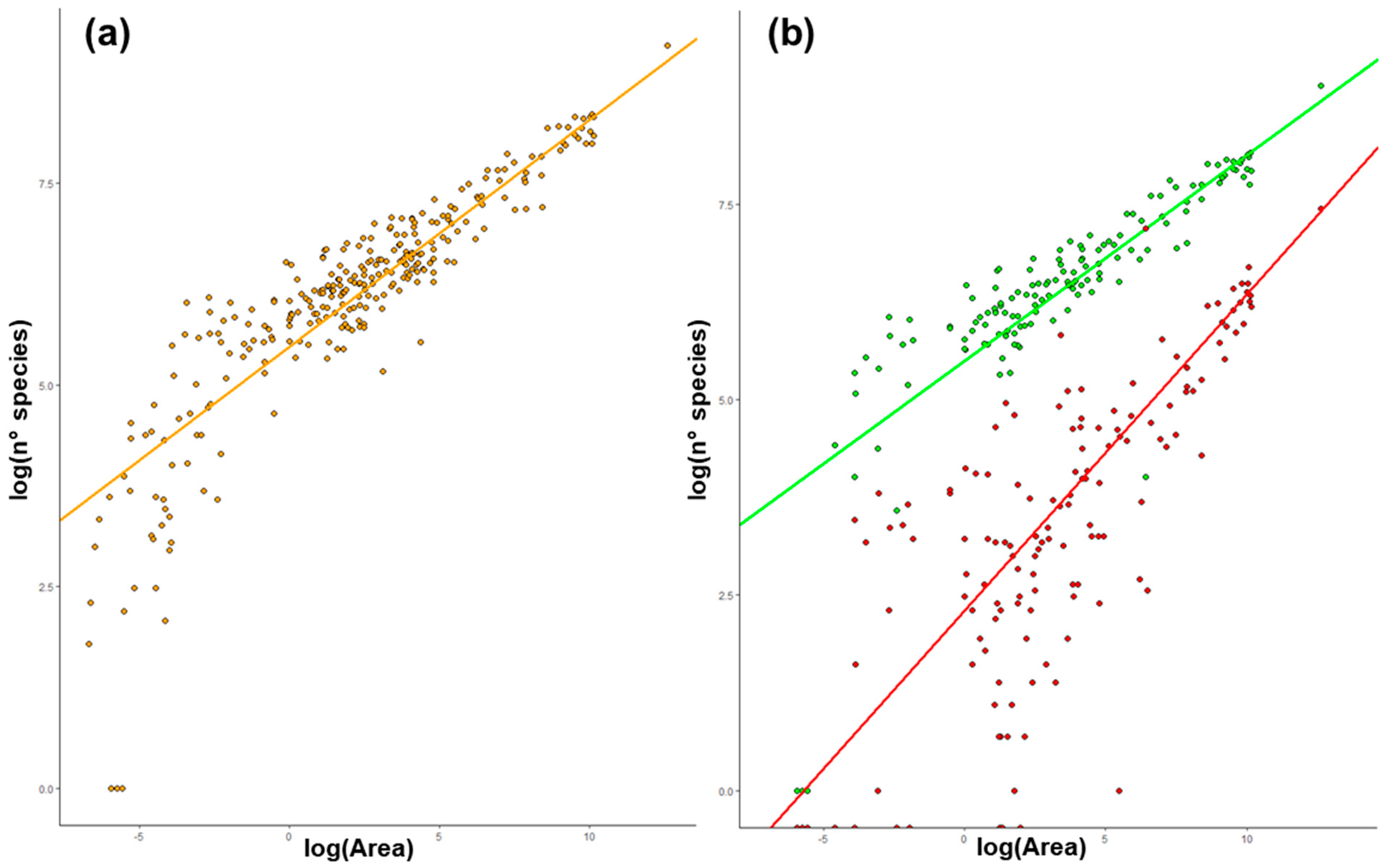
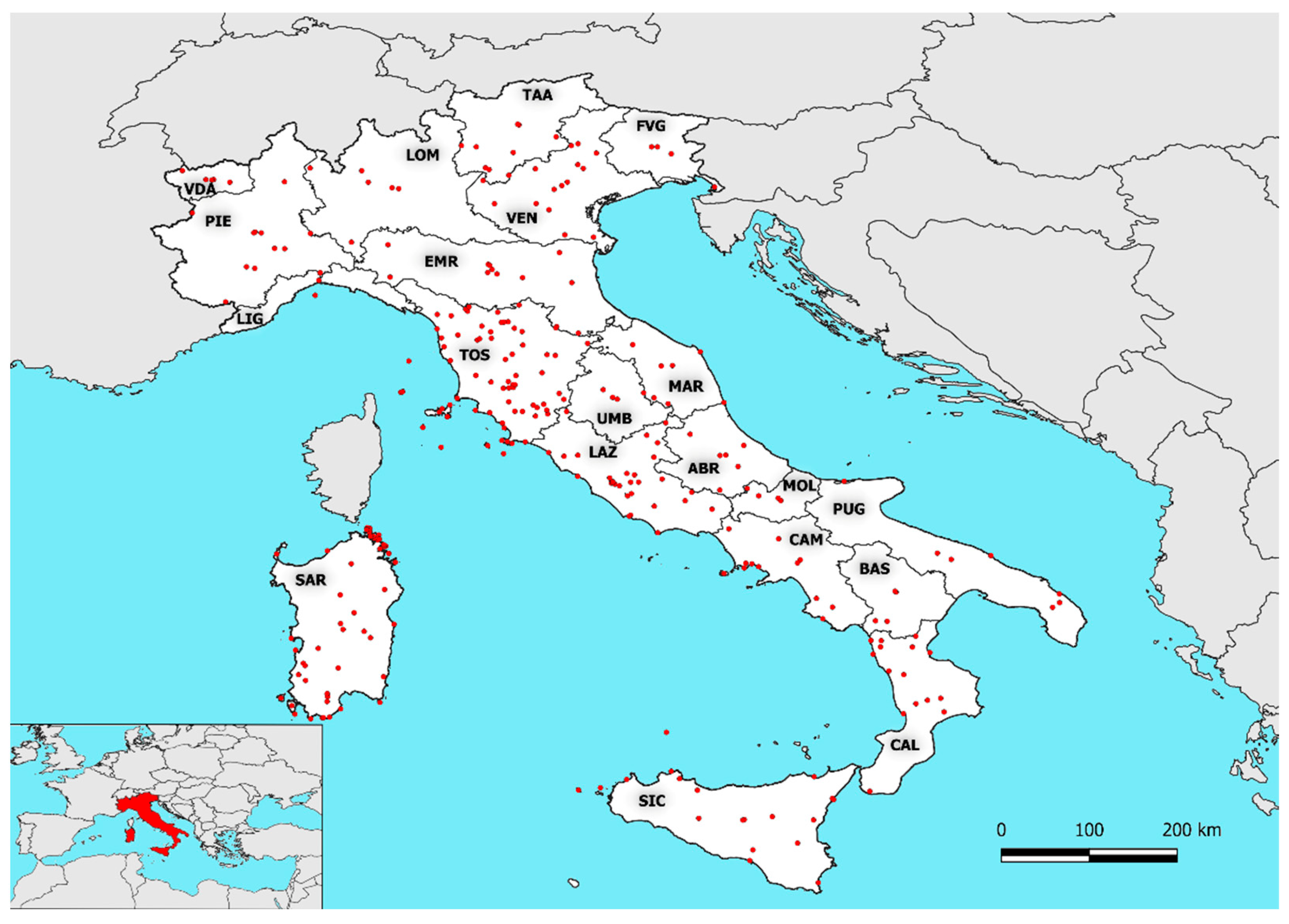
2.1. Species–Area Relationship (SAR) in Italy
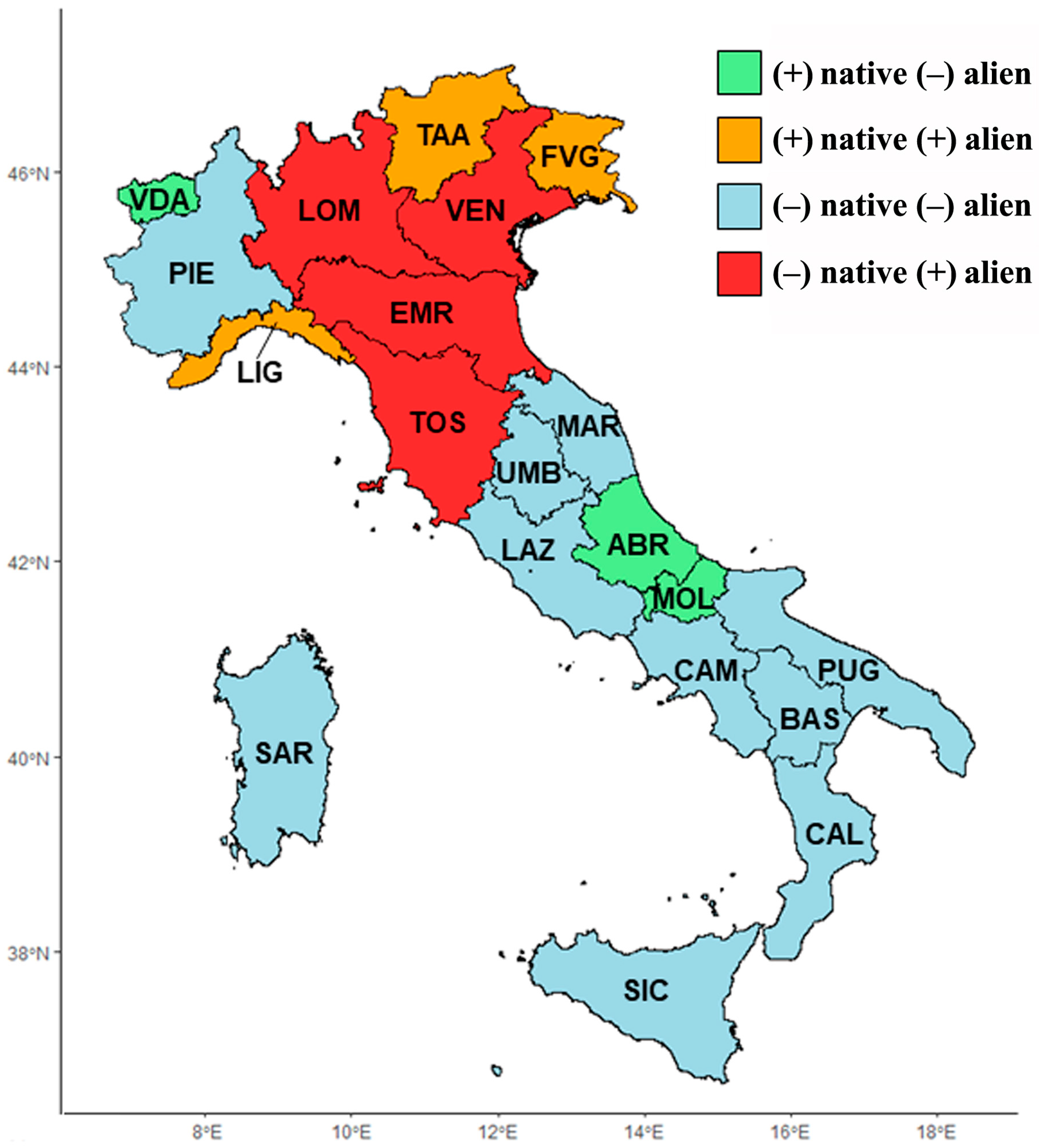
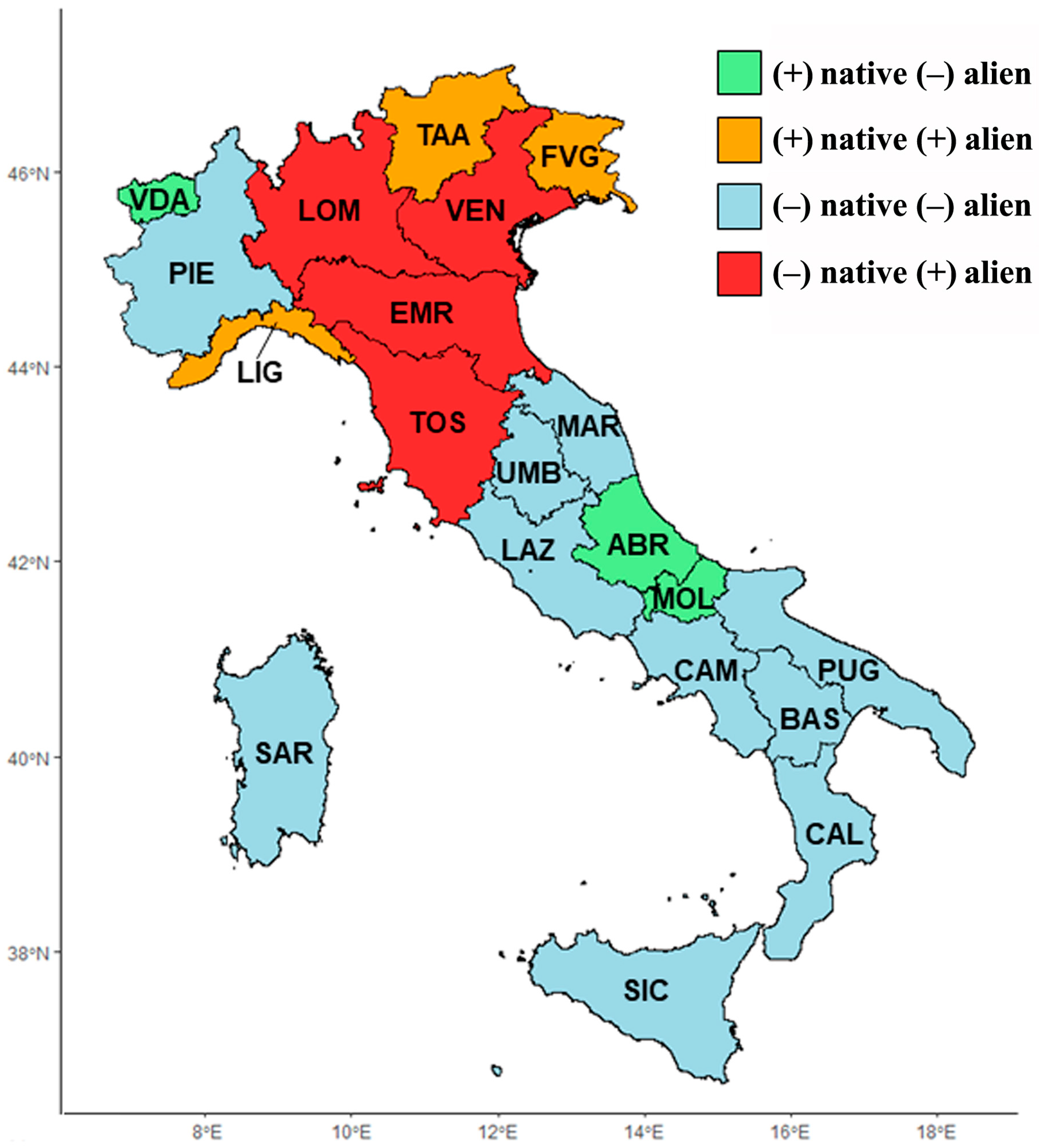
2.2. Floristic Richness Comparison among Italian Regions
3. Discussion
3.1. Species Area–Relationship (SAR) in Italy
3.2. Floristic Richness Comparison among Italian Regions
4. Materials and Methods
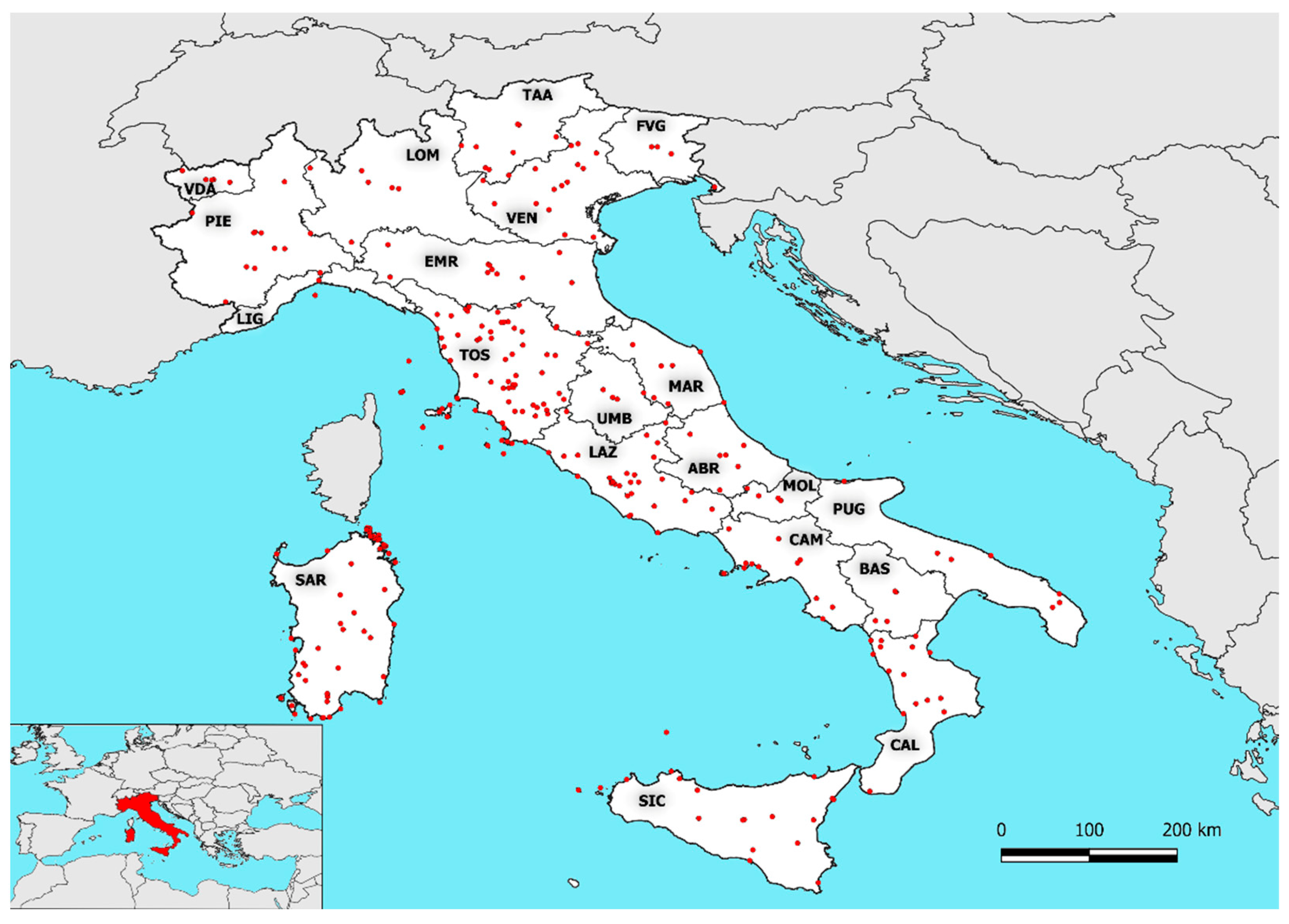
4.1. Study Area and Floristic Dataset
4.2. Species–Area Relationship (SAR)
4.3. Floristic Richness Comparison among Italian Regions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sabatini, F.M.; Jiménez-Alfaro, B.; Jandt, U.; Chytrý, M.; Field, R.; Kessler, M.; Lenoir, J.; Schrodt, F.; Wiser, S.K.; Arfin Khan, M.A.S.; et al. Global patterns of vascular plant alpha diversity. Nat. Commun. 2022, 13, 4683. [Google Scholar] [CrossRef] [PubMed]
- D’Antraccoli, M.; Roma-Marzio, F.; Carta, A.; Landi, S.; Bedini, G.; Chiarucci, A.; Peruzzi, L. Drivers of floristic richness in the Mediterranean: A case study from Tuscany. Biodiv. Cons. 2019, 28, 1411–1429. [Google Scholar] [CrossRef]
- Cowling, R.M.; Rundel, P.W.; Lamont, B.B.; Kalin Arroyo, M.; Arianoutsou, M. Plant diversity in Mediterranean-climate regions. Trends Ecol. Evol. 1996, 11, 362–366. [Google Scholar] [CrossRef]
- Thompson, J.D.; Lavergne, S.; Affre, L.; Gaudeul, M.; Debussche, M. Ecological differentiation of Mediterranean endemic plants. Taxon 2005, 54, 967–976. [Google Scholar] [CrossRef]
- D’Antraccoli, M.; Carta, A.; Astuti, G.; Franzoni, J.; Giacò, A.; Tiburtini, M.; Pinzani, L.; Peruzzi, L. A comprehensive approach to improving endemic plant species research, conservation, and popularization. J. Zool. Bot. Gard. 2023, 4, 490–506. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Peruzzi, L.; Conti, F.; Bartolucci, F. An inventory of vascular plants endemic to Italy. Phytotaxa 2014, 168, 1–75. [Google Scholar] [CrossRef]
- Brundu, G.; Peruzzi, L.; Domina, G.; Bartolucci, F.; Galasso, G.; Peccenini, S.; Raimondo, F.M.; Albano, A.; Alessandrini, A.; Banfi, E.; et al. At the intersection of cultural and natural heritage: Distribution and conservation of the type localities of the Italian endemic vascular plants. Biol. Cons. 2017, 214, 109–118. [Google Scholar] [CrossRef]
- Médail, F.; Quézel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Peruzzi, L. Floristic inventories and collaborative approaches: A new era for checklists and floras? Plant Biosyst. 2018, 152, 177–178. [Google Scholar] [CrossRef]
- D’Antraccoli, M.; Bedini, G.; Peruzzi, L. Next Generation Floristics: A workflow to integrate novel methods in traditional floristic research. Plant Biosyst. 2022, 156, 594–597. [Google Scholar] [CrossRef]
- D’Antraccoli, M.; Bacaro, G.; Tordoni, E.; Bedini, G.; Peruzzi, L. More species, less effort: Designing and comparing sampling strategies to draft optimised floristic inventories. Persp. Plant Ecol. Evol. Syst. 2020, 45, 125547. [Google Scholar] [CrossRef]
- Tjørve, E. Shapes and functions of Species–Area curves: A review of possible models. J. Biogeogr. 2003, 30, 827–835. [Google Scholar] [CrossRef]
- Dengler, J. Which function describes the Species–Area Relationship best? A review and empirical evaluation. J. Biogeogr. 2009, 36, 728–744. [Google Scholar] [CrossRef]
- Lomolino, M.V. The Species-Area Relationship: New challenges for an old pattern. Prog. Phys. Geogr. Earth Environ. 2001, 25, 1–21. [Google Scholar] [CrossRef]
- Arrhenius, O. Species and Area. J. Ecol. 1921, 9, 95–99. [Google Scholar] [CrossRef]
- Gleason, H.A. On the relation between species and area. Ecology 1922, 3, 158–162. [Google Scholar] [CrossRef]
- Iliadou, E.; Kallimanis, A.S.; Dimopoulos, P.; Panitsa, M. Comparing the two Greek archipelagos plant species diversity and endemism patterns highlight the importance of isolation and precipitation as biodiversity drivers. J. Biol. Res. 2014, 21, 16. [Google Scholar] [CrossRef]
- Chiarucci, A.; Fattorini, S.; Foggi, B.; Landi, S.; Lazzaro, L.; Podani, J.; Simberloff, D. Plant recording across two Centuries reveals dramatic changes in species diversity of a Mediterranean archipelago. Sci. Rep. 2017, 7, 5415. [Google Scholar] [CrossRef]
- Chiarucci, A.; Guarino, R.; Pasta, S.; Rosa, A.L.; Cascio, P.L.; Médail, F.; Pavon, D.; Fernández-Palacios, J.M.; Zannini, P. Species–Area Relationship and small-island effect of vascular plant diversity in a young volcanic archipelago. J. Biogeogr. 2021, 48, 2919–2931. [Google Scholar] [CrossRef]
- Testolin, R.; Attorre, F.; Bruzzaniti, V.; Guarino, R.; Jiménez-Alfaro, B.; Lussu, M.; Martellos, S.; Di Musciano, M.; Pasta, S.; Sabatini, F.M.; et al. Plant species richness hotspots and related drivers across spatial scales in small Mediterranean islands. J. Syst. Evol. 2023. [Google Scholar] [CrossRef]
- Panitsa, M.; Tzanoudakis, D.; Triantis, K.A.; Sfenthourakis, S. Patterns of species richness on very small islands: The plants of the Aegean Archipelago. J. Biogeogr. 2006, 33, 1223–1234. [Google Scholar] [CrossRef]
- Raus, T.; Karadimou, E.; Dimopoulos, P. Taxonomic and functional plant diversity of the Santorini-Christiana island group (Aegean Sea, Greece). Willdenowia 2019, 49, 363–381. [Google Scholar] [CrossRef]
- Bartolucci, F.; Galasso, G.; Peruzzi, L.; Conti, F. Report 2021 on plant biodiversity in Italy: Native and alien vascular flora. Nat. Hist. Sci. 2023, 10, 41–50. [Google Scholar] [CrossRef]
- Bonari, G.; Fiaschi, T.; Fanfarillo, E.; Roma-Marzio, F.; Sarmati, S.; Banfi, E.; Biagioli, M.; Zerbe, S.; Angiolini, C. Remnants of naturalness in a reclaimed land of central Italy. Ital. Bot. 2021, 11, 9–30. [Google Scholar] [CrossRef]
- Peruzzi, L. The vascular flora of Empoli (Tuscany, Central Italy). Ital. Bot. 2023, 15, 21–33. [Google Scholar] [CrossRef]
- Sólymos, P.; Lele, S.R. Global pattern and local variation in Species–Area Relationships. Glob. Ecol. Biogeogr. 2012, 21, 109–120. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the Twenty-First Century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef]
- Chiarucci, A.; Bacaro, G.; Filibeck, G.; Landi, S.; Maccherini, S.; Scoppola, A. Scale dependence of plant species richness in a network of protected areas. Biodiv. Cons. 2012, 21, 503–516. [Google Scholar] [CrossRef]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The world records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995; ISBN 978-0-521-49952-1. [Google Scholar]
- Crawley, M.J.; Harral, J.E. Scale dependence in plant biodiversity. Science 2001, 291, 864–868. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; von Holle, B. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Stohlgren, T.J.; Barnett, D.T.; Karts’, J.T. The rich get richer: Patterns of plant invasions in the United States. Front. Ecol. Environ. 2003, 1, 11–14. [Google Scholar] [CrossRef]
- Pierini, B.; Garbari, F.; Peruzzi, L. Flora vascolare del Monte Pisano (Toscana nord-occidentale). Inform. Bot. Ital. 2009, 41, 147–213. [Google Scholar]
- Stinca, A.; Musarella, C.M.; Rosati, L.; Laface, V.L.; Licht, W.; Fanfarillo, E.; Wagensommer, R.P.; Galasso, G.; Fascetti, S.; Esposito, A.; et al. Italian vascular flora: New findings, updates and exploration of floristic similarities between regions. Diversity 2021, 13, 600. [Google Scholar] [CrossRef]
- Roma-Marzio, F.; Bedini, G.; Müller, J.V.; Peruzzi, L. A critical checklist of the woody flora of Tuscany (Italy). Phytotaxa 2016, 287, 1–135. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. Taxon richness and climate in angiosperms: Is there a globally consistent relationship that precludes region effects? Am. Nat. 2004, 163, 773–779; discussion 780–785. [Google Scholar] [CrossRef]
- Gheyret, G.; Guo, Y.; Fang, J.; Tang, Z. Latitudinal and elevational patterns of phylogenetic structure in forest communities in China’s mountains. Sci. China Life Sci. 2020, 63, 1895–1904. [Google Scholar] [CrossRef]
- Harrison, S.; Spasojevic, M.J.; Li, D. Climate and plant community diversity in space and time. Proc. Natl. Acad. Sci. USA 2020, 117, 4464–4470. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.W. The peninsular effect in Baja California: An entomological assessment. J. Biogeogr. 1987, 14, 359–365. [Google Scholar] [CrossRef]
- Olivier, P.I.; Rolo, V.; van Aarde, R.J. Pattern or process? Evaluating the peninsula effect as a determinant of species richness in coastal dune forests. PLoS ONE 2017, 12, e0173694. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.-S.; Stevens, R.D.; Baccus, J.T. Peninsula effect and species richness gradient in terrestrial mammals on the Korean Peninsula and other peninsulas. Mam. Rev. 2017, 47, 266–276. [Google Scholar] [CrossRef]
- Sechrest, W.; Brooks, T.M.; da Fonseca, G.A.B.; Konstant, W.R.; Mittermeier, R.A.; Purvis, A.; Rylands, A.B.; Gittleman, J.L. Hotspots and the conservation of evolutionary history. Proc. Natl. Acad. Sci. USA 2002, 99, 2067–2071. [Google Scholar] [CrossRef] [PubMed]
- Pesaresi, S.; Galdenzi, D.; Biondi, E.; Casavecchia, S. Bioclimate of Italy: Application of the worldwide bioclimatic classification system. J. Maps 2014, 10, 538–553. [Google Scholar] [CrossRef]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.M.G.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Matthews, T.J.; Triantis, K.A.; Whittaker, R.J.; Guilhaumon, F. Sars: An R package for fitting, evaluating and comparing Species–Area Relationship models. Ecography 2019, 42, 1446–1455. [Google Scholar] [CrossRef]
- Scheiner, S.M. Six types of Species-Area curves. Glob. Ecol. Biogeogr. 2003, 12, 441–447. [Google Scholar] [CrossRef]
- Storch, D. The theory of the nested Species–Area Relationship: Geometric foundations of biodiversity scaling. J. Veg. Sci. 2016, 27, 880–891. [Google Scholar] [CrossRef]
- Fattorini, S. Detecting biodiversity hotspots by Species-Area Relationships: A case study of Mediterranean beetles. Cons. Biol. 2006, 20, 1169–1180. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Function Name | Total Species Adjusted R2 | Native Species Adjusted R2 | Alien Species Adjusted R2 |
|---|---|---|---|
| Asymptotic | 0.16 | 0.24 | 0.15 |
| Beta-P | * | * | 0.60 |
| Chapman–Richards | 0.44 | 0.44 | 0.41 |
| Logarithmic | 0.57 | 0.64 | 0.39 |
| Gompertz | 0.49 | 0.58 | * |
| Kobayashi | 0.80 | 0.78 | 0.68 |
| Linear | 0.46 | 0.49 | 0.46 |
| Logistic | 0.92 | 0.91 | 0.73 |
| Monod | 0.66 | 0.63 | 0.66 |
| Negative Exponential | 0.60 | 0.58 | 0.65 |
| Power | 0.92 | 0.91 | 0.73 |
| Rational | 0.80 | 0.81 | 0.70 |
| Weibull-3 | 0.92 | 0.91 | 0.73 |
| Weibull-4 | 0.92 | 0.91 | 0.73 |
| Administrative Region | Area (km2) | Species Recorded | Species Expected | Residual |
|---|---|---|---|---|
| Liguria | 5418 | 3574 | 2701 | 32.3 |
| Friuli Venezia Giulia | 7924 | 3666 | 3006 | 22.0 |
| Trentino-Alto Adige | 13,606 | 4098 | 3499 | 17.1 |
| Abruzzo | 10,763 | 3604 | 3276 | 10.0 |
| Valle d’Aosta | 3263 | 2507 | 2343 | 7.0 |
| Veneto | 18,345 | 4003 | 3806 | 5.2 |
| Lombardia | 23,844 | 4242 | 4097 | 3.5 |
| Toscana | 22,985 | 4102 | 4055 | 1.2 |
| Molise | 4461 | 2525 | 2558 | −1.3 |
| Lazio | 17,242 | 3593 | 3740 | −3.9 |
| Campania | 13,590 | 3298 | 3498 | −5.7 |
| Marche | 9344 | 2946 | 3148 | −6.4 |
| Piemonte | 25,387 | 3836 | 4169 | −8.0 |
| Basilicata | 9995 | 2878 | 3209 | −10.3 |
| Umbria | 8456 | 2709 | 3061 | −11.5 |
| Calabria | 15,222 | 3158 | 3611 | −12.5 |
| Emilia-Romagna | 22,510 | 3418 | 4031 | −15.2 |
| Sicilia | 25,711 | 3262 | 4184 | −22.0 |
| Puglia | 19,541 | 2962 | 3874 | −23.5 |
| Sardegna | 24,090 | 2963 | 4108 | −27.9 |
| Administrative Region | Area (km2) | Species Recorded | Species Expected | Residual |
|---|---|---|---|---|
| Liguria | 5418 | 3035 | 2352 | 29.0 |
| Friuli Venezia Giulia | 7924 | 2984 | 2600 | 14.8 |
| Abruzzo | 10,763 | 3207 | 2818 | 13.8 |
| Valle d’Aosta | 3263 | 2299 | 2059 | 11.7 |
| Trentino-Alto Adige | 13,606 | 3119 | 2997 | 4.1 |
| Molise | 4461 | 2319 | 2235 | 3.7 |
| Toscana | 22,985 | 3422 | 3440 | −0.5 |
| Piemonte | 25,387 | 3486 | 3531 | −1.3 |
| Veneto | 18,345 | 3183 | 3242 | −1.8 |
| Lazio | 17,242 | 3045 | 3190 | −4.5 |
| Basilicata | 9995 | 2637 | 2764 | −4.6 |
| Lombardia | 23,844 | 3293 | 3474 | −5.2 |
| Campania | 13,590 | 2829 | 2996 | −5.6 |
| Marche | 9344 | 2528 | 2715 | −6.9 |
| Calabria | 15,222 | 2797 | 3087 | −9.4 |
| Umbria | 8456 | 2371 | 2645 | −10.3 |
| Emilia-Romagna | 22,510 | 2826 | 3421 | −17.4 |
| Sicilia | 25,711 | 2765 | 3543 | −22.0 |
| Puglia | 19,541 | 2562 | 3296 | −22.3 |
| Sardegna | 24,090 | 2330 | 3483 | −33.1 |
| Administrative Region | Area (km2) | Species Recorded | Species Expected | Residual |
|---|---|---|---|---|
| Liguria | 5418 | 492 | 326 | 51.1 |
| Lombardia | 23,844 | 807 | 593 | 36.2 |
| Friuli Venezia Giulia | 7924 | 509 | 380 | 34.0 |
| Trentino-Alto Adige | 13,606 | 616 | 472 | 30.4 |
| Veneto | 18,345 | 656 | 533 | 23.1 |
| Toscana | 22,985 | 657 | 584 | 12.5 |
| Emilia-Romagna | 22,510 | 585 | 579 | 1.0 |
| Lazio | 17,242 | 516 | 520 | −0.7 |
| Marche | 9344 | 400 | 406 | −1.5 |
| Campania | 13,590 | 465 | 472 | −1.5 |
| Piemonte | 25,387 | 560 | 608 | −7.9 |
| Abruzzo | 10,763 | 378 | 430 | −12.0 |
| Sardegna | 24,090 | 520 | 595 | −12.6 |
| Sicilia | 25,711 | 485 | 611 | −20.6 |
| Umbria | 8456 | 306 | 390 | −21.5 |
| Puglia | 19,541 | 390 | 547 | −28.7 |
| Calabria | 15,222 | 352 | 494 | −28.8 |
| Molise | 4461 | 192 | 301 | −36.2 |
| Valle d’Aosta | 3263 | 166 | 265 | −37.4 |
| Basilicata | 9995 | 248 | 417 | −40.5 |
| Name | Shape | Parameters | Formula |
|---|---|---|---|
| Asymptotic | convex | 3 (c, d, z) | S = d − c × zA |
| Beta-P | sigmoid | 4 (c, d, z, f) | S = d × (1 − (1 + (A/c)z)(−f)) |
| Chapman–Richards | sigmoid | 3 (c, d, z) | S = d × (1 − exp(−z × A)c) |
| Logarithmic | convex | 2 (c, z) | S = c + z × log(A) |
| Gompertz | sigmoid | 3 (c, d, z) | S = d × exp(−exp(−z × (A − c))) |
| Kobayashi | convex | 2 (c, z) | S = c × log(1 + A/z) |
| Linear | linear | 2 (c, z) | S = c + z × A |
| Logistic | sigmoid | 3 (c, f, z) | S = c/(f + A(−z)) |
| Monod | convex | 2 (c, d) | S = d/(1 + c × A(−1)) |
| Negative Exponential | convex | 2 (d, z) | S = d × (1 − exp(−z × A)) |
| Power | convex | 2 (c, z) | S = c × Az |
| Rational | convex | 3 (c, d, z) | S = (c + z × A)/(1 + d × A) |
| Weibull-3 | sigmoid | 3 (c, d, z) | S = d × (1 − exp(−c × Az)) |
| Weibull-4 | sigmoid | 4 (c, d, f, z) | S = d × (1 − exp(−c × Az))f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Antraccoli, M.; Peruzzi, L.; Conti, F.; Galasso, G.; Roma-Marzio, F.; Bartolucci, F. Floristic Richness in a Mediterranean Hotspot: A Journey across Italy. Plants 2024, 13, 12. https://doi.org/10.3390/plants13010012
D’Antraccoli M, Peruzzi L, Conti F, Galasso G, Roma-Marzio F, Bartolucci F. Floristic Richness in a Mediterranean Hotspot: A Journey across Italy. Plants. 2024; 13(1):12. https://doi.org/10.3390/plants13010012
Chicago/Turabian StyleD’Antraccoli, Marco, Lorenzo Peruzzi, Fabio Conti, Gabriele Galasso, Francesco Roma-Marzio, and Fabrizio Bartolucci. 2024. "Floristic Richness in a Mediterranean Hotspot: A Journey across Italy" Plants 13, no. 1: 12. https://doi.org/10.3390/plants13010012
APA StyleD’Antraccoli, M., Peruzzi, L., Conti, F., Galasso, G., Roma-Marzio, F., & Bartolucci, F. (2024). Floristic Richness in a Mediterranean Hotspot: A Journey across Italy. Plants, 13(1), 12. https://doi.org/10.3390/plants13010012