
Bacilli Rhizobacteria as Biostimulants of Growth and Production of Sesame Cultivars under Water Deficit

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results and Discussion
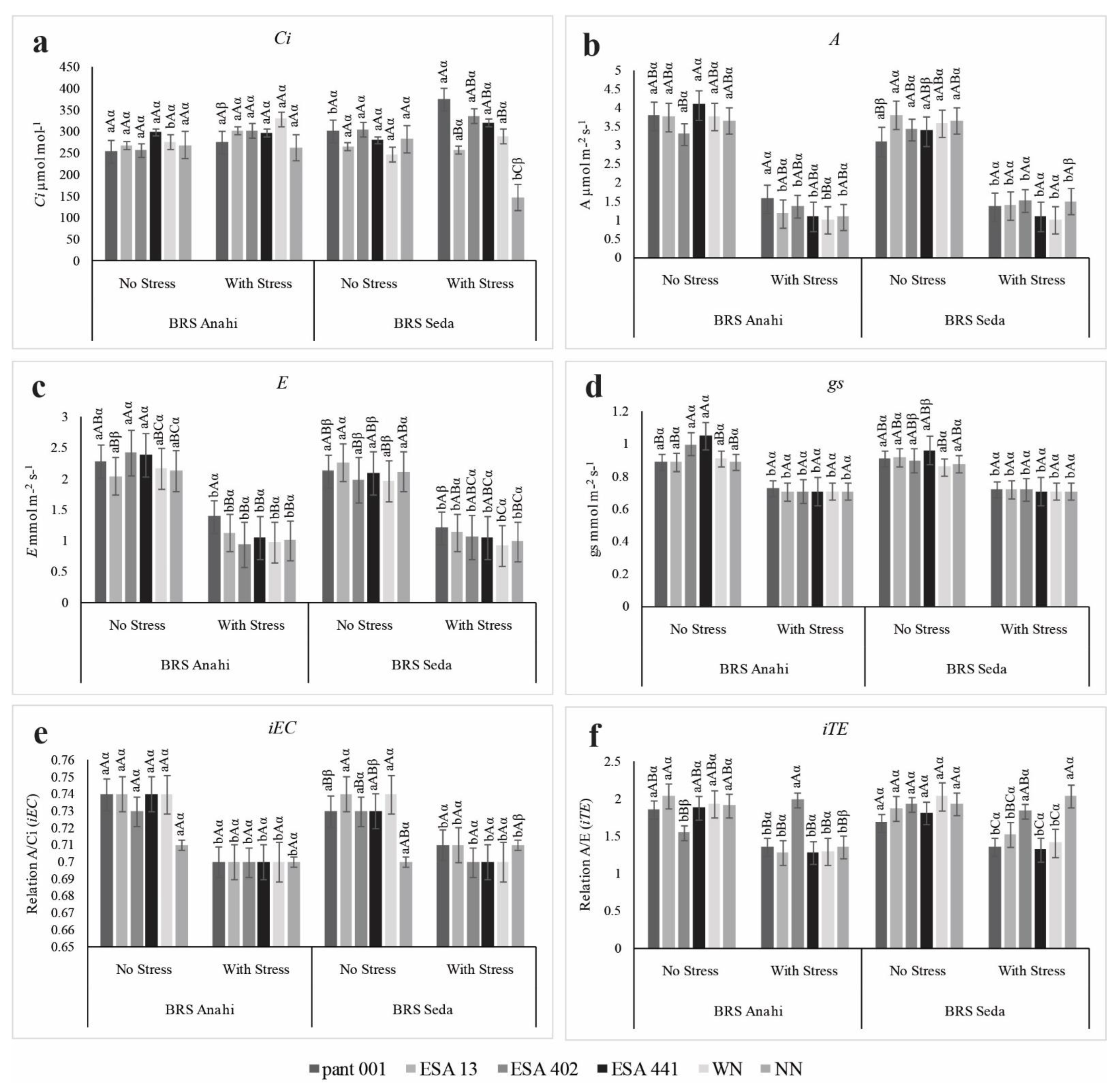
2.1. Gas Exchange and Fluorescence
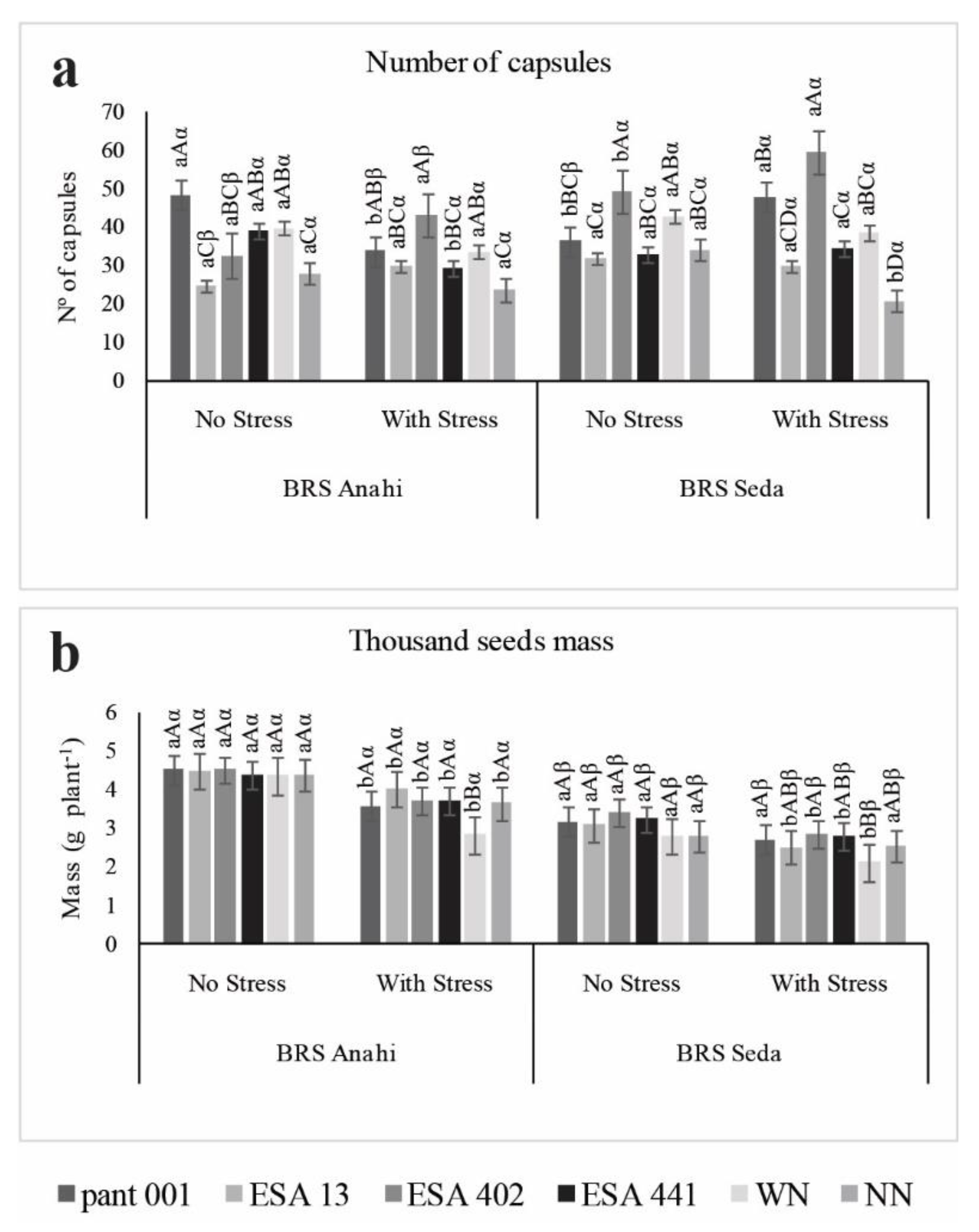
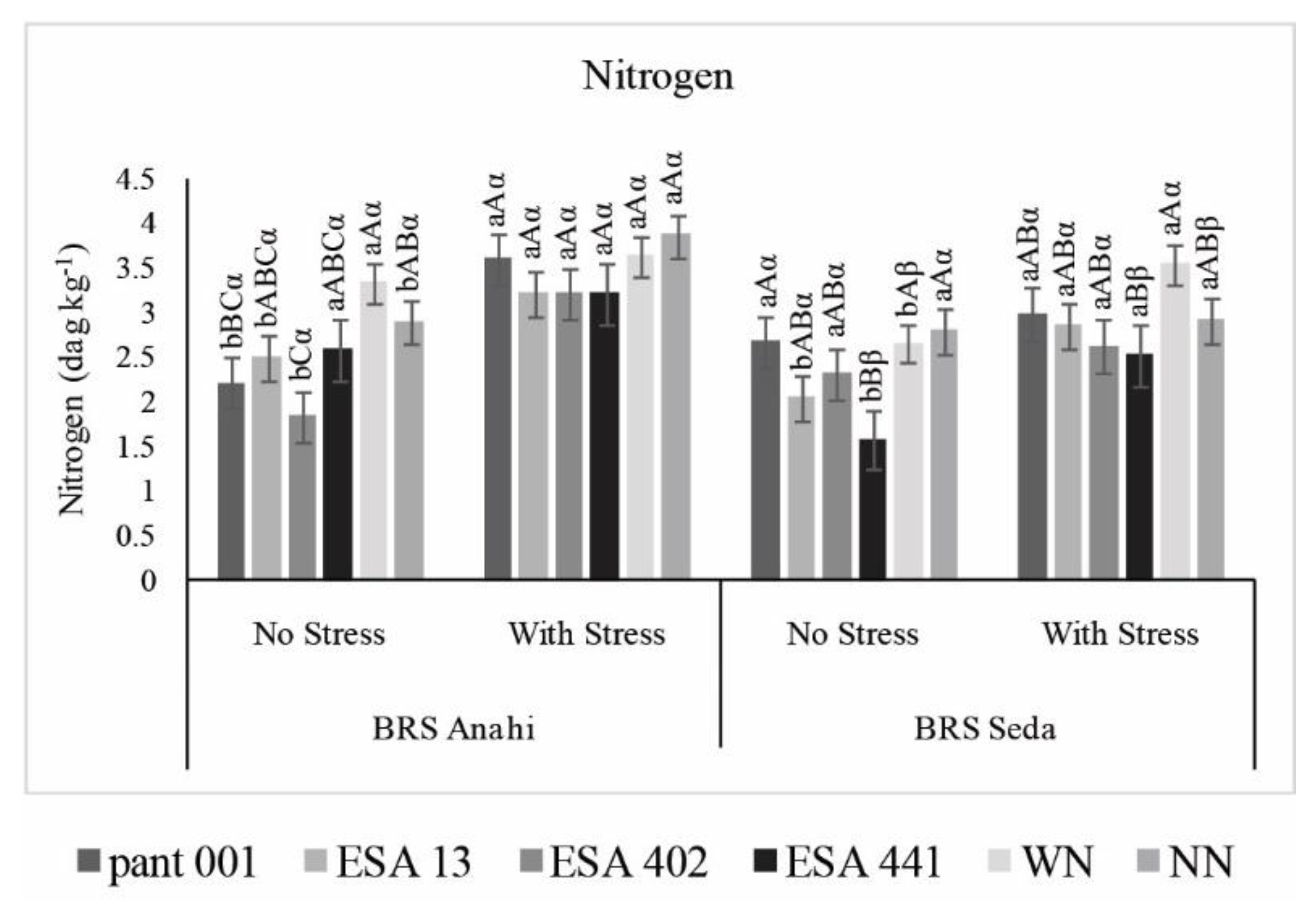
2.2. Biomass, Growth Measures and Nitrogen
2.3. Osmoregulation, Antioxidant Enzyme Complex and Chlorophyll Content
3. Materials and Methods
3.1. Cultivation of Bacteria and Preparation of Sesame Seeds
3.2. Implementation and Conduction of the Experiment in a Greenhouse
3.3. Physiological Measures
3.4. Nitrogen Content of the Shoot
3.5. Antioxidant Activities and Proline Content
3.6. Chlorophyll and Carotenoid Content
3.7. Biomass and Growth Measures
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Haque, M.M.; Mosharaf, M.K.; Khatun, M.; Haque, M.A.; Biswas, M.S.; Islam, M.S.; Islam, M.M.; Shozib, H.B.; Miah, M.M.U.; Molla, A.H.; et al. Biofilm producing rhizobacteria with multiple plant growth-promoting traits promote growth of tomato under water-deficit tress. Front. Microbiol. 2020, 11, 20–53. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2019, 128, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 17–109. [Google Scholar] [CrossRef]
- Kuss, A.V.; Kuss, V.V.; Lovato, T.; Flores, M.L. Fixação de nitrogênio e produção de ácido indolacético in vitro por bactérias diazotróficas endolíticas. Pesq. Agropec. Bras. 2007, 42, 1459–1465. [Google Scholar] [CrossRef]
- Kavamura, V.N.; Santos, S.N.; Silva, J.L.; Parma, M.M.; Ávila, L.A.; Visconti, A.; Zucchi, T.D.; Taketani, R.G.; Andreote, F.D.; Melo, I.S. Screening of brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol. Res. 2013, 168, 183–191. [Google Scholar] [CrossRef]
- Fernandes-Júnior, P.I.; Aidar, S.T.; Morgante, C.V.; Gava, C.A.T.; Zilli, J.É.; Souza, L.S.B.; Marinho, R.C.N.; Nóbrega, R.S.A.; Brasil, M.S.; Seido, S.L.; et al. The resurrection plant Tripogon Spicatus (Poaceae) harbors a diversity of plant growth promoting bacteria in Northeastern Brazilian Caatinga. Rev. Bras. Ciênc. Solo 2015, 39, 993–1002. [Google Scholar] [CrossRef]
- Santana, S.R.A.; Voltolini, T.V.; Antunes, G.R.; Silva, V.M.; Simões, W.L.; Morgante, C.V.; Freitas, A.D.S.; Chaves, A.R.M.; Aidar, S.T.; Fernandes, P.I., Jr. Inoculation of plant growth-promoting bacteria attenuates the negative effects of drought on sorghum. Arch. Microbiol. 2020, 202, 1015–1024. [Google Scholar] [CrossRef]
- Santos, A.F.J.; Morais, J.S.; Miranda, J.S.; Moreira, Z.P.M.; Feitoza, A.F.A.; Leite, J.; Fernandes, P.I., Jr. Cacti-associated rhizobacteria from Brazilian caatinga biome induce maize growth promotion and alleviate abiotic stress. Rev. Bras. Ciênc. Agrár. 2020, 15, e8221. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Dixon, R. Biotechnological solutions to the nitrogen problem. Curr. Opin. Biotechnol. 2014, 26, 19–24. [Google Scholar] [CrossRef]
- Bravo, G.; Vega-Celedón, P.; Gentina, J.C.; Seeger, M. Bioremediation by Cupriavidus metallidurans strain msr33 of mercury-polluted agricultural soil in a rotary drum bioreactor and its effects on nitrogen cycle microorganisms. Microorganisms 2020, 8, 1952. [Google Scholar] [CrossRef]
- Rahman, M.; Miah, M.N.A.; Dudding, W. Mechanisms involved with bacilli-mediated biotic and abiotic stress tolerance in plants. In Bacilli in Agrobiotechnology: Bacilli in Climate Resilient Agriculture and Bioprospecting; Islam, M.T., Rahman, M., Pandey, P., Eds.; Springer: Cham, Switzerland, 2022; pp. 169–197. [Google Scholar]
- Tsotetsi, T.; Nephali, L.; Malebe, M.; Tugizimana, F. Bacillus for plant growth promotion and stress resilience: What have we learned? Plants 2022, 11, 2482. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, R.C.; Cavalcanti, M.I.P.; Correia, A.J.; Escobar, I.E.C.; de Freitas, A.D.S.; Nóbrega, R.S.A.; Fernandes, P.I., Jr. Maize-associated bacteria from the brazilian semiarid region boost plant growth and grain yield. Symbiosis 2021, 83, 347–359. [Google Scholar] [CrossRef]
- Silva, J.F.; Silva, T.R.; Escobar, I.E.C.; Fraiz, A.C.R.; Santos, J.W.M.; Nascimento, T.R.; Santos, J.M.R.; Peters, S.J.W.; Melo, R.F.; Signor, D.; et al. Screening of plant growth promotion ability among bacteria isolated from field-grown sorghum under different managements in brazilian drylands. World J. Microbiol. Biotechnol. 2018, 34, 186. [Google Scholar] [CrossRef] [PubMed]
- Grover, M.; Bana, R.S.; Bodhankar, S. Prevalance of multifunctional Azospirillum Formosense strains in the rhizosphere of pearl millet across diverse edaphoclimatic regions of India. Vegetos 2022, 35, 1–11. [Google Scholar] [CrossRef]
- Nithyapriya, S.; Lalitha, S.; Sayyed, R.Z.; Reddy, M.S.; Dailin, D.J.; Enshasy, H.A.E.; Suriani, N.L.; Herlambang, S. Production, purification, and characterization of bacillibactin siderophore of Bacillus subtilis and its application for improvement in plant growth and oil content in sesame. Sustainability 2021, 13, 5394. [Google Scholar] [CrossRef]
- Varma, P.K.; Yamuna, C.; Suresh, V.; Teja, M.R.; Kumar, K.V.K. Potentiality of native Bacillus species in enhancing sesame seed germination and their antagonism against Macrophomina phaseolina under in vitro conditions. J. Oilseeds Res. 2017, 34, 98–102. [Google Scholar]
- Ferreira, M.D.; Spricigo, P.C. Colorimetria—Princípios e aplicações na agricultura. In Instrumentação pós-Colheita em Frutas e Hortaliças; Ferreira, M.D., Ed.; Embrapa Instrumentação: São Carlos, Brasil, 2017; pp. 209–220. [Google Scholar]
- Beltrão, N.E.M.; Oliveira, M.I.P. Ecofisiologia das Culturas de Algodão, Amendoim, Gergelim, Mamona, Pinhão-Manso e Sisal; Embrapa Informação Tecnológica: Brasília, Brazil, 2011. [Google Scholar]
- Arriel, N.H.C.; Beltrão, N.E.M.; Firmino, P.T. Clima. O produtor pergunta a Embrapa responde. In Gergelim; Beltrão, N.E.M., Arriel, N.H.C., Lima, R.L.S., Eds.; Embrapa Informação Tecnológica: Brasília, Brazil, 2009; pp. 57–88. [Google Scholar]
- Marulanda, A.; Porcel, R.; Barea, J.M.; Azcón, R. Drought tolerance and antioxidant activities in lavender plants colonized by native drought-tolerant or drought-sensitive. Microb. Ecol. 2007, 54, 543–552. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Allah, E.F.A. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi. J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Patel, R.R.; Patel, D.D.; Thakor, P.; Patel, B.; Thakkar, V.R. Alleviation of salt stress in germination of Vigna radiata L. by two halotolerant Bacilli sp. isolated from saline habitats of Gujarat. Plant Growth Regul. 2015, 76, 51–60. [Google Scholar] [CrossRef]
- Patel, R.R.; Patel, D.D.; Bhatt, J.; Thakor, P.; Triplett, L.R.; Thakkar, V.R. Induction of pre-chorismate, jasmonate and salicylate pathways by Burkholderia sp. RR18 in peanut seedlings. J. Appl. Microbiol. 2021, 131, 1417–1430. [Google Scholar] [CrossRef]
- Suassuna, J.; Fernandes, P.; Brito, M.; Arriel, N.M.A.; Fernandes, J. Tolerance to salinity of sesame genotypes in different phenological stages. Am. J. Plant Sci. 2017, 8, 1904–1920. [Google Scholar] [CrossRef]
- Lopes, M.S.; Araus, J.L.; Heerden, P.D.R.V.; Foyer, C.H. Enhancing drought tolerance in C4 crops. J. Exp. Bot. 2011, 62, 3135–3153. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.S.; Lima, G.S.L.; Gheyi, H.R.; Nobre, R.G.; Fernandes, P.D.; Silva, F.A.Z. Trocas gasosas e eficiência fotoquímica do gergelim sob estresse salino e adubação com nitrato-amônio. Irriga 2018, 23, 220–234. [Google Scholar] [CrossRef]
- Sousa, G.G.; Viana, T.V.A.; Dias, C.N.; Silva, G.L.S.; Azevedo, B.M. Lâminas de irrigação para cultura do gergelim com biofertilizante bovino. Magistra 2014, 26, 347–356. [Google Scholar]
- Lacerda, C.F.; Oliveira, E.V.; Neves, A.L.; Gheyi, H.R.; Bezerra, M.A.; Costa, C.A. Morphophysiological responses and mechanisms of salt tolerance in four ornamental perennial species under tropical climate. Rev. Bras. Eng. Agric. Ambient. 2020, 24, 656–663. [Google Scholar] [CrossRef]
- Silva, A.A.R.; Lacerda, C.N.; Lima, G.S.; Soares, L.A.A.; Gheyi, H.R.; Fernandes, P.D. Morfofisiologia de Cultivares de gergelim submetidos a diferentes estratégias de uso de água salina. Irriga 2021, 1, 42–55. [Google Scholar] [CrossRef]
- Agurla, S.; Gahir, S.; Munemasa, S.; Murata, Y.; Raghavendra, A.S. Mechanism of stomatal closure in plants exposed to drought and cold stress. Adv. Exp. Med. Biol. 2018, 1081, 215–232. [Google Scholar] [PubMed]
- Endres, L.; Souza, J.L.S.; Teodoro, I.; Marroquim, P.M.G.; Santos, C.M.S.; Brito, J.E.D. Gas exchange alteration caused by water deficit during the bean reproductive stage. Rev. Bras. Eng. Agric. Ambient. 2010, 14, 11–16. [Google Scholar] [CrossRef]
- Silva, E.M.; Lima, G.S.; Gheyi, H.R.; Nobre, R.G.; Sá, F.V.S.; Souza, L.P. Growth and gas exchanges in soursop under irrigation with saline water and nitrogen sources. Rev. Bras. Eng. Agric. Ambient. 2018, 22, 776–781. [Google Scholar] [CrossRef]
- Feitosa, S.S.; Albuquerque, M.B.; Oliveira, A.P.; Pereira, W.E.; Brito Neto, J.F. Fisiologia do Sesamum indicum L. sob estresse hídrico e aplicação de ácido salicílico. Irriga 2016, 21, 711–723. [Google Scholar] [CrossRef]
- Silva, E.M.; Ribeiro, R.V.; Ferreira-Silva, S.L.; Viégas, R.A.; Silveira, J.A.G. Salt stress induced damages on the photosynthesis of physic nut young plants. Sci. Agric. 2011, 68, 62–68. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Lin, Y.; Watts, D.B.; Kloepper, J.W.; Torbert, H.A. Influence of plant growth-promoting rhizobacteria on corn growth under different fertility sources. Commun. Soil Sci. Plant Anal. 2018, 49, 1239–1255. [Google Scholar] [CrossRef]
- Mesquita, J.B.R.; Azevedo, B.M.A.; Campelo, A.R.; Fernandes, C.N.V.; Viana, T.V.A. Crescimento e produtividade da cultura do gergelim (Sesamum indicum L.) sob diferentes níveis de irrigação. Irriga 2013, 18, 364–375. [Google Scholar] [CrossRef][Green Version]
- Grilo, J.A.; Azevedo, P.V. Crescimento, desenvolvimento e produtividade do gergelim ‘BRS Seda’ na agrovila de Canudos, em Ceará Mirim (RN). Holos 2013, 2, 19–33. [Google Scholar] [CrossRef]
- Antunes, G.R.; Santana, S.R.A.; Escobar, I.E.C.; Brasil, M.S.; Araújo, G.G.L.; Voltolini, T.V.; Fernandes, P.I., Jr. Associative diazotrophic bacteria from forage grasses in the Brazilian semiarid region are effective plant growth promoters. Crop Pasture Sci. 2019, 70, 899–907. [Google Scholar] [CrossRef]
- Brito, S.L.; Santos, A.B.; Barbosa, D.D.; Fernandes, P.D.; Fernandes, P.I., Jr.; Lima, L.M. Bradyrhizobium spp. as attenuators of water deficit stress in runner peanut genotypes based on physiological and gene expression responses. Genet. Mol. Res. 2019, 18, 1–12. [Google Scholar] [CrossRef]
- Mitter, B.; Brader, G.; Pfaffenbichler, N.; Sessitsch, A. Next generation microbiome applications for crop production-limitations and the need of knowledge-based solutions. Curr. Opin. Microbiol. 2019, 49, 59–65. [Google Scholar] [CrossRef]
- Jadhav, S.R.; Naiknaware, M.D.; Pawar, G.R. Effect of nitrogen, phosphorus and biofertilizers on growth, yield and quality of summer sesamum (Sesamum indicum L.). Int. J. Trop. Agric. 2015, 33, 475–480. [Google Scholar]
- Campos, L.N.; Guilherme, M.F.S.; Oliveira, H.M.; Costa, V.F.; Silva, E. Avaliação biométrica de sementes de Sesamum indicum L. In Proceedings of the Congresso Nordestino de Biólogos, João Pessoa, PB, Brazil, 27–28 March 2014. [Google Scholar]
- Melo, E.B.S.; Lima, L.M.; Fernandes-Junior, P.I.; Aidar, S.T.; Freire, M.A.O.; Freire, R.M.M.; Santos, R.C. Nodulation, gas exchanges and production of peanut cultivated with Bradyrhizobium in soils with different textures. Comun. Sci. 2016, 7, 160–166. [Google Scholar] [CrossRef]
- Couch, A.; Jani, A.; Mulvaney, M.; Hochmuth, G.; Bennett, J.; Gloaguen, R.; Langham, R.; Rowland, D. Nitrogen accumulation, partitioning, and remobilization by diverse sesame cultivars in the humid southeastern USA. Field Crops Res. 2017, 203, 55–64. [Google Scholar] [CrossRef]
- Dias, M.C.; Oliveira, H.; Costa, A.; Santos, C. Improving elms performance under drought stress: The retreatment with abscisic acid. Environ. Exp. Bot. 2014, 100, 64–73. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Lin, C.C.; Hsu, Y.T.; Kao, C.H. The effect of NaCl on proline accumulation in rice leaves. Plant Growth Regul. 2002, 36, 275–285. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Sankar, B.; Kishorekumar, A.; Gopi, R.; Somasundaram, R.; Panneerselvam, R. Pseudomonas fluorescens enhances biomass yield and ajmalicine production in Catharanthus roseus under water deficit stress. Colloids Surf. B Biointerfaces 2007, 60, 7–11. [Google Scholar] [CrossRef]
- Ozturk, M.; Unal, B.T.; Carrapós, G.P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Gupta, D.K.; Plama, J.M.; Corpas, F.J. Antioxidants and Antioxidant Enzymes in Higher Plants; Springer International Publishing: Berlim, Germany, 2018. [Google Scholar]
- Manivannan, P.; Jaleel, C.A.; Somasundaram, R.; Panneerselvam, R. Osmoregulation and antioxidant metabolism in drought-stressed Helianthus annuus under triadimefon drenching. C. R. Biol. 2008, 331, 418425. [Google Scholar] [CrossRef]
- Kadkhodaie, A.; Razmjoo, J.; Zahedi, M. Peroxisase, ascorbate peroxidase e catalase activities in drought sensitive, intermediate and resistence sesame (Sesamum indicum L. ) genotypes. Int. J. Agron. Plant Prod. 2013, 4, 3012–3021. [Google Scholar]
- Gholinezhad, E.; Darvishzadeh, R.; Moghaddam, S.S.; Popović-Djordjević, J. Effect of mycorrhizal inoculation in reducing water stress in sesame (Sesamum indicum L.): The assessment of agrobiochemical traits and enzymatic antioxidant activity. Agric. Water Manag. 2020, 238, 106–234. [Google Scholar] [CrossRef]
- Sousa, C.C.M.; Pedrosa, E.M.R.; Rolim, M.M.; Oliveira Filho, R.A.; Souza, M.A.L.M.; Pereira Filho, J.V. Crescimento e respostas enzimáticas do feijoeiro caupi sob estresse hídrico e nematoide de galhas. Rev. Bras. Eng. Agric. Ambient. 2015, 19, 113–118. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutase (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Hussein, Y.; Amin, J.; Azeb, A.; Gahin, H. Antioxidant activities during drought stress resistance of sesame (Sesamum indicum L.) plant by salicylic acid and kinetin. Res. J. Bot. 2016, 11, 1–8. [Google Scholar] [CrossRef][Green Version]
- Yousefzadeh-Najafabadi, M.; Ehsanzadeh, P. Photosynthetic and antioxidative upregulation in drought-stressed sesame (Sesamum indicum L.) subjected to foliar applied salicylic acid. Photosynthetica 2017, 55, 611–622. [Google Scholar] [CrossRef]
- Varamin, J.K.; Fanoodi, F.; Sinaki, J.M.; Rezvan, S.; Damavandi, A. Foliar application of chitosan and nano-magnesium fertilizers influence on seed yield, oil content, photosynthetic pigments, antioxidant enzyme activities of sesame (Sesamum indicum L.) under water-limited conditions. Not. Bot. Horti Agrobot. 2020, 48, 2228–2243. [Google Scholar] [CrossRef]
- Breusegem, F.V.; Vranová, E.; Dat, J.F.; Inzé, D. The role of active oxygen species in plant signal transduction. Plant Sci. 2001, 161, 405–414. [Google Scholar] [CrossRef]
- Carneiro, M.M.C.; Deuner, S.; Oliveira, P.V.; Teixeira, S.B.; Sousa, C.P.; Bacarin, M.A.; Moraes, D.M. Atividade antioxidante e viabilidade de sementes de girassol após estresse hídrico e salino. Rev. Bras. Sementes 2011, 33, 752–761. [Google Scholar] [CrossRef]
- Beltrão, N.E.M.; Vieira, D.J. O agronegócio do Gergelim No Brasil; Embrapa Informação Tecnológica: Brasília, Brazil, 2001. [Google Scholar]
- Perin, A.; Cruvinel, J.D.; Silva, W.J. Sesame performance as a function of NPK fertilization and soil fertility level. Acta Sci. Agron. 2010, 32, 93–98. [Google Scholar]
- Coelho, A.P.; Leal, F.P.; Filla, V.A.; Dalri, A.B.; Faria, R.T. Estimativa da produtividade de grãos da aveia-branca cultivada sob níveis de irrigação utilizando clorofilômetro portátil. Rev. Cient. Fac. Educ. Meio Amb. 2018, 9, 662–667. [Google Scholar] [CrossRef]
- Anaya, F.; Fghire, R.; Wahbi, S.; Loutfi, K. Influence of salicylic acid on seed germination of Vicia faba L. under salt stress. J. Saudi Soc. Agric. Sci. 2018, 17, 1–8. [Google Scholar] [CrossRef]
- Silva, J.F. Caracterização Polifásica de Bactérias Promotoras de Crescimento Vegetal Associados ao Sorgo (Sorghum Bicolor (L.) Moench) e ao milheto (Penissetum Glaucum (L.) R. Brown) Cultivados no sertão de Pernambuco; UNIVASF: Petrolina, Brazil, 2017. [Google Scholar]
- Vincent, J.M. A Manual for the Practical Study of Nodule Bacteria; Blackkwell Science Publication: Edinburgh, UK, 1970; p. 164. [Google Scholar]
- Carvalho, M.A.C.; Sá, M.E.; Campos, D.T.S.; Machado, A.P.; Chagas, A.F., Jr. Bacillus subtilis UFMT-Pant001 as a plant growth promoter in soybean in a greenhouse. Afr. J. Agric. Res. 2023, 19, 161–169. [Google Scholar]
- Silveira, C.A.P.; Bamberg, A.L.; Martinazzo, R.; Pillon, C.N.; Martins, E.S.; Piana, C.F.B.; Ferreira, L.H.G.; Pereira, I.S.; Stumpf, L. Instruções para Planejamento e Condução de Experimentos com Fertilizantes, Inoculantes, Corretivos, Biofertilizantes, Remineralizadores e Substratos para Plantas. Available online: https://www.gov.br/agricultura/pt-br/assuntos/insumos-agropecuarios/insumos-agricolas/fertilizantes/registro-estab-e-prod/registro-produtos/instrucoes-para-conducao-de-experimentos.pdf (accessed on 23 February 2023).
- Silva, F.C. Manual de Análises Químicas de Solos, Plantas e Fertilizantes; Embrapa Informação Tecnológica: Brasília, Brazil, 2009. [Google Scholar]
- Gomes, R.V.; Coutinho, J.L.B. Gergelim. In Recomendações de Adubação Para o Estado de Pernambuco, 2nd ed.; Cavalcanti, F.J.A., Ed.; Instituto Agronômico de Pernambuco: Recife, Brazil, 1998; p. 144. [Google Scholar]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How plants cope with stress in the field: Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef]
- Larbi, A.; Abadıa, A.; Abadıa, J.; Morales, F. Down co-regulation of light absorption, photochemistry, and carboxylation in Fe-deficient plants growing in different environments. Photosynth. Res. 2006, 89, 113–126. [Google Scholar] [CrossRef]
- Rohácek, K. Chlorophyll fluorescence parameters: The definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Bezerra Neto, E.; Barreto, L.P. Análises Químicas e Bioquímicas em Plantas; Imprensa Universitária da UFRPE: Recife, Brazil, 2011. [Google Scholar]
- Alcantara, R.M.; Xavier, G.R.; Rumjanek, N.G.; Rocha, M.M.; Carvalho, J.S. Eficiência simbiótica de progenitores de cultivares brasileiras de feijão-caupi. Rev. Ciênc. Agron. 2014, 45, 1–9. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 7, 248–254. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide desmutases occurrence in higher plants. Plant Physiol. 1977, 59, 14–309. [Google Scholar] [CrossRef]
- Azevedo, R.A.; Alas, R.M.; Smith, R.J.; Lea, P.J. Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild type and a catalase deficient mutant of barley. Physiol. Plant 1998, 104, 280–292. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbato-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bates, L.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar]
- IPGRI (International Board for Plant Genetic Resources). Descriptors for sesame (Sesamum spp.); NBPGR: New Delhi, India, 2004; 76p. [Google Scholar]
- Ferreira, D.F. Sisvar: A guide for its bootstrap procedures in multiple comparisons. Ciênc. Agrotec. 2014, 38, 109–112. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ci | E | A | gs | Proline | SOD | APX | CAT | |
|---|---|---|---|---|---|---|---|---|
| BRS Anahí | ||||||||
| pant 001 | ˄ | ˅ | ˅ | ˅ | ˄ | ˄ | ˅ | ˄ |
| ESA 13 | ˄ | ˅ | ˅ | ˅ | ˄ | ˅ | ˄ | ˅ |
| ESA 402 | ˄ | ˅ | ˅ | ˅ | ˄ | ˄ | ˅ | ˄ |
| ESA 441 | ˄ | ˅ | ˅ | ˅ | ˄ | ˄ | ˅ | ˄ |
| Nitrogen | ˄ | ˅ | ˅ | ˅ | ˄ | ˄ | ˅ | ˄ |
| No Nitrogen | ˅ | ˅ | ˅ | ˅ | ˄ | ˄ | ˄ | ˅ |
| BRS Seda | ||||||||
| pant 001 | ˄ | ˅ | ˅ | ˅ | ˄ | ˄ | ˄ | ˅ |
| ESA 13 | ˅ | ˅ | ˅ | ˅ | ˄ | ˅ | ˅ | ˅ |
| ESA 402 | ˄ | ˅ | ˅ | ˅ | ˄ | ˄ | ˅ | ˄ |
| ESA 441 | ˄ | ˅ | ˅ | ˅ | ˄ | ˄ | ˅ | ˄ |
| Nitrogen | ˄ | ˅ | ˅ | ˅ | ˄ | ˅ | ˅ | ˅ |
| No Nitrogen | ˅ | ˅ | ˅ | ˅ | ˄ | ˄ | ˄ | ˄ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, G.B.P.d.; Gomes, E.F.; Rocha, G.M.G.d.; Silva, F.d.A.; Fernandes, P.D.; Machado, A.P.; Fernandes-Junior, P.I.; Melo, A.S.d.; Arriel, N.H.C.; Gondim, T.M.d.S.; et al. Bacilli Rhizobacteria as Biostimulants of Growth and Production of Sesame Cultivars under Water Deficit. Plants 2023, 12, 1337. https://doi.org/10.3390/plants12061337
Lima GBPd, Gomes EF, Rocha GMGd, Silva FdA, Fernandes PD, Machado AP, Fernandes-Junior PI, Melo ASd, Arriel NHC, Gondim TMdS, et al. Bacilli Rhizobacteria as Biostimulants of Growth and Production of Sesame Cultivars under Water Deficit. Plants. 2023; 12(6):1337. https://doi.org/10.3390/plants12061337
Chicago/Turabian StyleLima, Giliard Bruno Primo de, Erika Fernandes Gomes, Geisenilma Maria Gonçalves da Rocha, Francisco de Assis Silva, Pedro Dantas Fernandes, Alexandre Paulo Machado, Paulo Ivan Fernandes-Junior, Alberto Soares de Melo, Nair Helena Castro Arriel, Tarcisio Marcos de Souza Gondim, and et al. 2023. "Bacilli Rhizobacteria as Biostimulants of Growth and Production of Sesame Cultivars under Water Deficit" Plants 12, no. 6: 1337. https://doi.org/10.3390/plants12061337
APA StyleLima, G. B. P. d., Gomes, E. F., Rocha, G. M. G. d., Silva, F. d. A., Fernandes, P. D., Machado, A. P., Fernandes-Junior, P. I., Melo, A. S. d., Arriel, N. H. C., Gondim, T. M. d. S., & Lima, L. M. d. (2023). Bacilli Rhizobacteria as Biostimulants of Growth and Production of Sesame Cultivars under Water Deficit. Plants, 12(6), 1337. https://doi.org/10.3390/plants12061337