

The Dual Benefit of Plant Essential Oils against Tuta absoluta
 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Results
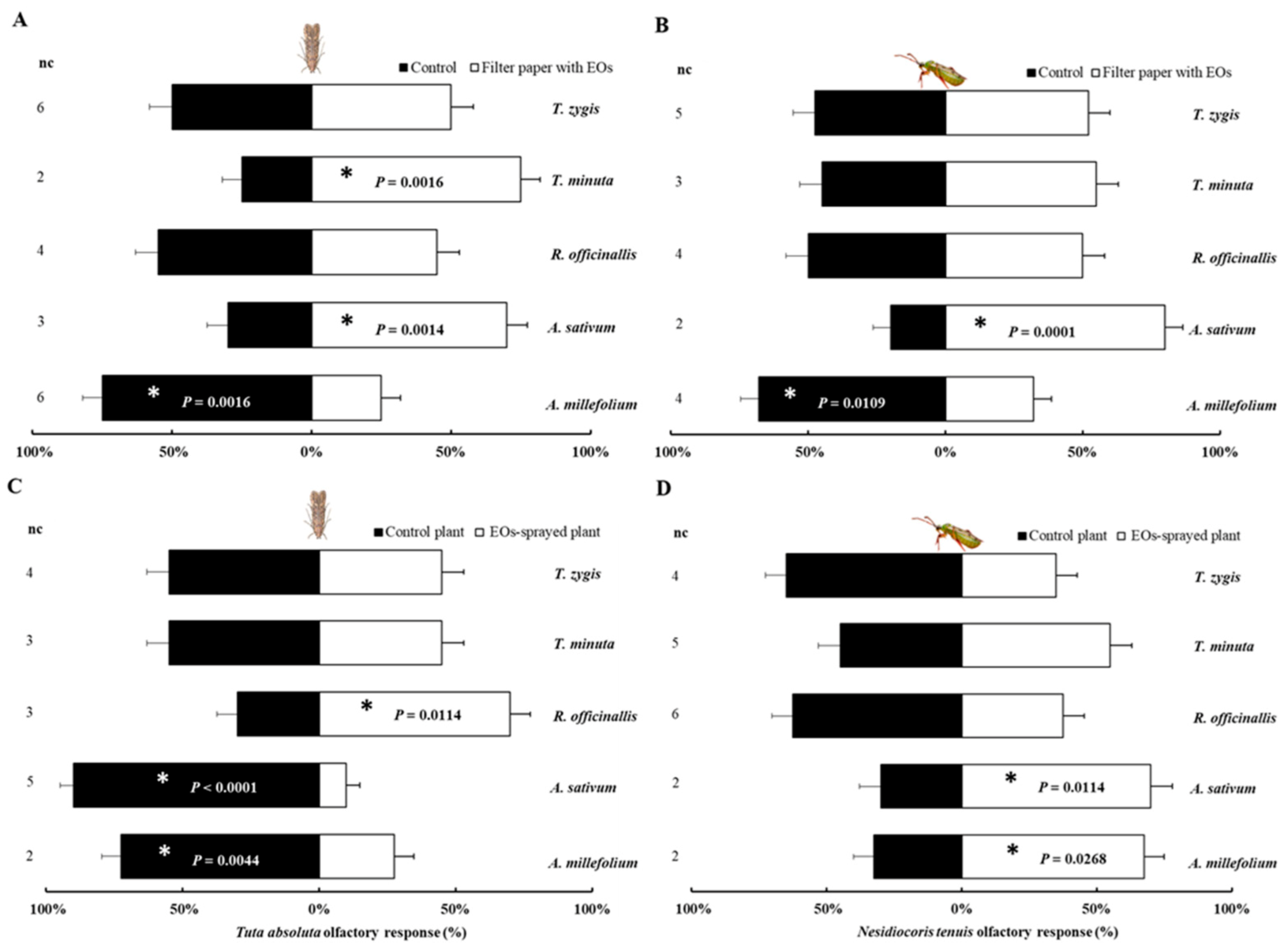
2.1. Olfactory Response of Tuta absoluta and Nesidiocoris tenuis to PEOs
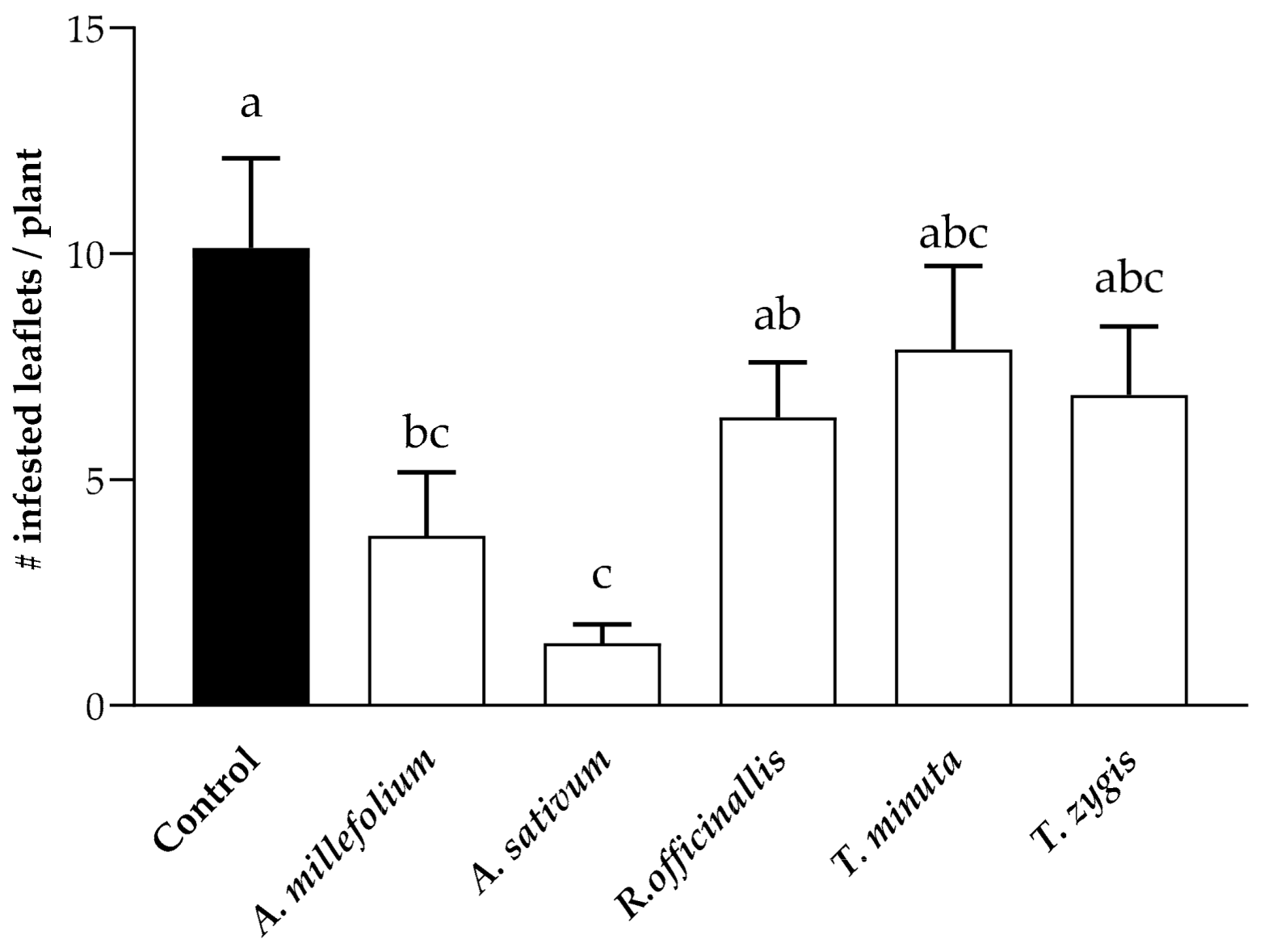
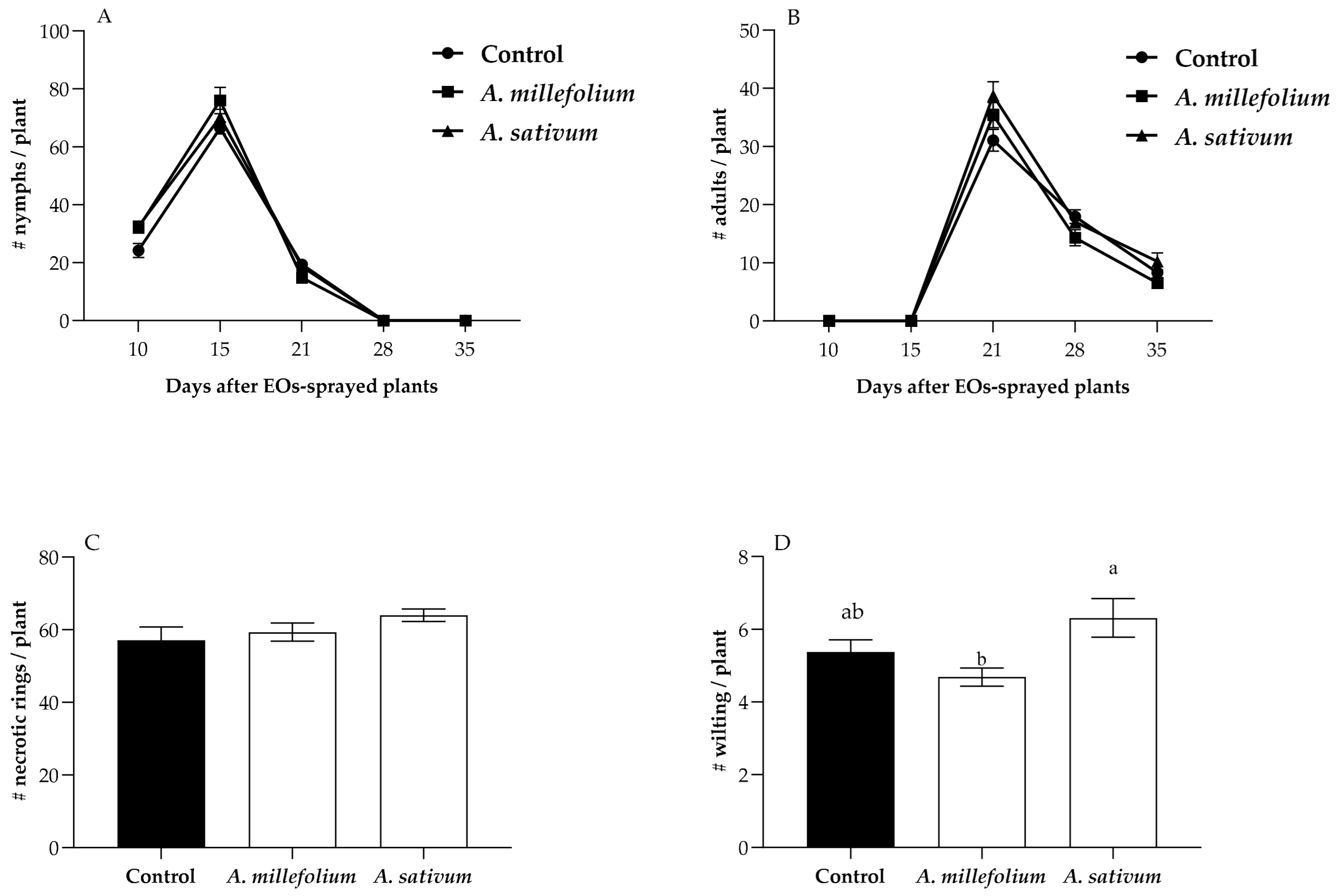
2.2. Effect of PEOs on the T. absoluta-Infested Leaflets
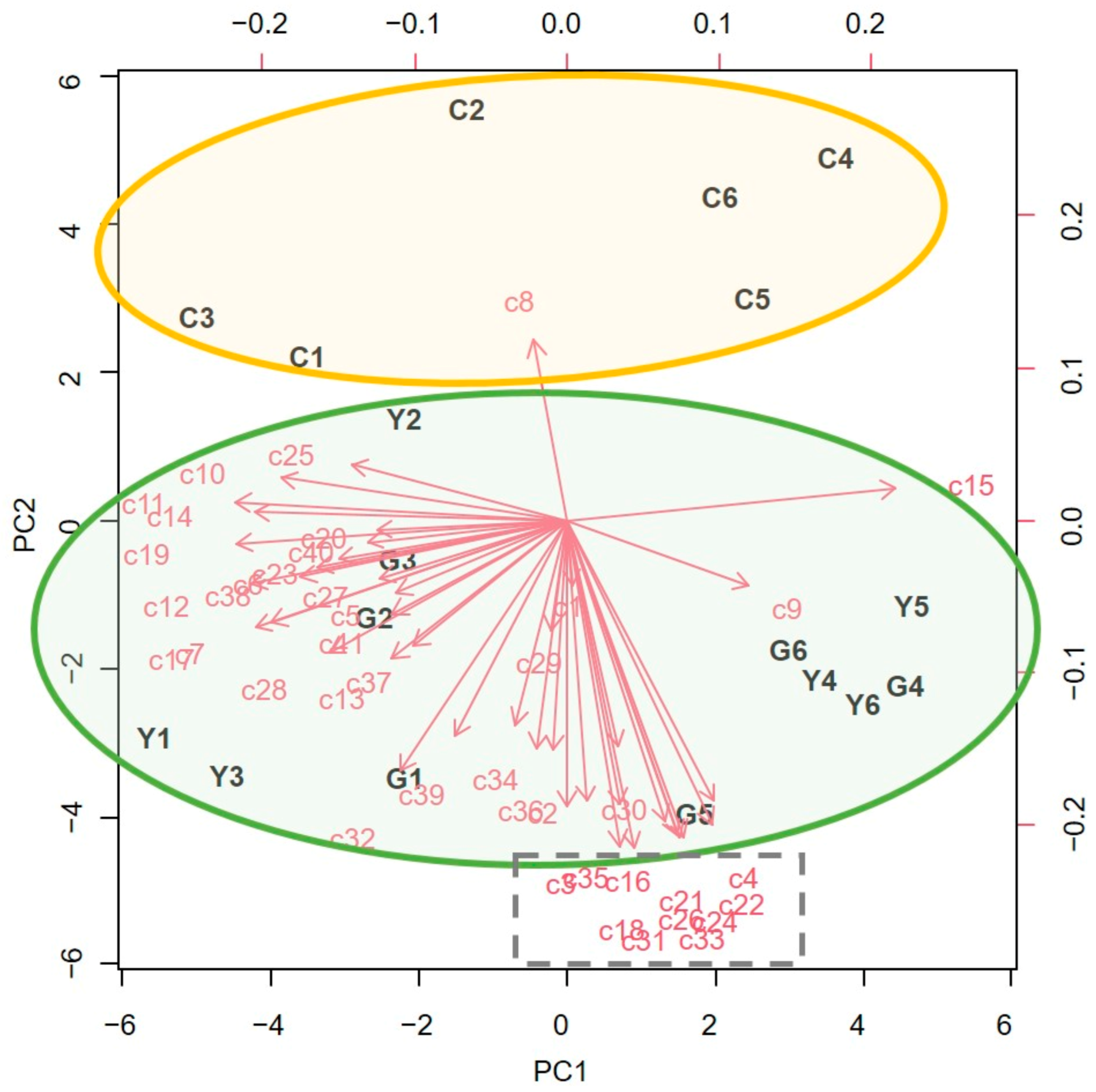
2.3. VOCs Emitted by A. millefolium or A. sativum- Sprayed Tomato Plants
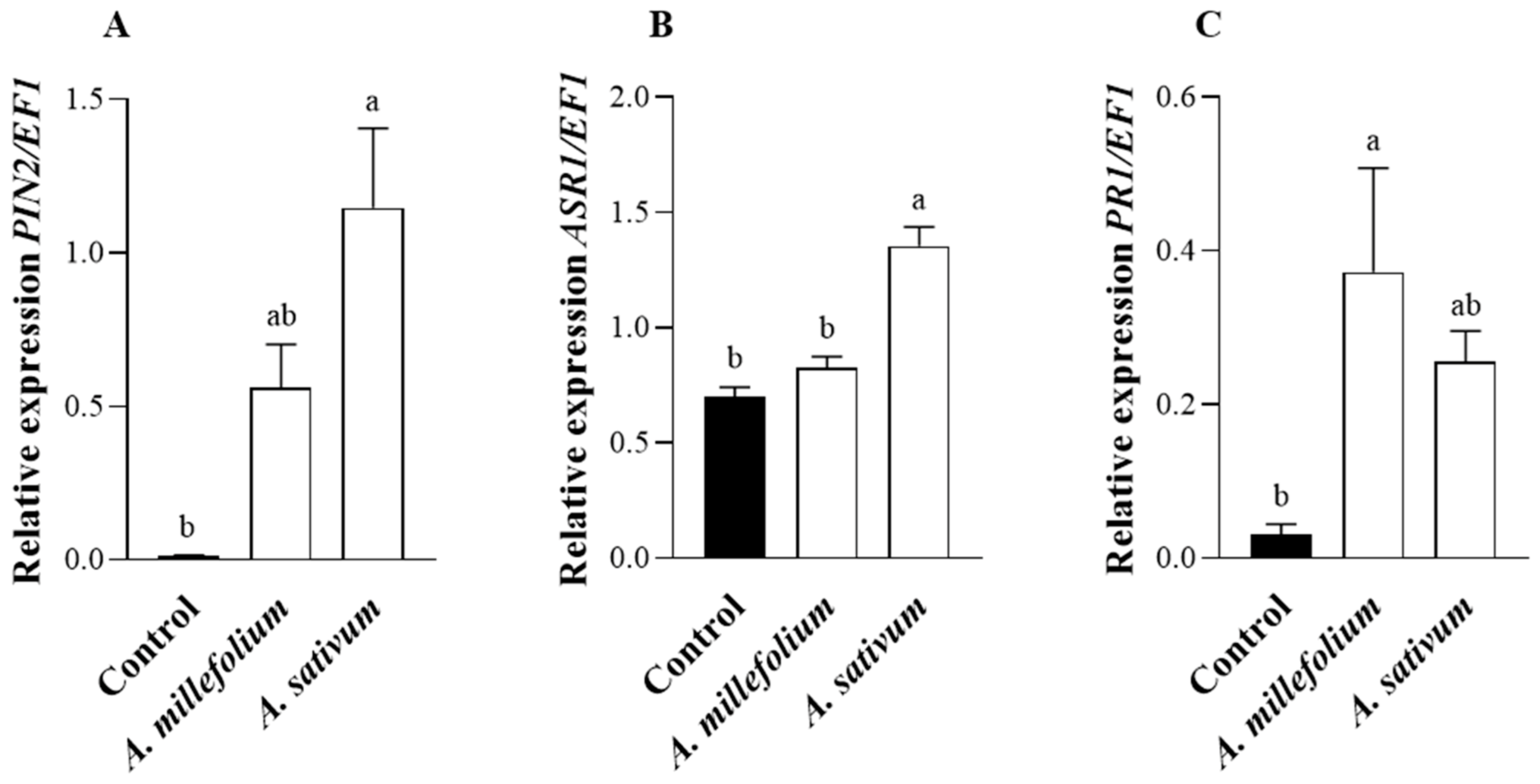
2.4. Defense Gene Expression in A. millefolium or A. sativum- Sprayed Tomato Plants
2.5. Side-Effect of A. millefolium and A. sativum on N. tenuis
3. Discussion
4. Materials and Methods
4.1. Plants, Plant Essential Oils, and Insects
4.2. Olfactory Response of T. absoluta and N. tenuis to PEOs
4.3. The Suitability of PEOs in Reducing T. absoluta-Infested Leaflets
4.4. Headspace Collection and Analysis of Volatile Compounds Induced by Plants Exposed to PEOs
4.5. Plant Gene Expression
4.6. Side-Effects of A. millefolium and A. sativum on N. tenuis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide Exposure, Safety Issues, and Risk Assessment Indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, A.; Benelli, G. Towards Pesticide-Free Farming? Sharing Needs and Knowledge Promotes Integrated Pest Management. Environ. Sci. Pollut. Res. 2018, 25, 13439–13445. [Google Scholar] [CrossRef] [PubMed]
- Jampílek, J.; Kráľová, K. Nanobiopesticides in Agriculture: State of the Art and Future Opportunities. In Nano-Biopesticides Today and Future Perspectives; Academic Press: Cambridge, MA, USA, 2019; pp. 397–447. ISBN 9780128158296. [Google Scholar]
- Moretti, M.D.L.; Sanna-Passino, G.; Demontis, S.; Bazzoni, E. Essential Oil Formulations Useful as a New Tool for Insect Pest Control. AAPS PharmSciTech 2002, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Sell, C. Chemistry of Essential Oils. In Handbook of Essential Oils: Science, Technology, and Applications, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 161–189. ISBN 9781466590472. [Google Scholar]
- Kesraoui, S.; Andrés, M.F.; Berrocal-Lobo, M.; Soudani, S.; Gonzalez-Coloma, A. Direct and Indirect Effects of Essential Oils for Sustainable Crop Protection. Plants 2022, 11, 2144. [Google Scholar] [CrossRef] [PubMed]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of Plant Defense against Insect Herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Commercial Development of Plant Essential Oils and Their Constituents as Active Ingredients in Bioinsecticides. Phytochem. Rev. 2020, 19, 235–241. [Google Scholar] [CrossRef]
- Isman, M.B. Bioinsecticides Based on Plant Essential Oils: A Short Overview. Z. Naturforsch. C 2020, 75, 179–182. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Oluwaseun Ademiluyi, A.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems; Universidade de Coimbra: Coimbra, Portugal, 2017; Volume 22, ISBN 9819919533. [Google Scholar]
- Pérez-Hedo, M.; Arias-Sanguino, Á.M.; Urbaneja, A. Induced Tomato Plant Resistance against Tetranychus urticae Triggered by the Phytophagy of Nesidiocoris tenuis. Front. Plant Sci. 2018, 9, 1419. [Google Scholar] [CrossRef]
- Bouagga, S.; Urbaneja, A.; Rambla, J.L.; Flors, V.; Granell, A.; Jaques, J.A.; Pérez-Hedo, M. Zoophytophagous Mirids Provide Pest Control by Inducing Direct Defences, Antixenosis and Attraction to Parasitoids in Sweet Pepper Plants. Pest Manag. Sci. 2018, 74, 1286–1296. [Google Scholar] [CrossRef]
- Cruz-Miralles, J.; Cabedo-López, M.; Pérez-Hedo, M.; Flors, V.; Jaques, J.A. Zoophytophagous Mites Can Trigger Plant-genotype Specific Defensive Responses Affecting Potential Prey beyond Predation: The Case of Euseius stipulatus and Tetranychus urticae in Citrus. Pest Manag. Sci. 2019, 75, 1962–1970. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Rambla, J.L.; Granell, A.; Urbaneja, A. Biological Activity and Specificity of Miridae-Induced Plant Volatiles. BioControl 2018, 63, 203–213. [Google Scholar] [CrossRef]
- Arraiza, M.P.; González-Coloma, A.; Andres, M.F.; Berrocal-Lobo, M.; Domínguez-Núñez, J.A.; da Costa, A.C., Jr.; Navarro-Rocha, J.; Calderón-Guerrero, C. Antifungal Effect of Essential Oils. In Potential of Essential Oils; El-Shemy, H.A., Ed.; IntechOpen Lim.: London, UK, 2018; pp. 145–164. [Google Scholar]
- Desneux, N.; Luna, M.G.; Guillemaud, T.; Urbaneja, A. The Invasive South American Tomato Pinworm, Tuta absoluta, Continues to Spread in Afro-Eurasia and beyond: The New Threat to Tomato World Production. J. Pest Sci. 2011, 84, 403–408. [Google Scholar] [CrossRef]
- Urbaneja, A.; González-Cabrera, J.; Arnó, J.; Gabarra, R. Prospects for the Biological Control of Tuta absoluta in Tomatoes of the Mediterranean Basin. Pest Manag. Sci. 2012, 68, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Catalán Ruescas, D.; Tabone, E.; Frandon, J.; et al. Biological Invasion of European Tomato Crops by Tuta absoluta: Ecology, Geographic Expansion and Prospects for Biological Control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Buragohain, P.; Saikia, D.K.; Sotelo-Cardona, P.; Srinivasan, R. Development and Validation of an Integrated Pest Management Strategy against the Invasive South American Tomato Leaf Miner, Tuta absoluta in South India. Crop Prot. 2021, 139, 105348. [Google Scholar] [CrossRef]
- Desneux, N.; Han, P.; Mansour, R.; Arnó, J.; Brévault, T.; Campos, M.R.; Chailleux, A.; Guedes, R.N.C.; Karimi, J.; Konan, K.A.J.; et al. Integrated Pest Management of Tuta absoluta: Practical Implementations across Different World Regions. J. Pest Sci. 2022, 95, 17–39. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Suay, R.; Alonso, M.; Ruocco, M.; Giorgini, M.; Poncet, C.; Urbaneja, A. Resilience and Robustness of IPM in Protected Horticulture in the Face of Potential Invasive Pests. Crop Prot. 2017, 97, 119–127. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Alonso-Valiente, M.; Vacas, S.; Gallego, C.; Pons, C.; Arbona, V.; Rambla, J.L.; Navarro-Llopis, V.; Granell, A.; Urbaneja, A. Plant Exposure to Herbivore-Induced Plant Volatiles: A Sustainable Approach through Eliciting Plant Defenses. J. Pest Sci. 2021, 94, 1221–1235. [Google Scholar] [CrossRef]
- Umpiérrez, M.L.; Lagreca, M.E.; Cabrera, R.; Grille, G.; Rossini, C. Essential Oils from Asteraceae as Potential Biocontrol Tools for Tomato Pests and Diseases. Phytochem. Rev. 2012, 11, 339–350. [Google Scholar] [CrossRef]
- Campolo, O.; Cherif, A.; Ricupero, M.; Siscaro, G.; Grissa-Lebdi, K.; Russo, A.; Cucci, L.M.; Di Pietro, P.; Satriano, C.; Desneux, N.; et al. Citrus Peel Essential Oil Nanoformulations to Control the Tomato Borer, Tuta absoluta: Chemical Properties and Biological Activity. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Ricupero, M.; Biondi, A.; Cincotta, F.; Condurso, C.; Palmeri, V.; Verzera, A.; Zappalà, L.; Campolo, O. Bioactivity and Physico-Chemistry of Garlic Essential Oil Nanoemulsion in Tomato. Entomol. Gen. 2022, 42, 921–930. [Google Scholar] [CrossRef]
- Goudarzvand Chegini, S.; Abbasipour, H. Chemical Composition and Insecticidal Effects of the Essential Oil of Cardamom, Elettaria Cardamomum on the Tomato Leaf Miner, Tuta absoluta. Toxin Rev. 2017, 36, 12–17. [Google Scholar] [CrossRef]
- Bouayad Alam, S.; Dib, M.E.A.; Djabou, N.; Tabti, B.; Gaouar Benyelles, N.; Costa, J.; Muselli, A. Essential Oils as Biocides for the Control of Fungal Infections and Devastating Pest (Tuta absoluta) of Tomato (Lycopersicon esculentum Mill.). Chem. Biodivers. 2017, 14, e1700065. [Google Scholar] [CrossRef] [PubMed]
- Essoung, F.R.E.; Tadjong, A.T.; Chhabra, S.C.; Mohamed, S.A.; Hassanali, A. Repellence and Fumigant Toxicity of Essential Oils of Ocimum gratissimum and Ocimum kilimandscharicum on Tuta absoluta (Lepidoptera: Gelechiidae). Environ. Sci. Pollut. Res. 2020, 27, 37963–37976. [Google Scholar] [CrossRef]
- Msisi, D.; Matojo, N.D.; Kimbokota, F. Attraction of Female Tomato Leaf Miner, Tuta absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae) to Shared Compounds from Hosts. Phytoparasitica 2021, 49, 153–162. [Google Scholar] [CrossRef]
- Yarou, B.B.; Bawin, T.; Boullis, A.; Heukin, S.; Lognay, G.; Verheggen, F.J.; Francis, F. Oviposition Deterrent Activity of Basil Plants and Their Essentials Oils against Tuta absoluta (Lepidoptera: Gelechiidae). Environ. Sci. Pollut. Res. 2018, 25, 29880–29888. [Google Scholar] [CrossRef]
- Galvan, T.L.; Koch, R.L.; Hutchison, W.D. Toxicity of Commonly Used Insecticides in Sweet Corn and Soybean to Multicolored Asian Lady Beetle (Coleoptera: Coccinellidae). J. Econ. Entomol. 2005, 98, 780–789. [Google Scholar] [CrossRef]
- Silva, D.B.; Weldegergis, B.T.; Van Loon, J.J.A.; Bueno, V.H.P. Qualitative and Quantitative Differences in Herbivore-Induced Plant Volatile Blends from Tomato Plants Infested by Either Tuta absoluta or Bemisia babaci. J. Chem. Ecol. 2017, 43, 53–65. [Google Scholar] [CrossRef]
- Abdelmaksoud, N.M.; Abdel-Aziz, N.F.; Sammour, E.A.; Agamy, E.A.E.M.; El-Bakry, A.M.; Kandil, M.A.-H. Influence of Insect Traps and Insecticides Sequential Application as a Tactic for Management of Tomato Leafminer, Tuta absoluta (Meyrick), (Lepidoptera: Gelechiidae). Bull. Natl. Res. Cent. 2020, 44, 123. [Google Scholar] [CrossRef]
- Cárdenas-Ortega, N.C.; González-Chávez, M.M.; Figueroa-Brito, R.; Flores-Macías, A.; Romo-Asunción, D.; Martínez-González, D.E.; Pérez-Moreno, V.; Ramos-López, M.A. Composition of the Essential Oil of Salvia ballotiflora (Lamiaceae) and Its Insecticidal Activity. Molecules 2015, 20, 8048–8059. [Google Scholar] [CrossRef]
- Luo, D.Y.; Yan, Z.T.; Che, L.R.; Zhu, J.J.; Chen, B. Repellency and Insecticidal Activity of Seven Mugwort (Artemisia argyi) Essential Oils against the Malaria Vector Anopheles sinensis. Sci. Rep. 2022, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bleeker, P.M.; Mirabella, R.; Diergaarde, P.J.; VanDoorn, A.; Tissier, A.; Kant, M.R.; Prins, M.; De Vos, M.; Haring, M.A.; Schuurink, R.C. Improved Herbivore Resistance in Cultivated Tomato with the Sesquiterpene Biosynthetic Pathway from a Wild Relative. Proc. Natl. Acad. Sci. USA 2012, 109, 20124–20129. [Google Scholar] [CrossRef] [PubMed]
- Piri, A.; Sahebzadeh, N.; Zibaee, A.; Sendi, J.J.; Shamakhi, L.; Shahriari, M. Toxicity and Physiological Effects of Ajwain (Carum Copticum, Apiaceae) Essential Oil and Its Major Constituents against Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Chemosphere 2020, 256, 127103. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Martínez, L.C.; Dos Santos, M.H.; Fernandes, F.L.; Wilcken, C.F.; Soares, M.A.; Serrão, J.E.; Zanuncio, J.C. Insecticidal Activity of Garlic Essential Oil and Their Constituents against the Mealworm Beetle, Tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae). Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.; Amini, J.; Saba, M.K.; Karimi, K.; Pertot, I. Preharvest and Postharvest Application of Garlic and Rosemary Essential Oils for Controlling Anthracnose and Quality Assessment of Strawberry Fruit During Cold Storage. Front. Microbiol. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Bhatnagar-Thomas, P.L.; Pal, A. Studies on the Insecticidal Activity of Garlic Oil. II. Mode of Action of the Oil as a Pesticide in Musca domestica Nebulo Fabr and Trogoderma granarium Everts. J. Food Sci. Technol. 1974, 11, 153–158. [Google Scholar]
- Singh, D. Enzyme Inhibition by Allicin, the Molluscicidal Agent of Allium sativum L. (Garlic). Phyther. Res. 1996, 10, 383–386. [Google Scholar] [CrossRef]
- Sukegawa, S.; Shiojiri, K.; Higami, T.; Suzuki, S.; Arimura, G.-I. Pest Management Using Mint Volatiles to Elicit Resistance in Soy: Mechanism and Application Potential. Plant J. 2018, 96, 910–920. [Google Scholar] [CrossRef]
- Okada, K.; Abe, H.; Arimura, G. Jasmonates Induce Both Defensive and Infochemical Strategies in Monocotyledonous and Dicotyledonous Plants. Plant Cell Physiol. 2015, 56, 16–27. [Google Scholar] [CrossRef]
- Zehra, A.; Meena, M.; Dubey, M.K.; Aamir, M.; Upadhyay, R.S. Synergistic Effects of Plant Defense Elicitors and Trichoderma harzianum on Enhanced Induction of Antioxidant Defense System in Tomato against Fusarium Wilt Disease. Bot. Stud. 2017, 58, 44. [Google Scholar] [CrossRef]
- Vucetic, A.; Dahlin, I.; Petrovic-Obradovic, O.; Glinwood, R.; Webster, B.; Ninkovic, V. Volatile Interaction between Undamaged Plants Affects Tritrophic Interactions through Changed Plant Volatile Emission. Plant Signal. Behav. 2014, 9, e29517. [Google Scholar] [CrossRef]
- Riahi, C.; González-Rodríguez, J.; Alonso-Valiente, M.; Urbaneja, A.; Pérez-Hedo, M. Eliciting Plant Defenses through Herbivore-Induced Plant Volatiles’ Exposure in Sweet Peppers. Front. Ecol. Evol. 2022, 9, 776827. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Alonso-Valiente, M.; Vacas, S.; Gallego, C.; Rambla, J.L.; Navarro-Llopis, V.; Granell, A.; Urbaneja, A. Eliciting Tomato Plant Defenses by Exposure to Herbivore Induced Plant Volatiles. Entomol. Gen. 2021, 41, 209–218. [Google Scholar] [CrossRef]
- Silva, D.B.; Urbaneja, A.; Pérez-Hedo, M. Response of Mirid Predators to Synthetic Herbivore-Induced Plant Volatiles. Entomol. Exp. Appl. 2021, 169, 125–132. [Google Scholar] [CrossRef]
- Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.; Schuurink, R. Green Leaf Volatiles: A Plant’s Multifunctional Weapon against Herbivores and Pathogens. Int. J. Mol. Sci. 2013, 14, 17781–17811. [Google Scholar] [CrossRef]
- Cofer, T.M.; Seidl-Adams, I.; Tumlinson, J.H. From Acetoin to (Z)-3-Hexen-1-Ol: The Diversity of Volatile Organic Compounds That Induce Plant Responses. J. Agric. Food Chem. 2018, 66, 11197–11208. [Google Scholar] [CrossRef] [PubMed]
- Piesik, D.; Pańka, D.; Jeske, M.; Wenda-Piesik, A.; Delaney, K.J.; Weaver, D.K. Volatile Induction of Infected and Neighbouring Uninfected Plants Potentially Influence Attraction/Repellence of a Cereal Herbivore. J. Appl. Entomol. 2013, 137, 296–309. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Y.; Wu, K.; Gao, X.W.; Guo, Y.Y. Field-Testing of Synthetic Herbivore-Induced Plant Volatiles as Attractants for Beneficial Insects. Environ. Entomol. 2008, 37, 1410–1415. [Google Scholar] [CrossRef] [PubMed]
- Kempraj, V.; Park, S.J.; Cameron, D.N.S.; Taylor, P.W. 1-Octanol Emitted by Oecophylla smaragdina Weaver Ants Repels and Deters Oviposition in Queensland Fruit Fly. Sci. Rep. 2022, 12, 15768. [Google Scholar] [CrossRef]
- Yu, J.; Yang, B.; Chang, Y.; Zhang, Y.; Wang, G. Identification of a General Odorant Receptor for Repellents in the Asian Corn Borer Ostrinia furnacalis. Front. Physiol. 2020, 11, 176. [Google Scholar] [CrossRef]
- Chinchilla-Ramírez, M.; Pérez-Hedo, M.; Pannebakker, B.A.; Urbaneja, A. Genetic Variation in the Feeding Behavior of Isofemale Lines of Nesidiocoris tenuis. Insects 2020, 11, 513. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Hedo, M.; Urbaneja, A. Prospects for Predatory Mirid Bugs as Biocontrol Agents of Aphids in Sweet Peppers. J. Pest Sci. 2015, 88, 65–73. [Google Scholar] [CrossRef]
- Chinchilla-Ramírez, M.; Garzo, E.; Fereres, A.; Gavara-Vidal, J.; ten Broeke, C.J.; van Loon, J.J.A.; Urbaneja, A.; Pérez-Hedo, M. Plant Feeding by Nesidiocoris tenuis: Quantifying Its Behavioral and Mechanical Components. Biol. Control 2021, 152, 104402. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| rt (min) 2 | Code 3 | Name | Treatment | ANOVA | ||
|---|---|---|---|---|---|---|
| Control | A. millefolium | A. sativum | ||||
| 4.77 | c1 | Z-3-hexenal | 0.079 ± 0.053 a | 0.056 ± 0.015 a | 0.033 ± 0.012 a | F 2,14 = 0.44, p = 0.6528 |
| 4.83 | c2 | hexanal | 0.064 ± 0.014 a | 0.156 ± 0.028 a | 0.080 ± 0.015 a | F 2,15 = 0.66, p = 0.5333 |
| 6.72 | c3 | Z-3-hexen-1-ol | 0.087 ± 0.066 a | 0.336 ± 0.066 b | 0.179 ± 0.054 ab | F 2,14 = 5.59, p = 0.0165 |
| 8.14 | c4 | heptanal | 0.003 ± 0.001 a | 0.089 ± 0.034 b | 0.089 ± 0.025 b | F 2,13 = 4.61, p = 0.0306 |
| 8.76 | c5 | 3-thujene | 0.047 ± 0.007 a | 0.062 ± 0.009 a | 0.051 ± 0.006 a | F 2,15 = 0.99, p = 0.3929 |
| 8.96 | c6 | α-pinene | 2.529 ± 0.286 a | 2.838 ± 0.346 a | 3.083 ± 0.385 a | F 2,15 = 0.61, p = 0.5542 |
| 9.94 | c7 | 3,7,7-trimethyl-1,3,5-cycloheptatriene | 3.024 ± 0.663 a | 3.712 ± 0.693 a | 3.185 ± 0.436 a | F 2,15 = 0.34, p = 0.7139 |
| 10.10 | c8 | β-pinene | 0.427 ± 0.203 a | 0.339 ± 0.149 a | 0.171 ± 0.022 a | F 2,15 = 0.97, p = 0.4009 |
| 10.40 | c9 | β-myrcene | 2.170 ± 0.253 a | 1.929 ± 0.276 a | 2.586 ± 0.393 a | F 2,15 = 1.05, p = 0.3741 |
| 10.64 | c10 | 2-carene | 24.257 ± 3.267 a | 23.441 ± 2.950 a | 24.054 ± 2.653 a | F 2,15 = 0.02, p = 0.9796 |
| 10.80 | c11 | α-phellandrene | 6.610 ± 0.981 a | 6.182 ± 1.080 a | 5.917 ± 0.540 a | F 2,15 = 0.12, p = 0.8876 |
| 11.05 | c12 | α-terpinene | 2.361 ± 0.521 a | 2.340 ± 0.532 a | 2.426 ± 0.366 a | F 2,15 = 0.03, p = 0.9700 |
| 11.23 | c13 | p-cymene | 0.262 ± 0.057 a | 0.446 ± 0.079 a | 0.465 ± 0.092 a | F 2,15 = 2.65, p = 0.1034 |
| 11.42 | c14 | limonene | 16.916 ± 3.394 a | 18.124 ± 5.063 a | 17.761 ± 5.033 a | F 2,15 = 0.00, p = 0.9979 |
| 11.49 | c15 | β-phellandrene | 36.615 ± 8.794 a | 32.838 ± 11.003 a | 33.174 ± 8.396 a | F 2,15 = 0.08, p = 0.9248 |
| 11.70 | c16 | (Z)-β-ocimene | 0.714 ± 0.157 a | 1.128 ± 0.104 b | 1.276 ± 0.141 b | F 2,15 = 4.16, p = 0.0364 |
| 11.98 | c17 | γ-terpinene | 0.408 ± 0.070 a | 0.454 ± 0.087 a | 0.455 ± 0.040 a | F 2,15 = 0.20, p = 0.8207 |
| 12.24 | c18 | 1-octanol | 0.000 ± 0.000 a | 0.033 ± 0.005 b | 0.027 ± 0.005 b | F 2,11 = 19.24, p = 0.0003 |
| 12.56 | c19 | terpinolene | 0.657 ± 0.130 a | 0.652 ± 0.170 a | 0.609 ± 0.098 a | F 2,15 = 0.02, p = 0.9765 |
| 12.66 | c20 | 3,4-dimethylstyrene | 0.033 ± 0.009 a | 0.039 ± 0.005 a | 0.029 ± 0.006 a | F 2,15 = 0.61, p = 0.5559 |
| 12.81 | c21 | (Z)-3-hexenyl propanoate | nd | 0.011 ± 0.003 a | 0.027 ± 0.010 a | F 1,10 = 2.32, p = 0.1590 |
| 12.94 | c22 | nonanal | 0.055 ± 0.007 a | 1.369 ± 0.385 b | 1.119 ± 0.276 b | F 2,15 = 11.75, p = 0.0009 |
| 13.20 | c23 | isoterpinolene | 0.174 ± 0.040 a | 0.186 ± 0.052 a | 0.126 ± 0.035 a | F 2,15 = 0.66, p = 0.5309 |
| 14.06 | c24 | 2-nonenal | 0.008 ± 0.003 a | 0.036 ± 0.009 a | 0.026 ± 0.006 a | F 2,12 = 2.93, p = 0.0922 |
| 14.42 | c25 | unknown | 0.036 ± 0.010 a | 0.023 ± 0.006 a | 0.034 ± 0.005 a | F 2,15 = 0.97, p = 0.4004 |
| 14.54 | c26 | (Z)-3-hexenyl butanoate | nd | 0.039 ± 0.013 a | 0.069 ± 0.024 a | F 1,10 = 0.82, p = 0.3862 |
| 14.61 | c27 | dill ether | 0.093 ± 0.008 a | 0.088 ± 0.018 a | 0.102 ± 0.021 a | F 2,15 = 0.13, p = 0.8765 |
| 14.71 | c28 | methyl salicylate | 0.021 ± 0.007 a | 0.053 ± 0.020 a | 0.033 ± 0.016 a | F 2,15 = 1.09, p = 0.3602 |
| 14.81 | c29 | dodecane | 0.032 ± 0.004 a | 0.040 ± 0.004 a | 0.031 ± 0.004 a | F 2,15 = 1.59, p = 0.2370 |
| 14.93 | c30 | decanal | 0.037 ± 0.008 a | 0.099 ± 0.012 b | 0.062 ± 0.010 c | F 2,15 = 11.01, p = 0.0011 |
| 15.38 | c31 | (Z)-3-hexenyl-2-methylbutanoate | nd | 0.009 ± 0.001 a | 0.015 ± 0.005 a | F 1,10 = 0.60, p = 0.4571 |
| 16.76 | c32 | isoascaridol | 0.043 ± 0.005 a | 0.054 ± 0.007 ab | 0.065 ± 0.004 b | F 2,15 = 4.14, p = 0.0369 |
| 17.01 | c33 | (Z)-3-hexenyl 2-methyl-(E)-2-butenoate | nd | 0.011 ± 0.003 a | 0.017 ± 0.002 a | F 1,10 = 0.11, p = 0.7487 |
| 17.27 | c34 | δ-elemene | 0.175 ± 0.039 a | 0.214 ± 0.059 a | 0.387 ± 0.062 b | F 2,15 = 4.04, p = 0.0395 |
| 17.43 | c35 | epoxide | 0.011 ± 0.001 a | 0.018 ± 0.002 b | 0.029 ± 0.003 c | F 2,15 = 18.18, p = 0.0001 |
| 18.19 | c36 | β-elemene | 0.019 ± 0.003 a | 0.026 ± 0.007 ab | 0.041 ± 0.005 b | F 2,15 = 3.94, p = 0.0423 |
| 18.30 | c37 | tetradecane | 0.018 ± 0.005 a | 0.027 ± 0.006 a | 0.029 ± 0.009 a | F 2,15 = 0.79, p = 0.4727 |
| 18.73 | c38 | β-caryophyllene | 1.885 ± 0.283 a | 2.310 ± 0.559 a | 1.985 ± 0.262 a | F 2,15 = 0.13, p = 0.8754 |
| 19.04 | c39 | sesquiterpene 1 | 0.054 ± 0.005 a | 0.071 ± 0.010 a | 0.064 ± 0.006 a | F 2,15 = 1.14, p = 0.3455 |
| 19.61 | c40 | sesquiterpene 2 | 0.057 ± 0.012 a | 0.055 ± 0.014 a | 0.033 ± 0.006 a | F 2,15 = 1.25, p = 0.3138 |
| 19.71 | c41 | sesquiterpene 3 | 0.041 ± 0.007 a | 0.038 ± 0.009 a | 0.040 ± 0.007 a | F 2,15 = 0.07, p = 0.9323 |
| 20.97 | c42 | 4,8,12-trimethyltrideca-1,3,7,11-tetraene | 0.016 ± 0.005 a | 0.038 ± 0.026 a | 0.026 ± 0.009 a | F 2,10 = 0.11, p = 0.8949 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Abdallah, S.; Riahi, C.; Vacas, S.; Navarro-Llopis, V.; Urbaneja, A.; Pérez-Hedo, M. The Dual Benefit of Plant Essential Oils against Tuta absoluta. Plants 2023, 12, 985. https://doi.org/10.3390/plants12050985
Ben Abdallah S, Riahi C, Vacas S, Navarro-Llopis V, Urbaneja A, Pérez-Hedo M. The Dual Benefit of Plant Essential Oils against Tuta absoluta. Plants. 2023; 12(5):985. https://doi.org/10.3390/plants12050985
Chicago/Turabian StyleBen Abdallah, Saoussen, Chaymaa Riahi, Sandra Vacas, Vicente Navarro-Llopis, Alberto Urbaneja, and Meritxell Pérez-Hedo. 2023. "The Dual Benefit of Plant Essential Oils against Tuta absoluta" Plants 12, no. 5: 985. https://doi.org/10.3390/plants12050985
APA StyleBen Abdallah, S., Riahi, C., Vacas, S., Navarro-Llopis, V., Urbaneja, A., & Pérez-Hedo, M. (2023). The Dual Benefit of Plant Essential Oils against Tuta absoluta. Plants, 12(5), 985. https://doi.org/10.3390/plants12050985