
Potential of Bioassays to Assess Consequences of Cultivation of Acacia mangium Trees on Nitrogen Bioavailability to Eucalyptus Trees: Two Case-Studies in Contrasting Tropical Soils
 , and
, and
Abstract
1. Introduction
2. Results
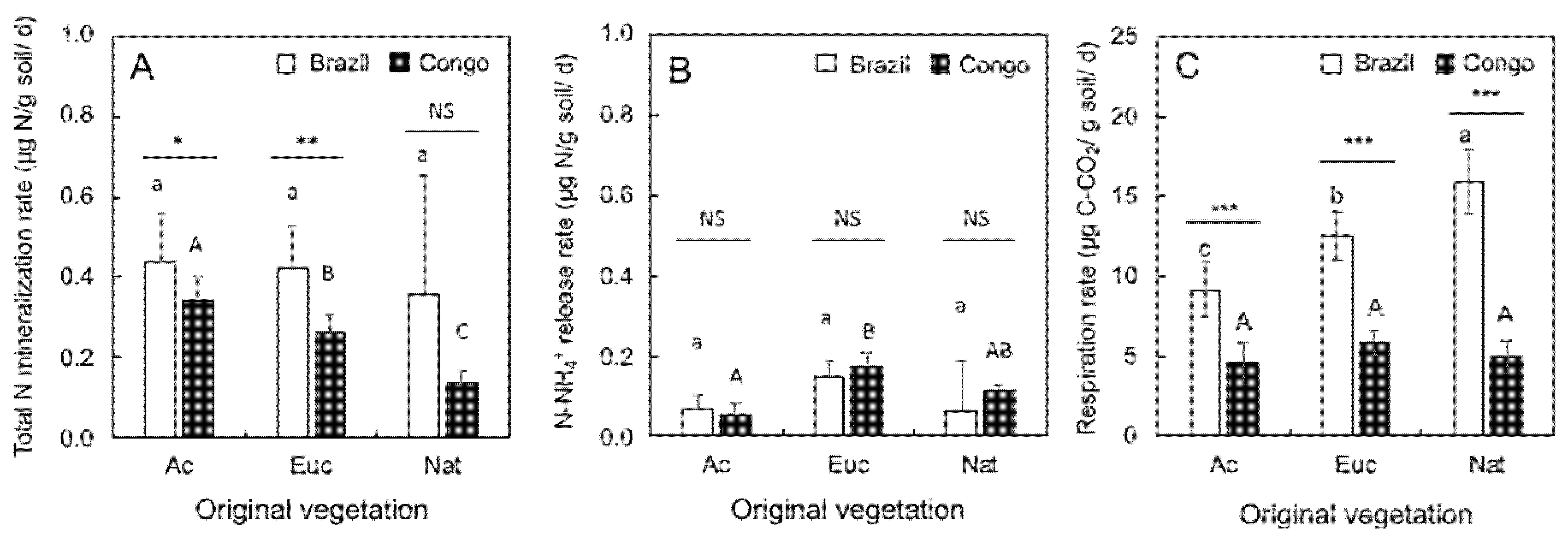
2.1. Soil N Mineralization Rates and Soil Respiration Rates
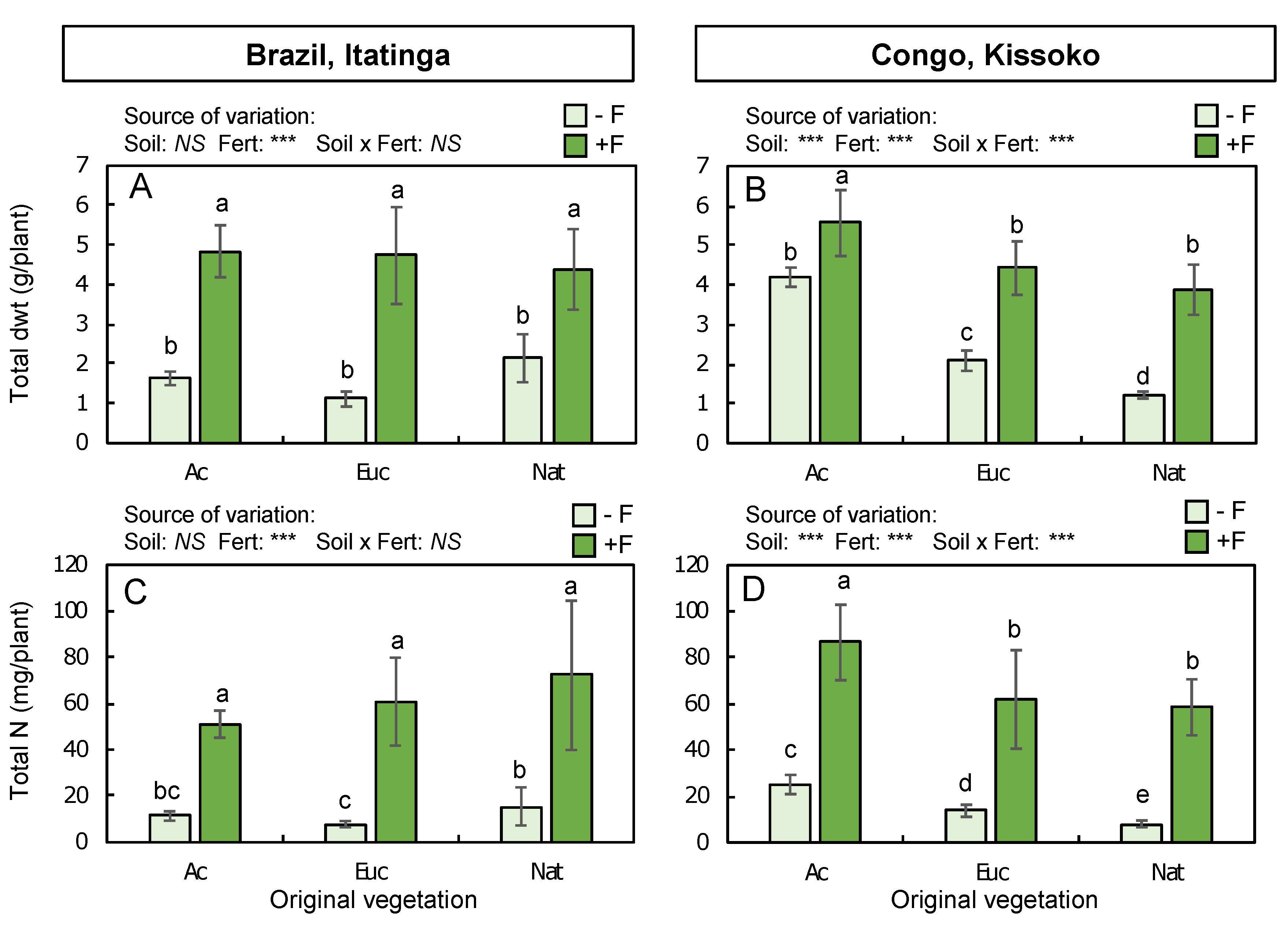
2.2. Eucalyptus Growth
2.3. N Accumulation in Eucalypt Plants
3. Discussion
3.1. Effect of Acacia Trees on N-Mineralization Rates
3.2. Effect of Acacia Trees on N Bioavailability for Eucalyptus Cultivation
4. Materials and Methods
4.1. Site Description
4.2. Soil Sampling
4.3. Measurement of Soil Respiration
4.4. Bioassay
4.5. Chemical Assays
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Global Forest Resources Assessment 2015; FAO Forestry Paper No. 1; UN Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of Global Forest Area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Booth, T.H. Eucalypt Plantations and Climate Change. For. Ecol. Manag. 2013, 301, 28–34. [Google Scholar] [CrossRef]
- de Moraes Gonçalves, J.L.; Alvares, C.A.; Higa, A.R.; Silva, L.D.; Alfenas, A.C.; Stahl, J.; de Barros Ferraz, S.F.; de Paula Lima, W.; Brancalion, P.H.S.; Hubner, A.; et al. Integrating Genetic and Silvicultural Strategies to Minimize Abiotic and Biotic Constraints in Brazilian Eucalypt Plantations. For. Ecol. Manag. 2013, 301, 6–27. [Google Scholar] [CrossRef]
- Stape, J.L.; Binkley, D.; Ryan, M.G.; Fonseca, S.; Loos, R.A.; Takahashi, E.N.; Silva, C.R.; Silva, S.R.; Hakamada, R.E.; Ferreira, J.M.d.A.; et al. The Brazil Eucalyptus Potential Productivity Project: Influence of Water, Nutrients and Stand Uniformity on Wood Production. For. Ecol. Manag. 2010, 259, 1684–1694. [Google Scholar] [CrossRef]
- Laclau, J.-P.; Ranger, J.; Deleporte, P.; Nouvellon, Y.; Saint-André, L.; Marlet, S.; Bouillet, J.-P. Nutrient Cycling in a Clonal Stand of Eucalyptus and an Adjacent Savanna Ecosystem in Congo: 3. Input–Output Budgets and Consequences for the Sustainability of the Plantations. For. Ecol. Manag. 2005, 210, 375–391. [Google Scholar] [CrossRef]
- Bouillet, J.-P.; Laclau, J.-P.; Gonçalves, J.L.d.M.; Voigtlaender, M.; Gava, J.L.; Leite, F.P.; Hakamada, R.; Mareschal, L.; Mabiala, A.; Tardy, F.; et al. Eucalyptus and Acacia Tree Growth over Entire Rotation in Single- and Mixed-Species Plantations across Five Sites in Brazil and Congo. For. Ecol. Manag. 2013, 301, 89–101. [Google Scholar] [CrossRef]
- Santos, F.M.; Balieiro, F.d.C.; Ataíde, D.H.d.S.; Diniz, A.R.; Chaer, G.M. Dynamics of Aboveground Biomass Accumulation in Monospecific and Mixed-Species Plantations of Eucalyptus and Acacia on a Brazilian Sandy Soil. For. Ecol. Manag. 2016, 363, 86–97. [Google Scholar] [CrossRef]
- Dias, A.C.; Arroja, L. Environmental Impacts of Eucalypt and Maritime Pine Wood Production in Portugal. J. Clean. Prod. 2012, 37, 368–376. [Google Scholar] [CrossRef]
- Brentrup, F.; Pallière, C. GHG Emissions and Energy Efficiency in European Nitrogen Fertiliser Production and Use. In Proceedings of the International Fertiliser Society Conference, Cambridge, UK, 11 December 2008; pp. 1–25. [Google Scholar]
- Fuchs, K.; Merbold, L.; Buchmann, N.; Bellocchi, G.; Bindi, M.; Brilli, L.; Conant, R.T.; Dorich, C.D.; Ehrhardt, F.; Fitton, N.; et al. Evaluating the Potential of Legumes to Mitigate N2O Emissions from Permanent Grassland Using Process-Based Models. Glob. Biogeochem. Cycles 2020, 34, e2020GB006561. [Google Scholar] [CrossRef]
- Bouillet, J.-P.; Safou Matondo, R.; Laclau, J.-P.; Nzila, J.d.D.; Ranger, J.; Deleporte, P. Pour Une Production Durable Des Plantations d’eucalyptus Au Congo: La Fertilisation. Bois For. Des Trop. 2004, 279, 23–35. [Google Scholar] [CrossRef]
- da Silva, P.H.M.; Poggiani, F.; Libardi, P.L.; Gonçalves, A.N. Fertilizer Management of Eucalypt Plantations on Sandy Soil in Brazil: Initial Growth and Nutrient Cycling. For. Ecol. Manag. 2013, 301, 67–78. [Google Scholar] [CrossRef]
- Du Toit, B.; Arbuthnot, A.; Oscroft, D.; Job, R.A. The Effects of Remedial Fertilizer Treatments on Growth and Pulp Properties of Eucalyptus grandis Stands Established on Infertile Soils of the Zululand Coastal Plain. South. Afr. For. J. 2001, 2001, 9–18. [Google Scholar]
- Noble, A.D.; Herbert, M.A. Influence of Soil Organic Matter Content on the Responsiveness of Eucalyptus grandis to Nitrogen Fertiliser. S. Afr. For. J. 1991, 156, 23–27. [Google Scholar]
- Laclau, J.P.; Ranger, J.; de Moraes Gonçalves, J.L.; Maquère, V.; Krusche, A.V.; M’Bou, A.T.; Nouvellon, Y.; Saint-André, L.; Bouillet, J.P.; de Cassia Piccolo, M.; et al. Biogeochemical Cycles of Nutrients in Tropical Eucalyptus Plantations. Main Features Shown by Intensive Monitoring in Congo and Brazil. For. Ecol. Manag. 2010, 259, 1771–1785. [Google Scholar] [CrossRef]
- Gonçalves, J.L.M.; Stape, J.L.; Laclau, J.P.; Smethurst, P.; Gava, J.L. Silvicultural Effects on the Productivity and Wood Quality of Eucalypt Plantations. For. Ecol. Manag. 2004, 193, 45–61. [Google Scholar] [CrossRef]
- Christina, M.; Laclau, J.-P.; Gonçalves, J.L.M.; Jourdan, C.; Nouvellon, Y.; Bouillet, J.-P. Almost Symmetrical Vertical Growth Rates above and below Ground in One of the World’s Most Productive Forests. Ecosphere 2011, 2, art27. [Google Scholar] [CrossRef]
- Laclau, J.-P.; Silva, E.; Rodrigues Lambais, G.; Bernoux, M.; le Maire, G.; Stape, J.L.; Bouillet, J.-P.; Gonçalves, J.L.D.M.; Jourdan, C.; Nouvellon, Y. Dynamics of Soil Exploration by Fine Roots down to a Depth of 10 m throughout the Entire Rotation in Eucalyptus grandis Plantations. Front. Plant Sci. 2013, 4, 243. [Google Scholar] [CrossRef]
- Pinheiro, R.C.; de Deus, J.C., Jr.; Nouvellon, Y.; Campoe, O.C.; Stape, J.L.; Aló, L.L.; Guerrini, I.A.; Jourdan, C.; Laclau, J.-P. A Fast Exploration of Very Deep Soil Layers by Eucalyptus Seedlings and Clones in Brazil. For. Ecol. Manag. 2016, 366, 143–152. [Google Scholar] [CrossRef]
- Mareschal, L.; Laclau, J.-P.; Nzila, J.-D.-D.; Versini, A.; Koutika, L.-S.; Mazoumbou, J.-C.; Deleporte, P.; Bouillet, J.-P.; Ranger, J. Nutrient Leaching and Deep Drainage under Eucalyptus Plantations Managed in Short Rotations after Afforestation of an African Savanna: Two 7-Year Time Series. For. Ecol. Manag. 2013, 307, 242–254. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L. On the Success and Failure of Mixed-Species Tree Plantations: Lessons Learned from a Model System of Eucalyptus Globulus and Acacia Mearnsii. For. Ecol. Manag. 2005, 209, 147–155. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L. Nutrient Cycling in a Mixed-Species Plantation of Eucalyptus Globulus and Acacia Mearnsii. Can. J. For. Res. 2005, 35, 2942–2950. [Google Scholar] [CrossRef]
- Kelty, M.J. The Role of Species Mixtures in Plantation Forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Binkley, D.; Senock, R.; Bird, S.; Cole, T.G. Twenty Years of Stand Development in Pure and Mixed Stands of Eucalyptus Saligna and Nitrogen-Fixing Facaltaria Moluccana. For. Ecol. Manag. 2003, 182, 93–102. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-Species Plantations of Eucalyptus with Nitrogen-Fixing Trees: A Review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef]
- Laclau, J.P.; Bouillet, J.P.; Gonçalves, J.L.M.; Silva, E.V.; Jourdan, C.; Cunha, M.C.S.; Moreira, M.R.; Saint-André, L.; Maquère, V.; Nouvellon, Y.; et al. Mixed-Species Plantations of Acacia mangium and Eucalyptus grandis in Brazil. 1. Growth Dynamics and Aboveground Net Primary Production. For. Ecol. Manag. 2008, 255, 3905–3917. [Google Scholar] [CrossRef]
- Bouillet, J.P.; Laclau, J.P.; Gonçalves, J.L.M.; Moreira, M.Z.; Trivelin, P.C.O.; Jourdan, C.; Silva, E.V.; Piccolo, M.C.; Tsai, S.M.; Galiana, A. Mixed-Species Plantations of Acacia mangium and Eucalyptus grandis in Brazil. 2: Nitrogen Accumulation in the Stands and Biological N2 Fixation. For. Ecol. Manag. 2008, 255, 3918–3930. [Google Scholar] [CrossRef]
- Voigtlaender, M.; Laclau, J.-P.; de Moraes Goncalves, J.L.; Piccolo, M.D.C.; Moreira, M.Z.; Nouvellon, Y.; Ranger, J.; Bouillet, J.-P. Introducing Acacia mangium Trees in Eucalyptus grandis Plantations: Consequences for Soil Organic Matter Stocks and Nitrogen Mineralization. Plant Soil 2012, 352, 99–111. [Google Scholar] [CrossRef]
- Nichols, J.D.; Bristow, M.; Vanclay, J.K. Mixed-Species Plantations: Prospects and Challenges. For. Ecol. Manag. 2006, 233, 383–390. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The Role of Plantations in Managing the World’s Forests in the Anthropocene. Front. Ecol. Environ. 2010, 8, 27–34. [Google Scholar] [CrossRef]
- Tchichelle, S.V.; Epron, D.; Mialoundama, F.; Koutika, L.S.; Harmand, J.-M.; Bouillet, J.-P.; Mareschal, L. Differences in Nitrogen Cycling and Soil Mineralisation between a Eucalypt Plantation and a Mixed Eucalypt and Acacia mangium Plantation on a Sandy Tropical Soil. South. For. J. For. Sci. 2017, 79, 1–8. [Google Scholar] [CrossRef]
- Voigtlaender, M.; Brandani, C.B.; Caldeira, D.R.M.; Tardy, F.; Bouillet, J.-P.; Gonçalves, J.L.M.; Moreira, M.Z.; Leite, F.P.; Brunet, D.; Paula, R.R.; et al. Nitrogen Cycling in Monospecific and Mixed-Species Plantations of Acacia mangium and Eucalyptus at 4 Sites in Brazil. For. Ecol. Manag. 2019, 436, 56–67. [Google Scholar] [CrossRef]
- Waithaisong, K.; Robin, A.; L’Huillery, V.; Abadie, J.; Sauvage, F.-X.; Chemardin, P.; Mareschal, L.; Bouillet, J.-P.; Laclau, J.-P.; Gonçalves, J.L.M.; et al. Organic Phosphorus Immobilization in Microbial Biomass Controls How N2-Fixing Trees Affect Phosphorus Bioavailability in Two Tropical Soils. Environ. Adv. 2022, 8, 100247. [Google Scholar] [CrossRef]
- Waithaisong, K.; Robin, A.; Mareschal, L.; Bouillet, J.-P.; Laclau, J.-P.; Deleporte, P.; Gonçalves, J.L.d.M.; Harmand, J.-M.; Plassard, C. Introducing N2-Fixing Trees (Acacia mangium) in Eucalypt Plantations Rapidly Modifies the Pools of Organic P and Low Molecular Weight Organic Acids in Tropical Soils. Sci. Total Environ. 2020, 742, 140535. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Louche, J.; Legname, E.; Duchemin, M.; Plassard, C. Pinus Pinaster Seedlings and Their Fungal Symbionts Show High Plasticity in Phosphorus Acquisition in Acidic Soils. Tree Physiol. 2009, 29, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Louche, J.; Duchemin, M.; Plassard, C. Positive Growth Response of Pinus Pinaster Seedlings in Soils Previously Subjected to Fertilization and Irrigation. For. Ecol. Manag. 2014, 318, 62–70. [Google Scholar] [CrossRef]
- Binkley, D. Bioassays of the Influence of Eucalyptus Saligna and Albizia Falcataria on Soil Nutrient Supply and Limitation. For. Ecol. Manag. 1997, 91, 229–234. [Google Scholar] [CrossRef]
- Graciano, C.; Goya, J.F.; Frangi, J.L.; Guiamet, J.J. Fertilization with Phosphorus Increases Soil Nitrogen Absorption in Young Plants of Eucalyptus grandis. For. Ecol. Manag. 2006, 236, 202–210. [Google Scholar] [CrossRef]
- Wan Juliana, W.A.; Burslem, D.F.R.P.; Swaine, M.D. Nutrient Limitation of Seedling Growth on Contrasting Soils from Pasoh Forest Reserve, Peninsular Malaysia. J. Trop. For. Sci. 2009, 21, 316–327. [Google Scholar]
- Giller, K.E.; Cadisch, G. Future Benefits from Biological Nitrogen Fixation: An Ecological Approach to Agriculture. Plant Soil. 1995, 65, 255–277. [Google Scholar] [CrossRef]
- Khanna, P.K. Nutrient Cycling under Mixed-Species Tree Systems in Southeast Asia. Agrofor. Syst. 1998, 38, 99–120. [Google Scholar] [CrossRef]
- Binkley, D.; Dunkin, K.A.; DeBell, D.; Ryan, M.G. Production and Nutrient Cycling in Mixed Plantations of Eucalyptus and Albizia in Hawaii. For. Sci. 1992, 38, 393–408. [Google Scholar] [CrossRef]
- Khanna, P.K. Comparison of Growth and Nutrition of Young Monocultures and Mixed Stands of Eucalyptus Globulus and Acacia Mearnsii. For. Ecol. Manag. 1997, 94, 105–113. [Google Scholar] [CrossRef]
- Binkley, D.; Ryan, M.G. Net Primary Production and Nutrient Cycling in Replicated Stands of Eucalyptus Saligna and Albizia Facaltaria. For. Ecol. Manag. 1998, 112, 79–85. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Khanna, P.K. Growth Dynamics in a Mixed-Species Plantation of Eucalyptus Globulus and Acacia Mearnsii. For. Ecol. Manag. 2004, 193, 81–95. [Google Scholar] [CrossRef]
- Colman, B.P.; Schimel, J.P. Drivers of Microbial Respiration and Net N Mineralization at the Continental Scale. Soil Biol. Biochem. 2013, 60, 65–76. [Google Scholar] [CrossRef]
- Tchichelle, S.V.; Mareschal, L.; Koutika, L.-S.; Epron, D. Biomass Production, Nitrogen Accumulation and Symbiotic Nitrogen Fixation in a Mixed-Species Plantation of Eucalypt and Acacia on a Nutrient-Poor Tropical Soil. For. Ecol. Manag. 2017, 403, 103–111. [Google Scholar] [CrossRef]
- Lensi, R.; Domenach, A.M.; Abbadie, L. Field Study of Nitrification and Denitrification in a Wet Savanna of West Africa (Lamto, Côte d’Ivoire). Plant Soil 1992, 147, 107–113. [Google Scholar] [CrossRef]
- Abbadie, L.; Mariotti, A.; Menaut, J.-C. Independence of Savanna Grasses from Soil Organic Matter for Their Nitrogen Supply. Ecology 1992, 73, 608–613. [Google Scholar] [CrossRef]
- Lata, J.-C.; Durand, J.; Lensi, R.; Abbadie, L. Stable Coexistence of Contrasted Nitrification Statuses in a Wet Tropical Savanna Ecosystem. Funct. Ecol. 1999, 13, 762–768. [Google Scholar] [CrossRef]
- Lata, J.-C.; Guillaume, K.; Degrange, V.; Abbadie, L.; Lensi, R. Relationships between Root Density of the African Grass Hyparrhenia Diplandra and Nitrification at the Decimetric Scale: An Inhibition–Stimulation Balance Hypothesis. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 595–600. [Google Scholar] [CrossRef]
- Rachid, C.; Balieiro, F.C.; Peixoto, R.S.; Pinheiro, Y.A.S.; Piccolo, M.C.; Chaer, G.M.; Rosado, A.S. Mixed Plantations Can Promote Microbial Integration and Soil Nitrate Increases with Changes in the N Cycling Genes. Soil Biol. Biochem. 2013, 66, 146–153. [Google Scholar] [CrossRef]
- Daly, A.B.; Jilling, A.; Bowles, T.M.; Buchkowski, R.W.; Frey, S.D.; Kallenbach, C.M.; Keiluweit, M.; Mooshammer, M.; Schimel, J.P.; Grandy, A.S. A Holistic Framework Integrating Plant-Microbe-Mineral Regulation of Soil Bioavailable Nitrogen. Biogeochemistry 2021, 154, 211–229. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, M.H.O.; Monteiro, R. Contribution of Forest Species to the Floristic Composition of a Forested Savanna in Southeastern Brazil. Braz. Arch. Biol. Technol. 2006, 49, 763–774. [Google Scholar] [CrossRef]
- Lilienfein, J.; Wilcke, W. Element Storage in Native, Agri-, and Silvicultural Ecosystems of the Brazilian Savanna. Plant Soil 2003, 254, 425–442. [Google Scholar] [CrossRef]
- Epron, D.; Nouvellon, Y.; Mareschal, L.; e Moreira, R.M.; Koutika, L.-S.; Geneste, B.; Delgado-Rojas, J.S.; Laclau, J.-P.; Sola, G.; de Moraes Gonçalves, J.L.; et al. Partitioning of Net Primary Production in Eucalyptus and Acacia Stands and in Mixed-Species Plantations: Two Case-Studies in Contrasting Tropical Environments. For. Ecol. Manag. 2013, 301, 102–111. [Google Scholar] [CrossRef]
- Versini, A. Effet de la Manipulation des Litières Aériennes sur les Cycles du C et de N dans les sols de Plantation d’Eucalyptus au Congo. Ph.D. Thesis, Université de Lorraine, Pont-à-Mousson, France, 2012; p. 221. [Google Scholar]
- FAO-UNESCO. Soil Map of the World at 1:5000000, 2nd ed.; Food and Agriculture Organization: Rome, Italy, 1989. [Google Scholar]
- Paula, R.R.; Bouillet, J.-P.; Goncalves, J.L.d.M.; Trivelin, P.C.O.; Balieiro, F.d.C.; Nouvellon, Y.; Oliveira, J.d.C.; de Deus Junior, J.C.; Bordron, B.; Laclau, J.-P. Nitrogen Fixation Rate of Acacia mangium Wild at Mid Rotation in Brazil Is Higher in Mixed Plantations with Eucalyptus grandis Hill Ex Maiden than in Monocultures. Ann. For. Sci. 2018, 75. [Google Scholar] [CrossRef]
- Hamdi, S.; Chevallier, T.; Ben Aïssa, N.; Ben Hammouda, M.; Gallali, T.; Chotte, J.-L.; Bernoux, M. Short-Term Temperature Dependence of Heterotrophic Soil Respiration after One-Month of Pre-Incubation at Different Temperatures. Soil Biol. Biochem. 2011, 43, 1752–1758. [Google Scholar] [CrossRef]
- Gonçalves, J.L.M.; Stape, J.L.; Laclau, J.-P.; Bouillet, J.-P.; Ranger, J. Assessing the Effects of Early Silvicultural Management on Long-Term Site Productivity of Fast-Growing Eucalypt Plantations: The Brazilian Experience. South. For. J. For. Sci. 2008, 70, 105–118. [Google Scholar] [CrossRef]
- Almeida, J.C.R.; Laclau, J.-P.; Gonçalves, J.L.d.M.; Ranger, J.; Saint-André, L. A Positive Growth Response to NaCl Applications in Eucalyptus Plantations Established on K-Deficient Soils. For. Ecol. Manag. 2010, 259, 1786–1795. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fertilization and Original Vegetation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| −Fert | +Fert | Two-Way ANOVA | ||||||||
| Variable | Organ | Ac | Euc | Nat | Ac | Euc | Nat | Veg | Fert | Veg × Fert |
| dwt (g/plant) | Roots | 1.05 b (0.16) | 0.56 b (0.14) | 1.14 b (0.34) | 2.17 a (0.32) | 2.60 a (0.87) | 2.57 a (0.79) | NS | *** | NS |
| Stem | 0.18 b (0.04) | 0.16 b (0.05) | 0.35 b (0.20) | 1.04 a (0.27) | 0.93 a (0.10) | 0.87 a (0.35) | NS | *** | NS | |
| Leaves | 0.41 c (0.03) | 0.39 c (0.11) | 0.66 bc (0.24) | 1.63 a (0.33) | 1.36 a (0.38) | 1.16 ab (0.47) | NS | *** | * | |
| N conc. (mg/g dwt) | Roots | 6.25 b (1.60) | 6.69 b (2.07) | 6.67 b (1.84) | 9.37 ab (0.78) | 8.79 ab (1.30) | 12.70 a (5.04) | NS | *** | NS |
| Stem | 4.42 cd (0.69) | 4.51 cd (0.52) | 3.43 d (1.42) | 8.40 bc (2.31) | 11.38 b (3.21) | 17.61 a (8.78) | *** | *** | *** | |
| Leaves | 9.83 b (2.80) | 8.56 b (2.19) | 9.60 b (4.20) | 14.15 ab (4.86) | 22.21 a (4.37) | 22.60 a (7.40) | NS | *** | NS | |
| Fertilization and Original Vegetation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| −Fert | +Fert | Two-Way ANOVA | ||||||||
| Variable | Organ | Ac | Euc | Nat | Ac | Euc | Nat | Veg | Fert | Veg × Fert |
| dwt (g/plant) | Roots | 0.73 ab (0.12) | 0.45 c (0.08) | 0.27 d (0.06) | 0.79 a (0.13) | 0.60 bc (0.14) | 0.57 bc (0.25) | *** | *** | * |
| Stem | 1.2 b (0.13) | 0.54 c (0.06) | 0.31 d (0.04) | 1.77 a (0.26) | 1.54 a (0.20) | 1.15 b (0.29) | *** | *** | ** | |
| Leaves | 2.27 b (0.16) | 1.10 c (0.16) | 0.65 d (0.05) | 3.00 a (0.48) | 2.28 b (0.60) | 2.15 b (0.37) | *** | *** | ** | |
| N conc. (mg/g dwt) | Roots | 6.82 c (1.09) | 7.48 c (1.75) | 6.92 c (1.96) | 13.71 a (1.68) | 12.97 a (2.58) | 10.03 b (2.66) | * | *** | ** |
| Stem | 2.44 c (0.68) | 2.76 c (0.98) | 3.03 bc (1.07) | 4.90 ab (2.76) | 5.09 ab (1.42) | 5.86 a (2.20) | NS | *** | NS | |
| Leaves | 9.83 b (1.54) | 9.76 b (1.56) | 9.60 b (1.32) | 22.60 a (3.75) | 22.21 a (4.60) | 14.14 ab (3.53) | NS | *** | NS | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waithaisong, K.; Robin, A.; Mareschal, L.; Bouillet, J.-P.; Harmand, J.-M.; Bordron, B.; Laclau, J.-P.; Gonçalves, J.L.M.; Plassard, C. Potential of Bioassays to Assess Consequences of Cultivation of Acacia mangium Trees on Nitrogen Bioavailability to Eucalyptus Trees: Two Case-Studies in Contrasting Tropical Soils. Plants 2023, 12, 802. https://doi.org/10.3390/plants12040802
Waithaisong K, Robin A, Mareschal L, Bouillet J-P, Harmand J-M, Bordron B, Laclau J-P, Gonçalves JLM, Plassard C. Potential of Bioassays to Assess Consequences of Cultivation of Acacia mangium Trees on Nitrogen Bioavailability to Eucalyptus Trees: Two Case-Studies in Contrasting Tropical Soils. Plants. 2023; 12(4):802. https://doi.org/10.3390/plants12040802
Chicago/Turabian StyleWaithaisong, Kittima, Agnès Robin, Louis Mareschal, Jean-Pierre Bouillet, Jean-Michel Harmand, Bruno Bordron, Jean-Paul Laclau, José Leonardo Moraes Gonçalves, and Claude Plassard. 2023. "Potential of Bioassays to Assess Consequences of Cultivation of Acacia mangium Trees on Nitrogen Bioavailability to Eucalyptus Trees: Two Case-Studies in Contrasting Tropical Soils" Plants 12, no. 4: 802. https://doi.org/10.3390/plants12040802
APA StyleWaithaisong, K., Robin, A., Mareschal, L., Bouillet, J.-P., Harmand, J.-M., Bordron, B., Laclau, J.-P., Gonçalves, J. L. M., & Plassard, C. (2023). Potential of Bioassays to Assess Consequences of Cultivation of Acacia mangium Trees on Nitrogen Bioavailability to Eucalyptus Trees: Two Case-Studies in Contrasting Tropical Soils. Plants, 12(4), 802. https://doi.org/10.3390/plants12040802