SlIAA9 Mutation Maintains Photosynthetic Capabilities under Heat-Stress Conditions

 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Results
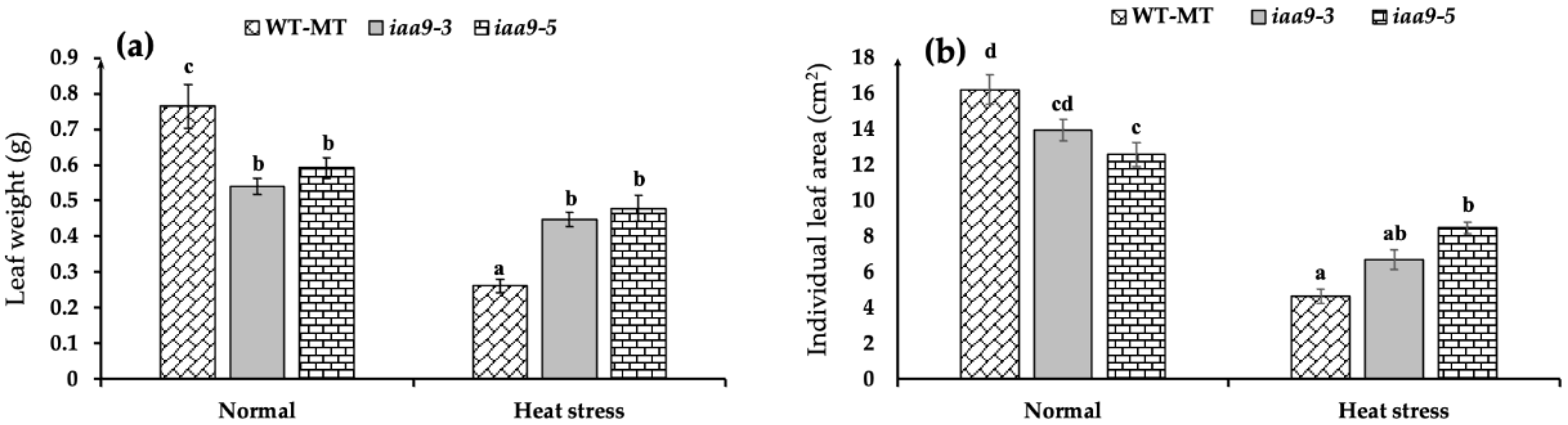
2.1. IAA9 Mutants Have Wider Individual Leaf Area and Heavier Leaf Weight under Heat Stress Conditions
2.2. IAA9 Mutants Have Higher Net Photosynthetic Rate, Stomatal Conductance, and Water Use Efficiency under Heat Stress Conditions
2.3. IAA9 Mutants Have Higher Leaf Chlorophyll and Carotenoid Content
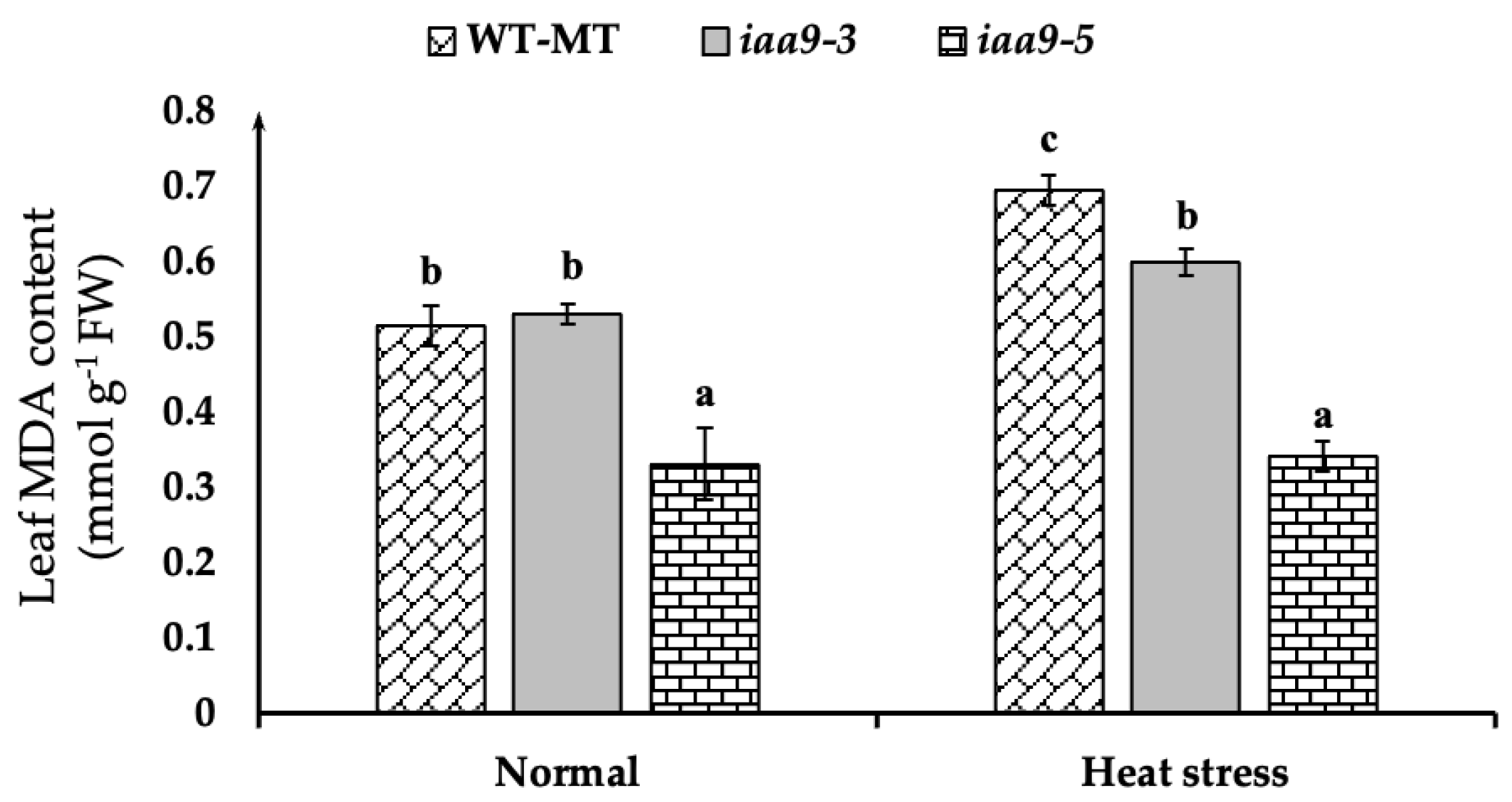
2.4. IAA9 Mutants Have Lower Leaf Malondialdehyde (MDA) Content during Heat Stress
2.5. IAA9 Mutants Shown Better Chlorophyll Fluorescence under Heat Stress Conditions
2.6. Multivariate Analysis of Analyzed Trait under Heat Stress Conditions
3. Discussion
4. Materials and Methods
4.1. Growth Condition and Plant Material
4.2. Individual Leaf Area and Leaf Weight
4.3. Plant Photosynthetic Rate, Stomatal Conductance, and Leaf Temperature
4.4. Leaf Water Use Efficiency
4.5. Total Leaf Chlorophyll and Total Carotenoid Content
4.6. Leaf MDA Content
4.7. Chlorophyll a Fluorescence
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Combet, E.; Jarlot, A.; Aidoo, K.E.; Lean, M.E. Development of a Nutritionally Balanced Pizza as a Functional Meal Designed to Meet Published Dietary Guidelines. Public Health Nutr. 2014, 17, 2577–2586. [Google Scholar] [CrossRef] [PubMed]
- Kamanga, R.M.; Mbega, E.; Ndakidemi, P. Drought Tolerance Mechanisms in Plants: Physiological Responses Associated with Water Deficit Stress in Solanum Lycopersicum. Adv. Crop Sci. Technol. 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Rajametov, S.N.; Yang, E.Y.; Jeong, H.B.; Cho, M.C.; Chae, S.Y.; Paudel, N. Heat Treatment in Two Tomato Cultivars: A Study of the Effect on Physiological and Growth Recovery. Horticulturae 2021, 7, 119. [Google Scholar] [CrossRef]
- Collins, M.; Knutti, R.; Arblaster, J.; Dufresne, J.-L.; Fichefet, T.; Friedlingstein, P.; Gao, X.; Gutowski, W.J.; Johns, T.; Krinner, G. Long-Term Climate Change: Projections, Commitments and Irreversibility. In Climate Change 2013-The Physical Science Basis: Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; pp. 1029–1136. [Google Scholar]
- Silva, R.S.; Kumar, L.; Shabani, F.; Picanço, M.C. Assessing the Impact of Global Warming on Worldwide Open Field Tomato Cultivation through CSIRO-Mk3·0 Global Climate Model. J. Agric. Sci. 2017, 155, 407–420. [Google Scholar] [CrossRef]
- Lohar, D.P.; Peat, W.E. Floral Characteristics of Heat-Tolerant and Heat-Sensitive Tomato (Lycopersicon esculentum Mill.) Cultivars at High Temperature. Sci. Hortic. 1998, 73, 53–60. [Google Scholar] [CrossRef]
- Sato, S.; Peet, M.M.; Thomas, J.F. Physiological Factors Limit Fruit Set of Tomato (Lycopersicon esculentum Mill.) under Chronic, Mild Heat Stress. Plant Cell Environ. 2000, 23, 719–726. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Angeles Morales, M.; Miguel Dell’Amico, J.; Torrecillas, A.; Alarcón, J.J. High Temperature Effects on Photosynthetic Activity of Two Tomato Cultivars with Different Heat Susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef]
- Qi, M.; Liu, Y.; Li, T. Nano-TiO2 Improve the Photosynthesis of Tomato Leaves under Mild Heat Stress. Biol. Trace Elem. Res. 2013, 156, 323–328. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Kjær, K.H.; Rosenqvist, E.; Ottosen, C.-O.; Wu, Z. Screening and Validation of Tomato Genotypes under Heat Stress Using Fv/Fm to Reveal the Physiological Mechanism of Heat Tolerance. Environ. Exp. Bot. 2015, 118, 1–11. [Google Scholar] [CrossRef]
- Poudyal, D.; Rosenqvist, E.; Ottosen, C.-O. Phenotyping from Lab to Field—Tomato Lines Screened for Heat Stress Using Fv/Fm Maintain High Fruit Yield during Thermal Stress in the Field. Funct. Plant Biol. 2019, 46, 44. [Google Scholar] [CrossRef]
- Hemantaranjan, A. Heat Stress Responses and Thermotolerance. Adv. Plants Agric. Res. 2014, 1, 62–70. [Google Scholar] [CrossRef]
- Mir, A.R.; Siddiqui, H.; Alam, P.; Hayat, S. Foliar Spray of Auxin/IAA Modulates Photosynthesis, Elemental Composition, ROS Localization and Antioxidant Machinery to Promote Growth of Brassica Juncea. Physiol. Mol. Biol. Plants 2020, 26, 2503–2520. [Google Scholar] [CrossRef]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Soliman, W.S.; Fujimori, M.; Tase, K.; Sugiyama, S. Oxidative Stress and Physiological Damage under Prolonged Heat Stress in C3 Grass Lolium Perenne. Grassl. Sci. 2011, 57, 101–106. [Google Scholar] [CrossRef]
- Bielach, A.; Hrtyan, M.; Tognetti, V. Plants under Stress: Involvement of Auxin and Cytokinin. Int. J. Mol. Sci. 2017, 18, 1427. [Google Scholar] [CrossRef]
- Tognetti, V.B.; Bielach, A.; Hrtyan, M. Redox Regulation at the Site of Primary Growth: Auxin, Cytokinin and ROS Crosstalk. Plant Cell Environ. 2017, 40, 2586–2605. [Google Scholar] [CrossRef]
- Tognetti, V.B.; Mühlenbock, P.; van Breusegem, F. Stress Homeostasis—The Redox and Auxin Perspective. Plant Cell Environ. 2012, 35, 321–333. [Google Scholar] [CrossRef]
- Singh, H.; Bhat, J.A.; Singh, V.P.; Corpas, F.J.; Yadav, S.R. Auxin Metabolic Network Regulates the Plant Response to Metalloids Stress. J. Hazard. Mater. 2021, 405, 124250. [Google Scholar] [CrossRef]
- Suminar, E.; Budiarto, R.; Nuraini, A.; Mubarok, S.; Ezura, H. Morpho-Physiological Responses of Iaa9 Tomato Mutants to Different Levels of PEG Simulated Drought Stress. Biodiversitas 2022, 23, 3115–3126. [Google Scholar] [CrossRef]
- Tyburski, J.; Tretyn, A. Ascorbate and Glutathione in Organogenesis, Regeneration and Differentiation in Plant In Vitro Cultures. In Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Springer Netherlands: Dordrecht, The Netherlands, 2010; pp. 55–90. [Google Scholar]
- Tyburski, J.; Dunajska, K.; Mazurek, P.; Piotrowska, B.; Tretyn, A. Exogenous Auxin Regulates H2O2 Metabolism in Roots of Tomato (Lycopersicon esculentum Mill.) Seedlings Affecting the Expression and Activity of CuZn-Superoxide Dismutase, Catalase, and Peroxidase. Acta Physiol. Plant. 2009, 31, 249–260. [Google Scholar] [CrossRef]
- Kim, J.I.; Baek, D.; Park, H.C.; Chun, H.J.; Oh, D.-H.; Lee, M.K.; Cha, J.-Y.; Kim, W.-Y.; Kim, M.C.; Chung, W.S.; et al. Overexpression of Arabidopsis YUCCA6 in Potato Results in High-Auxin Developmental Phenotypes and Enhanced Resistance to Water Deficit. Mol. Plant 2013, 6, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Song; Fan; Jiao; Liu; Wang; Jia Overexpression of OsPT8 Increases Auxin Content and Enhances Tolerance to High-Temperature Stress in Nicotiana Tabacum. Genes 2019, 10, 809. [CrossRef] [PubMed]
- Esmaeili, N.; Yang, X.; Cai, Y.; Sun, L.; Zhu, X.; Shen, G.; Payton, P.; Fang, W.; Zhang, H. Co-Overexpression of AVP1 and OsSIZ1 in Arabidopsis Substantially Enhances Plant Tolerance to Drought, Salt, and Heat Stresses. Sci. Rep. 2019, 9, 7642. [Google Scholar] [CrossRef] [PubMed]
- Pantoja-Benavides, A.D.; Garces-Varon, G.; Restrepo-Díaz, H. Foliar Growth Regulator Sprays Induced Tolerance to Combined Heat Stress by Enhancing Physiological and Biochemical Responses in Rice. Front. Plant Sci. 2021, 12, 702892. [Google Scholar] [CrossRef]
- Saito, T.; Ariizumi, T.; Okabe, Y.; Asamizu, E.; Hiwasa-Tanase, K.; Fukuda, N.; Mizoguchi, T.; Yamazaki, Y.; Aoki, K.; Ezura, H. TOMATOMA: A Novel Tomato Mutant Database Distributing Micro-Tom Mutant Collections. Plant Cell Physiol. 2011, 52, 283–296. [Google Scholar] [CrossRef]
- Shikata, M.; Ezura, H. Micro-Tom Tomato as an Alternative Plant Model System: Mutant Collection and Efficient Transformation. In Plant Signal Transduction; Humana Press: New York, NY, USA, 2016; pp. 47–55. [Google Scholar]
- Fujita, K.; Horiuchi, H.; Takato, H.; Kohno, M.; Suzuki, S. Auxin-Responsive Grape Aux/IAA9 Regulates Transgenic Arabidopsis Plant Growth. Mol. Biol. Rep. 2012, 39, 7823–7829. [Google Scholar] [CrossRef]
- Ariizumi, T.; Shinozaki, Y.; Ezura, H. Genes That Influence Yield in Tomato. Breed. Sci. 2013, 63, 3–13. [Google Scholar] [CrossRef]
- Wang, H.; Jones, B.; Li, Z.; Frasse, P.; Delalande, C.; Regad, F.; Chaabouni, S.; Latché, A.; Pech, J.-C.; Bouzayen, M. The Tomato Aux/IAA Transcription Factor IAA9 Is Involved in Fruit Development and Leaf Morphogenesis. Plant Cell 2005, 17, 2676–2692. [Google Scholar] [CrossRef]
- Efendi, D.; Budiarto, R. Benefit and Challenges of Using Tropical Fruits as Ornamental Trees for Green City. Acta Hortic. 2022, 1334, 369–378. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D. Morphological Evaluation and Determination Keys of 21 Citrus Genotypes at Seedling Stage. Biodiversitas 2021, 22, 1570–1579. [Google Scholar] [CrossRef]
- Hamdani, J.; Zahra, T.A.; Nuraini, A.; Budiarto, R. Production of G0 ‘Median’ Potato on Different Media Composition and Fertilizer Method. Asian J. Plant Sci. 2022, 21, 312–320. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D. Shoot Manipulations Improve Flushing and Flowering of Mandarin Citrus in Indonesia. J. Appl. Hortic. 2018, 20, 112–118. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Agronomical and Physiological Characters of Kaffir Lime (Citrus hystrix DC) Seedling under Artificial Shading and Pruning. Emir. J. Food Agric. 2019, 31, 222–230. [Google Scholar] [CrossRef]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The Relationship between Leaf Area Growth and Biomass Accumulation in Arabidopsis Thaliana. Front. Plant Sci. 2015, 6, 167. [Google Scholar] [CrossRef] [PubMed]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Comparative and Correlation Analysis of Young and Mature Kaffir Lime (Citrus Hystrix DC) Leaf Characteristics. Int. J. Plant Biol. 2022, 13, 270–280. [Google Scholar] [CrossRef]
- Budiarto, R.; Poerwanto, R.; Santosa, E.; Efendi, D.; Agusta, A. Short Communication: Allometric Model to Estimate Bifoliate Leaf Area and Weight of Kaffir Lime (Citrus Hystrix). Biodiversitas 2021, 22, 2815–2820. [Google Scholar] [CrossRef]
- Mubarok, S.; Wicaksono, F.Y.; Nuraini, A.; Rahmat, B.P.N.; Budiarto, R. Short Communication: Agronomical Characteristics of Sweet Corn under Different Plant Growth Regulators during the Dry Season. Biodiversitas 2022, 23. [Google Scholar] [CrossRef]
- Lestari, W.; Mubarok, S.; Nuraini, A. Respons Anatomis Dan Morfofisiologis Mutan Tomat Iaa9 (Iaa9-3 Dan Iaa9-5) Pada Kondisi Cekaman Suhu Tinggi (Heat Stress); Universitas Padjajaran: Sumedang, Indonesia, 2020. [Google Scholar]
- Naz, A.A.; Raman, S.; Martinez, C.C.; Sinha, N.R.; Schmitz, G.; Theres, K. Trifoliate Encodes an MYB Transcription Factor That Modulates Leaf and Shoot Architecture in Tomato. Proc. Natl. Acad. Sci. USA 2013, 110, 2401–2406. [Google Scholar] [CrossRef] [PubMed]
- Abdelmageed, A.H.A.; Gruda, N. Influence of High Temperatures on Gas Exchange Rate and Growth of Eight Tomato Cultivars under Controlled Heat Stress Conditions. Eur. J. Hortic. Sci. 2009, 74, 152. [Google Scholar]
- Abdellatif, I.M.Y.; Yuan, S.; Na, R.; Yoshihara, S.; Hamada, H.; Suzaki, T.; Ezura, H.; Miura, K. Functional Characterization of Tomato Phytochrome A and B1B2 Mutants in Response to Heat Stress. Int. J. Mol. Sci. 2022, 23, 1681. [Google Scholar] [CrossRef]
- von Caemmerer, S.; Farquhar, G.D. Some Relationships between the Biochemistry of Photosynthesis and the Gas Exchange of Leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Nankishore, A.; Farrell, A.D. The Response of Contrasting Tomato Genotypes to Combined Heat and Drought Stress. J. Plant Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Feller, U. Drought Stress and Carbon Assimilation in a Warming Climate: Reversible and Irreversible Impacts. J. Plant Physiol. 2016, 203, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y. Cytokinin and Auxin Inhibit Abscisic Acid-Induced Stomatal Closure by Enhancing Ethylene Production in Arabidopsis. J. Exp. Bot. 2006, 57, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Hayat, Q.; Hayat, S.; Ali, B.; Ahmad, A. Auxin Analogues and Nitrogen Metabolism, Photosynthesis, and Yield of Chickpea. J. Plant Nutr. 2009, 32, 1469–1485. [Google Scholar] [CrossRef]
- Netto, A.T.; Campostrini, E.; de Oliveira, J.G.; Bressan-Smith, R.E. Photosynthetic Pigments, Nitrogen, Chlorophyll a Fluorescence and SPAD-502 Readings in Coffee Leaves. Sci. Hortic. 2005, 104, 199–209. [Google Scholar] [CrossRef]
- Dao, G.-H.; Wu, G.-X.; Wang, X.-X.; Zhuang, L.-L.; Zhang, T.-Y.; Hu, H.-Y. Enhanced Growth and Fatty Acid Accumulation of Microalgae Scenedesmus Sp. LX1 by Two Types of Auxin. Bioresour. Technol. 2018, 247, 561–567. [Google Scholar] [CrossRef]
- Yuan, Y.; Mei, L.; Wu, M.; Wei, W.; Shan, W.; Gong, Z.; Zhang, Q.; Yang, F.; Yan, F.; Zhang, Q.; et al. SlARF10, an Auxin Response Factor, Is Involved in Chlorophyll and Sugar Accumulation during Tomato Fruit Development. J. Exp. Bot. 2018, 69, 5507–5518. [Google Scholar] [CrossRef]
- Ivanchenko, M.G.; den Os, D.; Monshausen, G.B.; Dubrovsky, J.G.; Bednářová, A.; Krishnan, N. Auxin Increases the Hydrogen Peroxide (H2O2) Concentration in Tomato (Solanum lycopersicum) Root Tips While Inhibiting Root Growth. Ann. Bot. 2013, 112, 1107–1116. [Google Scholar] [CrossRef]
- Ait Bihi, M.; Ain-Lhout, F.; Hatimi, A.; Fahmi, F.; Tahrouch, S. Ecophysiological Response and Morphological Adjustment of Argania Spinosa L. Skeels under Contrasting Climates: Case Study of Marginal Populations. Int. J. Plant Biol. 2022, 12, 9404. [Google Scholar] [CrossRef]
- Shah, S.; Houborg, R.; McCabe, M. Response of Chlorophyll, Carotenoid and SPAD-502 Measurement to Salinity and Nutrient Stress in Wheat (Triticum aestivum L.). Agronomy 2017, 7, 61. [Google Scholar] [CrossRef]
- Akter, S.; Khan, M.S.; Smith, E.N.; Flashman, E. Measuring ROS and Redox Markers in Plant Cells. RSC Chem. Biol. 2021, 2, 1384–1401. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Jiang, F.; Wen, J.; Wu, Z. Overexpression of Solanum Habrochaites MicroRNA319d (Sha-MiR319d) Confers Chilling and Heat Stress Tolerance in Tomato (S. lycopersicum). BMC Plant Biol. 2019, 19, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lestari, F.W.; Suminar, E.; Nuraini, A.; Ezura, H.; Mubarok, S. Perubahan Viabilitas Pollen Dan Anatomi Stomata Pada Dua Mutan Tomat, Iaa9-3 Dan Iaa9-5, Akibat Cekaman Suhu Tinggi. Agrikultura 2020, 31, 25. [Google Scholar] [CrossRef]
- Gao, J.; Guo, G.; Guo, Y.; Wang, X. Measuring Plant Leaf Area by Scanner and ImageJ Software. China Veg. 2011, 2, 73–77. [Google Scholar]
- Hendry, G.A.; Grime, J.P. Methods in Comparative Plant Ecology: A Laboratory Manual; Springer Science & Business Media: Berlin, Germany, 1993; ISBN 0412462303. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CHL | CR | MDA | CI | SC | WUE | LT | PN | LW | LA | |
|---|---|---|---|---|---|---|---|---|---|---|
| CHL | 0.97 | |||||||||
| CR | 0.86 | 0.97 | ||||||||
| MDA | −0.89 | −0.78 | 0.98 | |||||||
| CI | 0.89 | 0.71 | −0.84 | 0.99 | ||||||
| SC | 0.65 | 0.70 | −0.51 | 0.47 | 1.00 | |||||
| WUE | 0.64 | 0.38 | −0.31 | 0.52 | 0.36 | 0.97 | ||||
| LT | −0.93 | −0.81 | 0.74 | −0.80 | −0.68 | −0.69 | 0.97 | |||
| PN | 0.88 | 0.77 | −0.87 | 0.78 | 0.58 | 0.59 | −0.89 | 0.98 | ||
| LW | 0.92 | 0.80 | −0.74 | 0.85 | 0.66 | 0.65 | −0.97 | 0.93 | 0.94 | |
| LA | 0.88 | 0.84 | −0.89 | 0.79 | 0.54 | 0.40 | −0.87 | 0.92 | 0.79 | 0.97 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmat, B.P.N.; Octavianis, G.; Budiarto, R.; Jadid, N.; Widiastuti, A.; Matra, D.D.; Ezura, H.; Mubarok, S. SlIAA9 Mutation Maintains Photosynthetic Capabilities under Heat-Stress Conditions. Plants 2023, 12, 378. https://doi.org/10.3390/plants12020378
Rahmat BPN, Octavianis G, Budiarto R, Jadid N, Widiastuti A, Matra DD, Ezura H, Mubarok S. SlIAA9 Mutation Maintains Photosynthetic Capabilities under Heat-Stress Conditions. Plants. 2023; 12(2):378. https://doi.org/10.3390/plants12020378
Chicago/Turabian StyleRahmat, Bayu Pradana Nur, Grace Octavianis, Rahmat Budiarto, Nurul Jadid, Ani Widiastuti, Deden Derajat Matra, Hiroshi Ezura, and Syariful Mubarok. 2023. "SlIAA9 Mutation Maintains Photosynthetic Capabilities under Heat-Stress Conditions" Plants 12, no. 2: 378. https://doi.org/10.3390/plants12020378
APA StyleRahmat, B. P. N., Octavianis, G., Budiarto, R., Jadid, N., Widiastuti, A., Matra, D. D., Ezura, H., & Mubarok, S. (2023). SlIAA9 Mutation Maintains Photosynthetic Capabilities under Heat-Stress Conditions. Plants, 12(2), 378. https://doi.org/10.3390/plants12020378