

Identification of Newer Stable Genetic Sources for High Grain Number per Panicle and Understanding the Gene Action for Important Panicle Traits in Rice

 , and
, and
Abstract
1. Introduction
2. Results
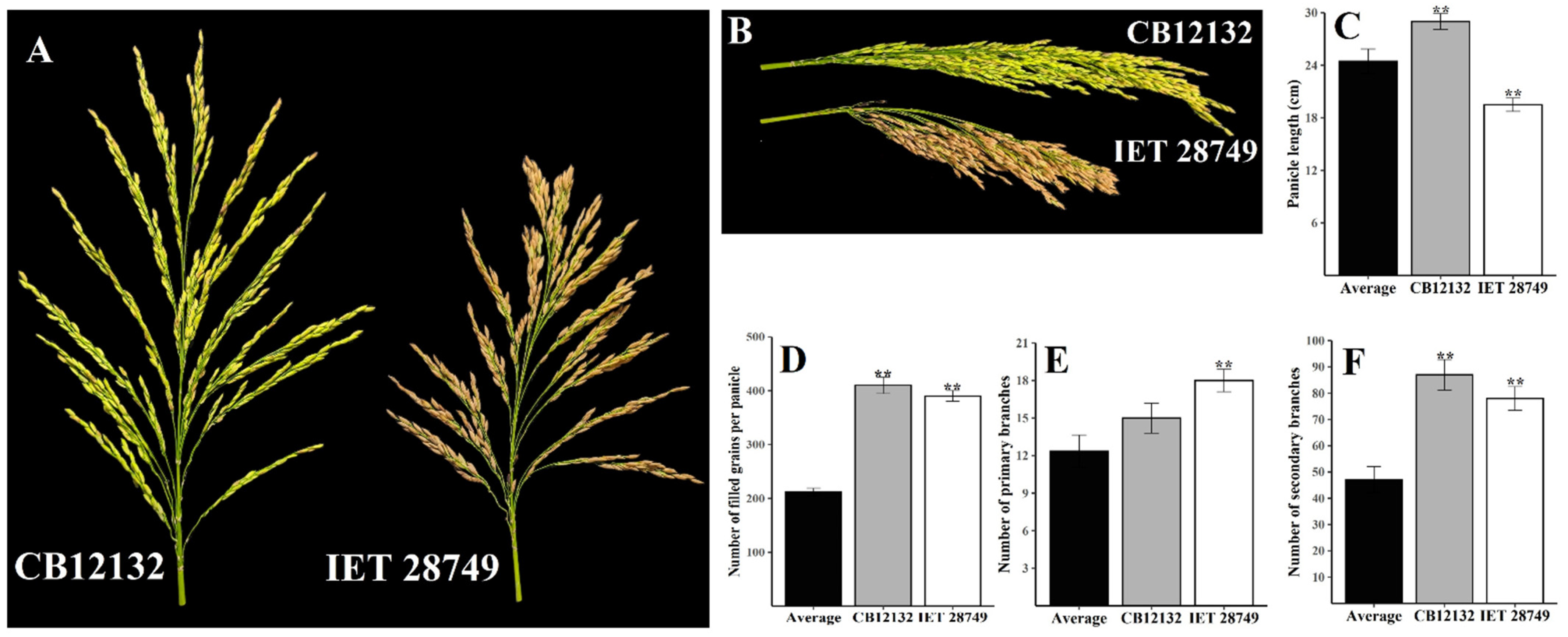
2.1. Per Se Performance and Genetic Variability
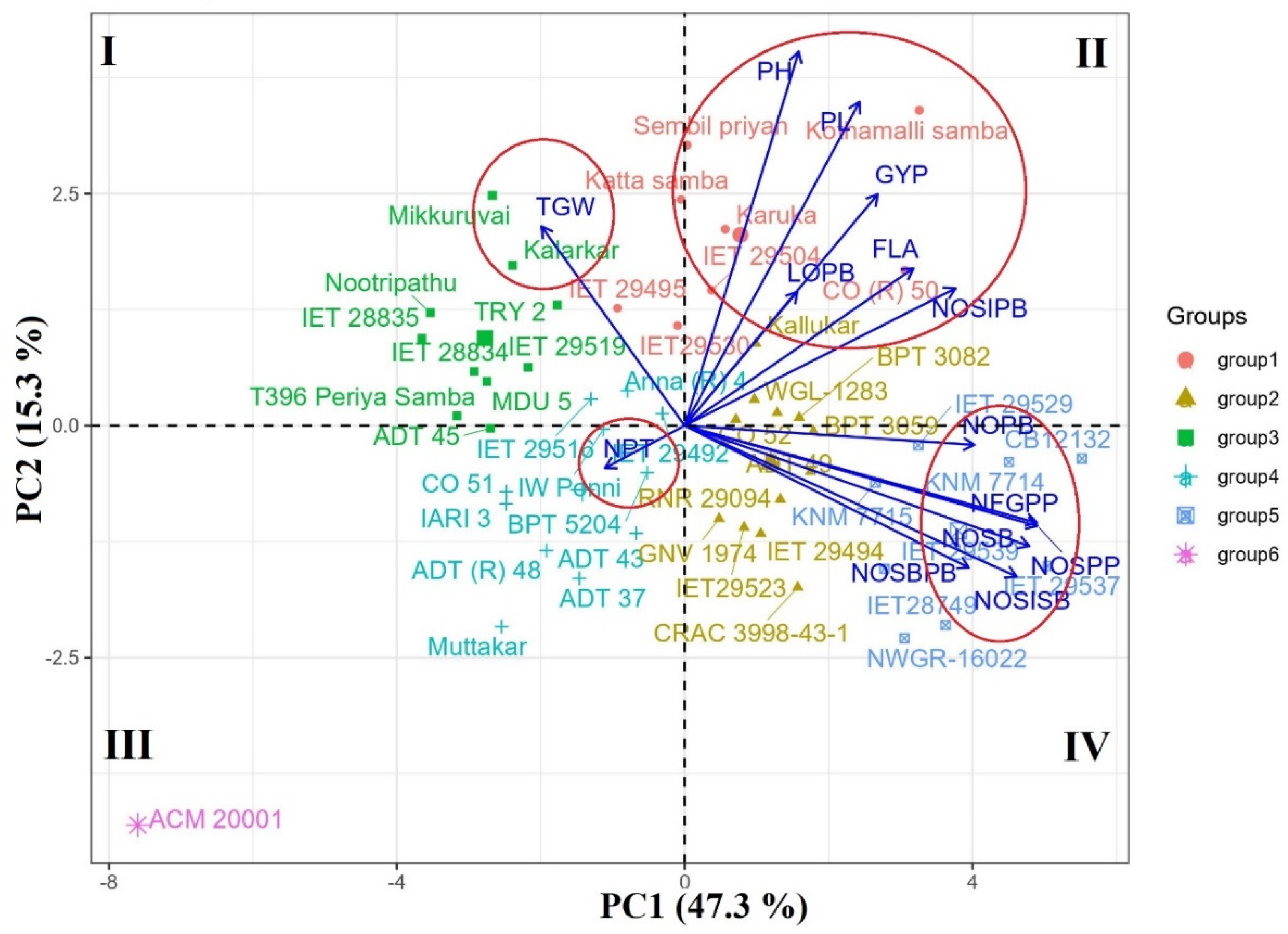
2.2. Genetic Diversity and Principal Component Analysis
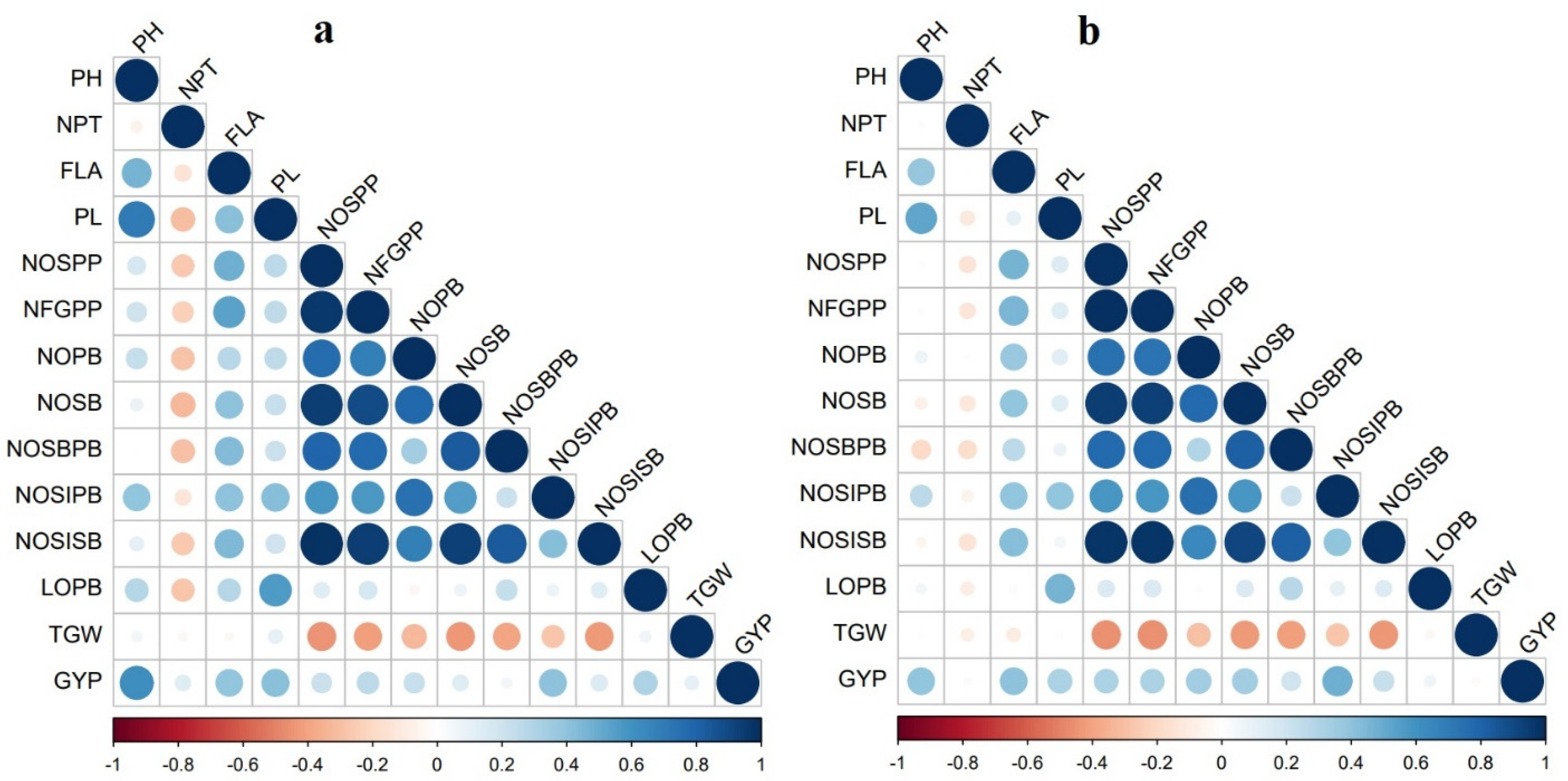
2.3. Correlation among Panicle- and Yield-Related Traits
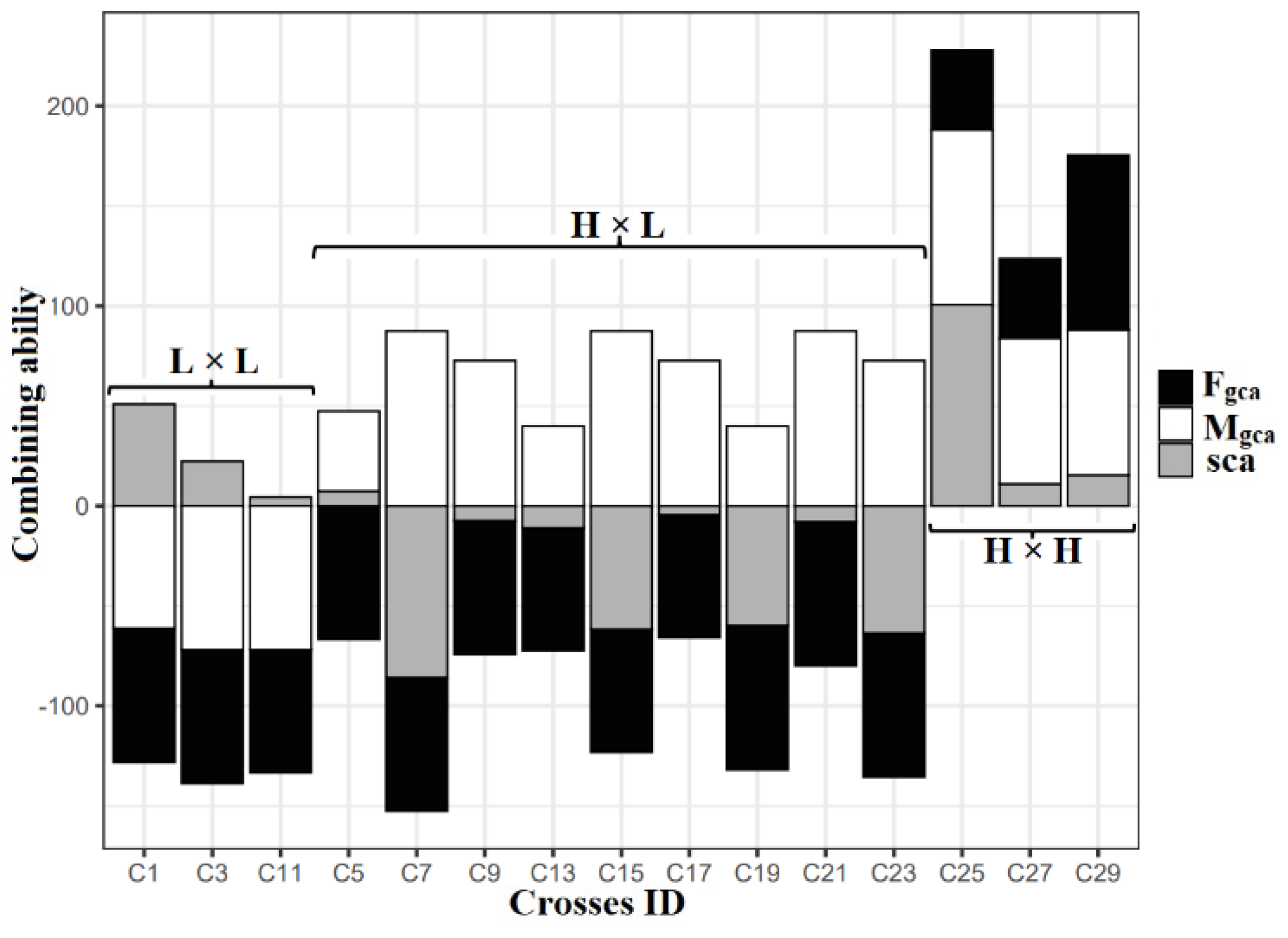
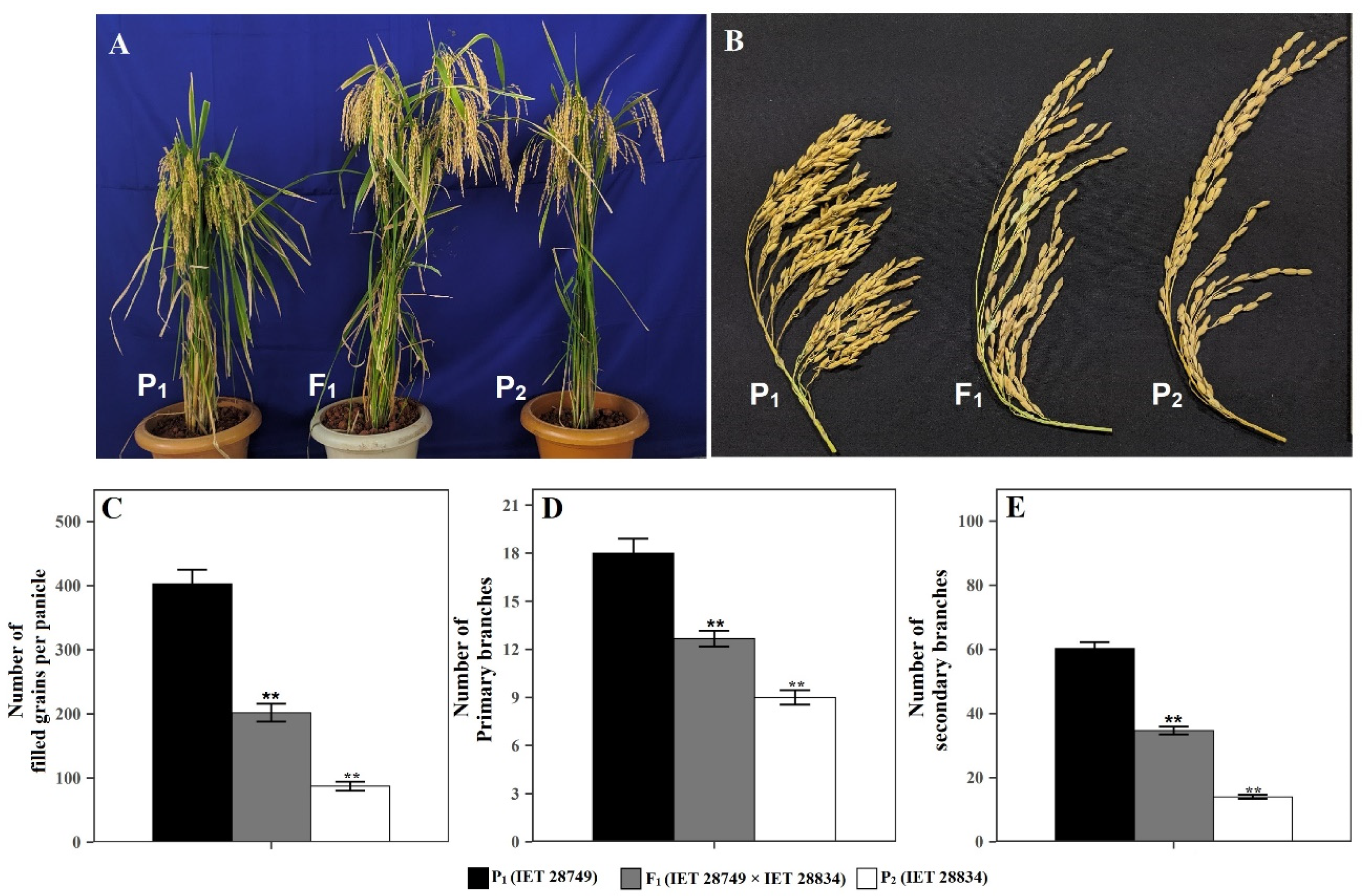
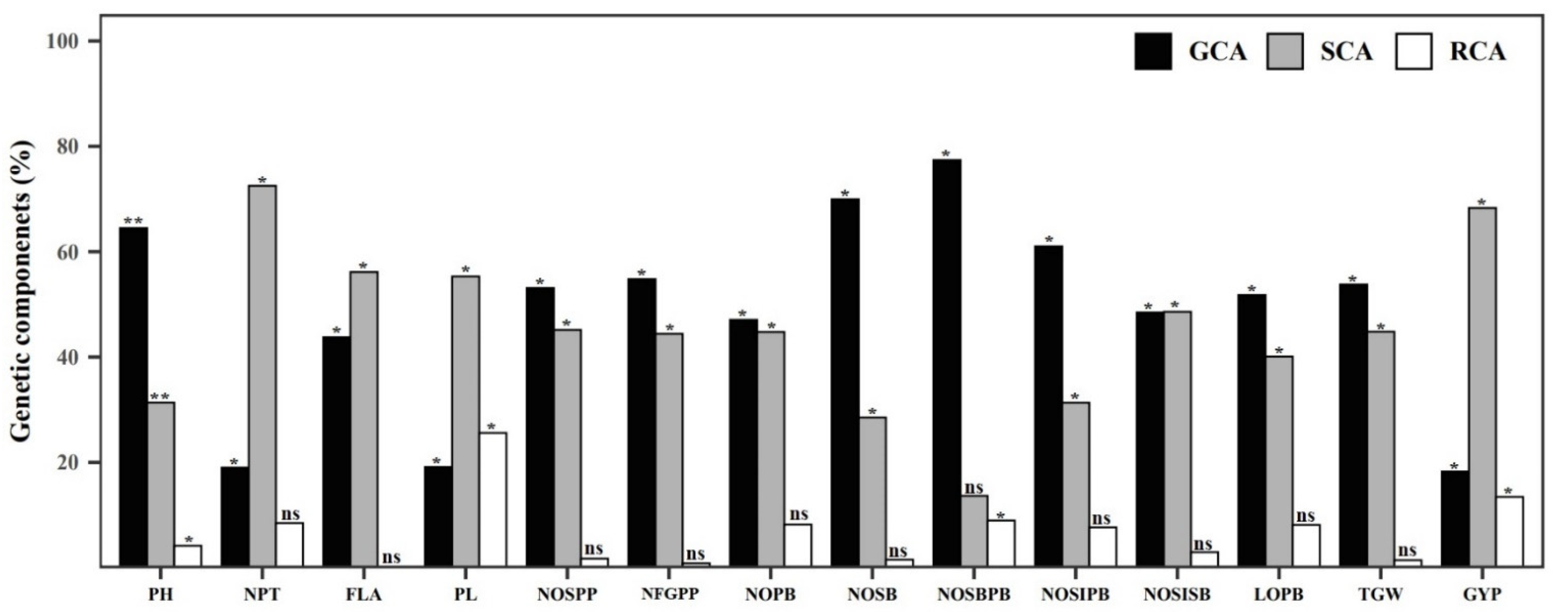
2.4. Diallel Analysis
3. Discussion
Diallel Analysis
4. Materials and Methods
4.1. Plant Genetic Material and Experimental Site
4.2. Experiments 1 and 2
4.3. Evaluation of Morphological Traits
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carcea, M. Value of wholegrain rice in a healthy human nutrition. Agriculture 2021, 11, 720. [Google Scholar] [CrossRef]
- Seck, P.A.; Diagne, A.; Mohanty, S.; Wopereis, M. Crops that feed the world 7: Rice. Food Secur. 2012, 4, 7–24. [Google Scholar] [CrossRef]
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for global nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Rao, I.; Rao, H.S.; Vani, C.S. Sources of growth in rice production in India: State wise component analysis. Int. J. Agric. Environ. Biotechnol. 2018, 11, 121–125. [Google Scholar]
- Xie, J.; Li, F.; Khan, N.U.; Zhu, X.; Wang, X.; Zhang, Z.; Ma, X.; Zhao, Y.; Zhang, Q.; Zhang, S. Identifying natural genotypes of grain number per panicle in rice (Oryza sativa L.) by association mapping. Genes Genom. 2019, 41, 283–295. [Google Scholar] [CrossRef]
- Ikeda, M.; Hirose, Y.; Takashi, T.; Shibata, Y.; Yamamura, T.; Komura, T.; Doi, K.; Ashikari, M.; Matsuoka, M.; Kitano, H. Analysis of rice panicle traits and detection of QTLs using an image analyzing method. Breed. Sci. 2010, 60, 55–64. [Google Scholar] [CrossRef]
- Zhong, H.; Liu, S.; Meng, X.; Sun, T.; Deng, Y.; Kong, W.; Peng, Z.; Li, Y. Uncovering the genetic mechanisms regulating panicle architecture in rice with GPWAS and GWAS. BMC Genom. 2021, 22, 86. [Google Scholar]
- Dhakal, A.; Pokhrel, A.; Sharma, S.; Poudel, A. Multivariate analysis of phenotypic diversity of rice (Oryza sativa L.) landraces from Lamjung and Tanahun Districts, Nepal. Int. J. Agron. 2020, 2020, 8867961. [Google Scholar] [CrossRef]
- Singh, H.P.; Raigar, O.P.; Chahota, R.K. Estimation of genetic diversity and its exploitation in plant breeding. Bot. Rev. 2022, 88, 413–435. [Google Scholar] [CrossRef]
- Tiwari, S.; Singh, Y.; Upadhyay, P.; Koutu, G. Principal component analysis and genetic divergence studies for yield and quality-related attributes of rice restorer lines. Indian J. Genet. Plant Breed. 2022, 82, 94–98. [Google Scholar]
- Lakshmi, V.I.; Sreedhar, M.; Vanisri, S.; Anantha, M.; Rao, L.S.; Gireesh, C. Multivariate analysis and selection criteria for identification of African rice (Oryza glaberrima) for genetic improvement of indica rice cultivars. Plant Genet. Resour. 2019, 17, 499–505. [Google Scholar] [CrossRef]
- Bassuony, N.N.; Zsembeli, J. Inheritance of some flag leaf and yield characteristics by half-diallel analysis in rice crops (Oryza Sativa L.). Cereal Res. Commun. 2021, 49, 503–510. [Google Scholar] [CrossRef]
- Ben Hassen, M.; Cao, T.-V.; Bartholome, J.; Orasen, G.; Colombi, C.; Rakotomalala, J.; Razafinimpiasa, L.; Bertone, C.; Biselli, C.; Volante, A. Rice diversity panel provides accurate genomic predictions for complex traits in the progenies of biparental crosses involving members of the panel. Theor. Appl. Genet. 2018, 131, 417–435. [Google Scholar] [CrossRef]
- Haghighi Hasanalideh, A.; Farshadfar, E.; Allahgholipour, M. Genetic Analysis and Heterosis for Viscosity Parameters in Rice (Oryza sativa L.) through North Carolina III Mating Design. Plant Genet. Res. 2020, 6, 129–140. [Google Scholar] [CrossRef]
- Kour, A.; Kumar, B.; Singh, B. Genetic evaluation of yield and yield attributing traits in rice (Oryza sativa L.) using line x tester analysis. Electron. J. Plant Breed. 2019, 10, 39–46. [Google Scholar] [CrossRef]
- Yao, W.; Zhang, Y.; Kang, M.; Chen, H.; Liu, L.; Yu, L.; Fan, X. Diallel analysis models: A comparison of certain genetic statistics. Crop Sci. 2013, 53, 1481–1490. [Google Scholar] [CrossRef]
- Raj, A.C.; Sharangi, A.B.; Das, A.; Pramanik, K.; Upadhyay, T.K.; Almutairi, M.; Khan, M.I.; Ahmad, I.; Kausar, M.A.; Saeed, M. Assessing the genetic divergence of onion (Allium Cepa L.) through morpho-physiological and molecular markers. Sustainability 2022, 14, 1131. [Google Scholar] [CrossRef]
- Madahana, S.L.; Owuoche, J.O.; Oyoo, M.E.; Macharia, G.K.; Randhawa, M.S. Evaluation of Kenya Stem Rust Observation Nursery Wheat Genotypes for Yield and Yield Components under Artificial Rust Conditions. Agronomy 2021, 11, 2394. [Google Scholar] [CrossRef]
- Guo, T.; Chen, K.; Dong, N.-Q.; Shi, C.-L.; Ye, W.-W.; Gao, J.-P.; Shan, J.-X.; Lin, H.-X. GRAIN SIZE AND NUMBER1 negatively regulates the OsMKKK10-OsMKK4-OsMPK6 cascade to coordinate the trade-off between grain number per panicle and grain size in rice. Plant Cell 2018, 30, 871–888. [Google Scholar] [CrossRef]
- Xu, Z.; Miao, Y.; Chen, Z.; Gao, H.; Wang, R.; Zhao, D.; Zhang, B.; Zhou, Y.; Tang, S.; Zhang, H. Identification and fine mapping of qGN1c, a QTL for grain number per panicle, in rice (Oryza sativa). Mol. Breed. 2019, 39, 129. [Google Scholar] [CrossRef]
- Niu, X.; Zhu, Y.; Sun, Z.; Yu, S.; Zhuang, J.; Fan, Y. Identification and validation of quantitative trait loci for grain number in rice (Oryza sativa L.). Agronomy 2020, 10, 180. [Google Scholar] [CrossRef]
- Wang, S.-S.; Chen, R.-K.; Chen, K.-Y.; Liu, C.-Y.; Kao, S.-M.; Chung, C.-L. Genetic mapping of the qSBN7 locus, a QTL controlling secondary branch number per panicle in rice. Breed. Sci. 2017, 67, 17007. [Google Scholar] [CrossRef] [PubMed]
- Adriani, D.E.; Dingkuhn, M.; Dardou, A.; Adam, H.; Luquet, D.; Lafarge, T. Rice panicle plasticity in Near Isogenic Lines carrying a QTL for larger panicle is genotype and environment dependent. Rice 2016, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A. Varietal differences in tiller and panicle development determining the total number of spikelets per unit area in rice. Plant Prod. Sci. 2019, 22, 192–201. [Google Scholar] [CrossRef]
- Malik, N.; Ranjan, R.; Parida, S.K.; Agarwal, P.; Tyagi, A.K. Mediator subunit OsMED14_1 plays an important role in rice development. Plant J. 2020, 101, 1411–1429. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Chu, C. Gibberellin metabolism and signaling: Targets for improving agronomic performance of crops. Plant Cell Physiol. 2020, 61, 1902–1911. [Google Scholar] [CrossRef]
- Yagioka, A.; Hayashi, S.; Kimiwada, K.; Kondo, M. Sink production and grain-filling ability of a new high-yielding rice variety, Kitagenki. Field Crops Res. 2021, 260, 107991. [Google Scholar] [CrossRef]
- Kim, S.-R.; Ramos, J.M.; Hizon, R.J.M.; Ashikari, M.; Virk, P.S.; Torres, E.A.; Nissila, E.; Jena, K.K. Introgression of a functional epigenetic OsSPL14WFP allele into elite indica rice genomes greatly improved panicle traits and grain yield. Sci. Rep. 2018, 8, 3833. [Google Scholar] [CrossRef]
- Singh, V.K.; Ellur, R.K.; Singh, A.K.; Nagarajan, M.; Singh, B.D.; Singh, N.K. Effect of qGN4. 1 QTL for grain number per panicle in genetic backgrounds of twelve different mega varieties of rice. Rice 2018, 11, 8. [Google Scholar] [CrossRef]
- Reyes, V.P.; Angeles-Shim, R.B.; Mendioro, M.S.; Manuel, M.; Carmina, C.; Lapis, R.S.; Shim, J.; Sunohara, H.; Nishiuchi, S.; Kikuta, M. Marker-assisted introgression and stacking of major QTLs controlling grain number (Gn1a) and number of primary branching (WFP) to NERICA cultivars. Plants 2021, 10, 844. [Google Scholar] [CrossRef]
- Reyes, V.P.; Angeles-Shim, R.B.; Lapis, R.S.; Shim, J.; Sunohara, H.; Jena, K.K.; Ashikari, M.; Doi, K. Improvement of Asian rice cultivars through marker-assisted introgression of yield QTLs, Grain Number 1A (GN1A) and Wealthy Farmer’s Panicle (WFP). Philipp. J. Biochem. Mol. Biol. 2021, 2, 29. [Google Scholar]
- Feng, X.; Wang, C.; Nan, J.; Zhang, X.; Wang, R.; Jiang, G.; Yuan, Q.; Lin, S. Updating the elite rice variety Kongyu 131 by improving the Gn1a locus. Rice 2017, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Wu, S.; Zhu, Z.; Liu, F.; Fu, Y.; Cai, H.; Sun, X.; Gu, P.; Xie, D.; Tan, L. NOG1 increases grain production in rice. Nat. Commun. 2017, 8, 1497. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhai, L.; Chen, K.; Shen, C.; Liang, Y.; Wang, C.; Zhao, X.; Wang, S.; Xu, J. Natural sequence variations and combinations of GNP1 and NAL1 determine the grain number per panicle in rice. Rice 2020, 13, 14. [Google Scholar] [CrossRef]
- Beena, R.; Veena, V.; Jaslam, M.; Nithya, N.; Adarsh, V. Germplasm innovation for high-temperature tolerance from traditional rice accessions of Kerala using genetic variability, genetic advance, path coefficient analysis and principal component analysis. J. Crop Sci. Biotechnol. 2021, 24, 555–566. [Google Scholar] [CrossRef]
- Panda, D.; Sahu, N.; Behera, P.K.; Lenka, K. Genetic variability of panicle architecture in indigenous rice landraces of Koraput region of Eastern Ghats of India for crop improvement. Physiol. Mol. Biol. Plants 2020, 26, 1961–1971. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Miah, G.; Hussin, G.; Ramli, A. Genotype× Environment interaction and stability analyses of yield and yield components of established and mutant rice genotypes tested in multiple locations in Malaysia. Acta Agric. Scand. Sect. B Soil Plant Sci. 2017, 67, 590–606. [Google Scholar] [CrossRef]
- Sabri, R.S.; Rafii, M.Y.; Ismail, M.R.; Yusuff, O.; Chukwu, S.C.; Hasan, N.A. Assessment of agro-morphologic performance, genetic parameters and clustering pattern of newly developed blast resistant rice lines tested in four environments. Agronomy 2020, 10, 1098. [Google Scholar] [CrossRef]
- Awad-Allah, M.M.; Elekhtyar, N.M.; El-Abd, M.A.-E.-M.; Abdelkader, M.F.; Mahmoud, M.H.; Mohamed, A.H.; El-Diasty, M.Z.; Said, M.M.; Shamseldin, S.A.; Abdein, M.A. Development of New Restorer Lines Carrying Some Restoring Fertility Genes with Flowering, Yield and Grains Quality Characteristics in Rice (Oryza sativa L.). Genes 2022, 13, 458. [Google Scholar] [CrossRef]
- Singh, V.K.; Wahi, N.; Mishra, S.K.; Singh, B.; Singh, N.K. Studies on Genetic variability, correlation analysis, character association and path analysis of phenotypic characteristics of twelve mega varieties of rice and its near-isogenic lines carrying high grain number per panicle QTL qGN4. 1. Curr. Trends Biotechnol. Pharm. 2022, 16, 35–45. [Google Scholar]
- Revathi, S.; Sakthivel, K.; Manonmani, S.; Umadevi, M.; Ushakumari, R.; Robin, S. Genetics of wide compatible gene and variability studies in rice (Oryza sativa L.). J. Genet. 2016, 95, 463–467. [Google Scholar] [CrossRef]
- Khalil, M.; Hossain, M.; Chowdhury, A.; Hassan, M. Characterization of bangladeshi aus rice landraces under drought stress. SABRAO J. Breed. Genet. 2022, 54, 113–126. [Google Scholar] [CrossRef]
- Roy, D.; Gaur, A.K.; Pandey, I.D. Deciphering genetic diversity in ‘Antenna Panel’genotypes of IRRI’s Global Rice Array-IV for yield traits in Indo-Gangetic Plains. Electron. J. Plant Breed. 2022, 13, 425–431. [Google Scholar]
- Yano, K.; Morinaka, Y.; Wang, F.; Huang, P.; Takehara, S.; Hirai, T.; Ito, A.; Koketsu, E.; Kawamura, M.; Kotake, K. GWAS with principal component analysis identifies a gene comprehensively controlling rice architecture. Proc. Natl. Acad. Sci. USA 2019, 116, 21262–21267. [Google Scholar] [CrossRef] [PubMed]
- Jyothi, B.; Divya, B.; Rao, L.S.; Bhavani, P.L.; Revathi, P.; Rao, P.R.; Rachana, B.; Padmavathi, G.; Kumar, J.A.; Gireesh, C. New plant type trait characterization and development of core set among indica and tropical japonica genotypes of rice. Plant Genet. Resour. 2018, 16, 504–512. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, J.; Ren, T.; Hussain, S.; Guo, C.; Wang, S.; Cong, R.; Li, X. Effects of nitrogen and tiller type on grain yield and physiological responses in rice. AoB Plants 2017, 9, plx012. [Google Scholar] [CrossRef]
- Nahar, S.; Lahkar, L.; Islam, M.A.; Saikia, D.; Shandilya, Z.M.; Vemireddy, L.R.; Sahoo, L.; Tanti, B. Genetic diversity based on osmotic stress tolerance-related morpho-physiological traits and molecular markers in traditional rice cultivars. Biologia 2020, 75, 669–679. [Google Scholar] [CrossRef]
- Latif, M.; Rahman, M.; Kabir, M.; Ali, M.; Islam, M.; Rafii, M. Genetic diversity analyzed by quantitative traits among rice (Oryza sativa L.) genotypes resistant to blast. Afr. J. Microbiol. Res. 2011, 5, 4383–4391. [Google Scholar] [CrossRef]
- Tanaka, M.; Keira, M.; Yoon, D.-K.; Mae, T.; Ishida, H.; Makino, A.; Ishiyama, K. Photosynthetic enhancement, lifespan extension, and leaf area enlargement in flag leaves increased the yield of transgenic rice plants overproducing Rubisco under sufficient N fertilization. Rice 2022, 15, 10. [Google Scholar] [CrossRef]
- Donde, R.; Mohapatra, S.; Baksh, S.Y.; Padhy, B.; Mukherjee, M.; Roy, S.; Chattopadhyay, K.; Anandan, A.; Swain, P.; Sahoo, K.K. Identification of QTLs for high grain yield and component traits in new plant types of rice. PLoS ONE 2020, 15, e0227785. [Google Scholar] [CrossRef] [PubMed]
- Tu Anh, T.T.; Khanh, T.D.; Dat, T.D.; Xuan, T.D. Identification of phenotypic variation and genetic diversity in rice (Oryza sativa L.) mutants. Agriculture 2018, 8, 30. [Google Scholar] [CrossRef]
- Mohammadi, R.; Mendioro, M.S.; Diaz, G.Q.; Gregorio, G.B.; Singh, R.K. Genetic analysis of salt tolerance at seedling and reproductive stages in rice (O ryza sativa). Plant Breed. 2014, 133, 548–559. [Google Scholar] [CrossRef]
- Sabouri, H.; Sabouri, A.; Kavandi, R.; Katouzi, M.; Dadras, A.R. Genetic Analysis of Agronomic traits in Rice (Oryza sativa L.). Int. J. Agron. Plant Prod. 2013, 4, 1298–1304. [Google Scholar]
- AnandaLekshmi, L.; Geetha, S.; Amudha, K.; Muthuvijayaragavan, R.; Uma, D. Combining ability and gene action analysis for yield and yield attributing traits in rice (Oryza sativa. L). Electron. J. Plant Breed. 2020, 11, 901–906. [Google Scholar]
- Vadivel, K. Studies on combining ability and heterosis in rice (Oryza sativa L.). Electron. J. Plant Breed. 2018, 9, 1115–1121. [Google Scholar] [CrossRef]
- Zewdu, Z. Combining ability analysis of yield and yield components in selected rice (Oryza sativa L.) genotypes. Cogent Food Agric. 2020, 6, 1811594. [Google Scholar] [CrossRef]
- Dan, Z.; Hu, J.; Zhou, W.; Yao, G.; Zhu, R.; Huang, W.; Zhu, Y. Hierarchical additive effects on heterosis in rice (Oryza sativa L.). Front. Plant Sci. 2015, 6, 738. [Google Scholar] [CrossRef]
- Gramaje, L.V.; Caguiat, J.D.; Enriquez, J.O.S.; Millas, R.A.; Carampatana, J.E.; Tabanao, D.A.A. Heterosis and combining ability analysis in CMS hybrid rice. Euphytica 2020, 216, 14. [Google Scholar] [CrossRef]
- Azad, A.K.; Sarker, U.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almeer, R.; Sayed, A.A.; Peluso, I. Evaluation of Combining Ability and Heterosis of Popular Restorer and Male Sterile Lines for the Development of Superior Rice Hybrids. Agronomy 2022, 12, 965. [Google Scholar] [CrossRef]
- Huang, M.; Chen, L.-Y.; Chen, Z.-Q. Diallel analysis of combining ability and heterosis for yield and yield components in rice by using positive loci. Euphytica 2015, 205, 37–50. [Google Scholar] [CrossRef]
- Verma, O.; Srivastava, H. Genetic component and combining ability analyses in relation to heterosis for yield and associated traits using three diverse rice-growing ecosystems. Field Crops Res. 2004, 88, 91–102. [Google Scholar] [CrossRef]
- International Rice Research Institute (IRRI). Standard Evaluation System for Rice; International Rice Research Institute (IRRI): Los Banos, CA, USA, 2014; p. 57. ISBN 9789712203046. [Google Scholar]
- Burton, G.W.; Devane, D.E. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material 1. Agronomy 1953, 45, 478–481. [Google Scholar] [CrossRef]
- Johnson, H.W.; Robinson, H.; Comstock, R. Estimates of genetic and environmental variability in soybeans 1. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Robinson, H.; Comstock, R.E.; Harvey, P. Estimates of heritability and the degree of dominance in corn. Agron. J. 1949, 41, 353–359. [Google Scholar] [CrossRef]
- Sivasubramanian, S.; Menon, M. Heterosis and inbreeding depression in rice. Madras. Agric. J. 1973, 60, 1139–1140. [Google Scholar]
- Griffing, B. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | PH | NPT | FLA | PL | NOSPP | NFGPP | NOPB | NOSB | NOSBPB | NOSIPB | NOSISB | LOPB | TGW | GYP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sembil priyan | 163.5 | 12 | 39.02 | 28.3 | 210.5 | 202 | 11.5 | 37.5 | 3.26 | 67.5 | 143 | 11.25 | 20.4 | 45.38 |
| Karuka | 141 | 24 | 39.15 | 27.95 | 232 | 210 | 14.5 | 41.5 | 2.865 | 90.5 | 141.5 | 11.50 | 15.2 | 40.42 |
| Kallukar | 98 | 11 | 62.19 | 25.1 | 262 | 239 | 12.5 | 55 | 4.395 | 66.5 | 195.5 | 11.80 | 23.4 | 33.025 |
| Kalarkar | 123 | 21 | 33.64 | 27.3 | 131 | 120 | 7.5 | 27 | 3.6 | 34 | 97 | 10.85 | 21 | 31.625 |
| T396 Periya Samba | 113 | 19.5 | 26.95 | 23.8 | 152 | 144.5 | 8.5 | 23.5 | 2.765 | 38.5 | 113.5 | 8.50 | 19.6 | 17.48 |
| Nootripathu | 109.5 | 19 | 32.34 | 27.25 | 124 | 115 | 7.5 | 25 | 3.335 | 46.5 | 77.5 | 11.05 | 27.9 | 13.46 |
| Muttakar | 77 | 42.5 | 35.46 | 17.05 | 152.5 | 145 | 10 | 28.5 | 2.815 | 42 | 110.5 | 6.90 | 21.4 | 12.055 |
| Kothamalli samba | 154 | 15.5 | 45.13 | 28.85 | 283 | 257.5 | 17.5 | 55 | 3.14 | 113 | 170 | 11.80 | 17.3 | 48.935 |
| Katta samba | 127.5 | 16 | 49.73 | 28.95 | 189.5 | 177.5 | 14 | 39 | 2.785 | 82.5 | 107 | 10.55 | 21.25 | 25.75 |
| CO 52 | 105.5 | 15 | 30.45 | 27.75 | 285.5 | 260.5 | 13.5 | 60 | 4.445 | 67.5 | 218 | 8.85 | 22.5 | 33.5 |
| IET 29519 | 89.5 | 14 | 31.95 | 22 | 116 | 105 | 10 | 26 | 2.4 | 52 | 64 | 10.75 | 13.7 | 25.4 |
| Mikkuruvai | 134.5 | 24 | 33.47 | 25.15 | 125.5 | 119.5 | 8.5 | 19.5 | 2.3 | 49.5 | 76 | 10.60 | 23.3 | 35.6 |
| IW Ponni | 107.5 | 18.5 | 29.25 | 23.25 | 185 | 176.5 | 12.5 | 40.5 | 3.24 | 58 | 127 | 10.50 | 15 | 31.305 |
| IARI 3 | 104.5 | 18.5 | 19.22 | 21.85 | 156.5 | 150.5 | 11.5 | 31 | 2.71 | 23.5 | 133 | 10.85 | 16.9 | 18.015 |
| IET 28835 | 93.5 | 14 | 24.05 | 23 | 102 | 92.5 | 9 | 17 | 1.89 | 55.5 | 46.5 | 11.00 | 24 | 19.075 |
| IET 28834 | 101 | 11 | 17.68 | 25.55 | 122.5 | 112 | 11.5 | 27 | 2.345 | 57.5 | 65 | 9.50 | 22.4 | 11.775 |
| ADT (R) 48 | 74.5 | 18 | 12.75 | 23.75 | 180 | 172 | 14.5 | 41 | 2.835 | 50.5 | 129.5 | 8.75 | 18.4 | 21.5 |
| BPT 5204 | 93 | 14.5 | 28.88 | 22.25 | 207 | 194.5 | 11 | 38 | 3.455 | 75 | 132 | 9.00 | 14.05 | 24.415 |
| CB12132 | 112.5 | 16.5 | 44.61 | 28.9 | 430 | 409.5 | 15 | 87 | 5.97 | 73 | 357 | 13.40 | 15.9 | 37 |
| IET 28749 | 102 | 15.5 | 42.04 | 18.5 | 417.5 | 389.5 | 17 | 78 | 4.59 | 69 | 348.5 | 8.50 | 17.2 | 31.8 |
| CO 51 | 77 | 17 | 29.36 | 22 | 149 | 141.5 | 9 | 32 | 3.555 | 50 | 99 | 10.50 | 20 | 25.6 |
| CO (R) 50 | 107.5 | 23.5 | 42.69 | 29.25 | 300 | 276.5 | 14 | 63 | 4.5 | 81.5 | 218.5 | 13.90 | 21.3 | 52.5 |
| ADT 37 | 83 | 13 | 25.06 | 22.75 | 217 | 202.5 | 9 | 42.5 | 4.72 | 33.5 | 183.5 | 11.25 | 16.5 | 25.5 |
| MDU 5 | 75.5 | 20.5 | 18.36 | 27 | 140 | 127 | 10.5 | 28.5 | 2.725 | 37.5 | 102.5 | 14.35 | 21.8 | 16.5 |
| ADT 45 | 80.5 | 17.5 | 17.78 | 25.6 | 128.5 | 117.5 | 8 | 28.5 | 3.565 | 45.5 | 83 | 11.80 | 17.9 | 24.79 |
| ADT 43 | 80.5 | 16.5 | 23.16 | 24 | 203 | 193 | 8 | 42.5 | 5.315 | 47.5 | 155.5 | 12.50 | 15.1 | 21.355 |
| TRY 2 | 95.5 | 17 | 19.41 | 25.5 | 158.5 | 149.5 | 10 | 35.5 | 3.55 | 61 | 97.5 | 12.85 | 30.6 | 33.635 |
| IET 29537 | 88 | 10.5 | 28.15 | 25.9 | 385.5 | 344 | 18.5 | 81.5 | 5.765 | 102 | 283.5 | 8.70 | 12 | 34.83 |
| CRAC 3998-43-1 | 71 | 11 | 31.20 | 21.95 | 286 | 267.5 | 15 | 65.5 | 4.37 | 83 | 203 | 10.55 | 18.2 | 15.71 |
| IET 29494 | 93.5 | 18.5 | 30.27 | 22.65 | 264.5 | 239.5 | 13 | 60.5 | 4.655 | 58 | 206.5 | 10.60 | 16.4 | 31.745 |
| IET 29530 | 95.5 | 15 | 26.10 | 25.85 | 207 | 197 | 13 | 46 | 3.535 | 72 | 135 | 9.60 | 24.9 | 35.18 |
| NWGR-16022 | 87 | 10 | 51.30 | 22.4 | 354 | 326 | 12 | 72.5 | 6.04 | 51 | 303 | 10.65 | 12.3 | 22.79 |
| IET 29529 | 101.5 | 8.5 | 36.09 | 26.75 | 392.5 | 345 | 16 | 71.5 | 4.47 | 67 | 325.5 | 12.15 | 24.9 | 26.955 |
| IET29523 | 75 | 13 | 30.71 | 23 | 292 | 252 | 16 | 65 | 4.0625 | 84 | 208 | 11.85 | 15.4 | 20.33 |
| IET 29504 | 124.5 | 11 | 31.76 | 29 | 220 | 205 | 12.5 | 42 | 3.355 | 69 | 151 | 15.00 | 15.8 | 19.2 |
| IET 29539 | 81 | 18 | 29.36 | 21.15 | 340 | 315 | 17 | 72 | 4.24 | 76.5 | 263.5 | 11.85 | 15.9 | 24.995 |
| IET 29516 | 85.5 | 14.5 | 32.18 | 21.75 | 166 | 158 | 10.5 | 40 | 3.81 | 42 | 124 | 11.90 | 23.1 | 39.33 |
| IET 29495 | 98 | 10.5 | 33.92 | 22.8 | 153.5 | 143.5 | 10.5 | 38 | 3.61 | 51.5 | 102 | 14.20 | 22.3 | 37.51 |
| WGL-1283 | 103 | 15 | 36.15 | 24 | 240 | 215 | 15.5 | 56 | 3.61 | 66 | 174 | 9.60 | 20.12 | 42.355 |
| BPT 3082 | 126 | 18.5 | 27.32 | 26 | 276 | 248.5 | 12.5 | 61.5 | 4.915 | 57.5 | 218.5 | 11.55 | 15.9 | 24.25 |
| KNM 7714 | 93.5 | 27 | 38.53 | 27.6 | 380 | 360 | 16 | 76 | 4.75 | 106.5 | 273.5 | 13.40 | 13.1 | 37.64 |
| NP 9253-13 | 105 | 17.5 | 36.94 | 23.9 | 225 | 206 | 14.5 | 51 | 3.515 | 75 | 150 | 9.50 | 13 | 30.485 |
| ADT 49 | 114 | 21.5 | 31.06 | 25.55 | 268.5 | 249.5 | 13.5 | 67.5 | 5 | 56 | 212.5 | 11.55 | 13.2 | 28.205 |
| KNM 7715 | 104.5 | 19 | 37.74 | 26.75 | 333 | 307.5 | 13.5 | 62 | 4.59 | 69.5 | 263.5 | 10.15 | 12 | 41.25 |
| IET 29492 | 100 | 19.5 | 20.72 | 25 | 218 | 207 | 14 | 38.5 | 2.75 | 86 | 132 | 10.00 | 14.7 | 30 |
| GNV 1974 | 95 | 18 | 32.91 | 22.9 | 263 | 246 | 10 | 47.5 | 4.75 | 56.5 | 206.5 | 12.00 | 17 | 34.51 |
| RNR 29094 | 100 | 13.5 | 30.81 | 26.5 | 289 | 267.5 | 11.5 | 52.5 | 4.565 | 61.5 | 227.5 | 13.50 | 11.8 | 17.935 |
| BPT 3059 | 112.5 | 10.5 | 35.46 | 24.85 | 278.5 | 247 | 14.5 | 59.5 | 4.095 | 71 | 207.5 | 11.60 | 16.3 | 22.625 |
| Anna (R) 4 | 95.5 | 15 | 32.03 | 25 | 197.5 | 187.5 | 14.5 | 46.5 | 3.205 | 55 | 142.5 | 10.50 | 25.78 | 24.15 |
| ACM- 20001 | 30 | 35 | 3.50 | 8.5 | 40 | 30.5 | 7.5 | 10.5 | 1.405 | 22 | 18 | 7.50 | 16.508 | 3.5 |
| Maximum | 163.5 | 42.5 | 62.19 | 29.25 | 430 | 409.5 | 18.5 | 87 | 6.04 | 113 | 357 | 15.5 | 30.6 | 52.5 |
| Minimum | 30 | 8.5 | 3.5 | 8.5 | 40 | 30.5 | 7.5 | 10.5 | 1.405 | 22 | 18 | 6.70 | 11.8 | 3.5 |
| Pooled Mean | 100.07 | 17.11 | 32.13 | 24.45 | 228.59 | 211.29 | 12.35 | 47.06 | 3.76265 | 62.14 | 166.45 | 11.02 | 18.690 | 28.06 |
| CD @ 5% | 7.02 | 4.74 | 1.567 | 1.60 | 19.11 | 21.42 | 1.70 | 6.74 | 0.42 | 8.96 | 15.88 | 1.203 | 0.920 | 11.13 |
| CD @ 1% | 9.36 | 6.32 | 2.089 | 2.13 | 25.49 | 28.56 | 2.26 | 8.99 | 0.55 | 11.95 | 21.17 | 1.605 | 1.226 | 14.84 |
| GCV (%) | 21.79 | 34.15 | 14.19 | 13.96 | 34.35 | 36.61 | 22.76 | 37.74 | 27.07 | 30.78 | 45.71 | 15.36 | 24.132 | 33.02 |
| PCV (%) | 22.32 | 36.82 | 14.54 | 14.63 | 39.30 | 38.99 | 24.15 | 39.32 | 27.62 | 32.03 | 47.47 | 16.29 | 24.25 | 38.41 |
| h2 (bs) (%) | 95.32 | 86.01 | 95.54 | 91.04 | 76.41 | 88.18 | 88.81 | 92.11 | 96.05 | 92.36 | 92.72 | 88.50 | 98.98 | 74.05 |
| GASM (%) | 43.83 | 65.24 | 28.53 | 27.44 | 61.85 | 70.83 | 44.18 | 74.61 | 54.64 | 60.93 | 90.67 | 29.83 | 49.46 | 58.47 |
| Clusters | Number of Genotypes | Details of Genotypes |
|---|---|---|
| Cluster I | 29 | Sembil priyan, Karuka, Mikkuruvai, IET 29504, Kothamalli samba, T396 Periya Samba, BPT 3082, NP 9253-13, WGL-1283, IW Ponni, IET 29530, Anna (R) 4, ADT 49, IARI 3, IET29523, Kallukar, IET 29492, CO 52, BPT 5204, ADT 37, IET 29516, CO 51, ADT (R) 48, Muttakar, IET 29494, GNV 1974, BPT 3059, RNR 29094, CO (R) 50 |
| Cluster II | 11 | Kalarkar, Nootripathu, Katta samba, IET 28835, IET 28834, MDU 5, ADT 45, ADT 43, TRY 2, IET 29495, IET 29519 |
| Cluster III | 7 | CB12132, CRAC 3998-43-1, NWGR-16022, IET 29529, IET 29539, KNM 7714, KNM 7715, |
| Cluster IV | 1 | IET28749 |
| Cluster V | 1 | IET 29537 |
| Cluster VI | 1 | ACM-20001 |
| Clusters | PH | NPT | FLA | PL | NOSPP | NFGPP | NOPB | NOSB | NOSBPB | NOSIPB | NOSISB | LOPB | TGW | GYP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cluster I | 106.052 | 19.172 | 32.190 | 24.755 | 222.983 | 204.052 | 12.397 | 46.017 | 3.707 | 61.328 | 161.655 | 10.869 | 18.441 | 29.538 |
| Cluster II | 98.046 | 16.727 | 27.553 | 25.586 | 143.500 | 133.227 | 9.636 | 30.318 | 3.219 | 52.273 | 91.227 | 11.764 | 21.946 | 23.850 |
| Cluster III | 93.000 | 15.714 | 38.375 | 25.071 | 359.357 | 332.929 | 14.929 | 72.714 | 4.919 | 75.214 | 284.143 | 11.736 | 16.000 | 28.831 |
| Cluster IV | 102.000 | 15.500 | 42.150 | 18.500 | 417.500 | 389.500 | 17.000 | 78.000 | 4.590 | 69.000 | 348.500 | 8.500 | 17.500 | 31.800 |
| Cluster V | 88.000 | 10.500 | 28.180 | 25.900 | 385.500 | 344.000 | 18.500 | 106.500 | 5.765 | 102.000 | 283.500 | 8.700 | 12.450 | 34.830 |
| Cluster VI | 30.000 | 35.000 | 3.475 | 8.500 | 40.000 | 30.500 | 7.500 | 10.500 | 1.405 | 22.000 | 18.000 | 7.500 | 16.405 | 4.500 |
| Cluster I | Cluster II | Cluster III | Cluster IV | Cluster V | Cluster VI | |
|---|---|---|---|---|---|---|
| Cluster I | 635.2865 | 1309.547 | 3888.638 | 11,524.82 | 7891.748 | 3565.379 |
| Cluster II | - | 750.9652 | 7287.658 | 17,413.5 | 12,616.88 | 5179.511 |
| Cluster III | - | - | 1244.3348 | 3213.377 | 2119.803 | 3596.254 |
| Cluster IV | - | - | - | 0 | 1569.969 | 8143.019 |
| Cluster V | - | - | - | - | 0 | 5385.813 |
| Cluster VI | - | - | - | - | - | 0 |
| Particulars | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 | PC10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Eigen value | 6.627 | 2.147 | 1.321 | 0.968 | 0.762 | 0.703 | 0.542 | 0.42 | 0.25 | 0.14 |
| Variability (%) | 47.333 | 15.339 | 9.432 | 6.915 | 5.445 | 5.021 | 3.874 | 2.999 | 1.783 | 1.003 |
| Cumulative (%) | 47.333 | 62.672 | 72.104 | 79.02 | 84.464 | 89.486 | 93.36 | 96.358 | 98.141 | 99.144 |
| PH | −0.01 | 0.923 | 0.06 | −0.027 | 0.121 | 0.012 | 0.204 | 0.215 | 0.201 | 0.015 |
| NPT | −0.135 | −0.03 | −0.054 | 0.985 | −0.026 | −0.067 | 0.008 | −0.008 | −0.055 | 0.005 |
| FLA | 0.353 | 0.266 | 0.027 | −0.01 | 0.156 | 0.025 | 0.177 | 0.863 | 0.048 | 0.023 |
| PL | 0.155 | 0.435 | 0.331 | −0.108 | 0.21 | 0.054 | 0.16 | 0.063 | 0.767 | 0.005 |
| NOSPP | 0.928 | 0.072 | 0.061 | −0.068 | 0.259 | −0.125 | 0.091 | 0.149 | 0.047 | 0.041 |
| NFGPP | 0.926 | 0.083 | 0.067 | −0.055 | 0.261 | −0.129 | 0.093 | 0.139 | 0.051 | 0.036 |
| NOPB | 0.588 | 0.066 | −0.06 | 0.025 | 0.614 | −0.046 | 0.112 | 0.102 | 0.019 | 0.491 |
| NOSB | 0.9 | −0.036 | 0.059 | −0.05 | 0.285 | −0.114 | 0.152 | 0.086 | 0.06 | 0.137 |
| NOSBPB | 0.858 | −0.074 | 0.192 | −0.107 | −0.09 | −0.14 | 0.109 | 0.074 | 0.134 | −0.175 |
| NOSIPB | 0.349 | 0.169 | 0.067 | −0.049 | 0.856 | −0.092 | 0.229 | 0.144 | 0.168 | −0.021 |
| NOSISB | 0.969 | 0.038 | 0.052 | −0.065 | 0.071 | −0.119 | 0.044 | 0.132 | 0.01 | 0.052 |
| LOPB | 0.145 | 0.064 | 0.973 | −0.053 | 0.026 | 0.007 | 0.032 | 0.019 | 0.149 | −0.011 |
| TGW | −0.305 | 0.016 | 0.01 | −0.077 | −0.076 | 0.945 | 0.031 | 0.019 | 0.03 | −0.008 |
| GYP | 0.212 | 0.225 | 0.043 | 0.012 | 0.203 | 0.035 | 0.909 | 0.154 | 0.102 | 0.022 |
| Particulars | Mean Sum of Squares | GCA/SCA | ||||||
|---|---|---|---|---|---|---|---|---|
| RBD | Combing Ability | |||||||
| Replication | Genotypes | Error | GCA | SCA | RCA | Error | ||
| Degrees of Freedom | 2 | 35 | 70 | 5 | 15 | 15 | 35 | - |
| PH | 8.337 | 259.010 ** | 12.015 | 755.14 ** | 36.370 ** | 14.093 ** | 6.007 | 2.06 |
| NPT | 0.031 | 20.660 ** | 7.524 | 30.329 * | 12.206 * | 1.788 | 3.762 | 0.26 |
| FLA | 127.044 | 311.9755 | 22.0256 | 778.8348 * | 93.0763 * | 11.2841 | 11.0128 | 0.780 |
| PL | 2.495 | 29.355 ** | 2.935 | 40.138 * | 10.781 * | 10.087 * | 1.467 | 0.35 |
| NOSPP | 5373.389 | 27,462.184 ** | 1983.732 | 73,557.953 * | 6132.930 * | 1386.981 | 991.866 | 1.181 |
| NFGPP | 6815.281 | 24,995.463 ** | 2216.160 | 66,991.797 * | 5557.886 * | 1272.894 | 1108.080 | 1.23 |
| NOPB | 10.764 | 21.775 ** | 3.045 | 51.002 * | 5.445 * | 2.959 | 1.522 | 1.05 |
| NOSB | 113.351 | 718.196 ** | 52.101 | 2120.779 * | 97.240 * | 33.729 | 26.050 | 2.45 |
| NOSBPB | 0.009 | 1.350 ** | 0.150 | 3.886 * | 0.131 | 0.149 * | 0.075 | 5.67 |
| NOSIPB | 7.897 | 477.919 ** | 60.232 | 1257.444 * | 82.637 * | 55.788 | 30.116 | 1.95 |
| NOSISB | 5350.410 | 22,803.498 ** | 1639.771 | 58,634.324 * | 5650.208 * | 1409.104 | 819.88 | 1 |
| LOPB | 0.299 | 4.654 ** | 0.628 | 11.391 * | 1.029 * | 0.604 | 0.314 | 1.29 |
| TGW | 0.1569 | 29.9457 | 0.3921 | 84.863 ** | 6.0763 ** | 0.5723 | 0.1960 | 1.1999 |
| GYP | 9.297 | 67.758 ** | 6.727 | 96.108 * | 32.275 * | 14.740 * | 3.363 | 0.27 |
| Parents | PH | NPT | FLA | PL | NOSPP | NFGPP | NOPB | NOSB | NOSBPB | NOSIPB | NOSISB | LOPB | TGW | SPY |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IET 28834 | 5 ** | −1.271 * | −4.9228 ** | −0.817 * | −70.361 ** | −66.799 ** | −1.084 ** | −10.132 ** | −0.473 ** | −4.741 ** | −66.704 ** | −0.19 ns | 2.719 | −1.229 * |
| IET 28835 | 1.188 ns | −1.313 * | −4.3907 ** | −1.334 ** | −64.278 ** | −61.299 ** | −1.562 ** | −9.965 ** | −0.4 ** | −4.651 ** | −58.356 ** | −0.348* | 2.765 | −1.453 ** |
| ADT (R) 48 | −15.438 ** | 1.479 ** | −10.9311 ** | −1.769 ** | −75.611 ** | −71.944 ** | −2.743 ** | −15.25 ** | −0.625 ** | −16.157 ** | −61.35 ** | −1.355 ** | 1.523 | −3.768 ** |
| BPT 5204 | −0.417 ns | 2.438 ** | 2.7264 ** | 0.351 ns | 43.472 ** | 39.972 ** | 1.528 ** | 7.396 ** | 0.274 ** | 8.843 ** | 33.88 ** | −0.194 ns | −2.976 | 1.266 * |
| CB12132 | 6.063 ** | −0.292 ns | 8.1956 ** | 3.31 ** | 88.514 ** | 87.41 ** | 1.528 ** | 15.354 ** | 0.718 ** | 7.634 ** | 80.255 ** | 1.562 ** | −2.726 | 0.696 ns |
| IET 28749 | 3.604 ** | −1.042 * | 9.3226 ** | 0.26 ns | 78.264 ** | 72.66 ** | 2.333 ** | 12.597 ** | 0.504 ** | 9.072 ** | 72.275 ** | 0.525 ** | −2.726 | 4.488 ** |
| SE (gi) | 0.6459 | 0.5111 | 0.8745 | 0.1690 | 8.2994 | 8.7721 | 0.3251 | 1.3450 | 0.0721 | 1.4462 | 7.5456 | 0.1476 | −1.306 | 0.4833 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunasekaran, A.; Seshadri, G.; Ramasamy, S.; Muthurajan, R.; Karuppasamy, K.S. Identification of Newer Stable Genetic Sources for High Grain Number per Panicle and Understanding the Gene Action for Important Panicle Traits in Rice. Plants 2023, 12, 250. https://doi.org/10.3390/plants12020250
Gunasekaran A, Seshadri G, Ramasamy S, Muthurajan R, Karuppasamy KS. Identification of Newer Stable Genetic Sources for High Grain Number per Panicle and Understanding the Gene Action for Important Panicle Traits in Rice. Plants. 2023; 12(2):250. https://doi.org/10.3390/plants12020250
Chicago/Turabian StyleGunasekaran, Ariharasutharsan, Geetha Seshadri, Saraswathi Ramasamy, Raveendran Muthurajan, and Krishna Surendar Karuppasamy. 2023. "Identification of Newer Stable Genetic Sources for High Grain Number per Panicle and Understanding the Gene Action for Important Panicle Traits in Rice" Plants 12, no. 2: 250. https://doi.org/10.3390/plants12020250
APA StyleGunasekaran, A., Seshadri, G., Ramasamy, S., Muthurajan, R., & Karuppasamy, K. S. (2023). Identification of Newer Stable Genetic Sources for High Grain Number per Panicle and Understanding the Gene Action for Important Panicle Traits in Rice. Plants, 12(2), 250. https://doi.org/10.3390/plants12020250