Abstract
Reversible phosphorylation of proteins is a ubiquitous regulatory mechanism in vivo that can respond to external changes, and plays an extremely important role in cell signal transduction. Protein phosphatase 2C is the largest protein phosphatase family in higher plants. Recently, it has been found that some clade A members can negatively regulate ABA signaling pathways. However, the functions of several subgroups of Arabidopsis PP2C other than clade A have not been reported, and whether other members of the PP2C family also participate in the regulation of ABA signaling pathways remains to be studied. In this study, based on the previous screening and identification work of PP2C involved in the ABA pathway, the clade F member PIA1 encoding a gene of the PP2C family, which was down-regulated after ABA treatment during the screening, was selected as the target. Overexpression of PIA1 significantly down-regulated the expression of ABA marker gene RD29A in Arabidopsis protoplasts, and ABA-responsive elements have been found in the cis-regulatory elements of PIA1 by promoter analysis. When compared to Col-0, transgenic plants overexpressing PIA1 were less sensitive to ABA, whereas pia1 showed the opposite trait in seed germination, root growth, and stomatal opening experiments. Under drought stress, SOD, POD, CAT, and APX activities of PIA1 overexpression lines were lower than Col-0 and pia1, while the content of H2O2 was higher, leading to its lowest survival rate in test plants, which were consistent with the significant inhibition of the expression of ABA-dependent stress-responsive genes RD29B, ABI5, ABF3, and ABF4 in the PIA1 transgenic background after ABA treatment. Using yeast two-hybrid and luciferase complementation assays, PIA1 was found to interact with multiple ABA key signaling elements, including 2 RCARs and 6 SnRK2s. Our results indicate that PIA1 may reduce plant drought tolerance by functioning as a common negative regulator involved in ABA signaling pathway.
1. Introduction
Plants, on the one hand, are constantly challenged by various biological or abiotic stresses throughout their lifetime, while on the other hand, they continuously develop regulatory mechanisms that can respond to these stresses in order to survive [1]. Reversible protein phosphorylation is a ubiquitous regulatory mechanism in organisms that can promptly respond to external changes. Previous studies have revealed that it is not only an important part of the cell signal transduction pathway [2], but also plays a key role in plant responses to stress [3,4,5,6,7,8]. The completion of reversible protein phosphorylation depends mainly on protein kinases (PKs) and protein phosphatases (PPs), which act as “switches” in the signaling pathway to regulate protein activity [9,10,11]. Unlike that of PKs, which have been well studied, the function of most PPs and the link between protein dephosphorylation and signaling pathways are currently unknown due to functional redundancy among the phosphatase-encoding genes.
Based on the substrate specificity, PPs can be divided into protein tyrosine phosphatase (PTP), serine (Ser)/threonine (Thr)- specific protein phosphatase (PPP), and metal-dependent protein phosphatase (PPM) [12], among which type 2C protein phosphatase (PP2C) Ser/Thr protein phosphatase is the largest branch of the protein phosphatase family [13]. PP2C exists widely in organisms in the form of monomeric enzymes, and its activity is closely related to the metal ions such as Mg2+, Ca2+, Zn2+, and Mn2+ [14], but is not inhibited by known phosphatase inhibitors such as okadaic acid [15]. Studies have shown that PP2C can regulate cell growth and development through dephosphorylation and regulate changes in related genes in vivo through different signal transductions to resist stress [16]. In addition, compared to other organisms, PP2C in plants is the most abundant and widely present in a variety of species, including Arabidopsis [13], rice [13], maize [17], cabbage [18], and cotton [19]. The abundant number of PP2C indicates that the signal transduction mechanism in different tissues has a certain diversity, which makes it very likely to be involved in regulating plant growth and development and responding to stress through different signal pathways. At present, PP2C is usually associated with abscisic acid (ABA) in plant abiotic stress responses, which is considered to be an early discovered regulatory factor in ABA signaling pathway [7].
As an endogenous plant hormone, ABA has a marked effect on multiple physiological processes throughout the plant life cycle, including seed dormancy and germination, root growth, stomatal guard cell expansion, leaf senescence, and fruit ripening [20,21,22,23,24], and is also involved in plant responses to abiotic stress, such as osmotic stress, salt stress, drought stress, low-temperature stress, and oxidative stress [25,26,27,28,29,30,31,32]. Plants respond to ABA signaling pathways by recognizing ABA in plant cells and converting ABA stimuli into a series of physiological and biochemical responses through signal transduction [33]. In the classical ABA signaling model, PP2C physically interacts with SnRK2 (SNF1-related protein kinase 2) and thus inhibits SnRK2 kinase activity in the absence of ABA. In the presence of ABA, ABA molecules bind to the PYR/PYL/RCAR (Pyrabactin resistance/PYR-like protein/regulatory component of ABA receptor) to change the receptor conformation, and further combine with PP2C to form the ABA-PYR/PYL/RCAR-PP2C ternary complex to inhibit the activity of PP2C, thus causing the release of SnRK2. Subsequently, SnRK2 phosphorylates and activates the downstream targets in response to external stress [34,35,36,37].
According to the structural characteristics of Arabidopsis PP2C genes [38], they can be further divided into 10 groups (A-J), excluding 6 unclustered groups. At present, the research on the A clade members of PP2C is the most extensive, and a large number of studies have shown that the A clade members of PP2C are mainly involved in ABA signal regulation [39,40,41]. For example, ABI1, ABI2, and HAB1 are members of the PP2C A clade identified in Arabidopsis to negatively regulate the ABA signaling pathway [42,43,44], while AaPP2C1 in Artemisia annua had a similar function [45]. Maize ZmPP2C-A2, ZmPP2C-A6, and ZmPP2C-A10 and tomato SlPP2C3 can reduce plant drought tolerance by acting as negative regulators of ABA signals [46,47,48]. The relationship between the function of PP2C A clade members and ABA signaling has been well established, but whether other members of the PP2C family are also involved in the regulation of ABA signaling is unclear. In our previous study, we screened the non-A member genes of the Arabidopsis PP2C family that may be involved in the regulation of the ABA signaling pathway by using the protoplast transient expression system. Among the multiple non-A member PP2Cs identified in the screen, overexpression of F member encoding gene PIA1 can significantly down-regulate the expression level of ABA pathway marker genes in Arabidopsis protoplasts, but the specific mechanism involved remains to be elucidated.
In this study, on the basis of verification that overexpression of PIA1 can significantly inhibit the expression of ABA pathway reporter pRD29A using the Arabidopsis protoplast transient expression system and the presence of ABA-responsive elements in cis-regulatory elements of PIA1 through promoter analysis, the stable overexpression and functional deletion lines of PIA1 were further constructed, screened, and identified. Using them as materials, the biological function of PIA1 was analyzed by testing seed germination, root length, stomatal opening, and ABA-dependent stress response gene expression with/without ABA treatment, as well as by detecting the activity of various antioxidant enzymes and hydrogen peroxide levels under drought stress. In addition, the interactions between PP2C and RCARs or SnRK2s were analyzed by Y2H and LCA to determine their interaction networks under abiotic stress. Our data clearly indicate that PIA1 may reduce drought tolerance in plants by acting as a common negative regulator in ABA signaling pathways.
2. Results
2.1. PIA1 with ABA-Responsive Elements Down-Regulated pRD29A in Arabidopsis Protoplasts under ABA Treatment
To confirm the accuracy of our preliminary protoplast screening results, p35S::PIA1, pRD29A::LUC, and p35S::GUS were transformed together into Arabidopsis protoplasts under ABA treatment, and the effect of transient PIA1 expression on pRD29A levels was detected. Using the transformed p35S::HAHA empty vector as a control, the expression of pRD29A was found to be significantly inhibited by PIA1 (Figure 1a), which was consistent with the previous screening results.
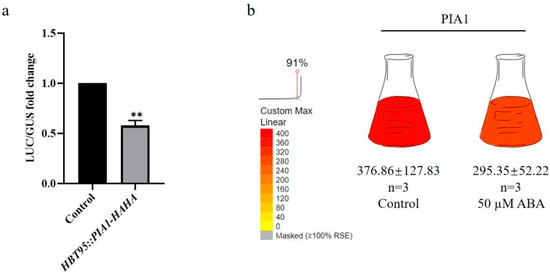
Figure 1.
Effect of PIA1 on ABA signaling pathway. (a) Effects of transient overexpression of PIA1 on ABA pathway reporter gene RD29A in Arabidopsis protoplasts. ** p < 0.01. (b) The transcription level of Arabidopsis PIA1. The data is from Böhmer et al.; the pictures were downloaded and modified from ePlant.
To determine whether the protein phosphatase PIA1 is directly involved in the abiotic stress response, the promoter sequence upstream of its start codon was analyzed using the PlantCARE database, and several abiotic stress responsive cis-elements (ABRE, TC rich repeats, CGTCA-motif, TGACG-motif, MBS, TCA-element) were identified. Moreover, PIA1 has one or more additional stress-responsive cis-elements in its promoter region, suggesting its potential involvement in various stress responses (Table 1). In addition, Böhmer et al. [49] found that the PIA1 transcript level was down-regulated in the Arabidopsis Col-0 cell line T87 after a 50 µM ABA treatment (Figure 1b), which was similar to our findings that transient-expression PIA1 in Arabidopsis protoplasts down-regulated ABA-dependent RD29A expression. To further identify the biological function of PIA1, we constructed stable OE PIA1s and identified a pia1 mutant from Arashare. Transgenic lines and mutants were verified by PCR and Western blot analysis (Figure S1).

Table 1.
Analysis of cis-acting elements in PIA1 promoter sequence.
2.2. PIA1 in Arabidopsis Affected ABA Sensitivity
To investigate whether PIA1 is related to plant response to ABA, the leaves of 4-week-old Col-0, OE PIA1, and pia1 were immersed in water and water supplemented with 20 μM ABA. Compared with Col-0, the leaves of OE PIA1 showed no significant difference, whereas the leaves of the mutant pia1 showed a whitening phenotype associated with ABA sensitivity (Figure 2a). Since seed germination is closely related to ABA response, statistical analyses of seed germination rates in Col-0, OE PIA1, and pia1 under different concentrations of ABA were performed. The results showed that the germination rate of PIA1 overexpression lines was significantly higher than that of Col-0 on 1, 3, and 6 μM ABA plates, whereas the germination rate of pia1 was significantly lower than that of Col-0 (Figure 2b,c). Root length was also used as an indicator of ABA response to detect the effect of PIA1 on plant response to ABA. Consistent with the above results, compared with Col-0, although the overexpression of PIA1 had no significant effect on root length under ABA treatment, the loss of PIA1 significantly inhibited root elongation (Figure 2d,e).
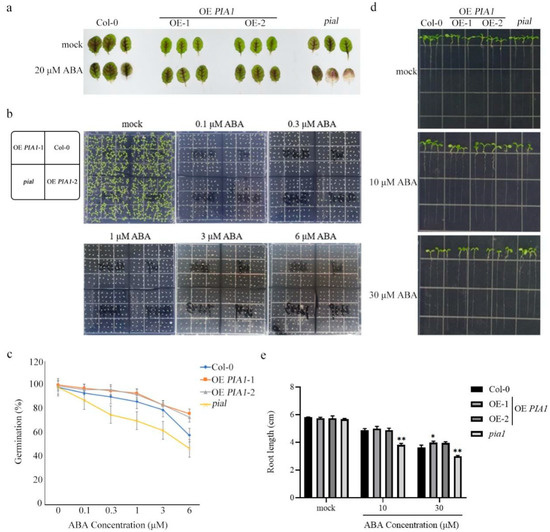
Figure 2.
The phenotype of leaves, seed germination, and root length. (a) The phenotype of leaves of Col-0, OE PIA1, and pia1 mutants. (b) The seed germination of Col-0, OE PIA1, and pia1 mutants. 100 seeds were germinated and grown on ½ MS plate containing ABA, and photographed on the 11th day. (c) Statistics of the seed germination. (d) The root length of Col-0, OE PIA1, and pia1 mutants. The seeds were germinated and grown on 1/2 MS plates for 7 days, then transferred to the plate containing ABA for 4 days, and then we took photos and count the root length. (e) Statistics of the root length. Data are means ± SE of three replicates. Significances of differences are indicated as * p < 0.05, ** p < 0.01.
2.3. PIA1-Regulated ABA-Mediated Stomatal Opening and Plant Drought Tolerance
It is known that plants can regulate stomatal opening and reduce transpiration by synthesizing endogenous ABA to improve their drought tolerance under stress [50]. Therefore, we further verified the effect of PIA1 on ABA-mediated stomatal opening, as well as plant drought tolerance under stress. Under normal conditions, the stomatal aperture of PIA1 transgenic seedlings was slightly larger than that of Col-0 and pia1. However, after ABA treatment, stomatal apertures of all tested seedlings decreased, with the largest reduction in the mutant, followed by Col-0, and transgenic plants was the least (Figure 3a,b). Similarly, PIA1 transgenic seedlings exhibited a more sensitive phenotype than Col-0 and pia1 in the soil drought stress test, with more leaves turning yellow and dying from dehydration (Figure 3c). Further water loss rate measurements confirmed the phenotype observed above (Figure 3d).
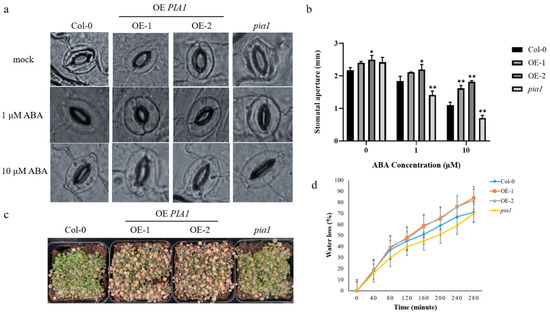
Figure 3.
The stomatal aperture comparison, soil drought test, and water loss rate analysis. (a) The stomatal aperture analyses of Col-0, OE PIA1, and pia1 mutants. (b) Statistics of the stomatal aperture. (c) Two-week-old plants of Col-0, OE PIA1, and pia1 mutants grown in short-light conditions were not irrigated for 15 days. (d) Analysis of water loss rate from detached leaves. Data are means ± SE of three replicates. Significances of differences are indicated as * p < 0.05, ** p < 0.01.
2.4. PIA1 Regulated the Activities of Antioxidant Enzymes and H2O2 Content under Drought Stress or Altered Expressions of Drought-Responsive Genes under ABA Treatment
The activities of SOD, POD, CAT, and APX and H2O2 content were measured to preliminarily analyze the physiological mechanism of PIA1 involved in plant responses to drought stress. As shown in Figure 4a–e, the decreased drought tolerance caused by overexpression of PIA1 under drought stress might be related to the significantly increased H2O2 content under a transgenic background, which was caused by the significantly decreased activities of SOD, POD, and APX, rather than CAT.
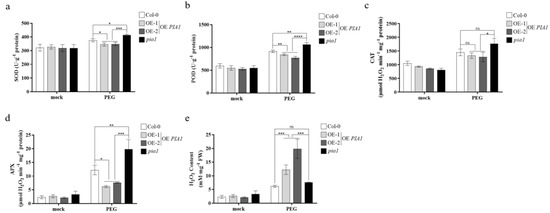
Figure 4.
Antioxidants and oxidant enzymes determination under drought condition. (a) SOD activity. (b) POD activity. (c) CAT activity. (d) APX activity. (e) H2O2 content. Significances of differences are indicated as ns indicates no significant, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
To identify molecular events involved in PIA1-mediated ABA signaling, expressions of several ABA-responsive genes, including RD29B, ABI5, ABF3, and ABF4 were investigated. The results showed that the expression levels of all detected genes were significantly down-regulated in PIA1 transgenic plants under ABA treatment and were lower than that of Col-0; results were the opposite in a pia1 mutant background (Figure 5a–d).
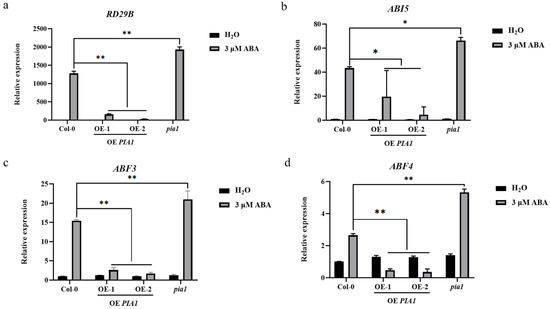
Figure 5.
Expression patterns of relevant genes in Col-0, OE PIA1, and pia1 seedlings. (a) RD29B. (b) ABI5. (c) ABF3. (d) ABF4. Significances of differences are indicated as * p < 0.05, ** p < 0.01.
2.5. PIA1 Interacted with Several Elements in the Classical ABA Signaling Model
Studies have found that ABA receptor proteins and SnRK2 family protein kinases participate in ABA signaling pathway and are regulated by group A PP2C [51,52,53,54,55]. In order to identify the target proteins of PIA1 involved in ABA signaling pathway, it was determined, by Y2H, whether the interactions were between PIA1 and 12 RCARs or 10 SnRK2s. On the premise of confirming no self-activation of BD-PIA1 (Figure 6a), we first examined the interactions between PIA1 and RCARs. It was found that PIA1 possibly interacted with RCAR1, RCAR2, and RCAR5 in the absence of exogenous ABA (Figure 6b). In order to verify the authenticity of the interactions, 3-AT was added to the SD/-Trp/-Leu/-His plates to exclude the influence of the Ade of yeast-competent cells, and X- α- Gal staining was performed, revealing that PIA1 interacted with RCAR1 and RCAR2 rather than RCAR5 (Figure 6c). Those interactions were then further confirmed not to be altered by the presence of ABA (Figure 6d).

Figure 6.
PIA1 interacts with 2 RCARs. (a) BD-PIA1 has no self-activation. (b–d) PIA1 interacts with RCAR1, RCAR2, but not with RCAR3, RCAR5, RCAR6, RCAR8, RCAR9, RCAR10, RCAR11, RCAR12, RCAR13, RCAR14.
Next, we examined the interactions between PIA1 and SnRK2s, and found that PIA1 interacted with six of the SnRK2s (SnRK2.1, SnRK2.4, SnRK2.5, SnRK2.6, SnRK2.8, and SnRK2.9) regardless of the absence/presence of ABA (Figure 7). Finally, we further demonstrated that PIAI interacted with several elements in the classical ABA signal transduction model, including 2 RCARs and 6 SnRK2s, by LCA (Figure 8, Figure S2).
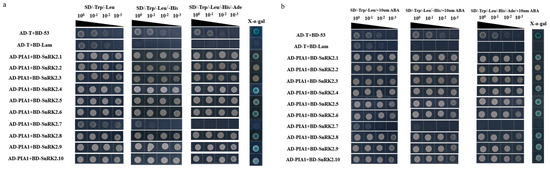
Figure 7.
PIA1 interacts with 6 SnRK2s. (a,b) PIA1 interacts with SnRK2.1, SnRK2.4, SnRK2.5, SnRK2.6, SnRK2.8, and SnRK2.9, but not with SnRK2.2, SnRK2.3, SnRK2.7, or SnRK2.10.
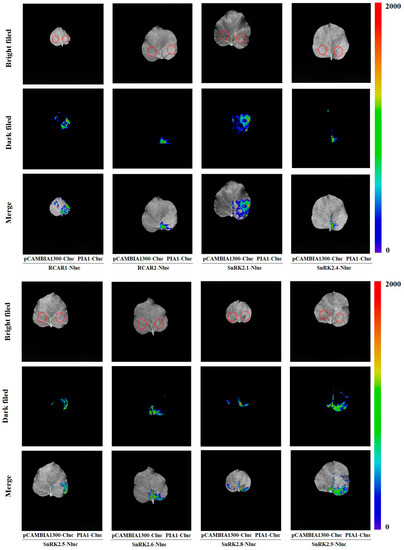
Figure 8.
PIA1 interacts with 2 RCARs and 6 SnRK2s.
3. Discussion
As a recognized stress hormone, ABA signaling pathway that response to abiotic stress in plants has been well elucidated, and key components involved have been identified, including ABA receptors, protein phosphatases, protein kinases, transcription factors, and other related elements [33]. On this basis, a classic ABA-PYR/PYL/RCAR-PP2C-SnRK2 signal transduction model has been established in Arabidopsis [34,35,36,37]. In this model, PP2C is the most diverse due to its unique physical and chemical properties, structural characteristics, and large quantities. At present, the studies on stress related to ABA regulation is mainly focused on class A PP2Cs [40,56,57,58,59], but whether other subfamily members were involved remains to be clarified. In previous studies, we used the protoplast transient expression system to systematically screen non-A class member genes of the Arabidopsis PP2C family that may be involved in ABA signaling regulation, and found several potential targets, including PIA1. In this study, on the basis of the verification of the preliminary transient screening result (Figure 1a), combined with promoter analysis and Böhmer et al.’s findings (Figure 1b), it was revealed that the cis-regulatory elements of the PIA1 promoter include ABRE and the ABA-responsive elements [60,61], and we speculated that PIA1 might play a negative regulatory role in ABA signaling.
It’s known that ABA plays an important role in the physiological processes of plant growth and development [62,63,64], and is also related to plant responses to abiotic stress [65], such as inhibiting seed germination and root growth and promoting stomatal closure to reduce water loss during drought. To test our hypothesis, we examined the effects of the overexpression and loss of function of PIA1 on ABA-induced seed dormancy (Figure 2b,c), root growth inhibition (Figure 2d,e), stomatal closure (Figure 3a,b), and drought stress gene expression (Figure 5), as well as on antioxidant enzyme activities and hydrogen peroxide content (Figure 4) under drought stress. It was found that overexpression of PIA1 reduced plant susceptibility to ABA, whereas pial exhibited increased susceptibility to ABA. Notably, the overexpression of PIA1 had no significant effect on root growth inhibition (Figure 2d,e), suggesting that PIA1 may not be a major negative regulatory gene involved in ABA regulation of the root growth pathway, but pial showed a different phenotype from Col-0, which might be due to the dose effects of hormones. Meanwhile, we also found that endogenous ABA-mediated drought tolerance was inhibited in the PIA1 transgenic background and enhanced in pia1 mutants under natural drought conditions (Figure 3c,d). So far, we have found that PIA1 can negatively regulate ABA signaling by down-regulating the positive regulatory factors encoding genes, including RD29B, ABI5, ABF3, and ABF4 in the ABA pathway [66,67], and inhibiting multiple antioxidant enzyme activities, which leads to an increase in intracellular hydrogen peroxide content and causes oxidative damage, thus increasing the sensitivity of plants to drought stress.
Since PPs function is mainly achieved through the dephosphorylation of substrates, understanding PP2C substrates is of great significance for exploring their mechanism of action. The established ABA-PYRs/PYLs/RCARs-PP2Cs-SnRK2s signaling model in Arabidopsis provides a basis for the screening of targets of PIA1 negatively regulating ABA signaling pathway. Using Y2H and LCA techniques, it was found that PIA1 interacted with both 2 RCARs and 6 SnRK2s, and their interactions were independent of ABA (Figure 6, Figure 7 and Figure 8, Figure S2). Because of the identification of multiple targets, the regulatory effect of PIA1 on ABA appears to be more general than specific. The subsequent detection of dephosphorylation of PIA1 and identification of key dephosphorylation sites are indispensable to elucidate its mechanism. Even more, future work will identify more PP2Cs that regulate the ABA signaling pathway, and completing their functional analysis will help to enrich the classical ABA signaling model.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
Arabidopsis plants used include: the Arabidopsis thaliana ecotype Colombia (Col-0), AT2G20630 overexpression lines (OE PIA1), and a pia1 mutant (SALK_105978C) from AraShare. Nicotiana benthamiana (N. benthamiana) plants used for the luciferase complementation assay (LCA) experiment were planted in nutrient soil and grown at 24 °C with 16 h of light for 5 weeks. Arabidopsis plants used for stomatal closure, soil drought test, and water loss rate analysis experiments were planted in nutrient soil and grown at 24 °C with 16 h of light for 4 weeks. For seed germination, root length, and qPCR experiments, seeds were sterilized with 75% alcohol and planted on 1/2 Murashige Skoog (MS) medium (1/2 MS medium containing 3% sucrose and 0.7% agar). After 2 days of vernalization at 4 °C, seeds were cultured vertically for 10 days at 24 °C with 16 h of light.
4.2. Vector Construction
The primers for constructing plant overexpression vectors, yeast two-hybrid (Y2H) vectors, and luciferase complementation vectors were designed according to the instructions of a one-step cloning kit (Vazyme, Nanjing, China). Table S1 summarizes primer and vector information.
4.3. Arabidopsis Protoplasts Transient Expression Assay
Transient expression in Arabidopsis protoplasts was performed as described previously [68]. HBT-PIA1-HAHA, pRD29A::LUC, and p35S::GUS were transformed together into Arabidopsis protoplasts under ABA treatment and cultured for 12 h before detection of pRD29A::LUC and GUS activity. HBT-HAHA was used as a control.
4.4. Promoter Cis-Acting Element Analysis
We used the PlantCARE (http://www.dna.affrc.go.jp/htdocs/PLACE/) (accessed on 28 May 2022) online prediction purpose gene promoter of all kinds of cis elements.
4.5. Phenotyping of Transgenic Arabidopsis
3 leaves of Col-0 mutants and transgenic plants grown for 4 weeks under short sunlight conditions were placed in an aqueous solution containing ABA at a final concentration of 20 µM or without ABA, respectively, and incubated in white light for 3 days and then photographed. The seedlings of Col-0, mutant, and transgenic plants grown for 3 weeks under short-day conditions were subjected to drought treatment without watering, and photographs were taken when obvious phenotypic differences appeared. 5 leaves of plants grown for 4 weeks under short-day conditions were cut and weighed at different time intervals to calculate the rate of water loss.
4.6. Seed Germination and Root Length
Seed germination and seedling growth were observed daily for 11 days after sowing on 1/2 MS plate with ABA. The root lengths of plants were measured by Image J.
4.7. Stomatal Opening Assay
4 leaves of Col-0, mutants, and transgenic plants grown for 4 weeks under short sunlight conditions were incubated in stomatal buffer (20 mM KCl, 1 mM CaCl2, 2.5 mM MES-KOH, pH 6.15) for 2–3 h in light (450 mol m−2 s−1). Water or ABA was added to the samples. After 2 h of continuous illumination, the lower epidermises of the leaves were observed under the microscope.
4.8. Enzymatic Determination
Col-0, mutant, and transgenic plants growing for 20 days were exposed to 20% PEG6000 to simulate drought stress. Leaves were collected at different times to prepare extracts, and the peroxidase activities (CAT, SOD, POD, APX) and hydrogen peroxide (H2O2) content were measured under specific wavelengths by colorimetry.
4.9. qPCR
The seeds of Col-0, OE PIA1 (OE-1, OE-2), and pia1 were grown vertically on 1/2 MS solid medium for 14 days. Then, 15 seedlings were transferred to 1/2 MS liquid medium for 12 h and treated with 3 μM ABA or water for 5 h. Experiments were performed under the instructions of a plant RNA rapid extraction kit (Mei5bio, Beijing, China). Reverse transcription was performed according to the instructions of hiscript III RT Supermix for the qPCR (+ gDNA wiper) kit (Vazyme). Primers are listed in Table S1.
4.10. Yeast Two-Hybrid and Luciferase Complementation Assay
Paired plasmids were transformed into the receptive states of Saccharomyces cerevisiae strain AH109 by the PEG/LiAC method [69]. After being coated on SD/-Trp/-Leu solid medium and cultured at 28 °C for 3 days, yeast cells were transferred to the specific selection media for yeast growth assessments. X-Gal staining was performed after 3 days of incubation at 28 °C [70]. The interaction was further verified by luciferase complementation assay (LCA), and the experiment was performed as described [71].
4.11. Statistical Analysis
All experiments were performed with three replicates. The data were shown as mean ± SD (standard deviation) and analyzed by using Student’s t-test at * p ≤ 0.05 and ** p ≤ 0.01.
5. Conclusions
PIA1, a clade F member of PP2C, plays a negative regulatory role in ABA signaling pathway. Its negative regulatory effect may be achieved by interacting with a variety of key ABA signaling elements to reduce the oxidative stress ability of plants under drought stress and down-regulate the expression of ABA response genes, and thereby reduce plant drought tolerance.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12142716/s1, Figure S1: OE PIA1 and pia1 mutant identified by PCR and Western blot; Figure S2: The luciferase activity measurement using a luminometer; Table S1: Primers information.
Author Contributions
R.Y. performed the experiments and analyzed the data. Y.H. and R.Y. completed the draft. Z.D., J.L., G.Y., H.Y., H.Z. and Y.C. designed the experiment. H.L., Y.Y. and S.G. worked together with R.Y. to finish the experiment. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Sichuan International Science and Technology Innovation Cooperation/Hong Kong, Macao and Taiwan Science and Technology Innovation Cooperation Project [2023YFHO103], the Sichuan Tianfu Emei Program, the Chengdu Science and Technology Bureau, Sichuan, China [2020-RC02-00060-CG], and the Henry Fok Foundation [151104].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bohnert, H.J.; Gong, Q.; Li, P.; Ma, S. Unraveling abiotic stress tolerance mechanisms--getting genomics going. Curr. Opin. Plant Biol. 2006, 9, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Li, T. A Casein Kinase II Phosphorylation Site in AtYY1 Affects Its Activity, Stability, and Function in the ABA Response. Front. Plant Sci. 2017, 8, 323. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P. The origins of protein phosphorylation. Nat. Cell Biol. 2002, 4, E127–E130. [Google Scholar] [CrossRef] [PubMed]
- Luan, S. Protein phosphatases in plants. Annu. Rev. Plant Biol. 2003, 54, 63–92. [Google Scholar] [CrossRef]
- Schweighofer, A.; Hirt, H.; Meskiene, I. Plant PP2C phosphatases: Emerging functions in stress signaling. Trends Plant Sci. 2004, 9, 236–243. [Google Scholar] [CrossRef]
- Uhrig, R.G.; Kerk, D.; Moorhead, G.B. Evolution of bacterial-like phosphoprotein phosphatases in photosynthetic eukaryotes features ancestral mitochondrial or archaeal origin and possible lateral gene transfer. Plant Physiol. 2013, 163, 1829–1843. [Google Scholar] [CrossRef]
- Humphrey, S.J.; James, D.E.; Mann, M. Protein Phosphorylation: A Major Switch Mechanism for Metabolic Regulation. Trends Endocrinol. Metab. 2015, 26, 676–687. [Google Scholar] [CrossRef]
- Kerk, D.; Bulgrien, J.; Smith, D.W.; Barsam, B.; Veretnik, S.; Gribskov, M. The complement of protein phosphatase catalytic subunits encoded in the genome of Arabidopsis. Plant Physiol. 2002, 129, 908–925. [Google Scholar] [CrossRef]
- Bai, G.; Yang, D.H.; Zhao, Y.; Ha, S.; Yang, F.; Ma, J.; Gao, X.S.; Wang, Z.M.; Zhu, J.K. Interactions between soybean ABA receptors and type 2C protein phosphatases. Plant Mol. Biol. 2013, 83, 651–664. [Google Scholar] [CrossRef]
- Cao, J.; Jiang, M.; Li, P.; Chu, Z. Genome-wide identification and evolutionary analyses of the PP2C gene family with their expression profiling in response to multiple stresses in Brachypodium distachyon. BMC Genom. 2016, 17, 175. [Google Scholar] [CrossRef]
- Barford, D.; Das, A.K.; Egloff, M.P. The structure and mechanism of protein phosphatases: Insights into catalysis and regulation. Annu. Rev. Biophys. Biomol. Struct. 1998, 27, 133–164. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Wang, D.; Zhang, S.; Ehlting, J.; Ni, F.; Jakab, S.; Zheng, C.C.; Zhong, Y. Genome-wide and expression analysis of protein phosphatase 2C in rice and Arabidopsis. BMC Genom. 2008, 9, 1471–2164. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kanwar, P.; Pandey, A.; Tyagi, A.K.; Sopory, S.K.; Kapoor, S.; Pandey, G.K. Comprehensive genomic analysis and expression profiling of phospholipase C gene family during abiotic stresses and development in rice. PLoS ONE 2013, 8, e62494. [Google Scholar] [CrossRef]
- Cohen, P. The structure and regulation of protein phosphatases. Annu. Rev. Biochem. 1989, 58, 453–508. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; He, Z.; Zhao, X.; Wang, Q.; Zhou, B.; Yu, D.S.; Huang, X.Q.; Tang, D.Y.; Guo, X.H.; et al. Molecular character of a phosphatase 2C (PP2C) gene relation to stress tolerance in Arabidopsis thaliana. Mol. Biol. Rep. 2013, 40, 2633–2644. [Google Scholar] [CrossRef]
- Wei, K.; Pan, S. Maize protein phosphatase gene family: Identification and molecular characterization. BMC Genom. 2014, 15, 773. [Google Scholar] [CrossRef]
- Khan, N.; Ke, H.; Hu, C.M.; Naseri, E.; Haider, M.S.; Ayaz, A.; Khan, W.A.; Wang, J.J.; Hou, X.L. Genome-Wide Identification, Evolution, and Transcriptional Profiling of PP2C Gene Family in Brassica rapa. BioMed Res. Int. 2019, 2019, 2965035. [Google Scholar] [CrossRef]
- Shazadee, H.; Khan, N.; Wang, J.; Wang, C.; Zeng, J.; Huang, Z.; Wang, X.Y. Identification and Expression Profiling of Protein Phosphatases (PP2C) Gene Family in Gossypium hirsutum L. Int. J. Mol. Sci. 2019, 20, 1395. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.I.; Allen, G.J.; Hugouvieux, V.; Kwak, J.M.; Waner, D. Guard Cell Signal Transduction. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 627–658. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Gampala, S.S.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14, 010441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yuan, B.; Leng, P. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit. J. Exp. Bot. 2009, 60, 1579–1588. [Google Scholar] [CrossRef]
- Ding, Z.; De Smet, I. Localised ABA signalling mediates root growth plasticity. Trends Plant Sci. 2013, 18, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Asad, M.A.U.; Zakari, S.A.; Zhao, Q.; Zhou, L.; Ye, Y.; Cheng, F. Abiotic Stresses Intervene with ABA Signaling to Induce Destructive Metabolic Pathways Leading to Death: Premature Leaf Senescence in Plants. Int. J. Mol. Sci. 2019, 20, 256. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Sun, X.; Hu, C.; Tan, Q.; Liu, J.; Liu, H. Effects of molybdenum on expression of cold-responsive genes in abscisic acid (ABA)-dependent and ABA-independent pathways in winter wheat under low-temperature stress. Ann. Bot. 2009, 104, 345–356. [Google Scholar] [CrossRef]
- Lee, S.C.; Luan, S. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 2012, 35, 53–60. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef]
- Xu, Z.; Ge, Y.; Zhang, W.; Zhao, Y.; Yang, G. The walnut JrVHAG1 gene is involved in cadmium stress response through ABA-signal pathway and MYB transcription regulation. BMC Plant Biol. 2018, 18, 19. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, G. The ABA signal transduction mechanism in commercial crops: Learning from Arabidopsis. Plant Cell Rep. 2012, 31, 1357–1369. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef]
- Rubio, S.; Rodrigues, A.; Saez, A.; Dizon, M.B.; Galle, A.; Kim, T.H.; Santiago, J.; Flexas, J.; Schroeder, S.I.; Rodriguez, P.L. Triple loss of function of protein phosphatases type 2C leads to partial constitutive response to endogenous abscisic acid. Plant Physiol. 2009, 150, 1345–1355. [Google Scholar] [CrossRef]
- Antoni, R.; Gonzalez-Guzman, M.; Rodriguez, L.; Rodrigues, A.; Pizzio, G.A.; Rodriguez, P.L. Selective inhibition of clade A phosphatases type 2C by PYR/PYL/RCAR abscisic acid receptors. Plant Physiol. 2012, 158, 970–980. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chan, Z.; Xing, L.; Liu, X.; Hou, Y.J.; Chinnusamy, V.; Wang, P.C.; Duan, C.G.; Zhu, J.K. The unique mode of action of a divergent member of the ABA-receptor protein family in ABA and stress signaling. Cell Res. 2013, 23, 1380–1395. [Google Scholar] [CrossRef]
- Haider, M.S.; Khan, N.; Pervaiz, T.; Liu, Z.J.; Nasim, M.; Jogaiah, S.; Mushtaq, N.; Jiu, S.T.; Fang, J.G. Genome-wide identification, evolution, and molecular characterization of the PP2C gene family in woodland strawberry. Gene 2019, 702, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Szostkiewicz, I.; Richter, K.; Kepka, M.; Demmel, S.; Ma, Y.; Korte, A.; Assaad, F.F.; Christmann, A.; Grill, E. Closely related receptor complexes differ in their ABA selectivity and sensitivity. Plant J. 2010, 61, 25–35. [Google Scholar] [CrossRef]
- Bhaskara, G.B.; Nguyen, T.T.; Verslues, P.E. Unique drought resistance functions of the highly ABA-induced clade A protein phosphatase 2Cs. Plant Physiol. 2012, 160, 379–395. [Google Scholar] [CrossRef]
- Couto, D.; Niebergall, R.; Liang, X.; Bücherl, C.A.; Sklenar, J.; Macho, A.P.; Ntoukakis, V.; Derbyshire, P.; Altenbach, D.; Maclean, D.; et al. The Arabidopsis Protein Phosphatase PP2C38 Negatively Regulates the Central Immune Kinase BIK1. PLoS Pathog. 2016, 12, e1005811. [Google Scholar] [CrossRef] [PubMed]
- Gosti, F.; Beaudoin, N.; Serizet, C.; Webb, A.A.; Vartanian, N.; Giraudat, J. ABI1 protein phosphatase 2C is a negative regulator of abscisic acid signaling. Plant Cell 1999, 11, 1897–1910. [Google Scholar] [CrossRef]
- Merlot, S.; Gosti, F.; Guerrier, D.; Vavasseur, A.; Giraudat, J. The ABI1 and ABI2 protein phosphatases 2C act in a negative feedback regulatory loop of the abscisic acid signalling pathway. Plant J. 2001, 25, 295–303. [Google Scholar] [CrossRef]
- Saez, A.; Apostolova, N.; Gonzalez-Guzman, M.; Gonzalez-Garcia, M.P.; Nicolas, C.; Lorenzo, O.; Rodriguez, P.L. Gain-of-function and loss-of-function phenotypes of the protein phosphatase 2C HAB1 reveal its role as a negative regulator of abscisic acid signalling. Plant J. 2004, 37, 354–369. [Google Scholar] [CrossRef]
- Zhang, F.; Fu, X.; Lv, Z.; Shen, Q.; Yan, T.; Jiang, W.; Wang, G.F.; Sun, X.F.; Tang, K.X. Type 2C phosphatase 1 of Artemisia annua L. is a negative regulator of ABA signaling. BioMed Res. Int. 2014, 2014, 521794. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Sun, X.; Gao, S.; Qin, F.; Dai, M. Deletion of an Endoplasmic Reticulum Stress Response Element in a ZmPP2C-A Gene Facilitates Drought Tolerance of Maize Seedlings. Mol. Plant 2017, 10, 456–469. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Wu, J.; Sun, X.; Dai, M. The Maize Clade A PP2C Phosphatases Play Critical Roles in Multiple Abiotic Stress Responses. Int. J. Mol. Sci. 2019, 20, 3573. [Google Scholar] [CrossRef]
- Liang, B.; Sun, Y.; Wang, J.; Zheng, Y.; Zhang, W.; Xu, Y.; Li, Q.; Leng, P. Tomato protein phosphatase 2C influences the onset of fruit ripening and fruit glossiness. J. Exp. Bot. 2021, 72, 2403–2418. [Google Scholar] [CrossRef]
- BöHmer, M.; Schroeder, J.I. Quantitative transcriptomic analysis of abscisic acid-induced and reactive oxygen species-dependent expression changes and proteomic profiling in Arabidopsis suspension cells. Plant J. 2011, 67, 105–118. [Google Scholar] [CrossRef]
- Hsu, P.K.; Dubeaux, G.; Takahashi, Y.; Schroeder, J.I. Signaling mechanisms in abscisic acid-mediated stomatal closure. Plant J. 2021, 105, 307–321. [Google Scholar] [CrossRef]
- Fujita, Y.; Nakashima, K.; Yoshida, T.; Katagiri, T.; Kidokoro, S.; Kanamori, N.; Umezawa, T.; Fujita, M.; Maruyama, K.; Ishiyama, K.V.; et al. Three SnRK2 protein kinases are the main positive regulators of abscisic acid signaling in response to water stress in Arabidopsis. Plant Cell Physiol. 2009, 50, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Fujita, Y.; Kanamori, N.; Katagiri, T.; Umezawa, T.; Kidokoro, S.; Maruyama, K.; Yoshida, T.; Ishiyama, K.; Kobayashi, M.; et al. Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signaling are essential for the control of seed development and dormancy. Plant Cell Physiol. 2009, 50, 1345–1363. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.; et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, T.; Sugiyama, N.; Mizoguchi, M.; Hayashi, S.; Myouga, F.; Yamaguchi-Shinozaki, K.; Ishihama, Y.; Hirayama, T.; Shinozaki, K. Type 2C protein phosphatases directly regulate abscisic acid-activated protein kinases in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 17588–17593. [Google Scholar] [CrossRef] [PubMed]
- Vlad, F.; Rubio, S.; Rodrigues, A.; Sirichandra, C.; Belin, C.; Robert, N.; Leung, J.; Rodriguez, P.L.; Laurière, C.; Merlot, S. Protein phosphatases 2C regulate the activation of the Snf1-related kinase OST1 by abscisic acid in Arabidopsis. Plant Cell 2009, 21, 3170–3184. [Google Scholar] [CrossRef]
- Kuhn, J.M.; Boisson-Dernier, A.; Dizon, M.B.; Maktabi, M.H.; Schroeder, J.I. The protein phosphatase AtPP2CA negatively regulates abscisic acid signal transduction in Arabidopsis, and effects of abh1 on AtPP2CA mRNA. Plant Physiol. 2006, 140, 127–139. [Google Scholar] [CrossRef]
- Nishimura, N.; Yoshida, T.; Kitahata, N.; Asami, T.; Shinozaki, K.; Hirayama, T. ABA-Hypersensitive Germination1 encodes a protein phosphatase 2C, an essential component of abscisic acid signaling in Arabidopsis seed. Plant J. 2007, 50, 935–949. [Google Scholar] [CrossRef]
- Rodrigues, A.; Adamo, M.; Crozet, P.; Margalha, L.; Confraria, A.; Martinho, C.; Elias, A.; Rabissi, A.; Lumbreras, V.; González-Guzmán, M.; et al. ABI1 and PP2CA phosphatases are negative regulators of Snf1-related protein kinase1 signaling in Arabidopsis. Plant Cell 2013, 25, 3871–3884. [Google Scholar] [CrossRef]
- Babula-SkowronSka, D.; LudwikoW, A.; CiesLa, A.; Olejnik, A.; Cegielska-Taras, T.; Bartkowiak-Broda, I.; Sadowski, J. Involvement of genes encoding ABI1 protein phosphatases in the response of Brassica napus L. to drought stress. Plant Mol. Biol. 2015, 88, 445–457. [Google Scholar] [CrossRef]
- Pla, M.; Vilardell, J.; Guiltinan, M.J.; Marcotte, W.R.; Niogret, M.F.; Quatrano, R.S.; Pagès, M. The cis-regulatory element CCACGTGG is involved in ABA and water-stress responses of the maize gene rab28. Plant Mol. Biol. 1993, 21, 259–266. [Google Scholar] [CrossRef]
- Choi, H.; Hong, J.; Ha, J.; Kang, J.; Kim, S.Y. ABFs, a family of ABA-responsive element binding factors. J. Biol. Chem. 2000, 275, 1723–1730. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef]
- Brookbank, B.P.; Patel, J.; Gazzarrini, S.; Nambara, E. Role of Basal ABA in Plant Growth and Development. Genes 2021, 12, 1936. [Google Scholar] [CrossRef]
- Sano, N.; Marion-Poll, A. ABA Metabolism and Homeostasis in Seed Dormancy and Germination. Int. J. Mol. Sci. 2021, 22, 5069. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kuma, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Mayank Pandey Abscisic Acid Signaling and Abiotic Stress Tolerance in Plants: A Review on Current Knowledge and Future Prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef]
- Liu, W.; Sikora, E.; Park, S.W. Plant growth-promoting rhizobacterium, Paenibacillus polymyxa CR1, upregulates dehydration-responsive genes, RD29A and RD29B, during priming drought tolerance in arabidopsis. Plant Physiol. Biochem. 2020, 156, 146–154. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Schiestl, R.H. Large-scale high-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 38–41. [Google Scholar] [CrossRef] [PubMed]
- MöCkli, N.; Auerbach, D. Quantitative beta-galactosidase assay suitable for high-throughput applications in the yeast two-hybrid system. Biotechniques 2004, 36, 872–876. [Google Scholar] [CrossRef]
- Zhou, Z.; Bi, G.; Zhou, J.M. Luciferase Complementation Assay for Protein-Protein Interactions in Plants. Curr. Protoc. Plant Biol. 2018, 3, 42–50. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).