
Evolution of the Xerocarpa clade (Opuntia; Opuntieae): Evidence for the Role of the Grand Canyon in the Biogeographic History of the Iconic Beavertail Cactus and Relatives

 ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. Sequencing, Assembly and Alignments
2.2. Phylogeny
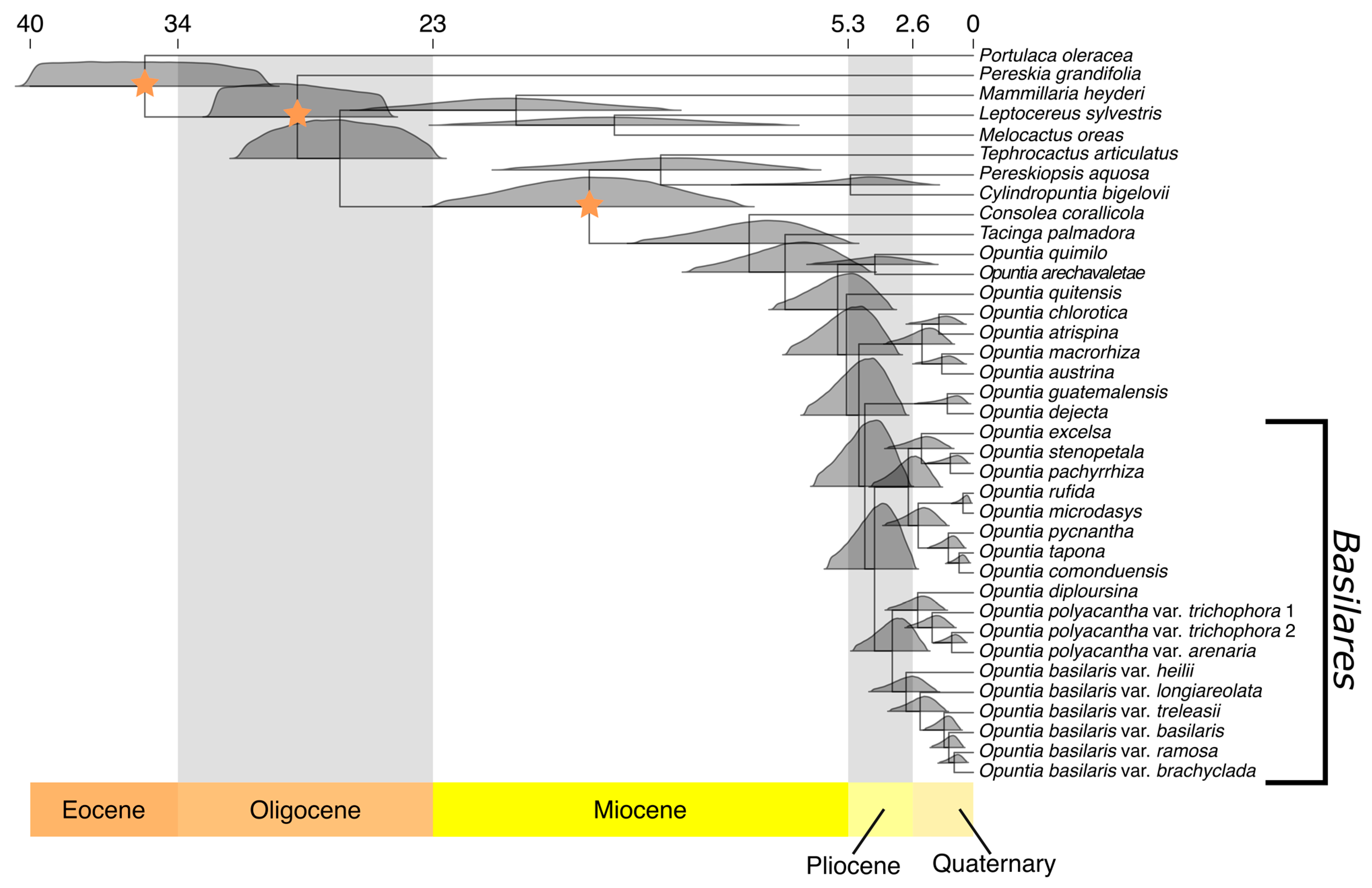
2.3. Divergence Time Estimation
2.4. Ancestral Area Estimation
2.5. Morphological Evolution
3. Discussion
3.1. Phylogeny
3.2. Biogeographic Patterns
3.3. Morphological Evolution
3.4. Taxonomy in the Xerocarpa Clade
4. Materials and Methods
4.1. Taxon Sampling
4.2. DNA Extraction, Sequencing, Alignment, and Phylogenetic Analysis
4.3. Divergence Time Estimation
4.4. Ancestral Area Estimation
4.5. Ancestral Character State Reconstruction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Wilson, J.S.; Pitts, J.P. Illuminating the Lack of Consensus among Descriptions of Earth History Data in the North American Deserts: A Resource for Biologists. Prog. Phys. Geogr. Earth Environ. 2010, 34, 419–441. [Google Scholar] [CrossRef]
- Morafka, D.J. A Biogeographical Analysis of the Chihuahuan Desert through Its Herpetofauna; Biogeographica: The Hague, The Netherlands, 1977; Volume 9. [Google Scholar]
- Petersen, K.L. Modern and Pleistocene climatic patterns in the west. In Natural History of the Colorado Plateau and Great Basin; Harper, K.T., St. Clair, L.L., Thorne, K.H., Hess, W.N., Eds.; University Press of Colorado: Denver, CO, USA, 1998; pp. 27–54. [Google Scholar]
- Graham, A. Late Cretaceous and Cenozoic History of North American Vegetation; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Beir, B.-A.; Nylander, J.A.A.; Chase, M.W.; Thulin, M. Phylogenetic relationships and biogeography of the desert plant Fagonia (Zygophyllaceae), inferred by parsimony and Bayesian model averaging. Mol. Phylogenet. Evol. 2004, 33, 91–108. [Google Scholar] [CrossRef]
- Moore, M.J.; Tye, A.; Jansen, R.K. Patterns of long-distance dispersal in Tiquilia subg. Tiquilia (Boraginaceae): Implications for the origins of amphitropical disjuncts and Galapagos Islands endemics. Am. J. Bot. 2006, 93, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.G.; Johnson, L.A.; Villaverde, T.; Guilliams, M. American amphitropical disjuncts: Perspectives from vascular plant analyses and prospects for future research. Am. J. Bot. 2017, 104, 1600–1650. [Google Scholar] [CrossRef]
- Majure, L.C.; Blankenship, A.; Grinage, A.; Noa-Monzón, A. Castela (Simaroubaceae), an impressive New World radiation of thorny shrubs destined for edaphically dry habitats. Braz. J. Bot. 2022, 45, 237–249. [Google Scholar] [CrossRef]
- Majure, L.C.; Puente, R.; Griffith, M.P.; Judd, W.S.; Soltis, P.S.; Soltis, D.S. Phylogeny of Opuntia s.s. (Cactaceae): Clade delineation, geographic origins, and reticulate evolution. Am. J. Bot. 2012, 99, 847–864. [Google Scholar] [CrossRef]
- Majure, L.C.; Baker, M.; Cloud-Hughes, M.; Salywon, A.; Neubig, K.M. Phylogenomics in Cactaceae: A case study using the chollas sensu lato (Cylindropuntieae, Opuntioideae) reveals a common pattern out of the Chihuahuan/Sonoran Desert. Am. J. Bot. 2019, 106, 1327–1345. [Google Scholar] [CrossRef]
- Hawkins, J.A.; Boutaoui, N.; Cheung, K.Y.; Van Klinken, R.D.; Hughes, C.E. Intercontinental dispersal prior to human translocation revealed in a cryptogenic invasive tree. New Phytol. 2007, 175, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Lia, V.V.; Confalonieri, V.A.; Comas, C.I.; Hunziker, J.H. Molecular phylogeny of Larrea and its allies (Zygophyllaceae): Reticulate evolution and the probable time of creosote bush arrival to North America. Mol. Phylogenet. Evol. 2001, 21, 309–320. [Google Scholar] [CrossRef]
- Simpson, B.B.; Tate, J.A.; Weeks, A. The biogeography of Hoffmannseggia (Leguminosae, Caesalpinoideae, Caesalpinieae): A tale of many travels. J. Biogeogr. 2005, 32, 15–27. [Google Scholar] [CrossRef]
- Gibson, A.C.; Nobel, P.S. The Cactus Primer; Harvard University Press: Cambridge, MA, USA, 1986. [Google Scholar]
- Majure, L.C.; Achá, S.; Baker, M.A.; Puente-Martínez, R.; Köhler, M.; Fehlberg, S.D. Phylogenomics of one of the world’s most intriguing groups of CAM plants, the Opuntioids (Opuntioideae: Cactaceae). Diversity 2023, 15, 570. [Google Scholar] [CrossRef]
- Hernández-Hernández, T.; Brown, J.W.; Schlumpberger, B.O.; Eguiarte, L.E.; Magallón, S. Beyond aridification: Multiple explanations for the elevated diversification of cacti in the New World succulent biome. New Phytol. 2014, 202, 1382–1397. [Google Scholar] [CrossRef] [PubMed]
- Hershkovitz, M.A.; Zimmer, E.A. On the evolutionary origins of the cacti. Taxon 1997, 46, 217–232. [Google Scholar] [CrossRef]
- Vázquez-Sánchez, M.; Terrazas, T.; Arias, S.; Ochoterena, H. Molecular phylogeny, origin and taxonomic implications of the tribe Cacteae (Cactaceae). Syst. Biod. 2013, 11, 103–116. [Google Scholar] [CrossRef]
- Ocampo, G.; Columbus, J.T. Molecular phylogenetics of suborder Cactinae (Caryophyllales), including insights into photosynthetic diversification and historical biogeography. Am. J. Bot. 2010, 97, 1827–1847. [Google Scholar] [CrossRef]
- Arakaki, M.; Christin, P.; Nyffeler, R.; Lendel, A.; Eggli, U.; Ogburn, R.M.; Spriggs, E.; Moore, M.J.; Edwards, E.J. Contemporaneous and recent radiations of the world’s major succulent plant lineages. Proc. Natl. Acad. Sci. USA 2011, 108, 8379–8384. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.P.; Porter, M. Phylogeny of Opuntioideae (Cactaceae). Int. J. Plant Sci. 2009, 170, 107–116. [Google Scholar] [CrossRef]
- Majure, L.C.; Puente, R. Phylogenetic relationships and morphological evolution in Opuntia s. str. and closely related members of tribe Opuntieae. Succul. Plant Res. 2014, 8, 9–30. [Google Scholar]
- Majure, L.C.; Judd, W.S.; Soltis, P.S.; Soltis, D.E. A taxonomic revision of the Opuntia humifusa complex (Opuntia s.s.: Cactaceae). Phytotaxa 2017, 290, 1–65. [Google Scholar] [CrossRef]
- Anderson, E.A. The Cactus Family; Timber Press: Portland, OR, USA, 2001. [Google Scholar]
- Hernández, H.M.; Gómez-Hinostrosa, C.; Bárcenas, R.T. Studies on Mexican Cactaceae. I. Opuntia pachyrrhiza, a new species from the Chihuahuan Desert, Mexico. Novon 2001, 11, 309–314. [Google Scholar]
- Engelmann, G. Synopsis of the Cactaceae of the territory of the United States and adjacent regions. Proc. Am. Acad. 1856, 3, 259–346. [Google Scholar]
- Baker, M.B.; Pinkava, D.J. Chromosome numbers in some cacti of western North America—IX. Haseltonia 2018, 25, 5–29. [Google Scholar] [CrossRef]
- Parfitt, B.D. Biosystematics of the Opuntia polyacantha (Cactaceae) Complex of Western North America. Ph.D. Dissertation, Arizona State University, Tempe, AZ, USA, 1991. [Google Scholar]
- Powell, A.M.; Weedin, J.F. Cacti of the Trans-Pecos & Adjacent Areas; Texas Tech University Press: Lubbock, TX, USA, 2004. [Google Scholar]
- Majure, L.C.; Puente, R.; Pinkava, D.J. Miscellaneous chromosome counts in Opuntieae DC. (Cactaceae) with a compilation of counts for the group. Haseltonia 2012, 18, 67–78. [Google Scholar] [CrossRef]
- Pinkava, D.J. (Ed.) Opuntia. In The Flora of North America Editorical Committee; 1993+; Flora of North America North of Mexico: New York, NY, USA, 2003; Volume 12, pp. 123–148. [Google Scholar]
- Stock, A.D.; Hussey, N.; Beckstrom, M.D. A new species of Opuntia (Cactaceae) from Mohave Co., Arizona. Cactus Succul. J. 2014, 86, 79–83. [Google Scholar] [CrossRef]
- Stock, A.D.; Lee, K.; Wellard, B.; Spallone, P. New nomenclatural combinations and types for Cactaceae in Utah. Haseltonia 2023, 30, 6–13. [Google Scholar] [CrossRef]
- Welsh, S.L.; Neese, E. New variety of Opuntia basilaris (Cactaceae) from Utah. Great Basin Nat. 1984, 43, 700. [Google Scholar]
- Parish, S.B. New California plants. Bull. Torrey Bot. Club 1892, 19, 91–93. [Google Scholar] [CrossRef]
- Parfitt, B.D. Opuntia pinkavae (Cactaceae), a new species from Arizona and Utah. Rhodora 1997, 99, 223–228. [Google Scholar]
- Breslin, P.; Romero, R.; Starr, G.; Watkins, V. Field Guide to Cacti and Other Succulents of Arizona; Tucson Cactus and Succulent Society: Tucson, AZ, USA, 2014. [Google Scholar]
- Pinkava, D.J. Chromosomes of the continental North American Oputioideae. Succul. Plant Res. 2002, 6, 59–98. [Google Scholar]
- Polyak, V.; Hill, C.; Asmerom, Y. Age and evolution of the Grand Canyon revealed by U-Pb dating of water table-type speleothems. Science 2008, 319, 1377–1380. [Google Scholar] [CrossRef]
- Karlstrom, K.E.; Lee, J.P.; Kelley, S.A.; Crow, R.S.; Crossey, L.J.; Young, R.A.; Lazear, G.; Beard, L.S.; Ricketts, J.W.; Fox, M.; et al. Formation of the Grand Canyon 5 to 6 million years ago through integration of older paleocanyons. Nat. Geosci. 2014, 7, 239–244. [Google Scholar] [CrossRef]
- Sanderson, M.J.; Copetti, D.; Búrquez, A.; Bustamante, E.; Charboneau, J.L.M.; Eguiarte, L.E.; Kumar, S.; Lee, H.O.; Lee, J.; McMahon, M.; et al. Exceptional reduction of the plastid genome of saguaro cactus (Carnegiea gigantea): Loss of the ndh gene suite and inverted repeat. Am. J. Bot. 2015, 102, 1115–1127. [Google Scholar] [CrossRef]
- Köhler, M.; Reginato, M.; Souza-Chies, T.T.; Majure, L.C. Next-generation sequencing across Opuntioideae (Cactaceae): Insights into plastome structural variation and phylogenetic markers in a prickly group. Front. Plant Sci. 2020, 11, 729. [Google Scholar] [CrossRef]
- Wilson, J.S.; Pitts, J.P. Identifying Pleistocene refugia in North American cold deserts using phylogeographic analyses and ecological niche modeling. Divers. Distrib. 2012, 18, 1139–1152. [Google Scholar] [CrossRef]
- Richardson, B.A.; Meyer, S.E. Paleoclimate effects and geographic barriers shape regional population genetic structure of blackbrush (Coleogyne ramosissima: Rosaceae). Botany 2012, 90, 293–299. [Google Scholar] [CrossRef]
- Wilder, B.T.; Becker, A.T.; Munguia-Vega, A.; Culver, M. Tracking the desert’s edge with a Pleistocene relict. J. Arid Environ. 2022, 196, 104653. [Google Scholar] [CrossRef]
- Majure, L.C.; Judd, W.S.; Soltis, P.S.; Soltis, D.E. Cytogeography of the Humifusa clade of Opuntia s.s. Mill. 1754 (Cactaceae: Opuntioideae): Correlations with geographic distributions and morphological differentiation of a polyploid complex. Comp. Cytogenet. 2012, 6, 53–77. [Google Scholar] [CrossRef] [PubMed]
- Frey, J.K. Modes of peripheral isolate formation and speciation. Syst. Biol. 1993, 42, 373–381. [Google Scholar] [CrossRef]
- Elizondo, J.L. Opuntia erinacea var. hystricina (Eng. & Bigelow) Benson en la reserva de la biosfera de Mapimí. Cactáceas Y Suculentas Mex. 1988, 33, 67–70. [Google Scholar]
- Dorsey, R.J.; O’Connell, B.; McDougall, K.; Homan, M.B. Punctuated sediment discharge during early Pliocene birth of the Colorado River: Evidence from regional stratigraphy, sedimentology, and paleontology. Sediment. Geol. 2018, 363, 1–33. [Google Scholar] [CrossRef]
- Beck, M.J.; Vander Wall, S.B. Seed dispersal by scatter-hoarding rodents in arid environments. J. Ecol. 2010, 98, 1300–1309. [Google Scholar] [CrossRef]
- Frego, K.A.; Staniforth, R.J. Factors determining the distribution of Opuntia fragilis in the boreal forest of southeastern Manitoba. Can. J. Bot. 1985, 63, 2377–2382. [Google Scholar] [CrossRef]
- Majure, L.C.; Fitzpatrick, P.; Ervin, G.N. Storm-driven maritime dispersal of prickly pear cacti (Opuntia species). In Proceedings of the USDA, Animal and Plant Health Inspection Service, International Cactoblastis cactorum Conference, Phoenix, AZ, USA, 7–10 May 2007. [Google Scholar]
- Crow, R.; Karlstrom, K.E.; McIntosh, W.; Peters, L.; Dunbar, N. History of Quaternary volcanism and lava dams in the western Grand Ganyon based on lidar analysis, 40Ar/39Ar dating, and field studies: Implications for flow stratigraphy, timing of volcanic events, and lava dams. Geosphere 2008, 4, 183–206. [Google Scholar] [CrossRef]
- Nobel, P.S. Spine influences on PAR interception, stem temperature, and nocturnal acid accumulation by cacti. Plant Cell Environ. 1983, 6, 153–159. [Google Scholar] [CrossRef]
- Nobel, P.S. Surface temperatures of cacti—Influences of envirvonmental and morphological factors. Ecol. Soc. Am. 1978, 59, 986–996. [Google Scholar]
- Nobel, P.S.; Bobich, E.G. Environmental Biology. In Cacti: Biology and Uses; Nobel, P.S., Ed.; University of California Press: Berkeley, CA, USA, 2001; pp. 57–74. [Google Scholar]
- Ellner, S.; Shmida, A. Why are adaptations for long-range seed dispersal rare in desert plants? Oecologia 1981, 51, 133–144. [Google Scholar]
- Maddox, J.C.; Carlquist, S. Wind dispersal in the Californian desert plants: Experimental studies and conceptual considerations. Aliso 1985, 11, 77–96. [Google Scholar]
- Bronstein, J.L.; Izhaki, I.; Nathan, R.; Tewksbury, J.J.; Spiegel, O.; Lotan, A.; Altstein, O. Seed dispersal: Theory and its application in a changing world. In Fleshy-Fruited Plants and Frugivores in Desert Ecosystems; Dennis, A.J., Schupp, E.W., Green, R.J., Westcott, D.A., Eds.; CABI: Cambridge, UK, 2007; pp. 148–177. [Google Scholar]
- Felger, R.S.; Rutman, S.; Malusa, J.; Baker, M.A. Ajo Peak to Tinajas Altas: A flora in southwestern Arizona. Part 7. Eudicots: Cactaceae–cactus family. Phytoneuron 2014, 69, 1–95. [Google Scholar]
- Majure, L.C.; Ribbens, E. Chromosome counts of Opuntia (Cactaceae), prickly pear cacti, in the Midwestern United States and environmental factors restricting the distribution of Opuntia Fragilis. Haseltonia 2012, 17, 58–65. [Google Scholar] [CrossRef]
- Pimienta-Barrios, E.; del Castillo, R.F. Reproductive Biology. In Cacti, Biology and Uses; Nobel, P.S., Ed.; University of California Press: Berkeley, CA, USA, 2001; pp. 75–90. [Google Scholar]
- McCarten, N.F. Fossil cacti and other succulents from the late Pleistocene. Cactus Succul. J. 1981, 53, 122–123. [Google Scholar]
- Van Devender, T.R.; Spaulding, W.G. Development of vegetation and climate in the southwestern United States. Science 1979, 204, 710. [Google Scholar] [CrossRef]
- Stebbins, G.L. Variation and Evolution in Plants; Columbia University Press: New York, NY, USA, 1950. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Straub, S.C.K.; Parks, M.; Weitemier, K.; Fishbein, M.; Cronn, R.C.; Liston, A. Navigating the tip of the genomic iceberg: Next-generation sequencing for plant systematics. Am. J. Bot. 2012, 99, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Neubig, K.M.; Whitten, W.M.; Abbott, J.R.; Elliott, S.; Soltis, D.E.; Soltis, P.S. Variables affecting DNA preservation in archival plant specimens. In DNA Banking for the 21st Century: Proceedings of the USWorkshop on DNA Banking; Missouri Botanical Garden; Applequist, W.L., Campbell., L.M., Eds.; William, L. Brown Center: St. Louis, MI, USA, 2014; pp. 81–112. [Google Scholar]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Blankenberg, D.; Gordon, A.; Von Kuster, G.; Coraor, N.; Taylor, J.; Nekrutenko, A. Manipulation of FASTQ data with Galaxy. Bioinformatics 2010, 26, 1783–1785. [Google Scholar] [CrossRef]
- Köhler, M.; Reginato, M.; Jin, J.-J.; Majure, L.C. More than a spiny morphology: Plastome variation in the prickly pear cacti (Opuntieae). Ann. Bot. 2023. [Google Scholar] [CrossRef]
- Stamatakis, A. RaxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Yang, Z.; Rannala, B. Bayesian estimation of species divergence Times under a Molecular Clock Using Multiple Fossil Calibrations with Soft Bounds. Mol. Biol. Evol. 2006, 23, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Puttick, M.; Title, P. MCMCtreeR: Prepare MCMCtree Analyses and Plot Bayesian Divergence Time Analyses Estimates on Trees. R Package Version 1.1. 2019. Available online: https://CRAN.R-project.org/package=MCMCtreeR (accessed on 1 December 2022).
- Griffith, G.E.; Omernik, J.M.; Azevedo, S.H. Ecological Classification of the Western Hemisphere; Unpublished Report; Environmental Protection Agency, Western Ecology Division: Corvallis, OR, USA, 1998; 49p. [Google Scholar]
- Matzke, N.J. Model selection in historical biogeography reveals that founder-event speciation is a crucial process in island clades. Syst. Biol. 2014, 63, 951–970. [Google Scholar] [CrossRef] [PubMed]
- Ree, R.H.; Moore, B.R.; Webb, C.O.; Donoghue, M.J. A likelihood framework for inferring the evolution of geographic range on phylogenetic trees. Evolution 2005, 59, 2299–2311. [Google Scholar] [PubMed]
- Ronquist, F. Dispersal-vicariance analysis: A new approach to the quantification of historical biogeography. Syst. Biol. 1997, 46, 195–203. [Google Scholar] [CrossRef]
- Landis, M.J.; Matzke, N.J.; Moore, B.R.; Huelsenbeck, J.P. Bayesian Analysis of Biogeography when the Number of Areas is Large. Syst. Biol. 2013, 62, 789–804. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 1 December 2022).
- Matzke, N.J. Probabilistic historical biogeography: New models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and model-testing. Front. Biogeogr. 2013, 5, 242–248. [Google Scholar] [CrossRef]
- Matzke, N. BioGeoBEARS: BioGeography with Bayesian (and Likelihood) Evolutionary Analysis in R Scripts; University of California: Berkeley, CA, USA, 2013. [Google Scholar]
- Matzke, N. cladoRcpp v0.15.1: C++ Implementations of Phylogenetic Cladogenesis Calculations; University of Auckland: Auckland, New Zealand, 2018. [Google Scholar]
- Sidje, R.B. A software package for computing matrix exponentials. ACM-Trans. Math. Softw. 1998, 24, 130–156. [Google Scholar] [CrossRef]
- Matzke, N.; Sidje, R.; Schmidt, D. rexpokit v0.26.6.6: R Wrappers for EXPOKIT; Other Matrix Functions; School of Biological Sciences, University of Auckland: Auckland, New Zealand, 2019. [Google Scholar]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.6. 2018. Available online: http://www.mesquiteproject.org (accessed on 1 November 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Number of Parameters | d | e | j | LnL | AICc |
|---|---|---|---|---|---|---|
| DEC | 2 | 0.03215266 | 4.375713 × 10−2 | 0 | −55.27566 | 115.2572 |
| DEC+J | 3 | 0.01711820 | 1.000000 × 10−12 | 0.03940934 | −50.40807 | 108.3161 |
| DIVALIKE | 2 | 0.03401536 | 2.001000 × 10−9 | 0 | −52.35097 | 109.4078 |
| DIVALIKE+J | 3 | 0.02024158 | 1.000000 × 10−12 | 0.02959976 | −49.65643 | 106.8129 |
| BAYAREALIKE | 2 | 0.03844895 | 3.691173 × 10−1 | 0 | −58.96130 | 122.6285 |
| BAYAREALIKE+J | 3 | 0.01266913 | 4.732005 × 10−2 | 0.05175805 | −53.18821 | 113.8764 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majure, L.C.; Murphy, T.H.; Köhler, M.; Puente, R.; Hodgson, W.C. Evolution of the Xerocarpa clade (Opuntia; Opuntieae): Evidence for the Role of the Grand Canyon in the Biogeographic History of the Iconic Beavertail Cactus and Relatives. Plants 2023, 12, 2677. https://doi.org/10.3390/plants12142677
Majure LC, Murphy TH, Köhler M, Puente R, Hodgson WC. Evolution of the Xerocarpa clade (Opuntia; Opuntieae): Evidence for the Role of the Grand Canyon in the Biogeographic History of the Iconic Beavertail Cactus and Relatives. Plants. 2023; 12(14):2677. https://doi.org/10.3390/plants12142677
Chicago/Turabian StyleMajure, Lucas C., Thomas H. Murphy, Matias Köhler, Raul Puente, and Wendy C. Hodgson. 2023. "Evolution of the Xerocarpa clade (Opuntia; Opuntieae): Evidence for the Role of the Grand Canyon in the Biogeographic History of the Iconic Beavertail Cactus and Relatives" Plants 12, no. 14: 2677. https://doi.org/10.3390/plants12142677
APA StyleMajure, L. C., Murphy, T. H., Köhler, M., Puente, R., & Hodgson, W. C. (2023). Evolution of the Xerocarpa clade (Opuntia; Opuntieae): Evidence for the Role of the Grand Canyon in the Biogeographic History of the Iconic Beavertail Cactus and Relatives. Plants, 12(14), 2677. https://doi.org/10.3390/plants12142677