
The Seasonal Divergence in the Weakening Relationship between Interannual Temperature Changes and Northern Boreal Vegetation Activity



Abstract
1. Introduction
2. Data and Methods
2.1. Data Sources
2.2. Methods
3. Results
3.1. Trends of Correlations between Vegetation Activity and Temperature Factors
3.1.1. Inter-Annual Changes in RNDVI−Tmean and RNDVI−DTR
3.1.2. Intra-Annual Changes in RNDVI−Tmean and RNDVI−DTR
3.2. Spatial Patterns of the Trends in the Correlations between Vegetation Activity and Temperature Factors
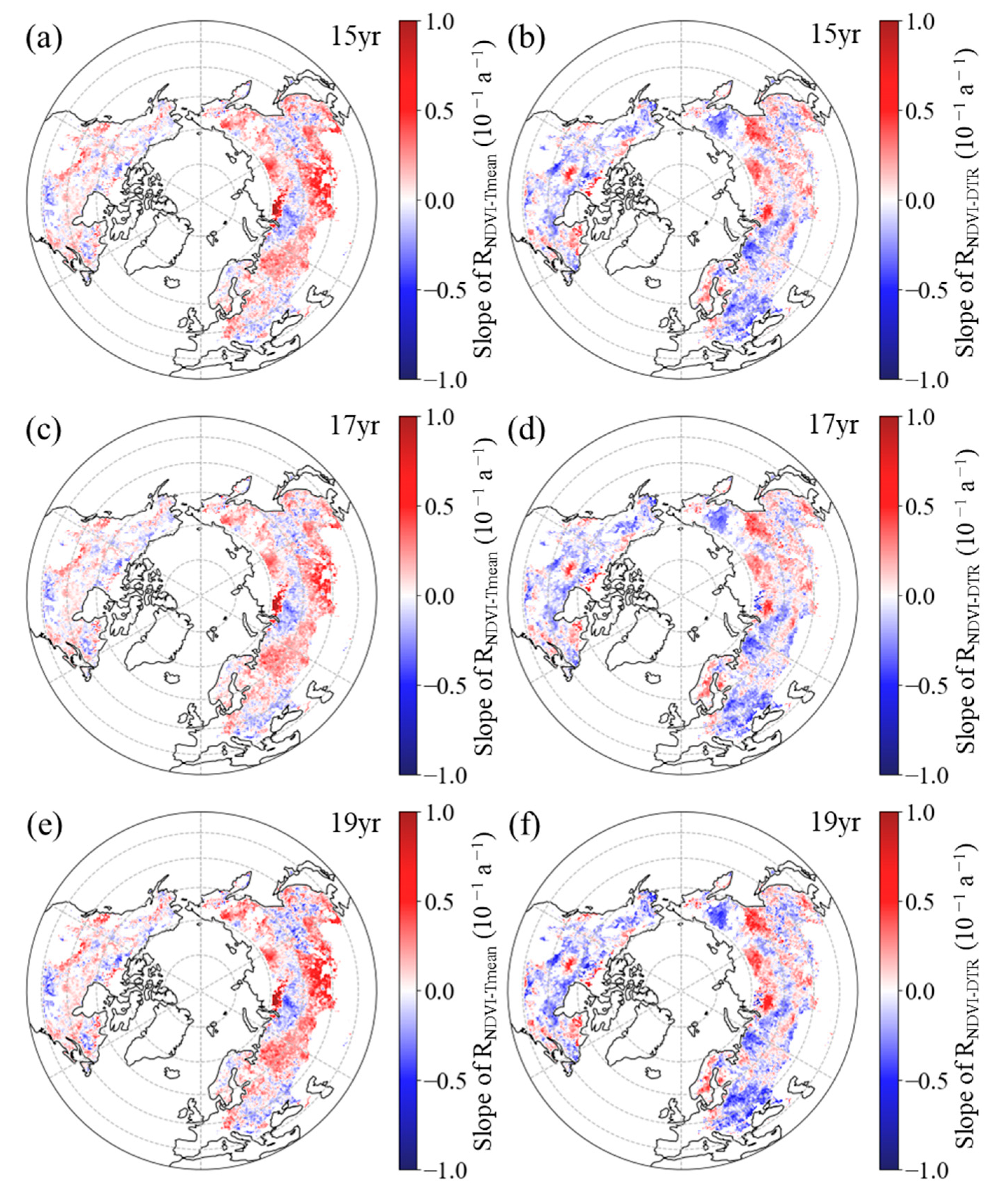
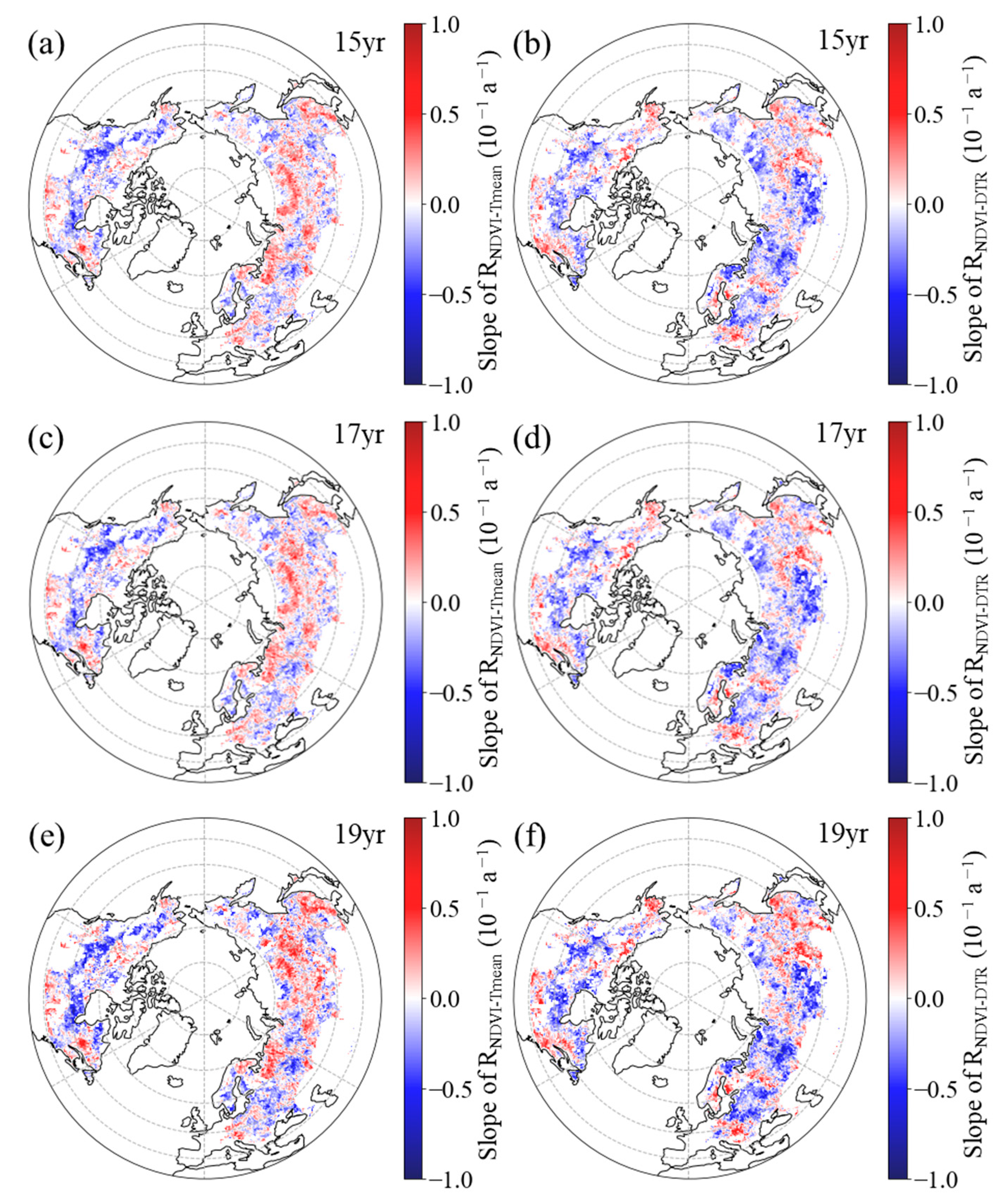
3.2.1. Inter-Annual Patterns of RNDVI−Tmean and RNDVI−DTR
3.2.2. Intra-Annual Patterns of RNDVI−Tmean and RNDVI−DTR
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B.; Pollard, D.; Thompson, S.L. Effects of boreal forest vegetation on global climate. Nature 1992, 359, 716–718. [Google Scholar] [CrossRef]
- Sulla-Menashe, D.; Woodcock, C.E.; Friedl, M.A. Canadian boreal forest greening and browning trends: An analysis of biogeographic patterns and the relative roles of disturbance versus climate drivers. Environ. Res. Lett. 2018, 13, 014007. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, L.; Hou, X.; Li, G.; Tian, Q.; Chan, E.; Ciais, P.; Yu, Q.; Yue, C. Fire Regime Impacts on Postfire Diurnal Land Surface Temperature Change Over North American Boreal Forest. J. Geophys. Res. Atmos. 2021, 126, e2021JD035589. [Google Scholar] [CrossRef]
- Deng, G.; Zhang, H.; Zhao, J.; Guo, X.; Biniyaz, E.; Zhao, H.; Guo, D.; Wu, R.; Li, H. Diverse variations in middle and high latitudes of the Northern Hemisphere spring phenology sensitivity to diurnal temperature during 1982–2015. Int. J. Clim. 2022, 42, 9385–9403. [Google Scholar] [CrossRef]
- Piao, S.; Nan, H.; Huntingford, C.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Peng, S.; Ahlström, A.; Canadell, J.G.; Cong, N.; et al. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nat. Commun. 2014, 5, 5018. [Google Scholar] [CrossRef]
- Tan, J.; Piao, S.; Chen, A.; Zeng, Z.; Ciais, P.; Janssens, I.A.; Mao, J.; Myneni, R.B.; Peng, S.; Peñuelas, J.; et al. Seasonally different response of photosynthetic activity to daytime and night-time warming in the Northern Hemisphere. Glob. Chang. Biol. 2015, 21, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Piao, S.; Janssens, I.A.; Zhu, Z.; Wang, T.; Wu, D.; Ciais, P.; Myneni, R.B.; Peaucelle, M.; Peng, S.; et al. Velocity of change in vegetation productivity over northern high latitudes. Nat. Ecol. Evol. 2017, 1, 1649–1654. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cook, B.I.; Allen, J.M.; Crimmins, T.M.; Betancourt, J.L.; Travers, S.E.; Pau, S.; Regetz, J.; Davies, T.J.; Kraft, N.J.B.; et al. Warming experiments underpredict plant phenological responses to climate change. Nature 2012, 485, 494–497. [Google Scholar] [CrossRef]
- Wu, Z.; Dijkstra, P.; Koch, G.W.; Peñuelas, J.; Hungate, B.A. Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef]
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shvidenko, A.Z.; Schepaschenko, D.G. Boreal forest health and global change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef]
- Du, Z.; Liu, X.; Wu, Z.; Zhang, H.; Zhao, J. Responses of Forest Net Primary Productivity to Climatic Factors in China during 1982–2015. Plants 2022, 11, 2932. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xiang, K.; Wu, Z.; Du, Z. Varying Responses of Vegetation Greenness to the Diurnal Warming across the Global. Plants 2022, 11, 2648. [Google Scholar] [CrossRef]
- Sun, X.; Ren, G.; You, Q.; Ren, Y.; Xu, W.; Xue, X.; Zhan, Y.; Zhang, S.; Zhang, P. Global diurnal temperature range (DTR) changes since 1901. Clim. Dyn. 2018, 52, 3343–3356. [Google Scholar] [CrossRef]
- Huang, Y.; Jiang, N.; Shen, M.; Guo, L. Effect of preseason diurnal temperature range on the start of vegetation growing season in the Northern Hemisphere. Ecol. Indic. 2020, 112, 106161. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.; Chen, A.; Wang, K.; Li, X.; Buermann, W.; Huntingford, C.; Peñuelas, J.; Xu, H.; Myneni, R.B. Seasonal biological carryover dominates northern vegetation growth. Nat. Commun. 2021, 12, 983. [Google Scholar] [CrossRef]
- Chen, X.; Yang, Y. Observed earlier start of the growing season from middle to high latitudes across the Northern Hemisphere snow-covered landmass for the period 2001–2014. Environ. Res. Lett. 2020, 15, 034042. [Google Scholar] [CrossRef]
- Shen, X.; Liu, B.; Henderson, M.; Wang, L.; Wu, Z.; Wu, H.; Jiang, M.; Lu, X. Asymmetric effects of daytime and nighttime warming on spring phenology in the temperate grasslands of China. Agric. For. Meteorol. 2018, 259, 240–249. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.B.; Chen, A.; Chevallier, F.; Dolman, A.J.; Janssens, I.A.; Peñuelas, J.; Zhang, G.; et al. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 2013, 501, 88–92. [Google Scholar] [CrossRef]
- Piao, S.; Wang, X.; Ciais, P.; Zhu, B.; Wang, T.A.O.; Liu, J.I.E. Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Glob. Chang. Biol. 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, X.; Pei, F.; Li, X.; Du, G. Non-uniform time-lag effects of terrestrial vegetation responses to asymmetric warming. Agric. For. Meteorol. 2018, 252, 130–143. [Google Scholar] [CrossRef]
- Cong, N.; Shen, M.; Yang, W.; Yang, Z.; Zhang, G.; Piao, S. Varying responses of vegetation activity to climate changes on the Tibetan Plateau grassland. Int. J. Biometeorol. 2017, 61, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Jin, N.; Zhao, J.; Feng, M.; Wang, C. Dynamics of the Response of Vegetation Activity to Air Temperature Change in Temperate China. Atmosphere 2022, 13, 1574. [Google Scholar] [CrossRef]
- Piao, S.L.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21, GB3018. [Google Scholar] [CrossRef]
- Zheng, B.; Ciais, P.; Chevallier, F.; Chuvieco, E.; Chen, Y.; Yang, H.; Lee, L.; Oliva, A.B.P.; Sato, K.; Martin-Pintado, N.; et al. Increasing forest fire emissions despite the decline in global burned area. Sci. Adv. 2021, 7, eabh2646. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Edith, B.A.I.; Xue, B. A review of influences of land use and land cover change on ecosystems. Chin. J. Plant Ecol. 2020, 44, 543–552. [Google Scholar] [CrossRef]
- Epstein, H.E.; Myers-Smith, I.; Walker, D.A. Recent dynamics of arctic and sub-arctic vegetation. Environ. Res. Lett. 2013, 8, 015040. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNDVI−Tmean | RNDVI−DTR | |||
|---|---|---|---|---|
| Positive | Negative | Positive | Negative | |
| 15-year | 40.2% (24.0%) | 59.8% (41.2%) | 40.9% (24.9%) | 59.1% (40.4%) |
| 17-year | 37.6% (22.3%) | 62.4% (44.4%) | 40.9% (25.8%) | 59.1% (41.2%) |
| 19-year | 40.0% (22.7%) | 60.0% (40.2%) | 41.4% (24.8%) | 58.6% (39.0%) |
| RNDVI−Tmean | RNDVI−DTR | |||
|---|---|---|---|---|
| Positive | Negative | Positive | Negative | |
| 15-year | 63.3% (45.7%) | 36.7% (22.1%) | 46.3% (28.0%) | 53.7% (34.0%) |
| 17-year | 61.7% (43.0%) | 38.3% (23.2%) | 46.1% (28.0%) | 53.9% (35.0%) |
| 19-year | 61.8% (42.0%) | 38.2% (22.1%) | 44.7% (25.3%) | 55.3% (34.5%) |
| RNDVI−Tmean | RNDVI−DTR | |||
|---|---|---|---|---|
| Positive | Negative | Positive | Negative | |
| 15-year | 39.8% (23.4%) | 60.2% (41.7%) | 45.7% (28.8%) | 54.3% (36.0%) |
| 17-year | 37.6% (22.0%) | 62.4% (45.2%) | 45.2% (28.7%) | 54.8% (36.9%) |
| 19-year | 39.8% (22.2%) | 60.2% (40.6%) | 46.5% (28.2%) | 53.5% (34.7%) |
| RNDVI−Tmean | RNDVI−DTR | |||
|---|---|---|---|---|
| Positive | Negative | Positive | Negative | |
| 15-year | 49.4% (30.4%) | 50.6% (31.5%) | 39.9% (23.0%) | 60.1% (40.8%) |
| 17-year | 48.8% (30.0%) | 51.3% (32.3%) | 39.8% (23.5%) | 60.2% (40.9%) |
| 19-year | 47.8% (29.4%) | 52.2% (32.5%) | 42.5% (24.4%) | 57.5% (37.2%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Jin, N.; Wang, X.; Fu, G.; Xiang, K.; Wang, L.; Zhao, J. The Seasonal Divergence in the Weakening Relationship between Interannual Temperature Changes and Northern Boreal Vegetation Activity. Plants 2023, 12, 2447. https://doi.org/10.3390/plants12132447
Zhao H, Jin N, Wang X, Fu G, Xiang K, Wang L, Zhao J. The Seasonal Divergence in the Weakening Relationship between Interannual Temperature Changes and Northern Boreal Vegetation Activity. Plants. 2023; 12(13):2447. https://doi.org/10.3390/plants12132447
Chicago/Turabian StyleZhao, Haijiang, Ning Jin, Xiurong Wang, Guiqin Fu, Kunlun Xiang, Liang Wang, and Jie Zhao. 2023. "The Seasonal Divergence in the Weakening Relationship between Interannual Temperature Changes and Northern Boreal Vegetation Activity" Plants 12, no. 13: 2447. https://doi.org/10.3390/plants12132447
APA StyleZhao, H., Jin, N., Wang, X., Fu, G., Xiang, K., Wang, L., & Zhao, J. (2023). The Seasonal Divergence in the Weakening Relationship between Interannual Temperature Changes and Northern Boreal Vegetation Activity. Plants, 12(13), 2447. https://doi.org/10.3390/plants12132447