
Exploring Antimicrobial Peptides Efficacy against Fire Blight (Erwinia amylovora)

 , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Results and Discussion
2.1. Screening of Potential AMPs against E. amylovora
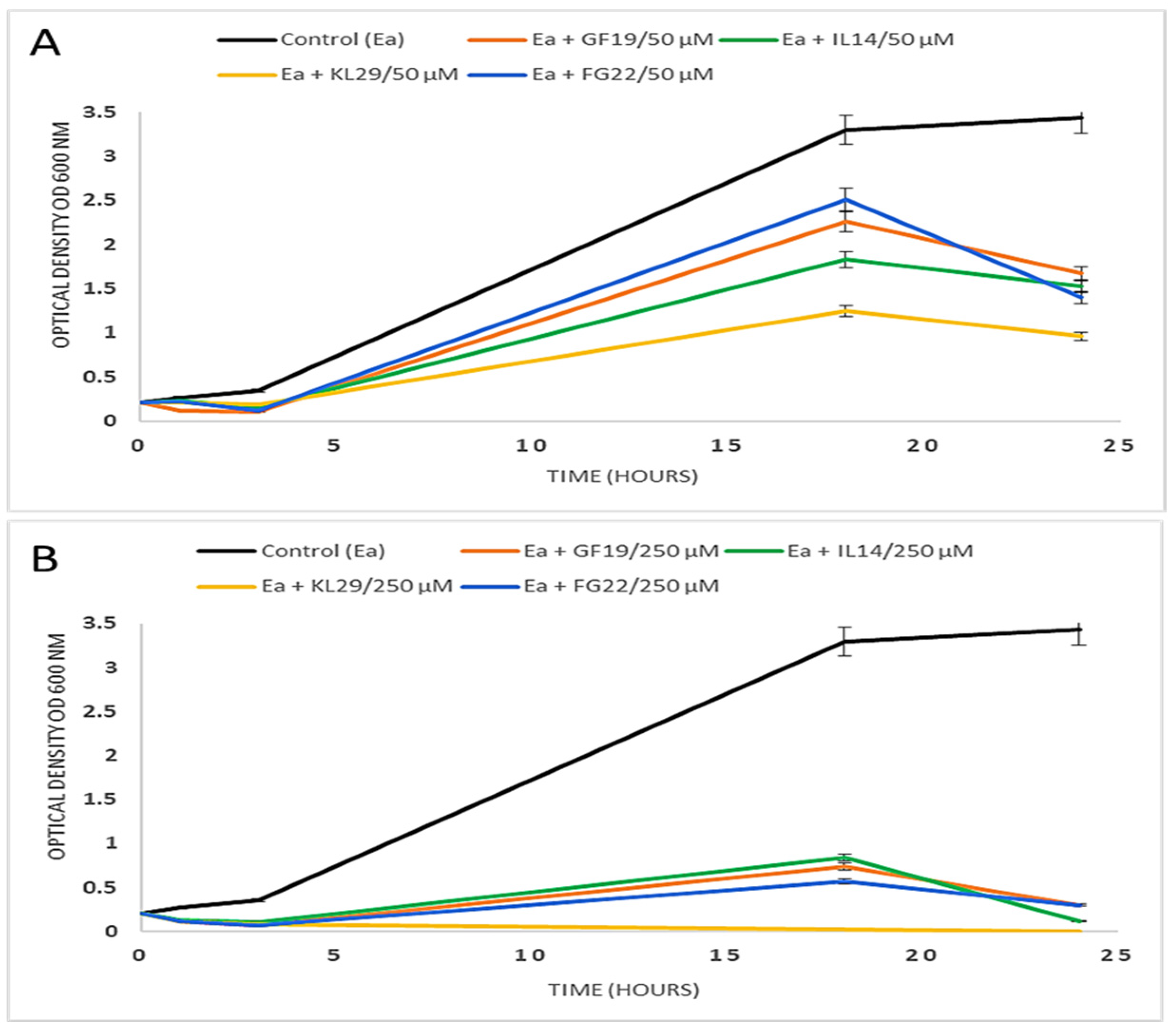
2.2. Effect of Selected AMPs on E. amylovora
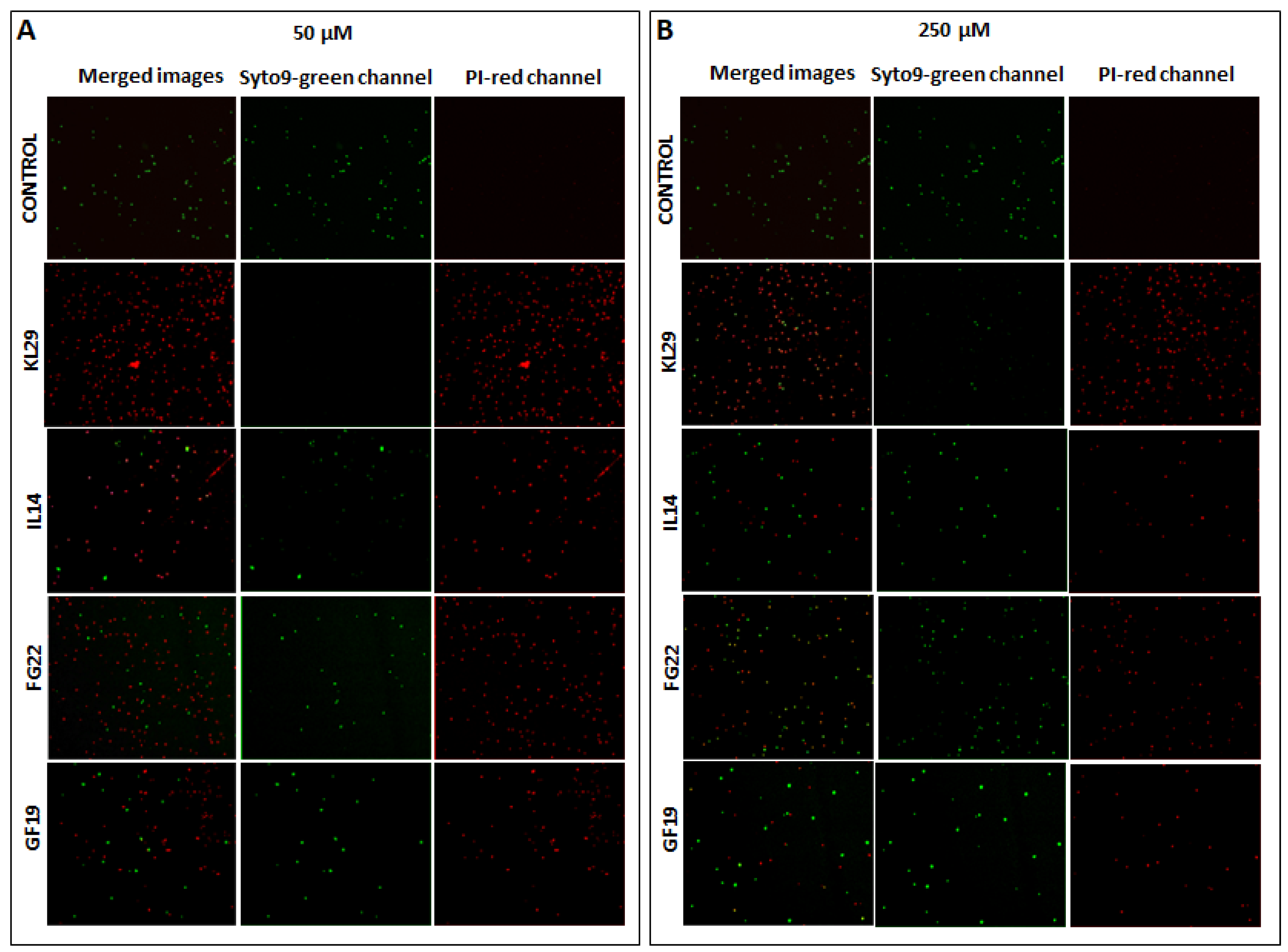
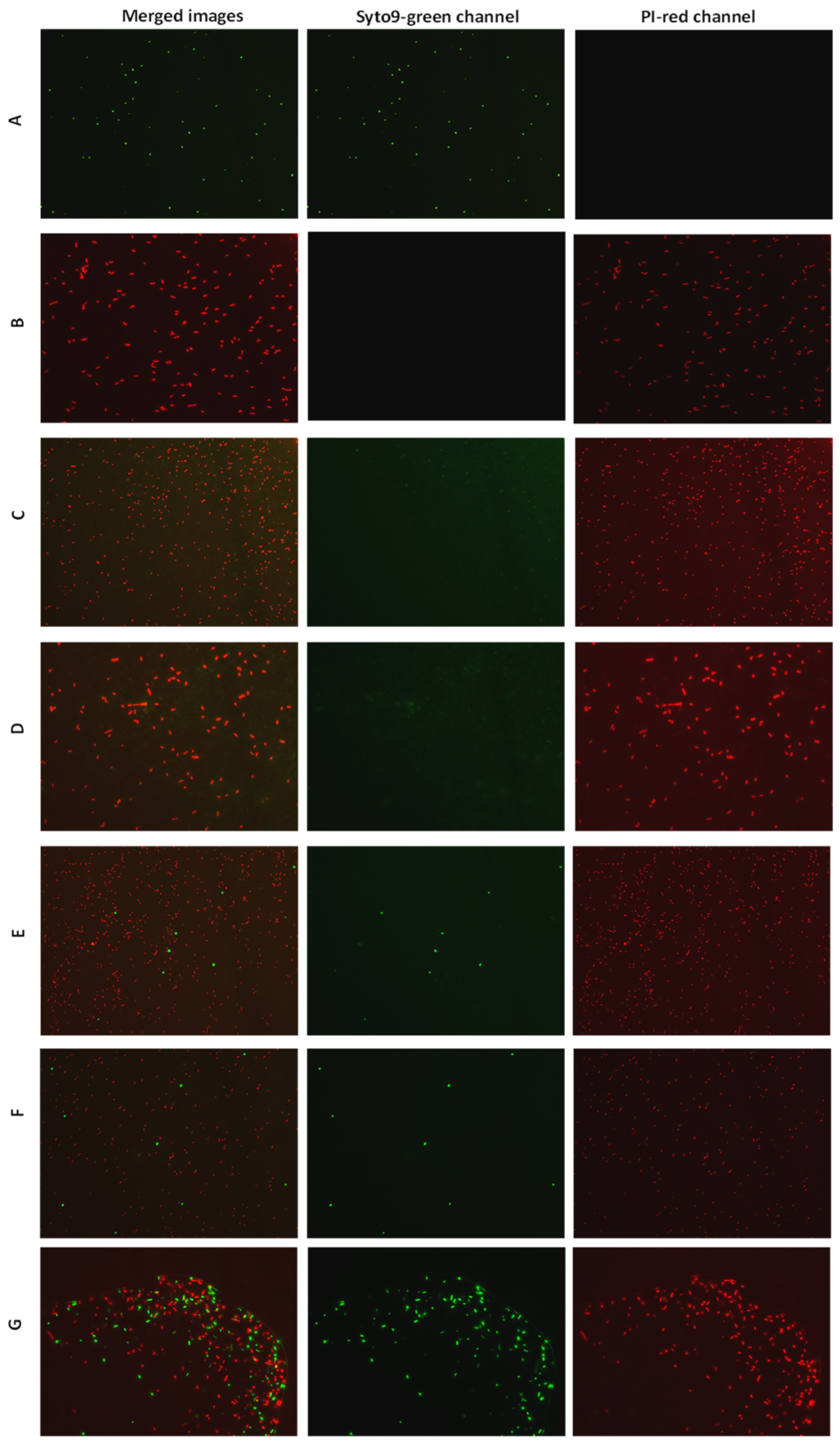
2.3. Bactericidal Activity of Selected AMPs
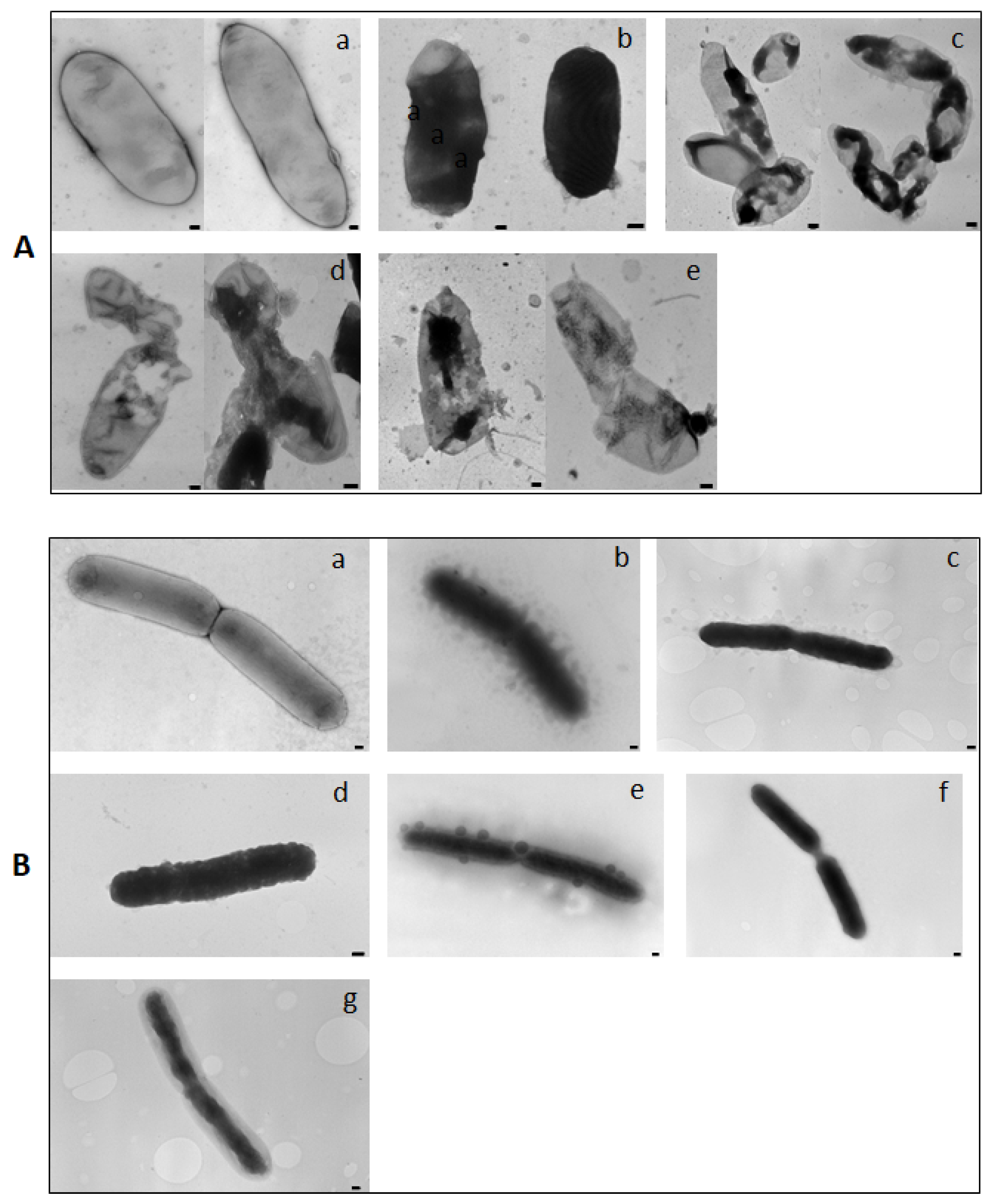
2.4. Lytic and Non-Lytic Activity of KL29
2.5. Selected AMPs Innocuous for Eukaryotic Cells
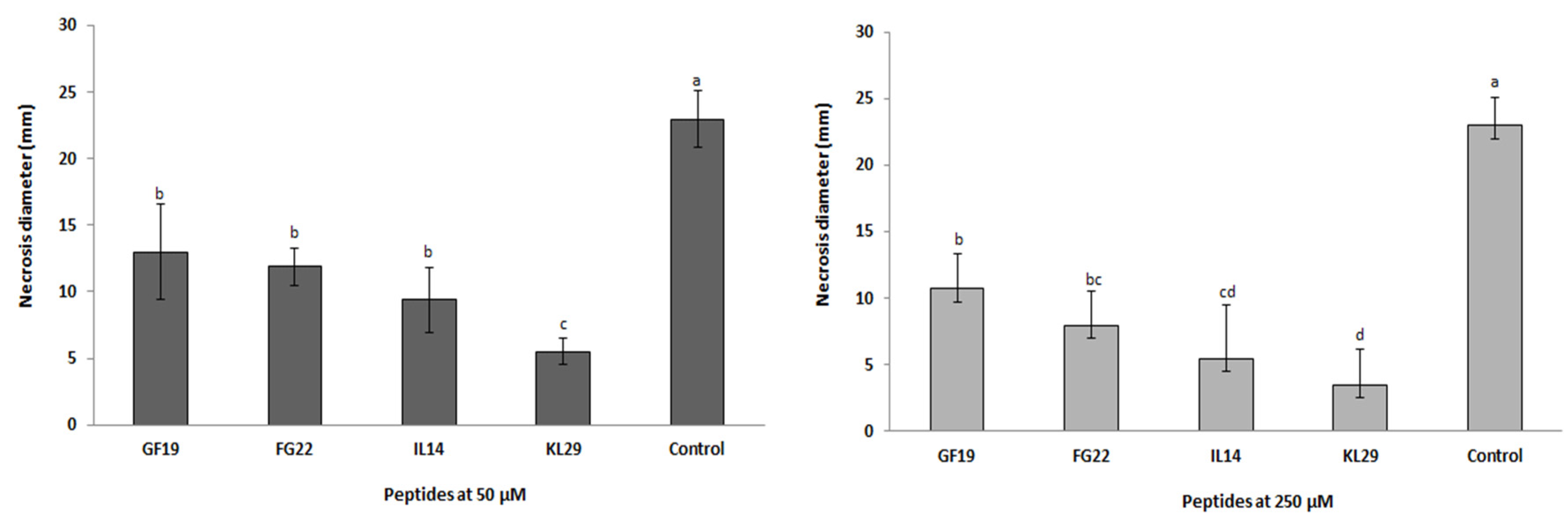
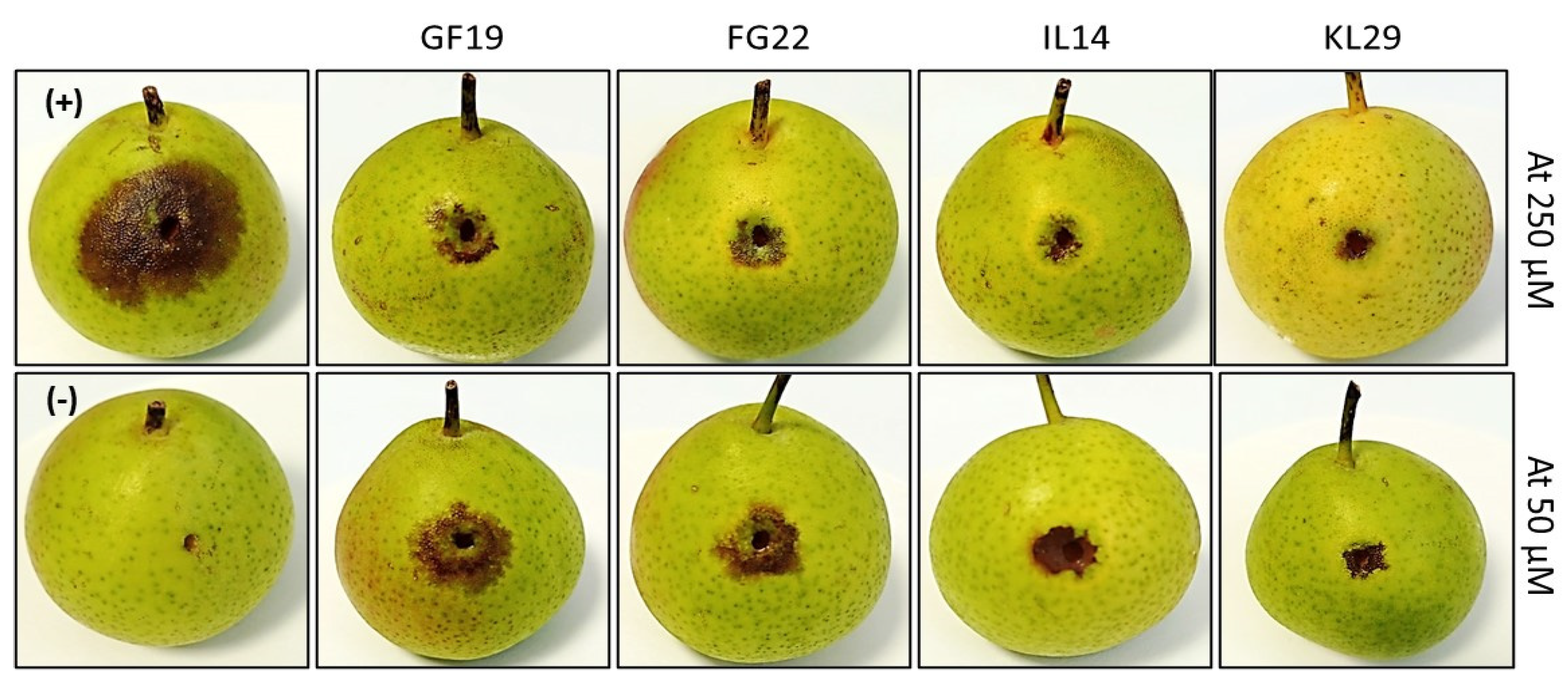
2.6. In Vivo Antibacterial Activity of Selected AMPs against E. amylovora Infection
3. Materials and Methods
3.1. AMP Synthesis and Bacterial Strains
3.2. Screening of Potential AMPs against E. amylovora
3.3. Turbidity Assay
3.4. Viable-Quantitative PCR (v-qPCR) and Fluorescence Microscopy (FM)
3.5. Transmission Electron Microscopy (TEM)
3.6. Evaluation of AMPs Toxicity
3.7. In Vivo Antibacterial Activity of AMPs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mendes, R.J.; Regalado, L.; Luz, J.P.; Tassi, N.; Teixeira, C.; Gomes, P.; Tavares, F.; Santos, C. In Vitro Evaluation of Five Antimicrobial Peptides against the Plant Pathogen Erwinia Amylovora. Biomolecules 2021, 11, 554. [Google Scholar] [CrossRef] [PubMed]
- Farkas, Á.; Mihalik, E.; Dorgai, L.; Bubán, T. Floral Traits Affecting Fire Blight Infection and Management. Trees 2012, 26, 47–66. [Google Scholar] [CrossRef]
- Zhao, Y.; Tian, Y.; Wang, L.; Geng, G.; Zhao, W.; Hu, B.; Zhao, Y. Fire Blight Disease, a Fast-Approaching Threat to Apple and Pear Production in China. J. Integr. Agric. 2019, 18, 815–820. [Google Scholar] [CrossRef]
- Pirc, M.; Ravnikar, M.; Dreo, T. Sensitivity Of Enrichment Isolation For Fire Blight Detection In Composite Twig Samples. Acta Hortic. 2014, 1056, 85–90. [Google Scholar] [CrossRef]
- Mendes, R.J.; Mariz-Ponte, N.; Correia, C.V.; Dias, M.C.; Sousa, M.L.D.; Tavares, F.; Santos, C. Fire Blight Management: Physiological Assessment of Cultural Control By Pruning in Pear Orchards. Agriculture 2020, 66, 128–136. [Google Scholar] [CrossRef]
- Sundin, G.W.; Wang, N. Antibiotic Resistance in Plant-Pathogenic Bacteria. Annu. Rev. Phytopathol. 2018, 56, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Sabri, M.; Benkirane, R.; Habbadi, K.; Sadik, S.; Ou-Zine, M.; Diouri, M.; Achbani, E.H. Phages as a Potential Biocontrol of Phytobacteria. Arch. Phytopathol. Plant Prot. 2021, 54, 1277–1291. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Tarighi, S.; Taheri, P. Effects of Plant Essential Oils on Growth and Virulence Factors of Erwinia Amylovora. J. Plant Pathol. 2020, 102, 409–419. [Google Scholar] [CrossRef]
- Mikiciński, A.; Puławska, J.; Molzhigitova, A.; Sobiczewski, P. Bacterial Species Recognized for the First Time for Its Biocontrol Activity against Fire Blight (Erwinia Amylovora). Eur. J. Plant Pathol. 2020, 156, 257–272. [Google Scholar] [CrossRef]
- Huang, Y.-T.; Kumar, S.R.; Chan, H.-C.; Jhan, Z.-H.; Chen, D.W.; Lue, S.J. Efficacy of Antimicrobial Peptides (AMPs) against Escherichia Coli and Bacteria Morphology Change after AMP Exposure. J. Taiwan Inst. Chem. Eng. 2021, 126, 307–312. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Xue, Z.; Jia, Y.; Li, R.; He, C.; Chen, H. The Structure-Mechanism Relationship and Mode of Actions of Antimicrobial Peptides: A Review. Trends Food Sci. Technol. 2021, 109, 103–115. [Google Scholar] [CrossRef]
- Menousek, J.; Mishra, B.; Hanke, M.L.; Heim, C.E.; Kielian, T.; Wang, G. Database Screening and in Vivo Efficacy of Antimicrobial Peptides against Methicillin-Resistant Staphylococcus Aureus USA300. Int. J. Antimicrob. Agents 2012, 39, 402–406. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente-Núñez, C.; Korolik, V.; Bains, M.; Nguyen, U.; Breidenstein, E.B.M.; Horsman, S.; Lewenza, S.; Burrows, L.; Hancock, R.E.W. Inhibition of Bacterial Biofilm Formation and Swarming Motility by a Small Synthetic Cationic Peptide. Antimicrob. Agents Chemother. 2012, 56, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Badosa, E.; Moiset, G.; Montesinos, L.; Talleda, M.; Bardají, E.; Feliu, L.; Planas, M.; Montesinos, E. Derivatives of the Antimicrobial Peptide BP100 for Expression in Plant Systems. PLoS ONE 2013, 8, e85515. [Google Scholar] [CrossRef] [PubMed]
- Moll, L.; Badosa, E.; Planas, M.; Feliu, L.; Montesinos, E.; Bonaterra, A. Antimicrobial Peptides With Antibiofilm Activity Against Xylella Fastidiosa. Front. Microbiol. 2021, 12, 3243. [Google Scholar] [CrossRef] [PubMed]
- Moll, L.; Baró, A.; Montesinos, L.; Badosa, E.; Bonaterra, A.; Montesinos, E. Induction of Defense Responses and Protection of Almond Plants Against Xylella Fastidiosa by Endotherapy with a Bifunctional Peptide. Phytopathology 2022, 112, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Elbeaino, T.; Incerti, O.; Dakroub, H.; Valentini, F.; Huang, Q. Development of an FTP-LAMP Assay Based on TaqMan Real-Time PCR and LAMP for the Specific Detection of Xylella Fastidiosa De Donno and Mulberry Strains in Both Plants and Insect Vectors. J. Microbiol. Methods 2020, 175, 105992. [Google Scholar] [CrossRef] [PubMed]
- Laforest, M.; Bisaillon, K.; Ciotola, M.; Cadieux, M.; Hébert, P.-O.; Toussaint, V.; Svircev, A.M. Rapid Identification of Erwinia Amylovora and Pseudomonas Syringae Species and Characterization of E. Amylovora Streptomycin Resistance Using Quantitative PCR Assays. Can. J. Microbiol. 2019, 65, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, L.; Gascón, B.; Ruz, L.; Badosa, E.; Planas, M.; Feliu, L.; Montesinos, E. A Bifunctional Synthetic Peptide with Antimicrobial and Plant Elicitation Properties that Protect Tomato Plants from Bacterial and Fungal Infections. Front. Plant Sci. 2021, 12, 756357. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Code | Sequence | Origin | References |
|---|---|---|---|---|
| Ascaphin-8 | GF19 | GFKDLLKGAAKALVKTVLF-NH2 | Frog | [12] |
| DASamP1 | FF13 | FFGKVLKLIRKIF-NH2 | Synthetic | [12] |
| DASamP2 | IL14 | IKWKKLLRAAKRIL-NH2 | Synthetic | [12] |
| Lycotoxin I | IL25 | IWLTALKFLGKHAAKHLAKQQLSKL | Spider | [12] |
| Maculatin 1.3 | GF21 | GLLGLLGSVVSHVVPAIVGHF-NH2 | Frog | [12] |
| Piscidin 1 | FG22 | FFHHIFRGIVHVGKTIHRLVTG | Fish | [12] |
| 1036 | VK13 | VQFRIRVRIVIRK-NH2 | Synthetic | [13] |
| BP178 | KL29 | KKLFKKILKYL-AGPA-GIGKFLHSAK-KDEL-OH | Synthetic | [14] |
| RIJK2 | RV12 | RIVWVRIRRWFV-NH2 | Synthetic | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabri, M.; El Handi, K.; Valentini, F.; De Stradis, A.; Achbani, E.H.; Benkirane, R.; Elbeaino, T. Exploring Antimicrobial Peptides Efficacy against Fire Blight (Erwinia amylovora). Plants 2023, 12, 113. https://doi.org/10.3390/plants12010113
Sabri M, El Handi K, Valentini F, De Stradis A, Achbani EH, Benkirane R, Elbeaino T. Exploring Antimicrobial Peptides Efficacy against Fire Blight (Erwinia amylovora). Plants. 2023; 12(1):113. https://doi.org/10.3390/plants12010113
Chicago/Turabian StyleSabri, Miloud, Kaoutar El Handi, Franco Valentini, Angelo De Stradis, El Hassan Achbani, Rachid Benkirane, and Toufic Elbeaino. 2023. "Exploring Antimicrobial Peptides Efficacy against Fire Blight (Erwinia amylovora)" Plants 12, no. 1: 113. https://doi.org/10.3390/plants12010113
APA StyleSabri, M., El Handi, K., Valentini, F., De Stradis, A., Achbani, E. H., Benkirane, R., & Elbeaino, T. (2023). Exploring Antimicrobial Peptides Efficacy against Fire Blight (Erwinia amylovora). Plants, 12(1), 113. https://doi.org/10.3390/plants12010113