
Extracellular Vesicle-Mediated Secretion of Protochlorophyllide in the Cyanobacterium Leptolyngbya boryana


Abstract
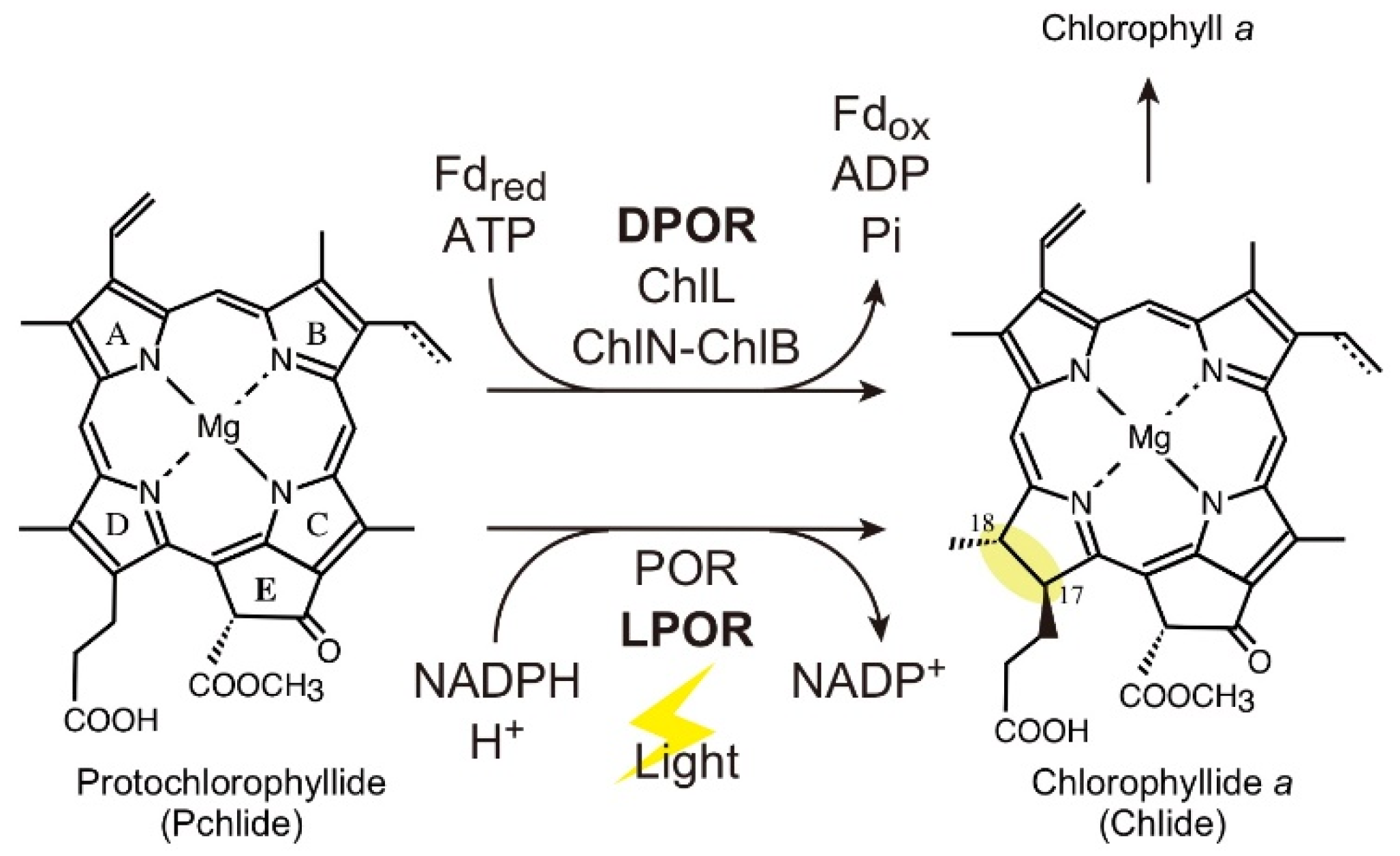
:1. Introduction
2. Results
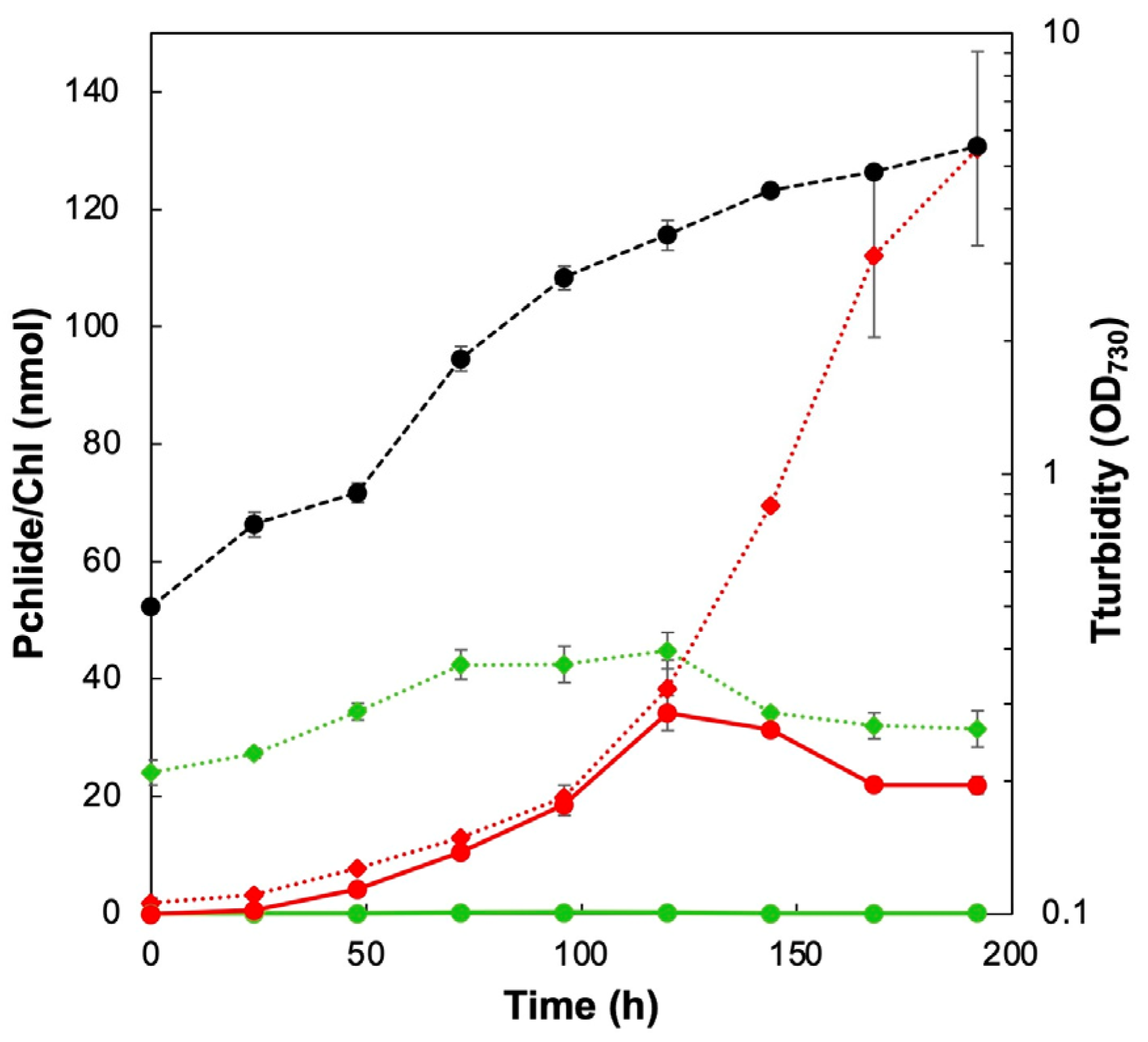
2.1. Accumulation of Pchlide in Culture Media
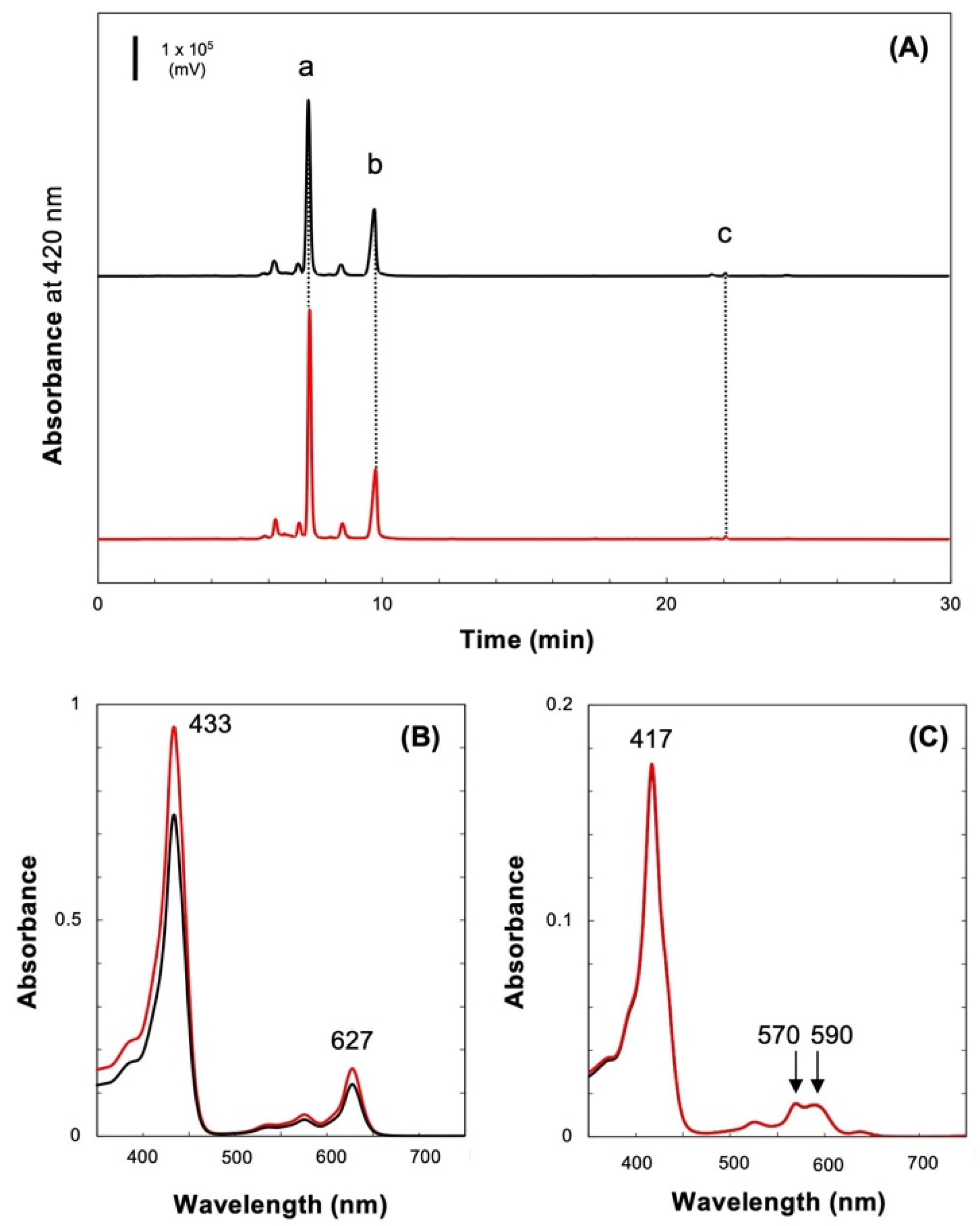
2.2. Fractionation of Pchlide in the Culture Medium
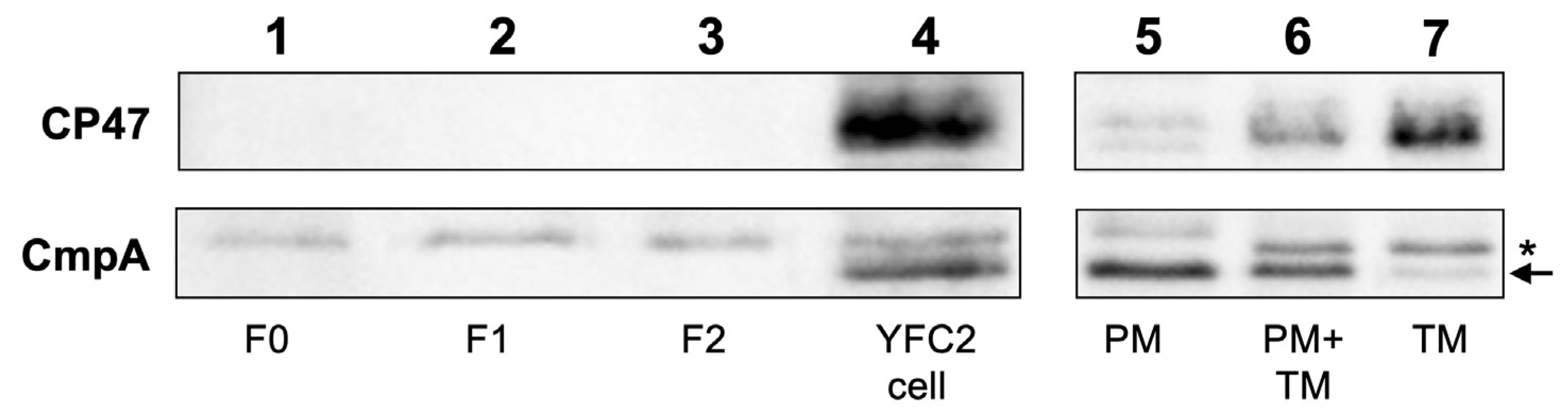
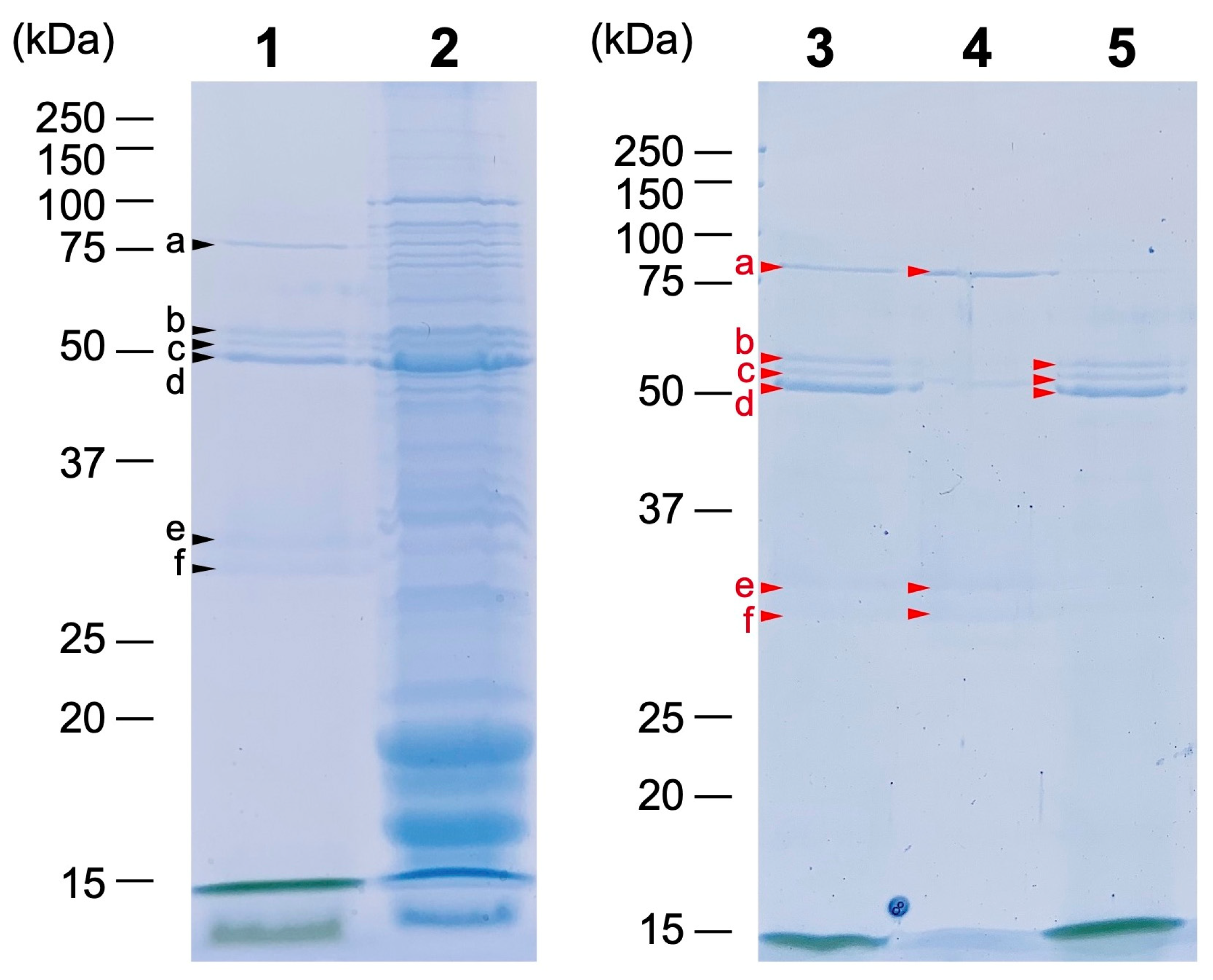
2.3. Protein Composition of the Pchlide Fractions
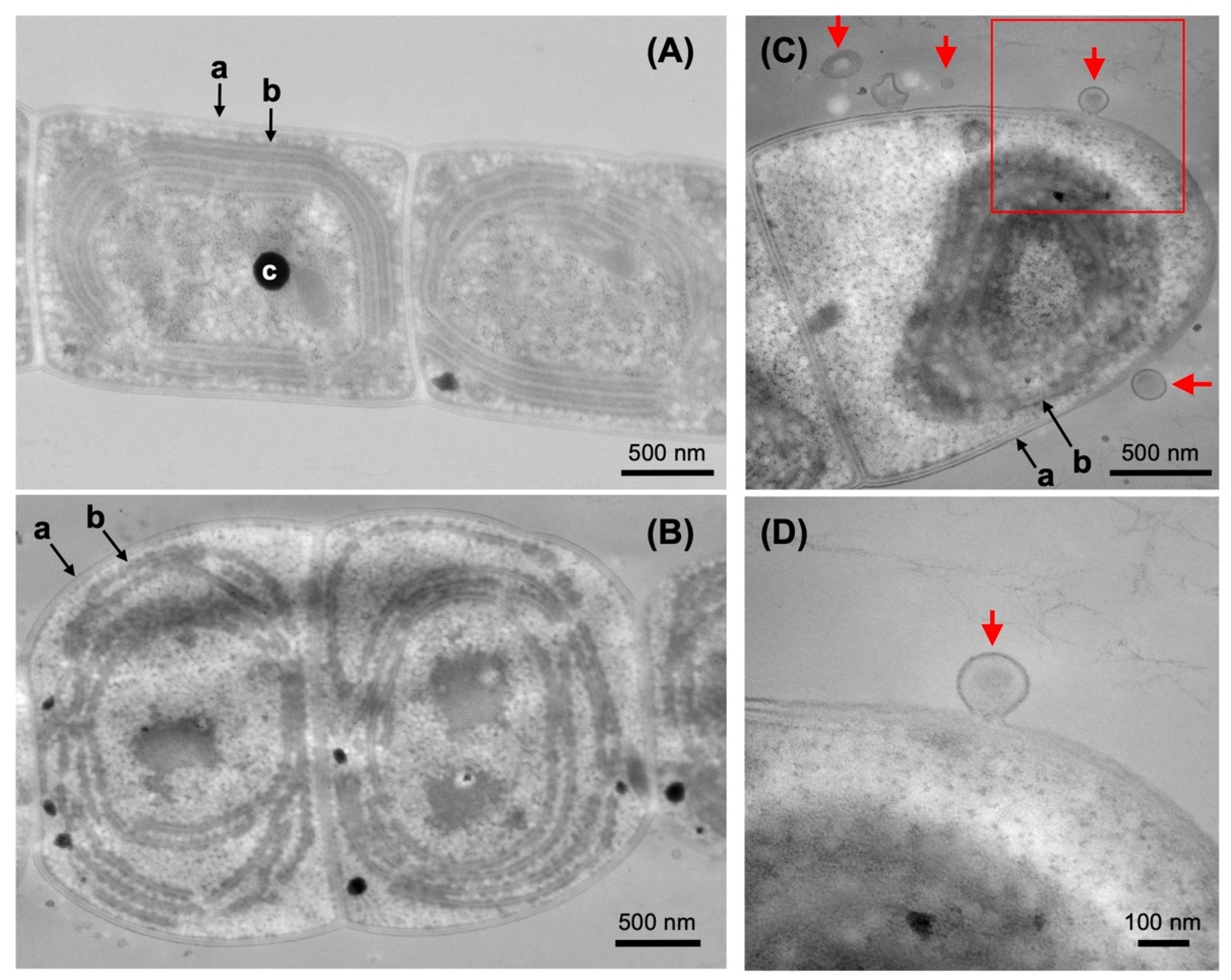
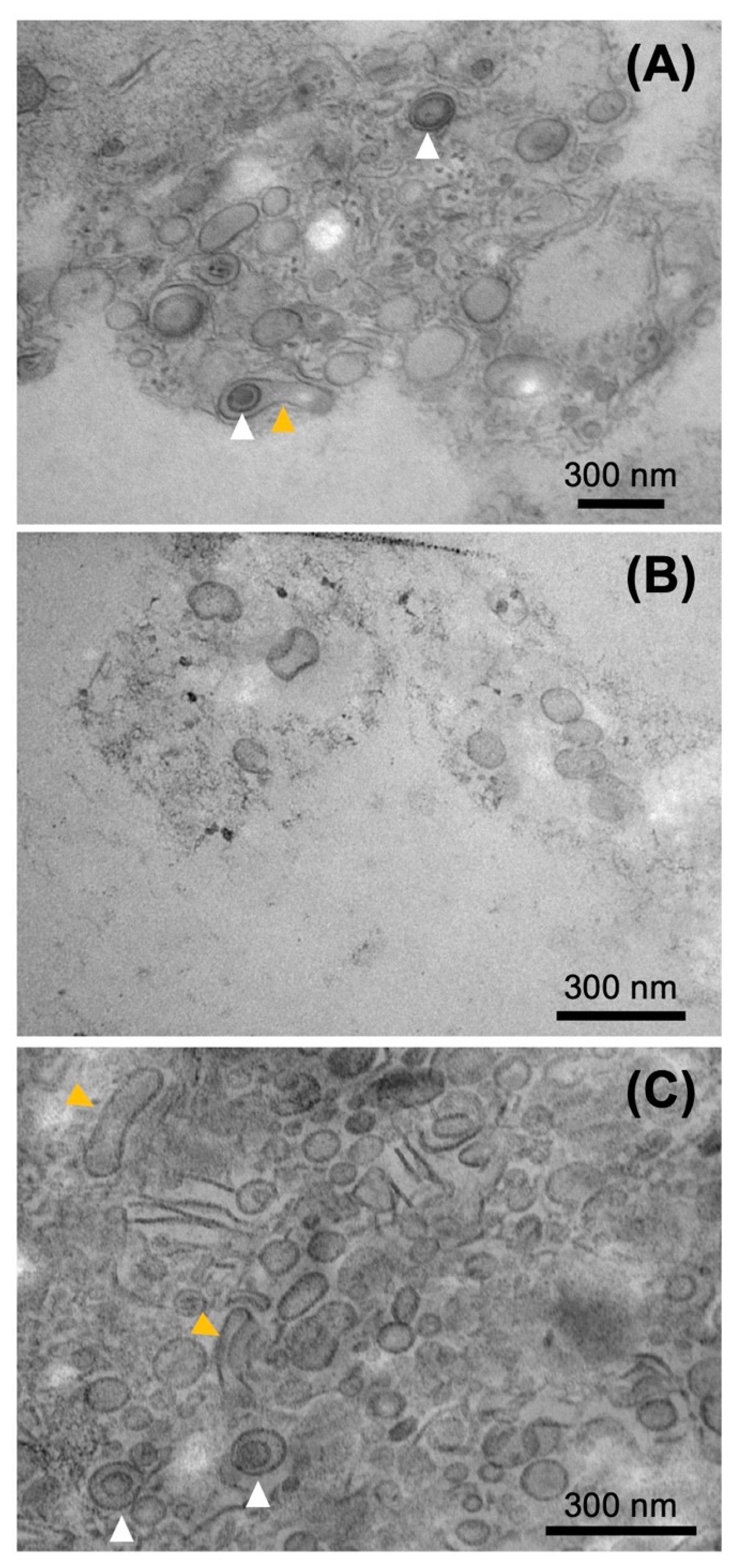
2.4. Electron Microscopy of the Pchlide Fractions
3. Discussion
4. Materials and Methods
4.1. Cyanobacterial Strains and Growth Conditions
4.2. Preparation of Pigments in the Culture Media
4.3. Preparation of Pchlide Fractions from Culture Media Using Sucrose Density Gradient Centrifugation
4.4. Pigment Analysis of the Culture Media
4.5. Preparation of Membrane Fractions from L. boryana Cells
4.6. SDS-PAGE and Western Blot Analysis
4.7. Mass Spectroscopic Analysis
4.8. Transmission Electron Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fujita, Y.; Yamakawa, H. Biochemistry of chlorophyll biosynthesis in photosynthetic prokaryotes. In Modern Topics in the Phototrophic Prokaryotes–Metabolism, Bioenergetics, and Omics; Hallenbeck, P., Ed.; Springer: Cham, Switzerland, 2017; pp. 67–122. [Google Scholar]
- Willows, R.D. The Mg branch of chlorophyll synthesis: Biosynthesis of chlorophyll a from protoporphyrin IX. In Advances in Botanical Research, Volume 90: Metabolism, Structure and Function of Plant Tetrapyrroles: Introduction, Microbial and Eukaryotic Chlorophyll Synthesis and Catabolism; Grimm, B., Ed.; Elsevier: London, UK, 2019; pp. 141–182. [Google Scholar]
- Czarnecki, O.; Grimm, B. Post-translational control of tetrapyrrole biosynthesis in plants, algae, and cyanobacteria. J. Exp. Bot. 2012, 63, 1675–1687. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Grimm, B. Organization of chlorophyll biosynthesis and insertion of chlorophyll into the chlorophyll-binding proteins in chloroplasts. Photosynth. Res. 2015, 126, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Komenda, J.; Sobotka, R. Chlorophyll-binding subunits of photosystem I and II: Biosynthesis, chlorophyll incorporation and assembly. In Advances in Botanical Research, volume 91: Metabolism, Structure and Function of Plant Tetrapyrroles: Control Mechanisms of Chlorophyll Biosynthesis and Analysis of Chlorophyll-binding Proteins; Grimm, B., Ed.; Elsevier: London, UK, 2019; pp. 195–223. [Google Scholar]
- Wang, P.; Grimm, B. Connecting chlorophyll metabolism with accumulation of the photosynthetic apparatus. Trends Plant Sci. 2021, 26, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.C.; Melo, A.A.; Kruk, J.; Frost, A.; Gabruk, M. Photocatalytic LPOR forms helical lattices that shape membranes for chlorophyll synthesis. Nat. Plants 2021, 7, 437–444. [Google Scholar] [CrossRef]
- Heyes, D.J.; Zhang, S.; Taylor, A.; Johannissen, L.O.; Hardman, S.J.O.; Hay, S.; Scrutton, N.S. Photocatalysis as the ‘master switch’ of photomorphogenesis in early plant development. Nat. Plants 2021, 7, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Muraki, N.; Nomata, J.; Ebata, K.; Mizoguchi, T.; Shiba, T.; Tamiaki, H.; Kurisu, G.; Fujita, Y. X-ray crystal structure of the light-independent protochlorophyllide reductase. Nature 2010, 465, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Moser, J.; Lange, C.; Krausze, J.; Rebelein, J.; Schubert, W.D.; Ribbe, M.W.; Heinz, D.W.; Jahn, D. Structure of ADP-aluminum fluoride-stabilized protochlorophyllide oxidoreductase complex. Proc. Natl. Acad. Sci. USA 2013, 110, 2094–2098. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Takagi, H.; Hase, T. Cloning of the gene encoding a protochlorophyllide reductase: The physiological significance of the co-existence of light-dependent and -independent protochlorophyllide reduction systems in the cyanobacterium Plectonema boryanum. Plant Cell Physiol. 1998, 39, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, S.; Nomata, J.; Fujita, Y. Differential operation of dual protochlorophyllide reductases for chlorophyll biosynthesis in response to environmental oxygen levels in the cyanobacterium Leptolyngbya boryana. Plant Physiol. 2006, 142, 911–922. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Takahashi, Y.; Chuganji, M.; Matsubara, H. The nifH-like (frxC) gene is involved in the biosynthesis of chlorophyll in the filamentous cyanobacterium Plectonema boryanum. Plant Cell Physiol. 1992, 81, 81–92. [Google Scholar]
- Fujita, Y.; Matsumoto, H.; Takahashi, Y.; Matsubara, H. Identification of a nifDK-like gene (ORF467) involved in the biosynthesis of chlorophyll in the cyanobacterium Plectonema boryanum. Plant Cell Physiol. 1993, 34, 305–314. [Google Scholar] [PubMed]
- Wu, Q.; Vermaas, W.F. Light-dependent chlorophyll a biosynthesis upon chlL deletion in wild-type and photosystem I-less strains of the cyanobacterium Synechocystis sp. PCC 6803. Plant Mol. Biol. 1995, 29, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Takagi, H.; Hase, T. Identification of the chlB gene and the gene product essential for the light-independent chlorophyll biosynthesis in the cyanobacterium Plectonema boryanum. Plant Cell Physiol. 1996, 37, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Manning, A.J.; Kuehn, M.J. Functional advantages conferred by extracellular prokaryotic membrane vesicles. J. Mol. Microbiol. Biotechnol. 2013, 23, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, J.; Park, J.; Gho, Y.S. Gram-negative and Gram-positive bacterial extracellular vesicles. Semin. Cell. Dev. Biol. 2015, 40, 97–104. [Google Scholar] [CrossRef]
- Pardo, Y.A.; Florez, C.; Baker, K.M.; Schertzer, J.W.; Mahler, G.J. Detection of outer membrane vesicles in Synechocystis PCC 6803. FEMS Microbiol. Lett. 2015, 362, fnv163. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, D.; Lima, S.; Matinha-Cardoso, J.; Tamagnini, P.; Oliveira, P. The role of outer membrane protein(s) harboring SLH/OprB-domains in extracellular vesicles’ production in Synechocystis sp. PCC 6803. Plants 2021, 10, 2757. [Google Scholar] [CrossRef]
- Kada, S.; Koike, H.; Satoh, K.; Hase, T.; Fujita, Y. Arrest of chlorophyll synthesis and differential decrease of Photosystems I and II in a cyanobacterial mutant lacking light-independent protochlorophyllide reductase. Plant Mol. Biol. 2003, 51, 225–235. [Google Scholar] [CrossRef]
- Lee, E.Y.; Bang, J.Y.; Park, G.W.; Choi, D.-S.; Kang, J.S.; Kim, H.-J.; Park, K.-S.; Lee, J.-O.; Kim, Y.-K.; Kwon, K.-H.; et al. Global proteomic profiling of native outer membrane vesicles derived from Escherichia coli. Proteomics 2007, 7, 3143–3153. [Google Scholar] [CrossRef]
- Koropatkin, N.M.; Koppeanaal, D.W.; Pakrasi, H.B.; Smith, T.J. The structure of a cyanobacterial bicarbonate transport protein, CmpA. J. Biol. Chem. 2007, 282, 2606–2614. [Google Scholar] [CrossRef] [Green Version]
- Velling, T.; Kusche-Gullberg, M.; Sejersen, T.; Gullberg, D. cDNA cloning and chromosomal localization of human α11 integrin. A collagen-binding, I domain-containing, β1-associated integrin α-chain present in muscle tissues. J. Biol. Chem. 1999, 274, 25735–25742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koronakis, V.; Sharff, A.; Koronakis, E.; Luisi, B.; Hughes, C. Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nature 2000, 405, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Sondermann, H.; O’Toole, G.A. Type 1 does the two-step: Type 1 secretion substrates with a functional periplasmic intermediate. J. Bacteriol. 2018, 200, e00168-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Kojima-Ando, H.; Ohki, K.; Fujita, Y. Formation of prolamellar-body-like ultrastructures in etiolated cyanobacterial cells overexpressing light-dependent protochlorophyllide oxidoreductase in Leptolyngbya boryana. J. Gen. Appl. Microbiol. 2020, 66, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Zarantonello, V.; Silva, T.P.; Noyma, N.P.; Gamalier, J.P.; Mello, M.M.; Marinho, M.M.; Melo, R.C.N. The cyanobacterium Cylindrospermopsis raciborskii (CYRF-01) responds to environmental stresses with increased vesiculation detected at single-cell resolution. Front. Microbiol. 2018, 9, 272. [Google Scholar] [CrossRef] [Green Version]
- Takaki, K.; Tahara, Y.O.; Nakamichi, N.; Hasegawa, Y.; Shintani, M.; Ohkuma, M.; Miyata, M.; Futamata, H.; Tashiro, Y. Multilamellar and multivesicular outer membrane vesicles produced by a Buttiauxella agrestis tolB mutant. Appl. Environ. Microbiol. 2020, 86, e01131-20. [Google Scholar] [CrossRef]
- Heyes, D.J.; Ruban, A.V.; Wilks, H.M.; Hunter, C.N. Enzymology below 200 K: The kinetics and thermodynamics of the photochemistry catalyzed by protochlorophyllide oxidoreductase. Proc. Natl. Acad. Sci. USA 2002, 99, 11145–11150. [Google Scholar] [CrossRef] [Green Version]
- Warshan, D.; Espinoza, J.L.; Stuart, R.K.; Richter, R.A.; Kim, S.-Y.; Shapiro, N.; Woyke, T.; Kyrpides, N.C.; Barry, K.; Singan, V.; et al. Feathermoss and epiphytic Nostoc cooperate differently: Expanding the spectrum of plant-cyanobacteria symbiosis. ISME J. 2017, 11, 2821–2833. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.M.; Bauer, C.E. Rhodobacter capsulatus genes involved in early steps of the bacteriochlorophyll biosynthetic pathway. J. Bacteriol. 1990, 172, 5001–5010. [Google Scholar] [CrossRef] [Green Version]
- Bollivar, D.; Suzuki, J.; Beatty, J.; Dobrowolski, J.; Bauer, C. Directed mutational analysis of bacteriochlorophyll a biosynthesis in Rhodobacter capsulatus. J. Mol. Biol. 1994, 237, 622–640. [Google Scholar] [CrossRef]
- Bollivar, D.W.; Bauer, C.E. Association of tetrapyrrole intermediates in the bacteriochlorophyll a biosynthetic pathway with the major outer-membrane porin protein of Rhodobacter capsulatus. Biochem. J. 1992, 282, 471–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biller, S.J.; Schubotz, F.; Roggensack, S.E.; Thompson, A.W.; Summons, R.E.; Chisholm, S.W. Bacterial vesicles in marine ecosystems. Science 2014, 343, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Bergman, B.; Chen, B.; Zheng, S.; Xiang, G.; Rasmussen, U. Cellular responses in the cyanobacterial symbiont during its vertical transfer between plant generations in the Azolla microphylla symbiosis. New Phytol. 2009, 181, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Chen, C.-Y.; Liu, Y.W.; Tan, Y.-J.; Deng, Z.-L.; Yang, F.; Huang, F.-Y.; Wen, C.; Rao, S.-S.; Luo, M.-J.; et al. Synechococcus elongatus PCC7942 secretes extracellular vesicles to accelerate cutaneous wound healing by promoting angiogenesis. Theranostics 2019, 9, 2678–2693. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kulp, A.; Kuehn, M.J. Modulation of bacterial outer membrane vesicle production by envelope structure and content. BMC Micobiol. 2014, 14, 324. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, P.; Martins, N.M.; Santos, M.; Pinto, F.; Büttel, Z.; Couto, N.A.S.; Wright, P.C.; Tamagnini, P. The versatile TolC-like Slr1270 in the cyanobacterium Synechocystis sp. PCC 6803. Environ. Microbiol. 2016, 18, 486–502. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Zapata, M.; Rodoriguez, R.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Stejskal, K.; Potěšil, D.; Zdráhal, Z. Suppression of peptide sample losses in autosampler vials. J. Proteome Res. 2013, 12, 3057–3062. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Band 1 | Peptides 2 | Accession Number | Annotation | Locus Tag 3 | Predicted Mol. Mass (kDa) |
|---|---|---|---|---|---|---|
| 0 | a | 26 | WP_017290631.1 | FG-GAP repeat-containing protein | 07530 | 80.4 |
| b | 19 | WP_017287236.1 | Carbohydrate-selective porin, OprB family | 40860 | 58.4 | |
| 11 | WP_017287165.1 | Glutamine synthetase (GlnA) | 41490 | 65.3 | ||
| 11 | WP_144056229.1 | Outer membrane efflux protein (TolC) | 23750 | 52.8 | ||
| c | 41 | WP_017288757.1 | Cyanobacterial porin | 25860 | 61.4 | |
| 10 | WP_017287236.1 | Carbohydrate-selective porin | 40860 | 58.4 | ||
| 9 | WP_017287123.1 | Carbohydrate-selective porin, OprB family | 41910 | 58.4 | ||
| d | 21 | WP_017287123.1 | Carbohydrate-selective porin, OprB family | 41910 | 58.4 | |
| 10 | WP_017288757.1 | Cyanobacterial porin | 25860 | 61.4 | ||
| 8 | WP_026148594.1 | Carbohydrate-selective porin, OprB family | 34630 | 60.8 | ||
| 1 | a | 17 | WP_017290631.1 | FG-GAP repeat-containing protein | 07530 | 80.4 |
| 2 | b | 10 | WP_017287236.1 | Carbohydrate-selective porin, OprB family | 40860 | 58.4 |
| c | 15 | WP_017288757.1 | Cyanobacterial porin | 25860 | 61.4 | |
| d | 10 | WP_017287123.1 | Carbohydrate-selective porin, OprB family | 41910 | 58.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usui, K.; Yamamoto, H.; Oi, T.; Taniguchi, M.; Mori, H.; Fujita, Y. Extracellular Vesicle-Mediated Secretion of Protochlorophyllide in the Cyanobacterium Leptolyngbya boryana. Plants 2022, 11, 910. https://doi.org/10.3390/plants11070910
Usui K, Yamamoto H, Oi T, Taniguchi M, Mori H, Fujita Y. Extracellular Vesicle-Mediated Secretion of Protochlorophyllide in the Cyanobacterium Leptolyngbya boryana. Plants. 2022; 11(7):910. https://doi.org/10.3390/plants11070910
Chicago/Turabian StyleUsui, Kentaro, Haruki Yamamoto, Takao Oi, Mitsutaka Taniguchi, Hitoshi Mori, and Yuichi Fujita. 2022. "Extracellular Vesicle-Mediated Secretion of Protochlorophyllide in the Cyanobacterium Leptolyngbya boryana" Plants 11, no. 7: 910. https://doi.org/10.3390/plants11070910
APA StyleUsui, K., Yamamoto, H., Oi, T., Taniguchi, M., Mori, H., & Fujita, Y. (2022). Extracellular Vesicle-Mediated Secretion of Protochlorophyllide in the Cyanobacterium Leptolyngbya boryana. Plants, 11(7), 910. https://doi.org/10.3390/plants11070910