
Characterisation of Grains and Flour Fractions from Field Grown Transgenic Oil-Accumulating Wheat Expressing Oat WRI1

 ,
,  , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Field Trial of Genetically Modified Wheat in Sweden
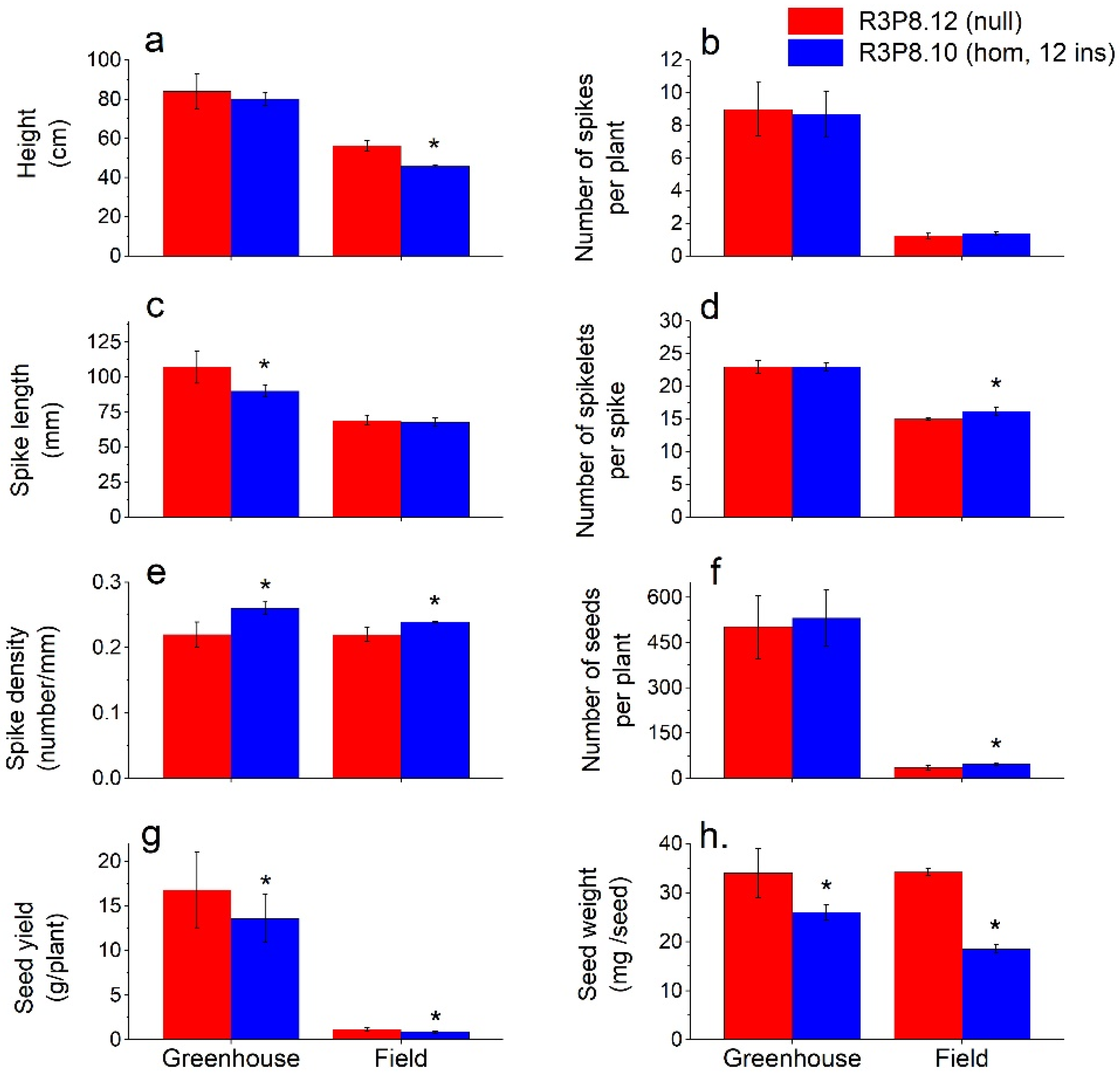
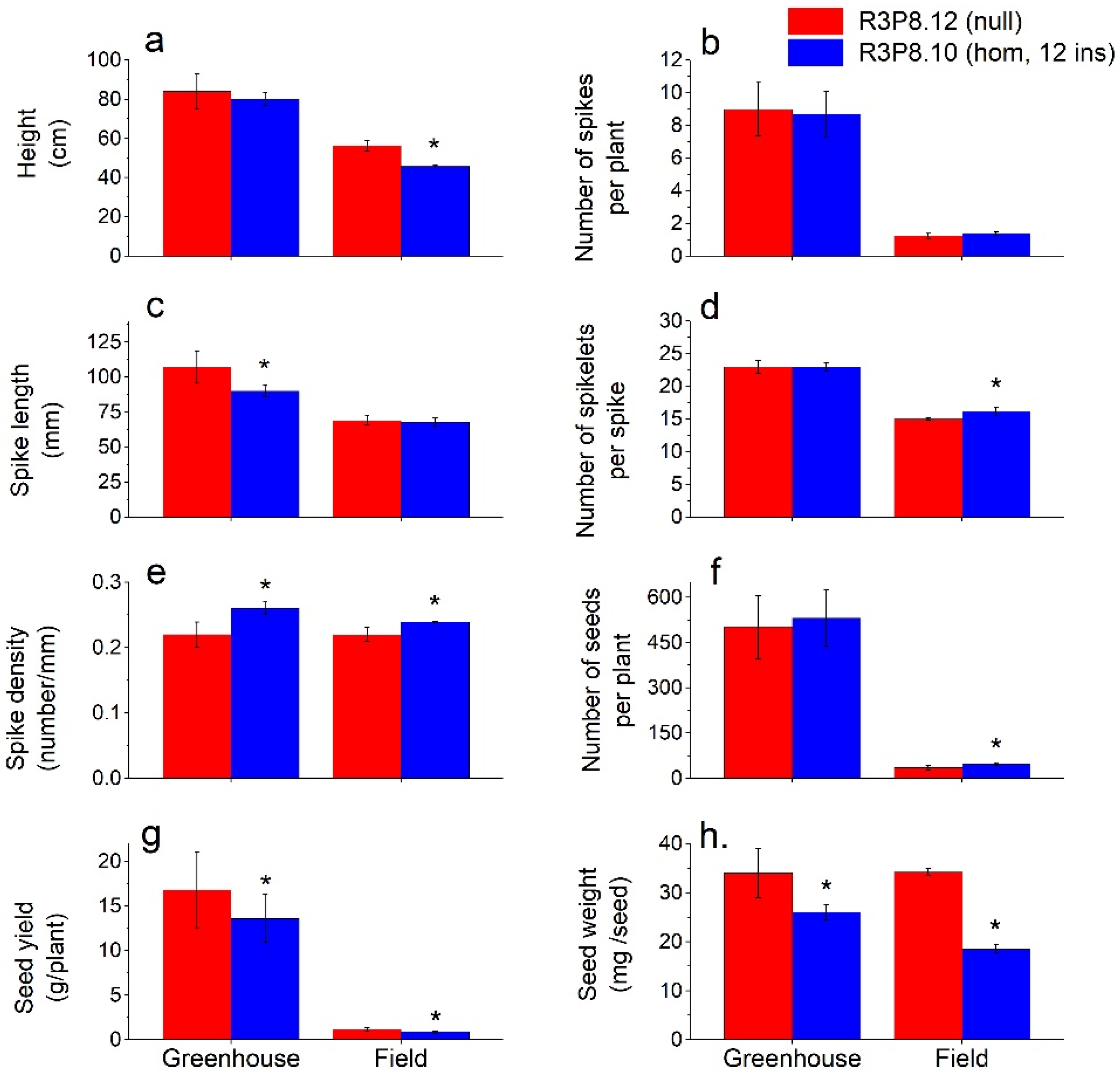
2.2. General Plant, Spike and Grain Characteristics from Field Grown and Greenhouse Grown Plants
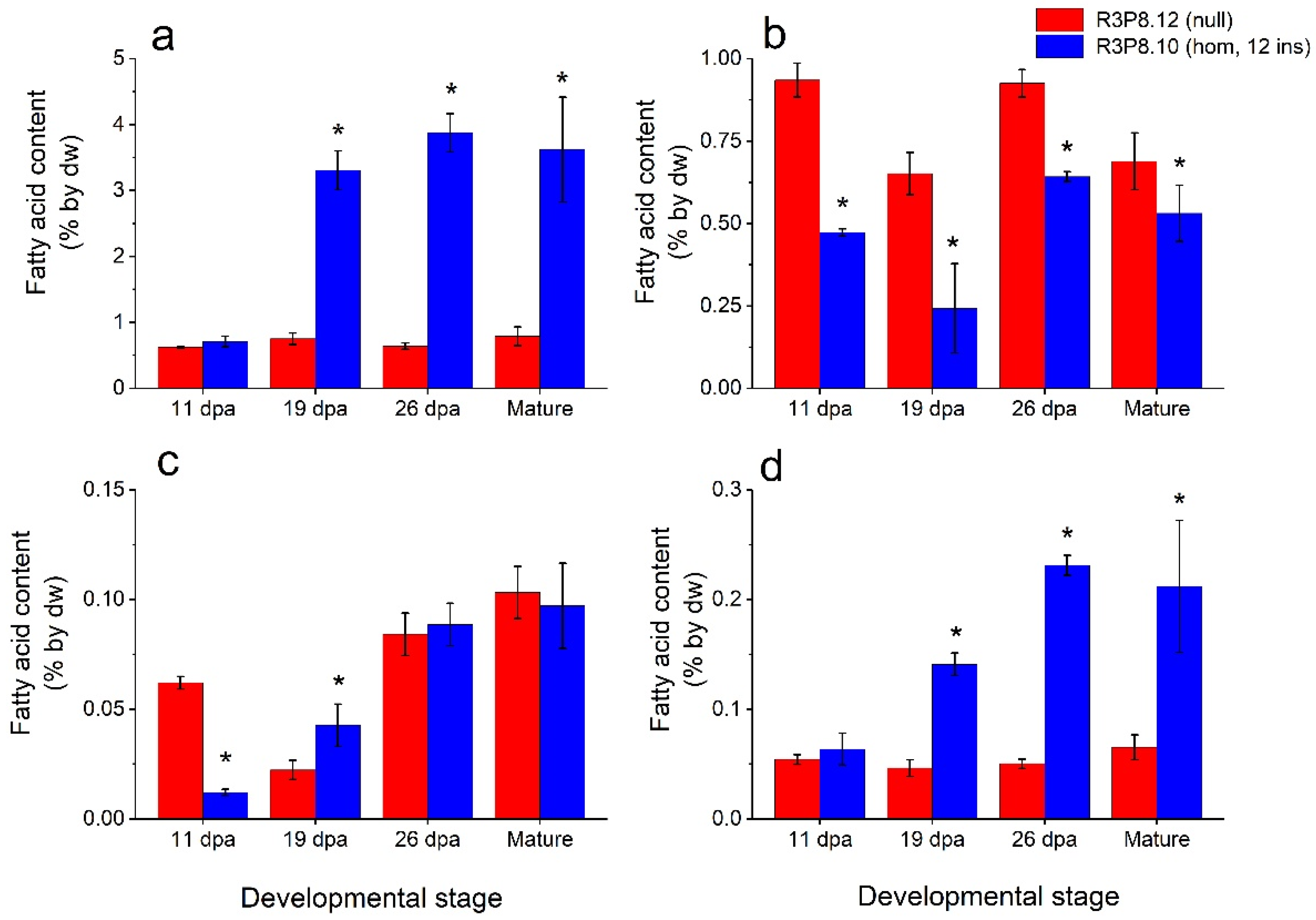
2.3. Lipid and Starch Contents of Endosperms from Developing Wheat Caryopses from Field Grown Plants
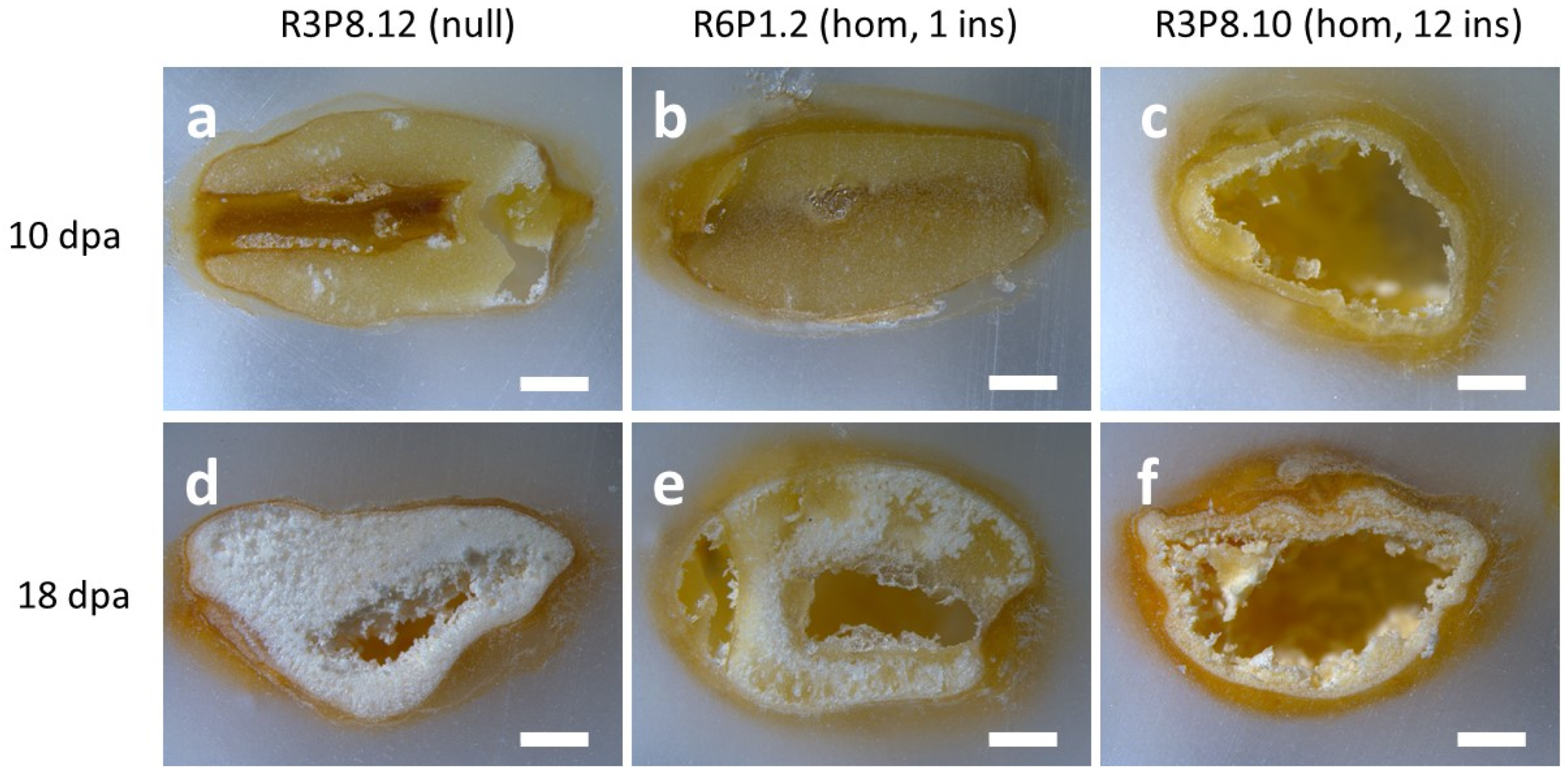
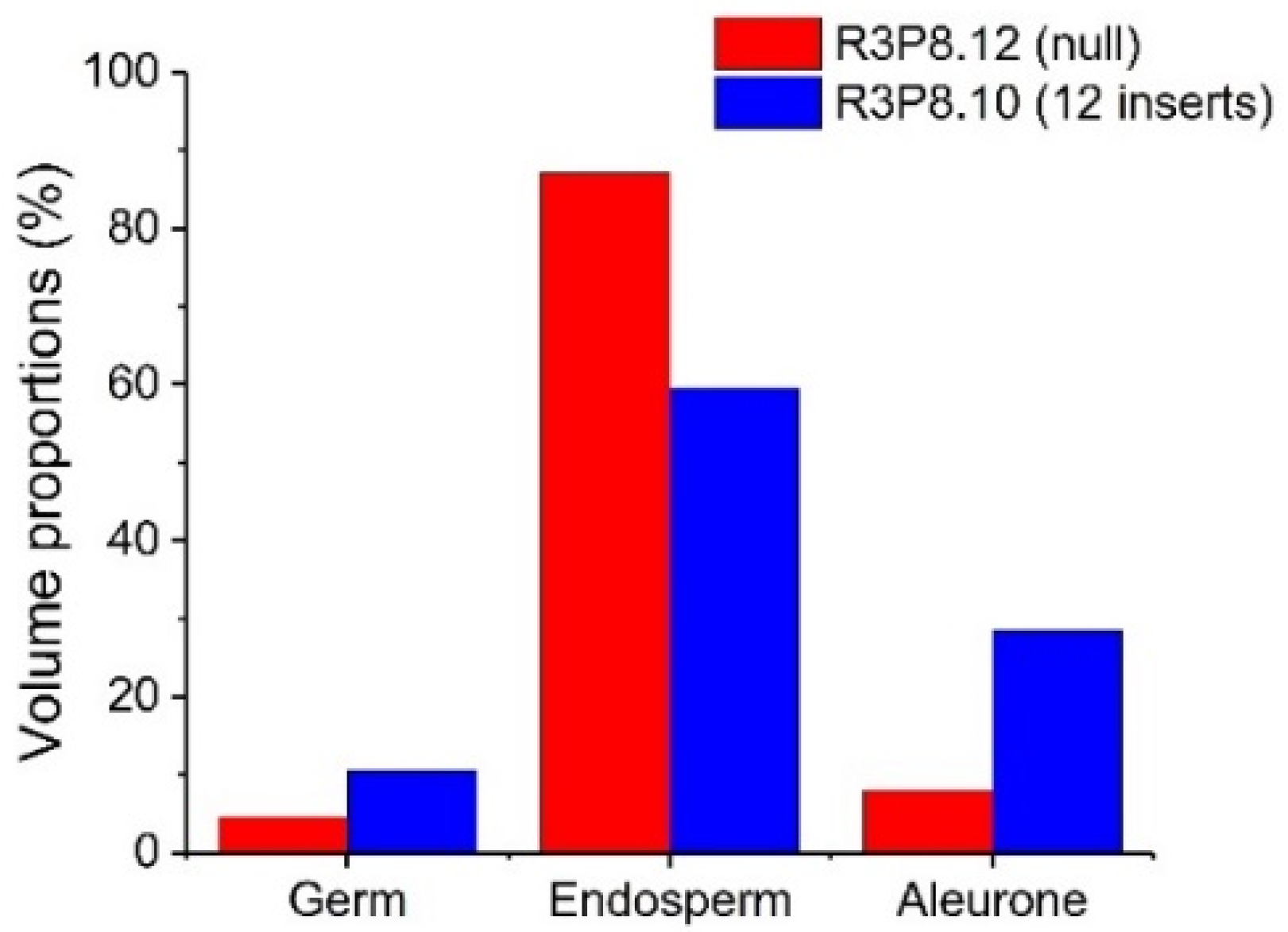
2.4. Morphological Characterisation of Wheat Grains Using X-ray Microtomography
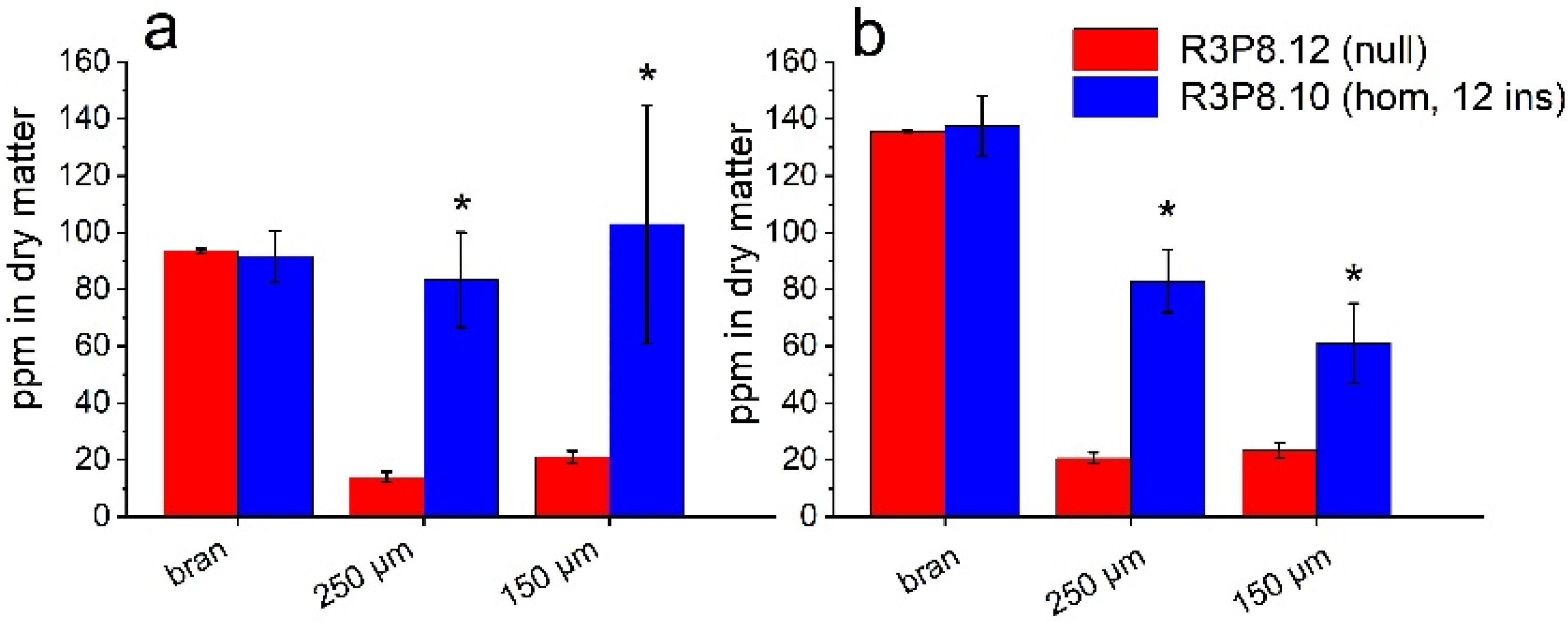
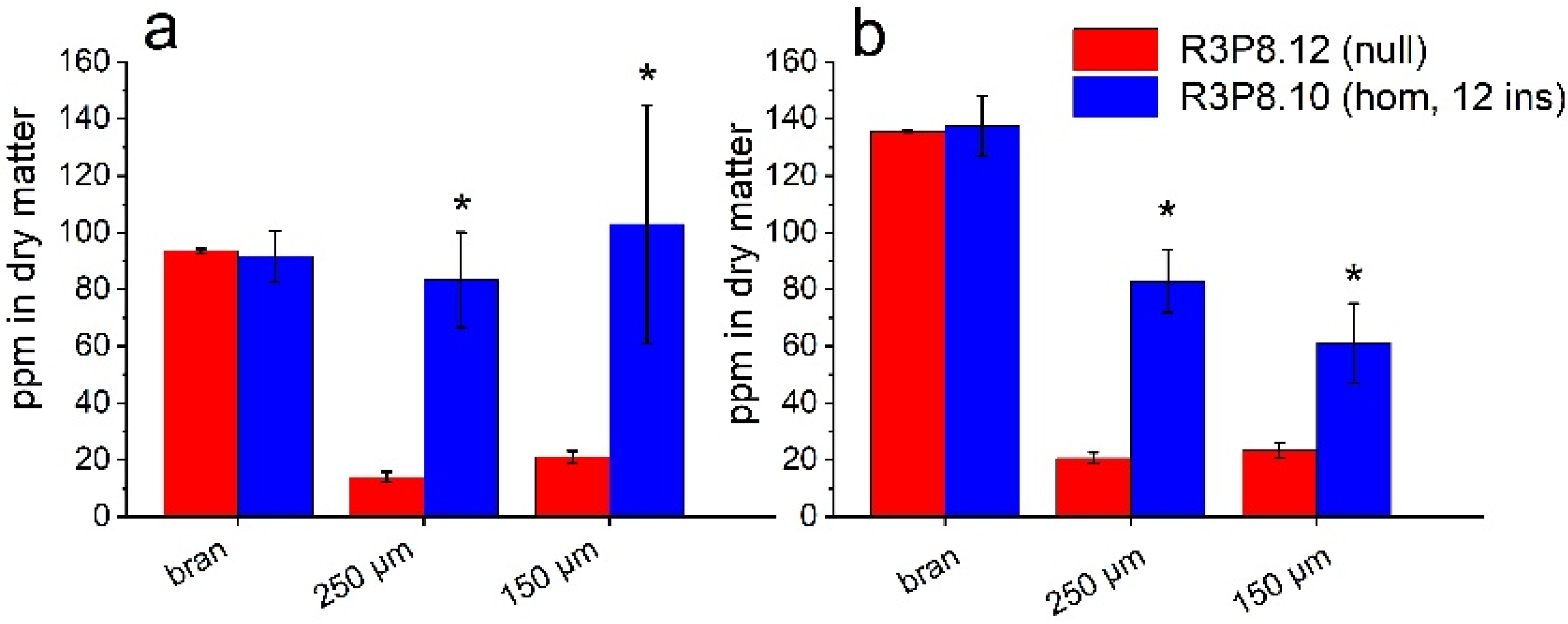
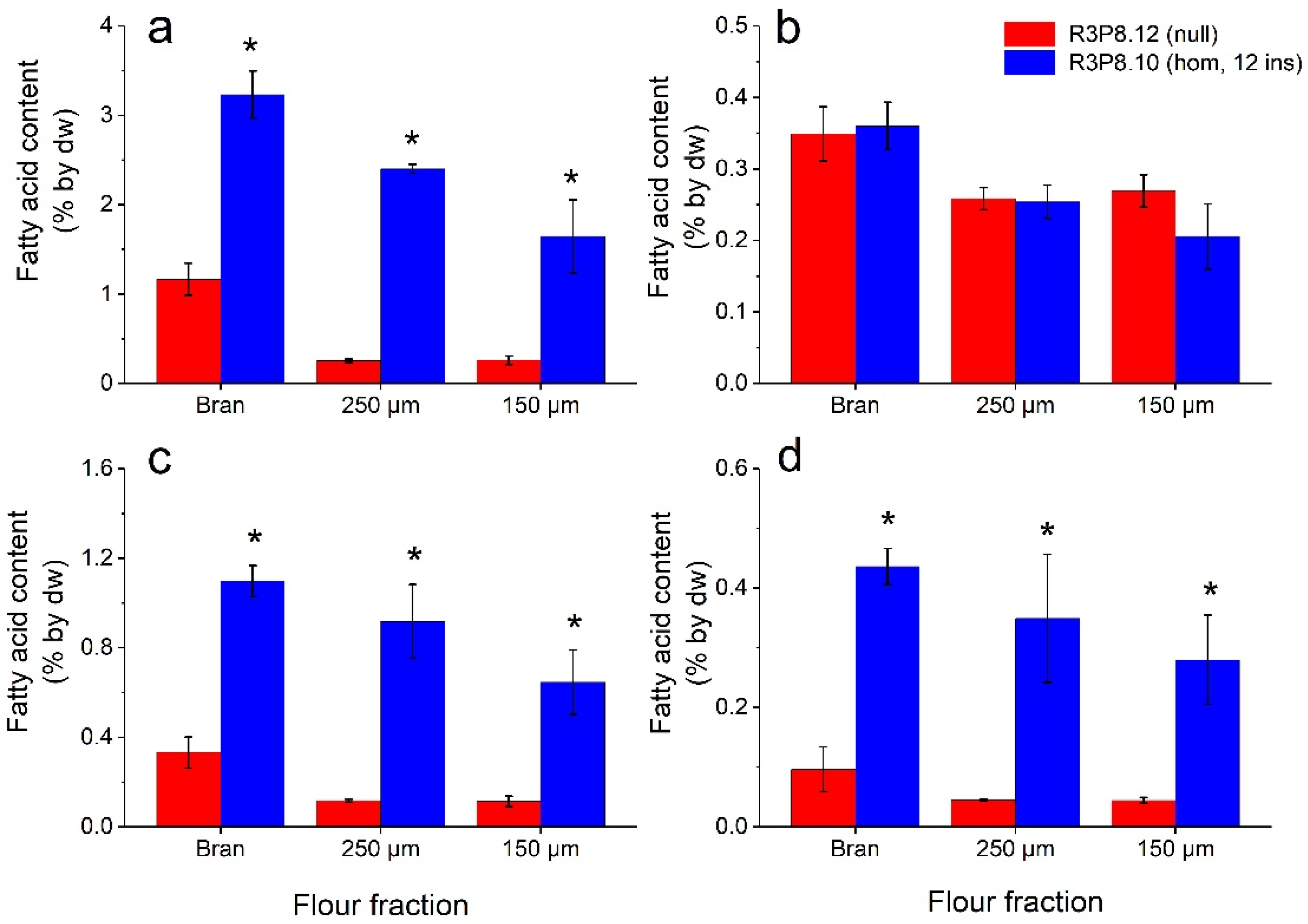
2.5. Lipid and Mineral Contents of Flour Fractions from Wheat Grains from Field Grown Plants
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Grain Characteristics Analyses and Flour Fractionation
4.3. Starch, Lipid and Mineral Analyses
4.4. Fixation of Caryopses and Light Microscopy
4.5. X-ray Microtomography
4.6. Analysis of Volumetric Images
4.7. Statistical Analyses of Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Tosi, P.; He, J.; Lovegrove, A.; Gonzáles-Thuillier, I.; Penson, S.; Shewry, P.R. Gradients in compositions in the starchy endosperm of wheat have implications for milling and processing. Trends Food Sci. Technol. 2018, 82, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chung, O.K.; Ohm, J.B.; Ram, M.; Park, S.; Howitt, C.A. Wheat Lipids. In Wheat: Chemistry and Technology: Fourth Edition; Khan, K., Shrewry, P.R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 363–399. ISBN 9780128104545. [Google Scholar]
- Banaś, K.; Harasym, J. Current Knowledge of Content and Composition of Oat Oil—Future Perspectives of Oat as Oil Source. Food Bioprocess Technol. 2021, 14, 232–247. [Google Scholar] [CrossRef]
- Barron, C.; Surget, A.; Rouau, X. Relative amounts of tissues in mature wheat (Triticum aestivum L.) grain and their carbohydrate and phenolic acid composition. J. Cereal Sci. 2007, 45, 88–96. [Google Scholar] [CrossRef]
- Hargin, K.D.; Morrison, W.R. The distribution of acyl lipids in the germ, aleurone, starch and non-starch endosperm of four wheat varieties. J. Sci. Food Agric. 1980, 31, 877–888. [Google Scholar] [CrossRef]
- Alexander, D.E.; Seif, R.D. Relation of Kernel Oil Content to Some Agronomic Traits in Maize. Crop Sci. 1963, 3, 354–355. [Google Scholar] [CrossRef] [Green Version]
- Khin, O.M.; Sato, M.; Li-Tao, T.; Matsue, Y.; Yoshimura, A.; Mochizuki, T. Close Association between Aleurone Traits and Lipid Contents of Rice Grains Observed in Widely Different Genetic Resources of Oryza sativa. Plant Prod. Sci. 2013, 16, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Heneen, W.K.; Banaś, A.; Leonova, S.; Carlsson, A.S.; Marttila, S.; Dębski, H.; Stymne, S. The distribution of oil in the oat grain. Plant Signal. Behav. 2009, 4, 55–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banaś, A.; Dębski, H.; Banaś, W.; Heneen, W.K.; Dahlqvist, A.; Bafor, M.; Gummeson, P.-O.; Marttila, S.; Ekman, Å.; Carlsson, A.S.; et al. Lipids in grain tissues of oat (Avena sativa): Differences in content, time of deposition, and fatty acid composition. J. Exp. Bot. 2007, 58, 2463–2470. [Google Scholar] [CrossRef]
- Liu, K. Comparison of Lipid Content and Fatty Acid Composition and Their Distribution within Seeds of 5 Small Grain Species. J. Food Sci. 2011, 76, C334–C342. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.M.; Wood, D.F. Composition and Structure of High-Oil Oat. J. Cereal Sci. 1997, 26, 121–128. [Google Scholar] [CrossRef]
- Focks, N.; Benning, C. wrinkled 1: A Novel, Low-Seed-Oil Mutant of Arabidopsis with a Deficiency in the Seed-Specific Regulation of Carbohydrate Metabolism. Plant Physiol. 1998, 118, 91–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruuska, S.A.; Girke, T.; Benning, C.; Ohlrogge, J.B. Contrapuntal Networks of Gene Expression during Arabidopsis Seed Filling. Plant Cell 2002, 14, 1191–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Hua, W.; Zhan, G.; Wei, F.; Wang, X.; Liu, G.; Wang, H. Increasing seed mass and oil content in transgenic Arabidopsis by the overexpression of wri1-like gene from Brassica napus. Plant Physiol. Biochem. 2010, 48, 9–15. [Google Scholar] [CrossRef]
- Troncoso-Ponce, M.A.; Kilaru, A.; Cao, X.; Durrett, T.P.; Fan, J.; Jensen, J.K.; Thrower, N.A.; Pauly, M.; Wilkerson, C.; Ohlrogge, J.B. Comparative deep transcriptional profiling of four developing oilseeds. Plant J. 2011, 68, 1014–1027. [Google Scholar] [CrossRef] [Green Version]
- Bourgis, F.; Kilaru, A.; Cao, X.; Ngando-Ebongue, G.F.; Drira, N.; Ohlrogge, J.B.; Arondel, V. Comparative transcriptome and metabolite analysis of oil palm and date palm mesocarp that differ dramatically in carbon partitioning. Proc. Natl. Acad. Sci. USA 2011, 108, 12527–12532. [Google Scholar] [CrossRef] [Green Version]
- Pouvreau, B.; Baud, S.; Vernoud, V.; Morin, V.; Py, C.; Gendrot, G.; Pichon, J.P.; Rouster, J.; Paul, W.; Rogowsky, P.M. Duplicate Maize Wrinkled1 Transcription Factors Activate Target Genes Involved in Seed Oil Biosynthesis. Plant Physiol. 2011, 156, 674–686. [Google Scholar] [CrossRef] [Green Version]
- Dussert, S.; Guerin, C.; Andersson, M.; Joët, T.; Tranbarger, T.J.; Pizot, M.; Sarah, G.; Omore, A.; Durand-Gasselin, T.; Morcillo, F. Comparative Transcriptome Analysis of Three Oil Palm Fruit and Seed Tissues That Differ in Oil Content and Fatty Acid Composition. Plant Physiol. 2013, 162, 1337–1358. [Google Scholar] [CrossRef] [Green Version]
- An, D.; Kim, H.; Ju, S.; Go, Y.S.; Kim, H.U.; Suh, M.C. Expression of Camelina WRINKLED1 Isoforms Rescue the Seed Phenotype of the Arabidopsis wri1 Mutant and Increase the Triacylglycerol Content in Tobacco Leaves. Front. Plant Sci. 2017, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, D.M.; Rolletschek, H.; Borisjuk, L.; Corwin, J.; Kliebenstein, D.J.; Grimberg, A.; Stymne, S.; Dehesh, K. Cofactome analyses reveal enhanced flux of carbon into oil for potential biofuel production. Plant J. 2011, 67, 1018–1028. [Google Scholar] [CrossRef]
- Grimberg, Å.; Carlsson, A.S.; Marttila, S.; Bhalerao, R.; Hofvander, P. Transcriptional transitions in Nicotiana benthamiana leaves upon induction of oil synthesis by WRINKLED1 homologs from diverse species and tissues. BMC Plant Biol. 2015, 15, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Liu, X.; Li, N.; Du, C.; Wang, K.; Zhao, C.; Wang, Z.; Hu, Y.; Zhang, M. WRINKLED1 homologs highly and functionally express in oil-rich endosperms of oat and castor. Plant Sci. 2019, 287, 110193. [Google Scholar] [CrossRef] [PubMed]
- Grimberg, Å.; Wilkinson, M.; Snell, P.; De Vos, R.P.; González-Thuillier, I.; Tawfike, A.; Ward, J.L.; Carlsson, A.S.; Shewry, P.; Hofvander, P. Transitions in wheat endosperm metabolism upon transcriptional induction of oil accumulation by oat endosperm WRINKLED1. BMC Plant Biol. 2020, 20, 235. [Google Scholar] [CrossRef] [PubMed]
- Becraft, P.W.; Yi, G. Regulation of aleurone development in cereal grains. J. Exp. Bot. 2011, 62, 1669–1675. [Google Scholar] [CrossRef] [Green Version]
- Baird, E.; Taylor, G. X-ray micro computed-tomography. Curr. Biol. 2017, 27, R289–R291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, M.K.; Smith, A.M.; Ellis, T.H.N.; Hedley, C.; Martin, C. The wrinkled-seed character of pea described by Mendel is caused by a transposon-like insertion in a gene encoding starch-branching enzyme. Cell 1990, 60, 115–122. [Google Scholar] [CrossRef]
- Baud, S.; Lepiniec, L. Physiological and developmental regulation of seed oil production. Prog. Lipid Res. 2010, 49, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, K.; Denyer, K. Starch synthesis in cereal grains. Adv. Bot. Res. 2003, 40, 1–61. [Google Scholar] [CrossRef]
- Shewry, P.R.; Wan, Y.; Hawkesford, M.J.; Tosi, P. Spatial distribution of functional components in the starchy endosperm of wheat grains. J. Cereal Sci. 2020, 91, 102869. [Google Scholar] [CrossRef] [PubMed]
- Bechtel, D.B.; Wilson, J.D. Amyloplast Formation and Starch Granule Development in Hard Red Winter Wheat. Cereal Chem. 2003, 80, 175–183. [Google Scholar] [CrossRef]
- Meziani, S.; Nadaud, I.; Tasleem-Tahir, A.; Nurit, E.; Benguella, R.; Branlard, G. Wheat aleurone layer: A site enriched with nutrients and bioactive molecules with potential nutritional opportunities for breeding. J. Cereal Sci. 2021, 100, 103225. [Google Scholar] [CrossRef]
- Brouns, F.; Hemery, Y.; Price, R.; Anson, N.M. Wheat Aleurone: Separation, Composition, Health Aspects, and Potential Food Use. Crit. Rev. Food Sci. Nutr. 2012, 52, 553–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evers, T.; Millar, S. Cereal Grain Structure and Development: Some Implications for Quality. J. Cereal Sci. 2002, 36, 261–284. [Google Scholar] [CrossRef]
- Gustin, J.L.; Jackson, S.; Williams, C.; Patel, A.; Armstrong, P.; Peter, G.F.; Settles, A.M. Analysis of Maize (Zea mays) Kernel Density and Volume Using Microcomputed Tomography and Single-Kernel Near-Infrared Spectroscopy. J. Agric. Food Chem. 2013, 61, 10872–10880. [Google Scholar] [CrossRef] [PubMed]
- Gargiulo, L.; Grimberg, Å.; Repo-Carrasco-Valencia, R.; Carlsson, A.S.; Mele, G. Morpho-densitometric traits for quinoa (Chenopodium quinoa Willd.) seed phenotyping by two X-ray micro-CT scanning approaches. J. Cereal Sci. 2019, 90, 102829. [Google Scholar] [CrossRef]
- Schoeman, L.; Williams, P.; du Plessis, A.; Manley, M. X-ray micro-computed tomography (μCT) for non-destructive characterisation of food microstructure. Trends Food Sci. Technol. 2016, 47, 10–24. [Google Scholar] [CrossRef]
- Schoeman, L.; du Plessis, A.; Manley, M. Non-destructive characterisation and quantification of the effect of conventional oven and forced convection continuous tumble (FCCT) roasting on the three-dimensional microstructure of whole wheat kernels using X-ray micro-computed tomography (μCT). J. Food Eng. 2016, 187, 1–13. [Google Scholar] [CrossRef]
- Larkin, P.J.; Liu, Q.; Vanhercke, T.; Zhou, X.R.; Bose, U.; Broadbent, J.A.; Colgrave, M.L.; Ral, J.P.; Reynolds, K.B.; Sun, M.; et al. Transgenic wheat with increased endosperm lipid—Impacts on grain composition and baking quality. J. Cereal Sci. 2021, 101, 103289. [Google Scholar] [CrossRef]
- Larkin, P.J.; Zhou, X.R.; Liu, Q.; Reynolds, K.; Vanhercke, T.; Ral, J.P.; Li, Z.; Wu, X.B.; Yu, R.; Luo, J.X.; et al. A transcriptional journey from sucrose to endosperm oil bodies in triple transgene oily wheat grain. J. Cereal Sci. 2021, 100, 103268. [Google Scholar] [CrossRef]
- Bosma, R.; Miazek, K.; Willemsen, S.M.; Vermuë, M.H.; Wijffels, R.H. Growth inhibition of Monodus subterraneus by free fatty acids. Biotechnol. Bioeng. 2008, 101, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-T.; Chiang, Y.-R.; Huang, W.-Y.; Jane, W.-N. Cytotoxic effects of free fatty acids on phytoplankton algae and cyanobacteria. Aquat. Toxicol. 2006, 80, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Yan, C.; Xu, C. Phospholipid:diacylglycerol acyltransferase-mediated triacylglycerol biosynthesis is crucial for protection against fatty acid-induced cell death in growing tissues of Arabidopsis. Plant J. 2013, 76, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Munz, J.; Cass, C.; Zienkiewicz, A.; Kong, Q.; Ma, W.; Sanjaya; Sedbrook, J.; Benning, C. Ectopic expression of WRINKLED1 affects fatty acid homeostasis in Brachypodium distachyon vegetative tissues. Plant Physiol. 2015, 169, 1836–1847. [Google Scholar] [CrossRef] [Green Version]
- Vanhercke, T.; Divi, U.K.; El Tahchy, A.; Liu, Q.; Mitchell, M.; Taylor, M.C.; Eastmond, P.J.; Bryant, F.; Mechanicos, A.; Blundell, C.; et al. Step changes in leaf oil accumulation via iterative metabolic engineering. Metab. Eng. 2017, 39, 237–246. [Google Scholar] [CrossRef] [PubMed]
- González-Thuillier, I.; Pellny, T.K.; Tosi, P.; Mitchell, R.A.C.; Haslam, R.; Shewry, P.R. Accumulation and deposition of triacylglycerols in the starchy endosperm of wheat grain. J. Cereal Sci. 2021, 98, 103167. [Google Scholar] [CrossRef] [PubMed]
- Lehtinen, P.; Èinen, K.K.; Èki, I.L.; Laakso, S. Effect of Heat Treatment on Lipid Stability in Processed Oats. J. Cereal Sci. 2003, 37, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhao, L.; Chen, H.; Sun, D.; Deng, B.; Li, J.; Liu, Y.; Wang, F. Inactivation of Lipase and Lipoxygenase of Wheat Germ with Temperature-Controlled Short Wave Infrared Radiation and Its Effect on Storage Stability and Quality of Wheat Germ Oil. PLoS ONE 2016, 11, e0167330. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Dong, Z.; Tian, D.; Dong, L.; Qian, W.; Liu, J.; Liu, X.; Qin, H.; Zhai, W.; Gao, C.; et al. Development and characterization of marker-free and transgene insertion site-defined transgenic wheat with improved grain storability and fatty acid content. Plant Biotechnol. J. 2020, 18, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- White, T.; Bursten, S.; Federighi, D.; Lewis, R.A.; Nudelman, E. High-Resolution Separation and Quantification of Neutral Lipid and Phospholipid Species in Mammalian Cells and Sera by Multi-One-Dimensional Thin-Layer Chromatography. Anal. Biochem. 1998, 258, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Limaye, A. Drishti: A volume exploration and presentation tool. In Proceedings of the Developments in X-ray Tomography VIII; SPIE: Bellingham, WA, USA, 2012; Volume 8506, pp. 191–199. [Google Scholar]
- Lösel, P.D.; Van De Kamp, T.; Jayme, A.; Ershov, A.; Faragó, T.; Pichler, O.; Tan Jerome, N.; Aadepu, N.; Bremer, S.; Chilingaryan, S.A.; et al. Introducing Biomedisa as an open-source online platform for biomedical image segmentation. Nat. Commun. 2020, 11, 5577. [Google Scholar] [CrossRef] [PubMed]
- Doebley, J.F.; Gaut, B.S.; Smith, B.D. The Molecular Genetics of Crop Domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snell, P.; Wilkinson, M.; Taylor, G.J.; Hall, S.; Sharma, S.; Sirijovski, N.; Hansson, M.; Shewry, P.R.; Hofvander, P.; Grimberg, Å. Characterisation of Grains and Flour Fractions from Field Grown Transgenic Oil-Accumulating Wheat Expressing Oat WRI1. Plants 2022, 11, 889. https://doi.org/10.3390/plants11070889
Snell P, Wilkinson M, Taylor GJ, Hall S, Sharma S, Sirijovski N, Hansson M, Shewry PR, Hofvander P, Grimberg Å. Characterisation of Grains and Flour Fractions from Field Grown Transgenic Oil-Accumulating Wheat Expressing Oat WRI1. Plants. 2022; 11(7):889. https://doi.org/10.3390/plants11070889
Chicago/Turabian StyleSnell, Per, Mark Wilkinson, Gavin J. Taylor, Stephen Hall, Shrikant Sharma, Nick Sirijovski, Mats Hansson, Peter R. Shewry, Per Hofvander, and Åsa Grimberg. 2022. "Characterisation of Grains and Flour Fractions from Field Grown Transgenic Oil-Accumulating Wheat Expressing Oat WRI1" Plants 11, no. 7: 889. https://doi.org/10.3390/plants11070889
APA StyleSnell, P., Wilkinson, M., Taylor, G. J., Hall, S., Sharma, S., Sirijovski, N., Hansson, M., Shewry, P. R., Hofvander, P., & Grimberg, Å. (2022). Characterisation of Grains and Flour Fractions from Field Grown Transgenic Oil-Accumulating Wheat Expressing Oat WRI1. Plants, 11(7), 889. https://doi.org/10.3390/plants11070889