
Crocus pallidus (Iridaceae)—A Neglected Species for the Bulgarian Flora and Critical Taxon in the Balkans


 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Description Based on Bulgarian Materials
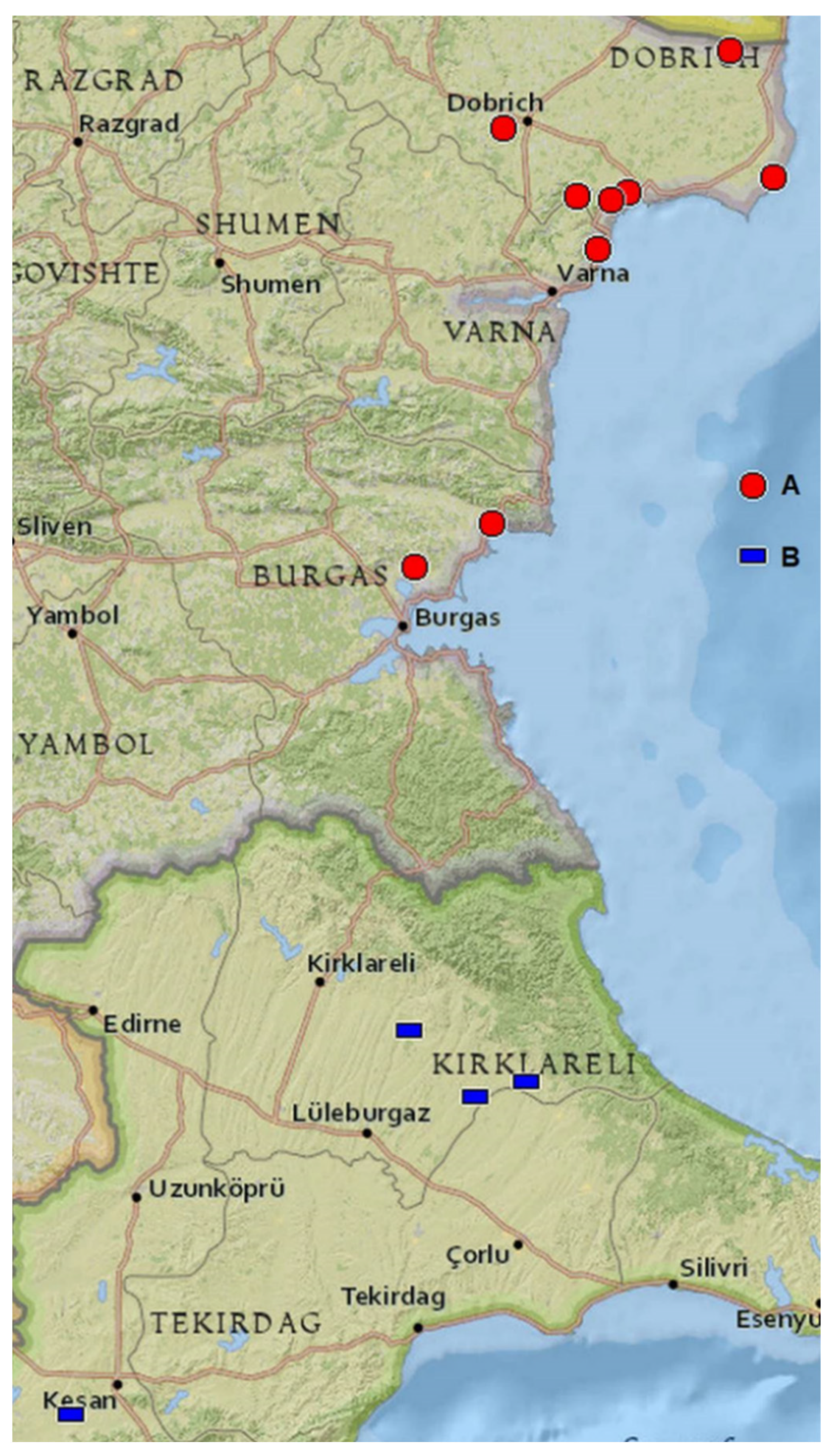
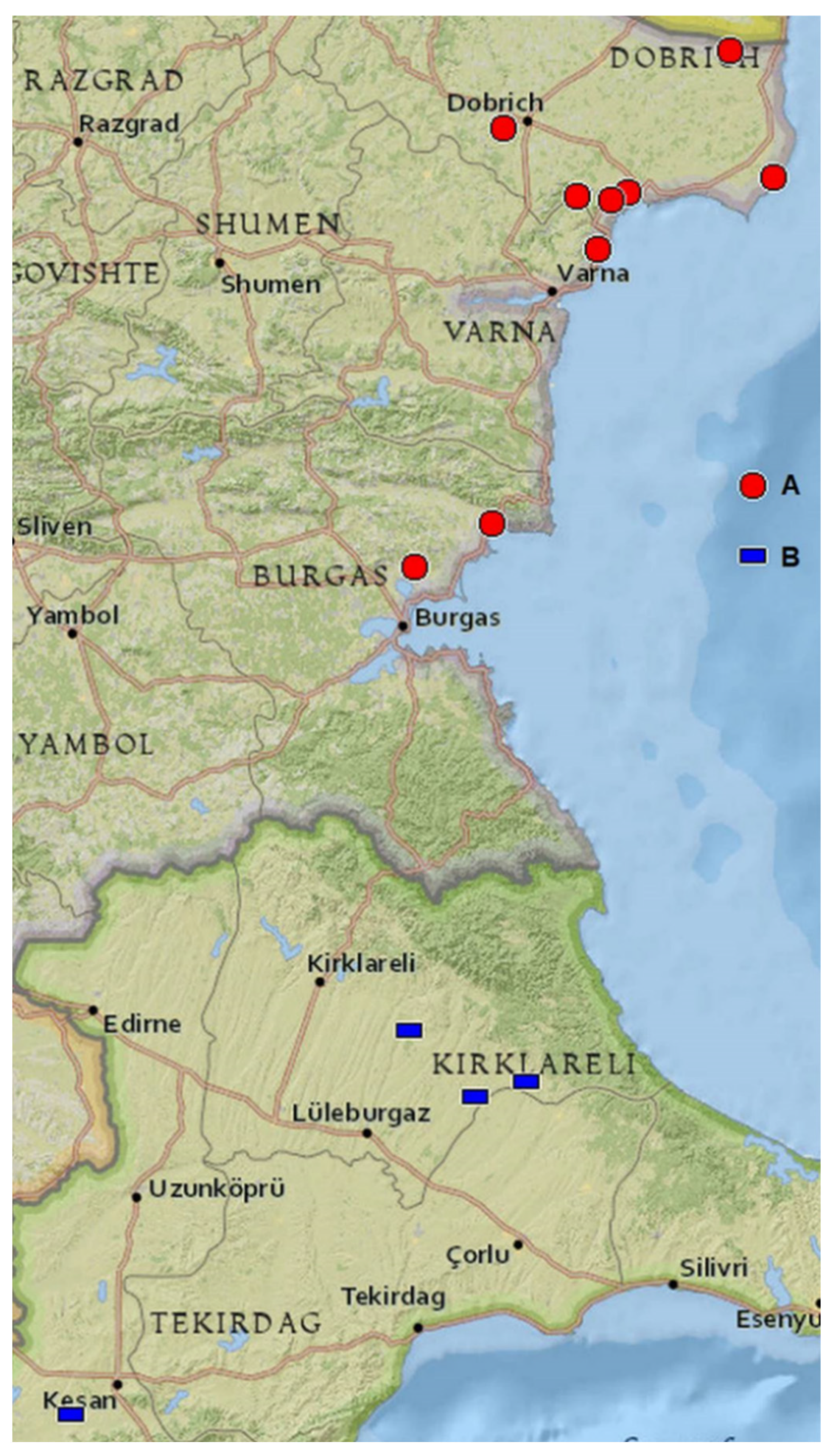
2.2. Distribution
2.3. Phenology and Habitats
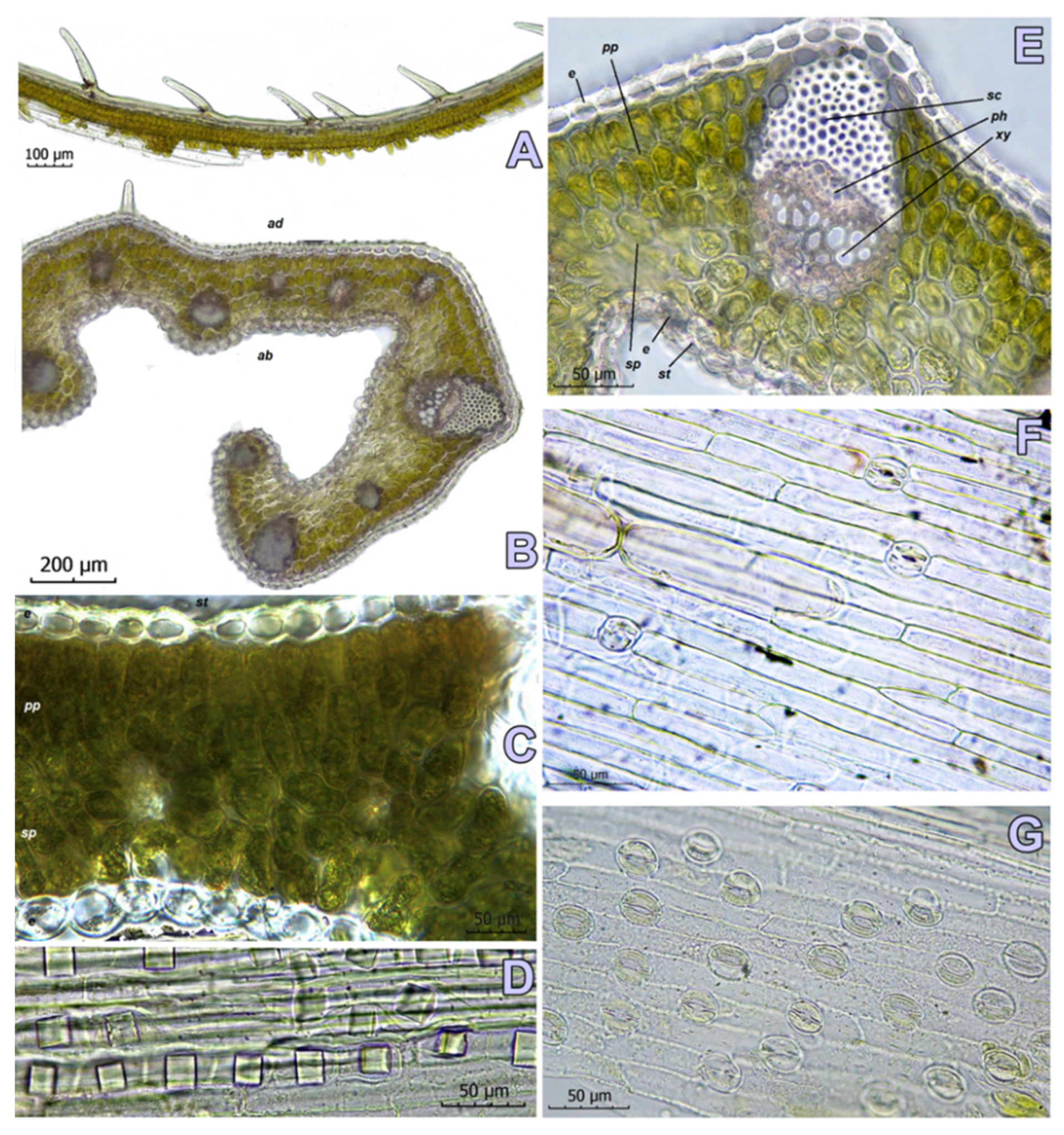
2.4. Leaf Anatomy
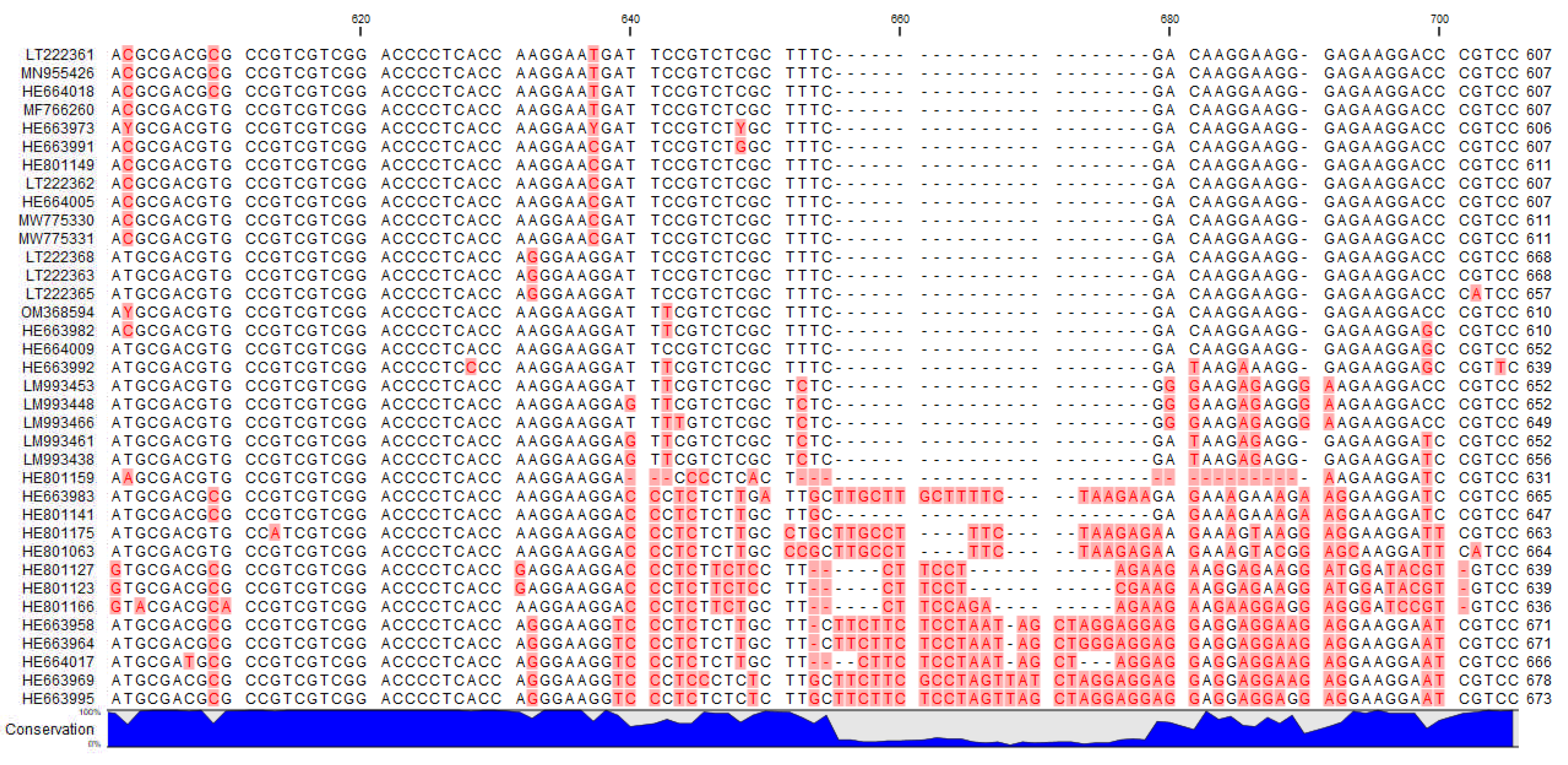
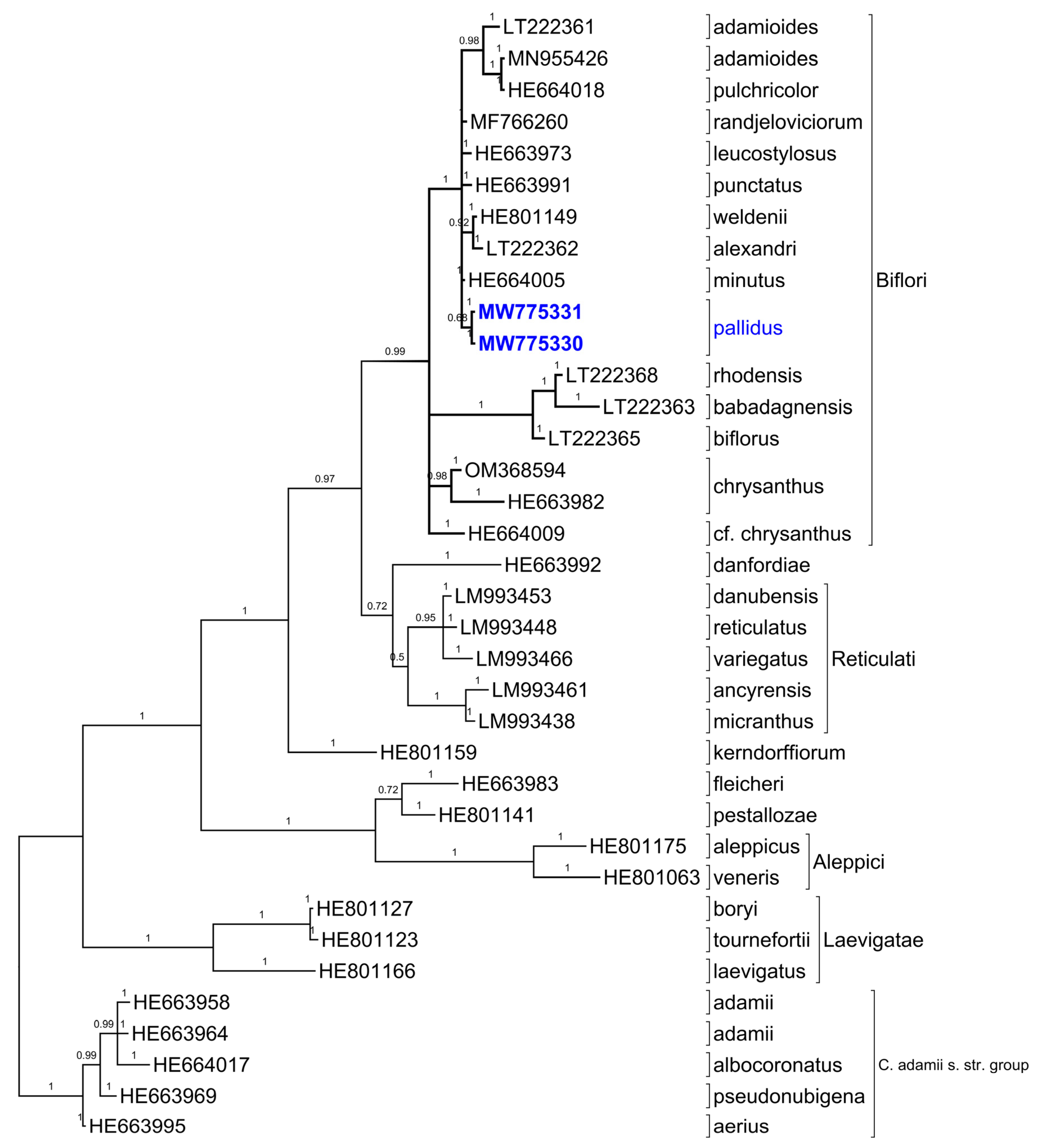
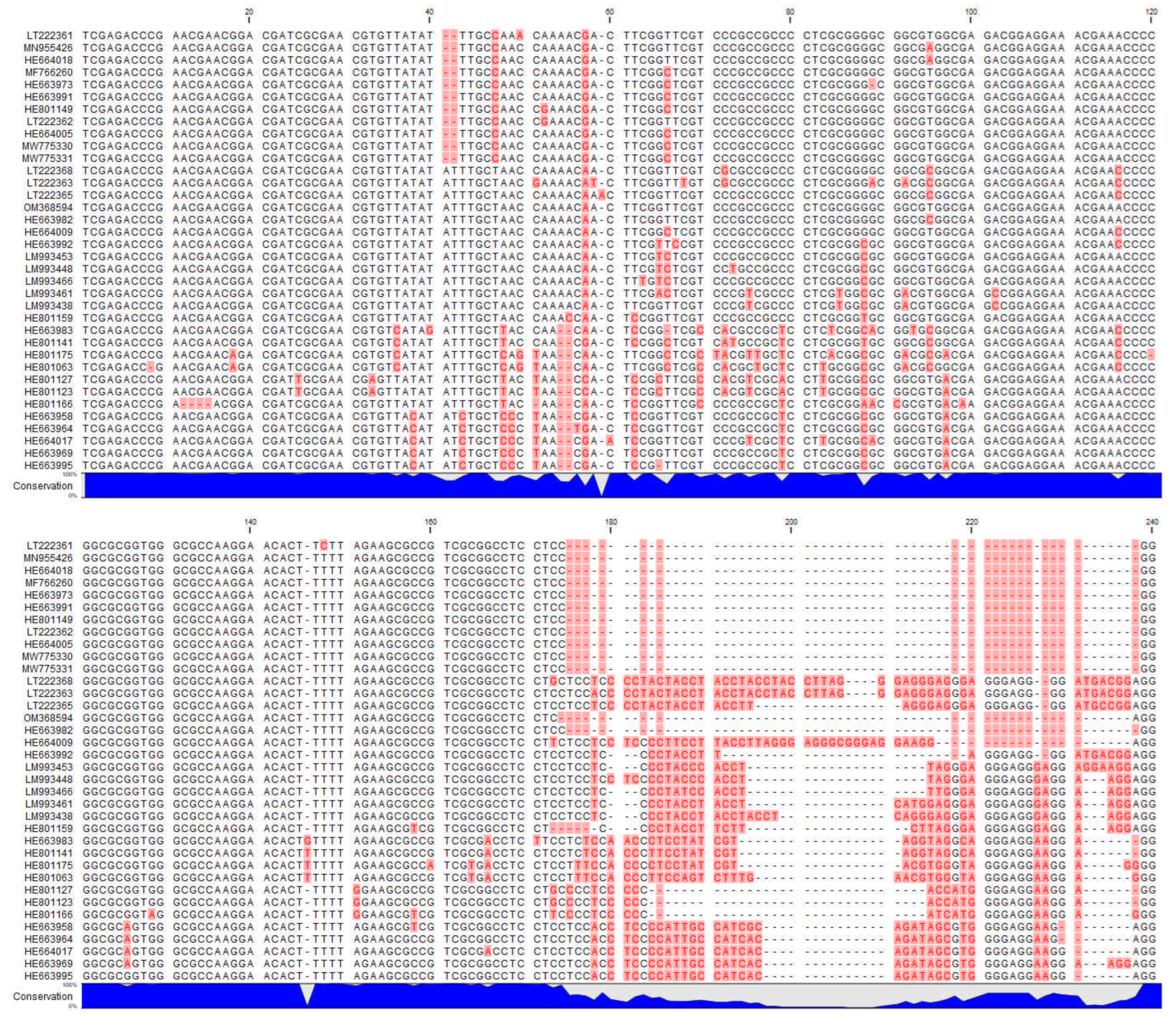
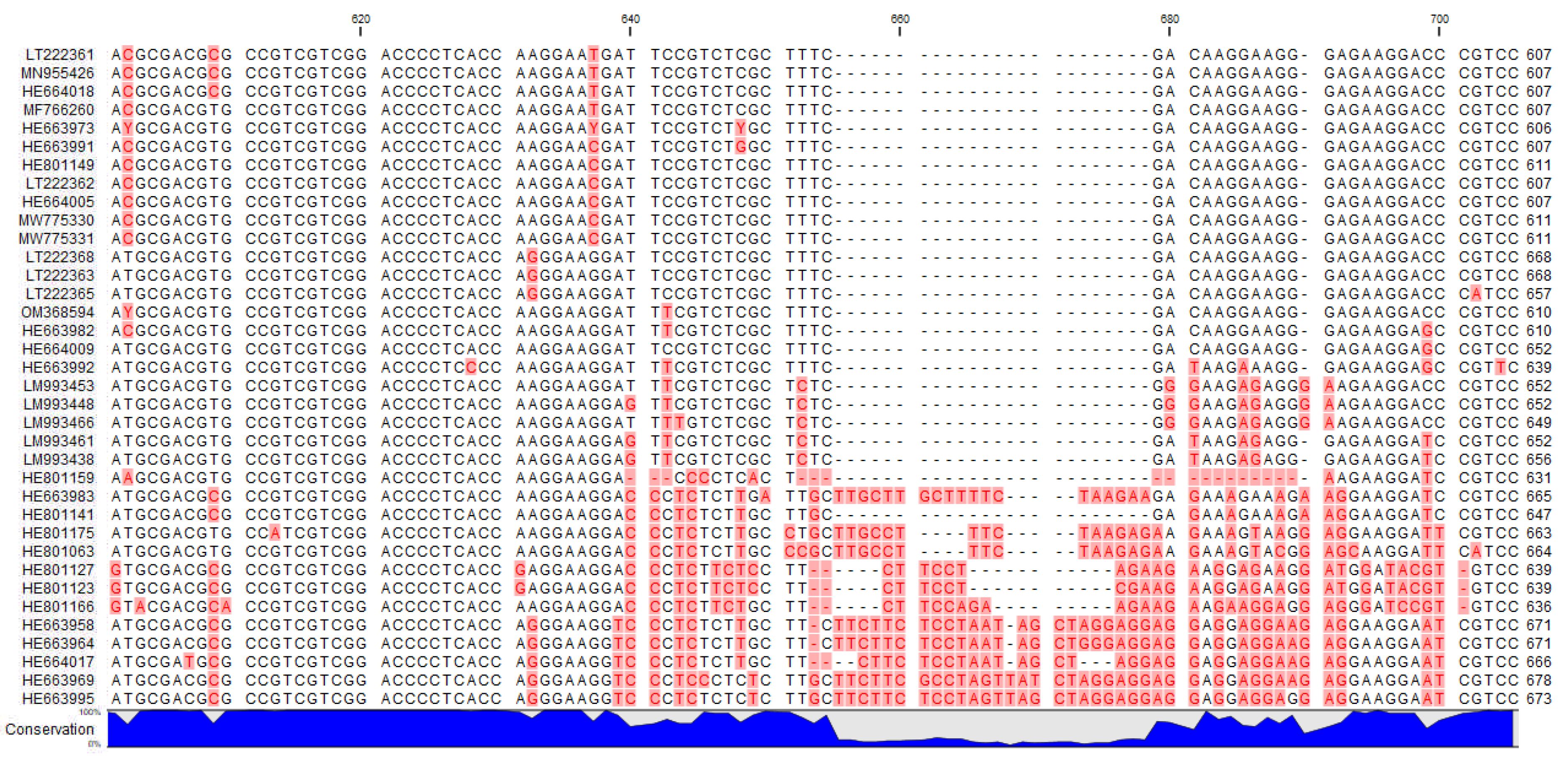
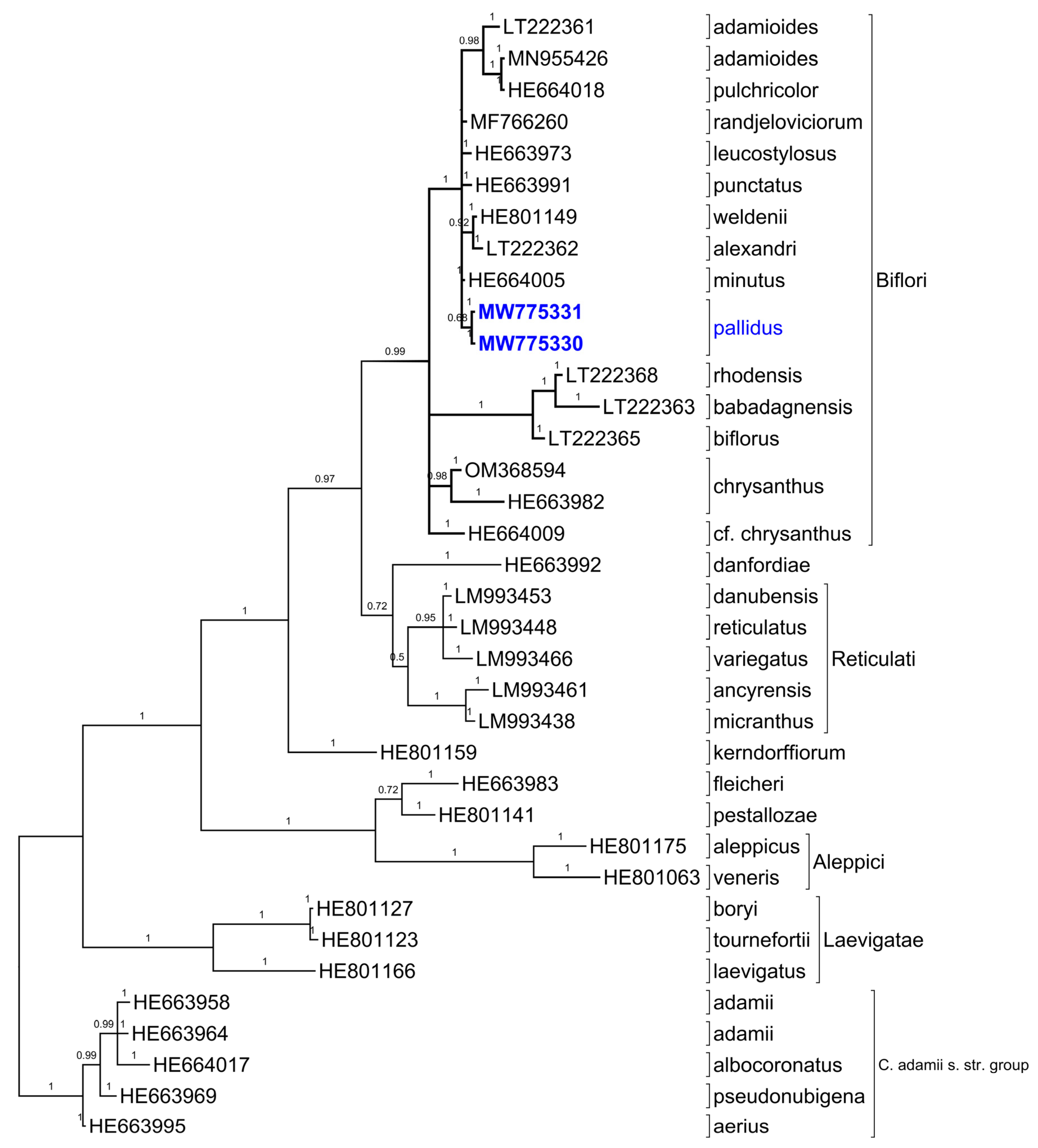
2.5. ITS Sequences
3. Discussion
4. Materials and Methods
4.1. Examined Specimens
- Crocus pallidus:
- Crocus thracicus (photo scans):
4.2. Anatomical Investigations
4.3. Molecular Methods
4.4. Phylogenetic Analyses
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | NCBI Genbank Accession Number | Authors | Country | Voucher Specimen |
|---|---|---|---|---|
| C. adamii | HE663958 | DH, SM, TR, HK, FB | Armenia | GAT 7161 -! |
| C. adamii | HE663964 | DH, SM, TR, HK, FB | Armenia | B 10 0355320 |
| C. adamioides | LT222361 | DH, HK, EP, LP | Turkey | GAT 7136 -! |
| C. adamioides | MN955426 | SN, EA, KS, TR | Bulgaria | SOA 062625 -! |
| C. aerius | HE663995 | DH, SM, TR, HK, FB | Turkey | GAT 7178 -! |
| C. albocoronatus | HE664017 | DH, SM, TR, HK, FB | GAT 7477 -! | |
| C. aleppicus | HE801175 | DH, SM, TR, HK, FB | Jordan | IABH 18357 |
| C. alexandri | LT222362 | DH, HK, EP, LP | Serbia | GAT 23024 -! |
| C. ancyrensis | LM993461 | DH, LP, HK, TK, TC, VR, NR, MJ, FB | Turkey | GAT 29387 -! |
| C. babadagnensis | LT222363 | DH, HK, EP, LP | GAT 7185 | |
| C. biflorus | LT222365 | DH, HK, EP, LP | Italy | GAT25812 -! |
| C. boryi | HE801127 | DH, SM, TR, HK, FB | cult. | GAT 7209 -! |
| C. chrysanthus | OM368594 | SN, EA, KS, TR | Bulgaria | SOA 062596 -! |
| C. chrysanthus | HE663982 | DH, SM, TR, HK, FB | Turkey | GAT 7138 -! |
| C. cf. chrysanthus | HE664009 | DH, SM, TR, HK, FB | Turkey | GAT 7449 -! |
| C. danfordiae | HE663992 | DH, SM, TR, HK, FB | Turkey | GAT 7123 -! |
| C. danubensis | LM993453 | DH, LP, HK, TK, TC, VR, NR, MJ, FB | Serbia | GAT 23019 -! |
| C. fleischeri | HE663983 | DH, SM, TR, HK, FB | Turkey | GAT 7139 -! |
| C. kerndorffiorum | HE801159 | DH, SM, TR, HK, FB | Turkey | K 00802496 -! |
| C. laevigatus | HE801166 | DH, SM, TR, HK, FB | cult. | GAT 7224 -! |
| C. leucostylosus | HE663973 | DH, SM, TR, HK, FB | Turkey | GAT 7471 -! |
| C. micranthus | LM993438 | DH, LP, HK, TK, TC, VR, NR, MJ, FB | Turkey | GAT 25831 -! |
| C. minutus | HE664005 | DH, SM, TR, HK, FB | Turkey | GAT 7470 -! |
| C. pallidus | MW775330 | SN, EA, KS, TR | Bulgaria | SOA 062797 -! |
| C. pallidus | MW775331 | SN, EA, KS, TR | Bulgaria | SOA 062791 -! |
| C. pestalozzae | HE801141 | DH, SM, TR, HK, FB | Turkey | GAT 7261 -! |
| C. pseudonubigena | HE663969 | DH, SM, TR, HK, FB | Turkey | GAT 7452 -! |
| C. pulchricolor | HE664018 | DH, SM, TR, HK, FB | Turkey | GAT 7188 -! |
| C. punctatus | HE663991 | DH, SM, TR, HK, FB | Turkey | GAT 7155 -! |
| C. randjeloviciorum | MF766260 | DH, HK, IR, EP | Serbia | GAT 23042 -! |
| C. reticulatus | LM993448 | DH, LP, HK, TK, TC, VR, NR, J, FB | Russia | GAT 23082 -! |
| C. rhodensis | LT222368 | DH, HK, EP, LP | Greece | GAT s/n |
| C. tournefortii | HE801123 | DH, SM, TR, HK, FB | Greece | GAT 7202 -! |
| C. variegatus | LM993466 | DH, LP, HK, TK, TC, VR, NR, MJ, FB | GAT 7264 | |
| C. veneris | HE801063 | DH, SM, TR, HK, FB | cult. | GAT 7201 -! |
| C. weldenii | HE801149 | DH, SM, TR, FB | cult. | GAT 7392 -! |
References
- Harpke, D.; Meng, S.; Rutten, T.; Kerndorff, H.; Blattner, F.R. Phylogeny of Crocus (Iridaceae) based on one chloroplast and two nuclear loci: Ancient hybridization and chromosome number evolution. Mol. Phylogenetics Evol. 2013, 66, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Rukšāns, J. The World of Crocuses; Latvian Academy of Sciences: Riga, Latvia, 2017; p. 568. [Google Scholar]
- Yüzbaşıoğlu, S. Crocus keltepensis (Iridaceae): A new Crocus species from the highest part of the Samanlı Mountains, NW Turkey. Phytotaxa 2019, 418, 230–236. [Google Scholar] [CrossRef]
- Raca, I.; Harpke, D.; Shuka, L.; Ranđelović, V. A new species of Crocus ser. Verni (Iridaceae) with 2n = 12 chromosomes from the Balkans. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2020, 1–7. [Google Scholar] [CrossRef]
- Velchev, V. Crocus L. In Flora Reipublicae Popularis Bulgaricae, 2nd ed.; Jordanov, D., Ed.; Academia Scientarum Bulgaricae: Sofia, Bulgaria, 1964; pp. 328–338. [Google Scholar]
- Kitanov, B.; Drenkovski, R. Über die Taxonomie einiger Sippen der Gattung Crocus L. auf der Balkanhalbinsel. In Problems of Balkan Flora and Vegetation, Proceedings of the First International Symposium on Balkan Flora and Vegetation, Varna, Bulgaria, 7–14 June 1973; Bulgarian Academy of Science: Sofia, Bulgaria, 1975; pp. 212–215. [Google Scholar]
- Kitanov, B.; Penev, I.; Krachunov, I. Materials on the flora of Dobroudja. Annu. Sofia Univ. St. Kliment Ohridski Fac. Biol. 1977, 70, 85–87. [Google Scholar]
- Kitanov, B.; Penev, I. Flora of Dobroudja; Naouka & Izkoustvo: Sofia, Bulgaria, 1980; p. 630. [Google Scholar]
- Delipavlov, D. Crocus L. In Key to the Plants in Bulgaria; Delipavlov, D., Cheschmedzhiev, I., Eds.; Agrarian University Academic Pres: Plovdiv, Bulgaria, 2003; pp. 456–457. [Google Scholar]
- Petrova, A.; Vladimirov, V. Balkan endemics in the Bulgarian flora. Phytol. Balc. 2010, 16, 293–311. [Google Scholar]
- Assyov, B.; Petrova, A. Conspectus of the Bulgarian Vascular Flora, 4th ed.; Distribution Maps and Floristic Elements; Bulgarian Biodiversity Foundation: Sofia, Bulgaria, 2012; pp. 156–157. [Google Scholar]
- Pulević, V. Two new species of the genus Crocus L. from Yugoslavia. Glas. Republičkog Zavoda Zaštitu Prir. Prir. Zbirke Titogr. 1976, 9, 39–43. [Google Scholar]
- Mathew, B. The Crocus, a Revision of the Genus Crocus; BT Batsford Ltd.: London, UK, 1982; p. 127. [Google Scholar]
- WCSP. World Checklist of Selected Plant Families. Facilitated by the Royal Botanic Gardens, Kew. 2021. Available online: http://wcsp.science.kew.org/ (accessed on 4 January 2021).
- Pavletić, Z.; Trinajstić, I. Prilog poznavanju horologije taksona Crocus weldenii Hoppe et Fürnrohr u srednjoj Dalmaciji. Acta Bot. Croat. 1979, 38, 163–166. [Google Scholar]
- Pulević, V. Prilog taksonomiji i horologiji nekih vrsta roda Crocus L. iz Jugoslavije (C. weldenii, C. alexandri i C. adamii sens. lat.). Glas. Republičkog Zavoda Zaštitu Prir. Prir. Zbirke Titogr. 1977, 10, 81–99. [Google Scholar]
- Ranđelović, N.; Hill, D.A.; Ranđelović, V. The Genus Crocus L. in Serbia; The Serbian Academy of Sciences and Arts: Belgrade, Serbia, 1990; p. 49. [Google Scholar]
- Rakaj, M. Floristic and chorological news from north Albania. Bot. Serbica 2009, 33, 177–183. [Google Scholar]
- Ranđelović, N.; Sekovski, Ž.; Dimeska, G. Systematic, chorological and genetic research into the genus Crocus L. in Macedonia. In Collection of Papers Devoted to Academician Kiril Micevski; Cyril and Methodius University in Skopje: Skopje, North Macedonia, 2007; pp. 98–131. [Google Scholar]
- Brighton, C.A.; Mathew, B.; Marchant, C. Chromosome counts in the genus Crocus (Iridaceae). Kew Bull. 1973, 28, 451–464. [Google Scholar] [CrossRef]
- Goldblatt, P.; Takei, M. Chromosome cytology of Iridaceae, patterns of variation, determination of ancestral base numbers, and modes of karyotype change. Ann. Mo. Bot. Gard. 1997, 84, 285–304. [Google Scholar] [CrossRef]
- Schneider, I.; Kerndorff, H.; Pasche, E. Chromosome numbers of Turkish Crocus (Liliiflorae, Iridaceae) and their geographical distribution. Feddes Repert. 2012, 123, 73–79. [Google Scholar] [CrossRef]
- Harpke, D.; Peruzzi, L.; Kerndorff, H.; Karamplianis, T.; Constantinidis, T.; Ranđelović, V.; Ranđelović, N.; Jušković, M.; Pasche, E.; Blattner, F.R. Phylogeny, geographic distribution and new taxonomic circumscription of the Crocus reticulatus species group (Iridaceae). Turk. J. Bot. 2014, 38, 1182–1198. [Google Scholar] [CrossRef]
- Harpke, D.; Kerndorff, H.; Pasche, E.; Peruzzi, L. Neotypification of the name Crocus biflorus Mill. (Iridaceae) and its consequences in the taxonomy of the genus. Phytotaxa 2016, 260, 131–143. [Google Scholar] [CrossRef]
- Mosolygó, Á.; Sramkó, G.; Barabás, S.; Czeglédi, L.; Jávor, A.; Molnár, A.; Surányi, G. Molecular genetic evidence for allotetraploid hybrid speciation in the genus Crocus L. (Iridaceae). Phytotaxa 2016, 258, 121–136. [Google Scholar] [CrossRef]
- NCBI GenBank Nucleotide Database. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 10 January 2022).
- Yüzbaşioğlu, S.; Aslan, S.; Özhatay, N. Crocus thracicus (Iridaceae), a new species from north-western Turkey. Phytotaxa 2015, 197, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Harpke, D.; Kerndorff, H.; Raca, I.; Pasche, E. A new Serbian endemic species of the genus Crocus (Iridaceae). Biol. Nyssana 2017, 8, 7–13. [Google Scholar] [CrossRef]
- Kerndorff, H.; Pasche, E.; Harpke, D. The Genus Crocus (Liliflorae, Iridaceae): Life-cycle, morphology, phenotypic characteristics, and taxonomical relevant parameters. Stapfia 2015, 103, 27–65. [Google Scholar]
- Aykurt, C.; Deniz, I.G.; Başaran, M.A.; Genç, G.E. Taxonomic and GIS-based ecologic studies on critically endangered Crocus wattiorum B. Mathew (Olympos saffron). Thaiszia-J. Bot. 2015, 25, 73–84. [Google Scholar]
- Hršak, V.; Lukač, G. The genus Crocus L. in the Paklenica National Park. Acta Bot. Croat. 2001, 60, 97–101. Available online: https://hrcak.srce.hr/160820 (accessed on 10 February 2021).
- Milović, M. Rod Crocus L. (Iridaceae) u flori Hrvatske. Glas. Hrvat. Bot. Društva 2016, 4, 4–20. Available online: https://hrcak.srce.hr/167881 (accessed on 28 January 2021).
- Kerndorff, H.; Pasche, E. Two New Taxa of Crocus (Liliiflorae, Iridaceae) from Turkey. Stapfia 2011, 95, 2–5. [Google Scholar]
- Leibniz Institute of Plant Genetics and Crop Plant Research (IPK). Herbarium GAT. Occurrence Dataset. GBIF.org. Available online: https://www.gbif.org/dataset/5d47cb09-3ef2-477d-a539-dd7f6955aefd (accessed on 20 January 2022).
- Royal Botanic Gardens, Kew. Royal Botanic Gardens, Kew-Herbarium Specimens. Occurrence Dataset. GBIF.org. 2021. Available online: https://www.gbif.org/dataset/cd6e21c8-9e8a-493a-8a76-fbf7862069e5 (accessed on 20 January 2022).
- Steiermärkisches Landesmuseum Joanneum-Herbarium GJO. Steiermärkisches Landesmuseum Joanneum-Herbarium GJO. Occurrence Dataset. GBIF.org. Available online: https://www.gbif.org/dataset/85a214d6-f762-11e1-a439-00145eb45e9a/ (accessed on 13 February 2022).
- Bijmoer, R.; Scherrenberg, M.; Creuwels, J. Naturalis Biodiversity Center (NL)-Botany. Naturalis Biodiversity Center. Occurrence Dataset. GBIF.org. 2022. Available online: https://www.gbif.org/dataset/15f819bd-6612-4447-854b-14d12ee1022d (accessed on 13 February 2022).
- University of Oslo. Vascular Plant Herbarium, Oslo (O) UiO. Version 1.1754. Occurrence Dataset. GBIF.org. 2022. Available online: https://www.gbif.org/dataset/e45c7d91-81c6-4455-86e3-2965a5739b1f/ (accessed on 13 February 2022).
- Van Westen, M. Micam 2.4. Available online: http://science4all.nl (accessed on 10 October 2019).
- Tirel, C.; Jérémie, J.; Lobreau-Callen, D. Corchorus neocaledonicus (Tiliaceae), véritable identité de l’’énigmatique Oceanopapaver. Bull. Du Muséum Natl. D’histoire Nat. 1996, 18, 35–43. [Google Scholar]
- NCBI. Standard Nucleotide BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 10 January 2022).
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. Available online: https://bmcbioinformatics.biomedcentral.com/track/pdf/10.1186/1471-2105-11-7.pdf (accessed on 10 June 2019). [CrossRef] [Green Version]
- CLC Sequence Viewer 8.0, QIAGEN. Available online: https://clc-sequence-viewer.software.informer.com/8.0/ (accessed on 27 October 2021).





| C. pallidus | C. weldenii | C thracicus | |
|---|---|---|---|
| Corm | 13.5–25 mm | 13–19.7 mm | 10–12 mm |
| Plant height | (8)10–20 cm | 8–17 cm | 7.35–12 mm |
| Teeth of the basal rings | Irregular | Regular/Irregular | Irregular |
| Count of leaves | 3–5 | 3–5 | 3–4(–5) |
| Leaf width | 0.84–2.3 mm | max 1 mm | 0.75–1.3 mm |
| Ribs on the abaxial leaf surface | Missing or vaguely visible | 1–2, clearly visible | Missing |
| Color of the perigone tube | White, rarely violet | White, often violet | White, rarely violet near the apex |
| Color of outer perigone segments | White or sprayed in violet | White | White or sprayed in violet |
| Size of perigone segments (out/in) | 19–33 × 6–11/15–26 × 5–12 mm | 19–37 × 4.8–12.8 | 17–24 × 6–9/14.6–23 × 5.7–8.7 mm |
| Perigone throat | Glabrous, white | Yellow | Glabrous, white |
| Filaments | 3.8–13.7 mm | 8–16 mm | 10–13.5 mm |
| Anthers | 8–16 mm, yellow, with blackish basal lobes, or entirely black edge | yellow, without blackish lobes | 7.3–11.8 mm, yellow, with blackish basal lobes. |
| Capsule | Ellipsoid; 15–21 mm long; 4.3–7.4 mm wide. | Ellipsoid, about 14 mm long | |
| Seeds | 2.2–3.4 × 1.5–2.1 mm, with convex caruncle | 2.5 mm in diameter, with convex caruncle | |
| Distribution | Bulgaria (Eastern parts), North Macedonia, Serbia. | Italy, Albania, Serbia. | Turkey in Europe (Thrakia) |
| Elevation | 30–190 m | 100–750 m | 45–170 m |
| Flowering period | February–March | February–March |
| Population * | 062791 Kamen Bryag | 062797 Pomorie Narrow | 063064 Roudnik |
|---|---|---|---|
| Section width, µm | 1701–2224 1927.6 ± 133 | 955–1722 1210.9 ± 193 | 910–2017 1432.6 ± 327.9 |
| Section height, µm | 709–807 761.1 ± 39.8 | 356–664 481.8 ± 89.2 | 480–633 562.8 ± 54.8 |
| Arm width, µm | 657–973 830.1 ± 96.3 | 385–696 547.2 ± 91.1 | 455–1071 703.4 ± 146.6 |
| White stripe to leaf width ratio, % | 15.1–21 17.4 ± 2 | 12.7–28.8 20 ± 4.2 | 14.9–19.3 16.64 ±1.5 |
| Vascular bundles, count | 23–25 23.8 ± 1 | 13–15 13.8 ± 1 | 15–17 15.5 ± 0.3 |
| Vascular bundles, height, μm | 25.9–177 81.7 ± 45.3 | 27.5–187 95.6 ± 45.8 | 33.2–190 104.6 ± 50.1 |
| Vascular bundles, width, μm | 17.9–103 59.2 ± 24.6 | 22.9–141 73.2 ±36.5 | 23.9–127 74 ± 29.8 |
| Vascular tissues in the bundles, % | 22.2–50.7 37.3 ± 8.2 | 7–61 19.9 ±9.3 | 4.1–30.8 19.4 ± 7.7 |
| Palisade tissue: thickness, μm | 13.7–77 44 ± 14.5 | 38–80 56.8 ± 8.8 | 25.1–75 57.9 ± 11.2 |
| Spongy tissue-thickness, μm | 25–91 48 ± 16.6 | 43–104 61.6 ± 13.8 | 33.8–65 46.3 ± 9.7 |
| White stripe, µm | 287–407 341 ± 49.2 | 150–295 241.1 ± 44.5 | 247–305 276.9 ± 22.8 |
| Adaxial epidermal cell: length, µm | 173–290 236 ± 47.3 | 98–431 237.2 ± 61.7 | 155–473 286.2 ± 66.8 |
| Adaxial epidermal cell: height, µm | 15–23.8 18.5 ± 1.96 | 11–22.6 17.83 ± 2.56 | 6.7–21 15.3 ± 3 |
| Adaxial epidermal cell: width, µm | 10.4–23.1 17.5 ± 2.93 | 11.5–20.8 16.45 ± 2.4 | 9.8–23.5 16.8 ± 3 |
| Palisade cell: height, µm | 13.8–36.4 26.4 ± 4.25 | 21.3–40.7 32.1 ± 6.49 | 17.2–41.3 30.3 ± 5.3 |
| Palisade cell: width, µm | 8.8–21.2 13.9 ± 2.47 | 9.6–18.6 13.53 ± 2.46 | 8.9–17.5 12.7 ± 2 |
| Spongy cell: height, µm | 14.4–29.6 21.4 ± 3.14 | 19.1–25.1 22.36 ± 2.07 | 13.9–28.6 20.7 ± 4.1 |
| Spongy cell: width, µm | 8.3–23.1 14.7 ± 3.39 | 13.7–17.9 15.74 ± 1.57 | 7.4–21.5 14.2 ± 2.6 |
| Abaxial epidermal cell: height, µm | 11.2–24.3 19.6 ± 2.92 | 15–28 18.2 ± 3.33 | 7.7–18.1 13.5 ± 2.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raycheva, T.; Stoyanov, K.; Naimov, S.; Apostolova-Kuzova, E.; Marinov, Y. Crocus pallidus (Iridaceae)—A Neglected Species for the Bulgarian Flora and Critical Taxon in the Balkans. Plants 2022, 11, 686. https://doi.org/10.3390/plants11050686
Raycheva T, Stoyanov K, Naimov S, Apostolova-Kuzova E, Marinov Y. Crocus pallidus (Iridaceae)—A Neglected Species for the Bulgarian Flora and Critical Taxon in the Balkans. Plants. 2022; 11(5):686. https://doi.org/10.3390/plants11050686
Chicago/Turabian StyleRaycheva, Tsvetanka, Kiril Stoyanov, Samir Naimov, Elena Apostolova-Kuzova, and Yulian Marinov. 2022. "Crocus pallidus (Iridaceae)—A Neglected Species for the Bulgarian Flora and Critical Taxon in the Balkans" Plants 11, no. 5: 686. https://doi.org/10.3390/plants11050686
APA StyleRaycheva, T., Stoyanov, K., Naimov, S., Apostolova-Kuzova, E., & Marinov, Y. (2022). Crocus pallidus (Iridaceae)—A Neglected Species for the Bulgarian Flora and Critical Taxon in the Balkans. Plants, 11(5), 686. https://doi.org/10.3390/plants11050686