
Effective Resistance to Four Fungal Foliar Diseases in Samples of Wild Triticum L. Species from the VIR (N.I. Vavilov All-Russian Institute of Plant Genetic Resources) Collection: View from Vavilov’s Concepts of Plant Immunity


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Pathogen Material
2.3. Seedling Resistance Screening

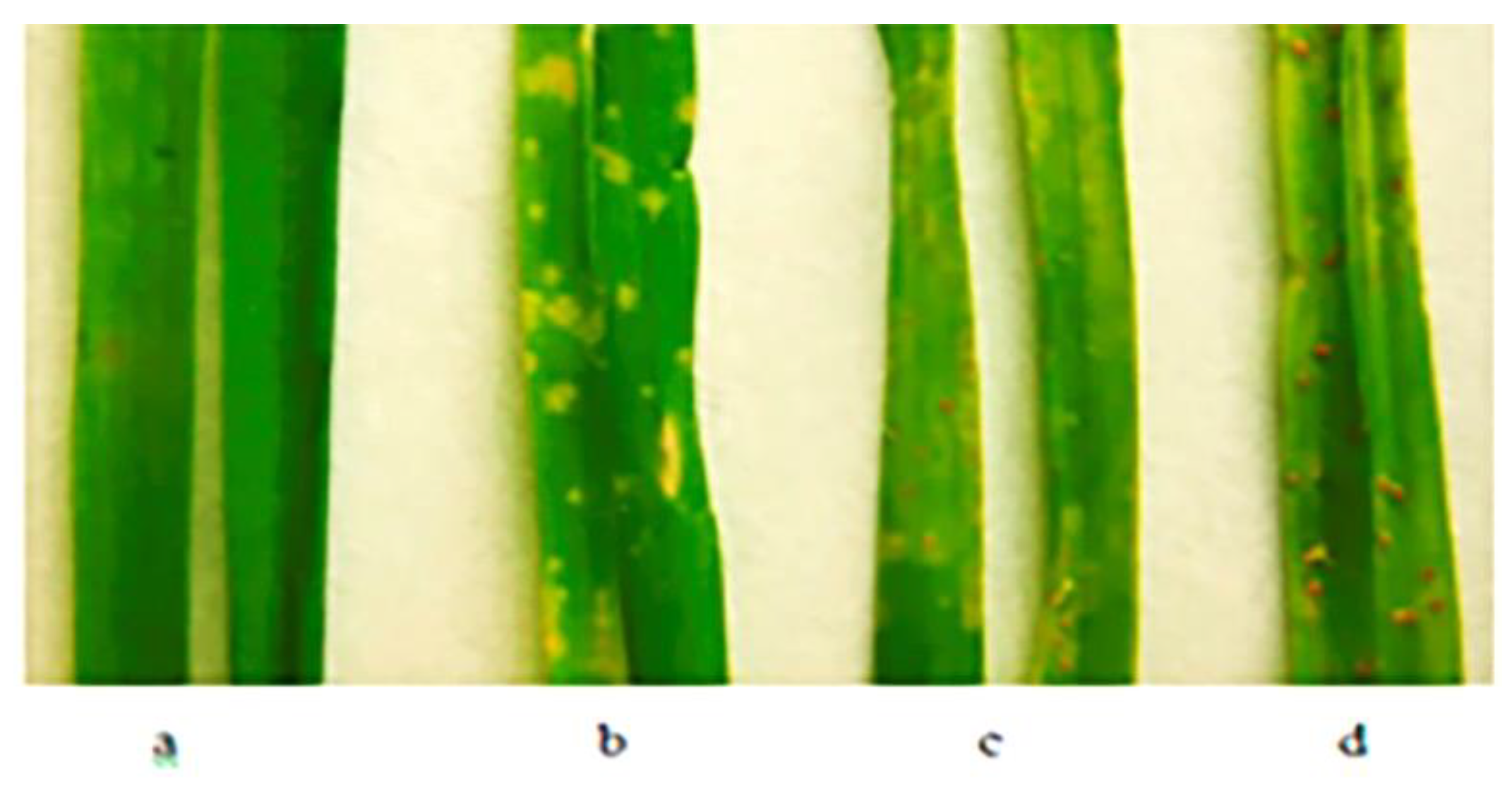{kind=link}
{kind=link}
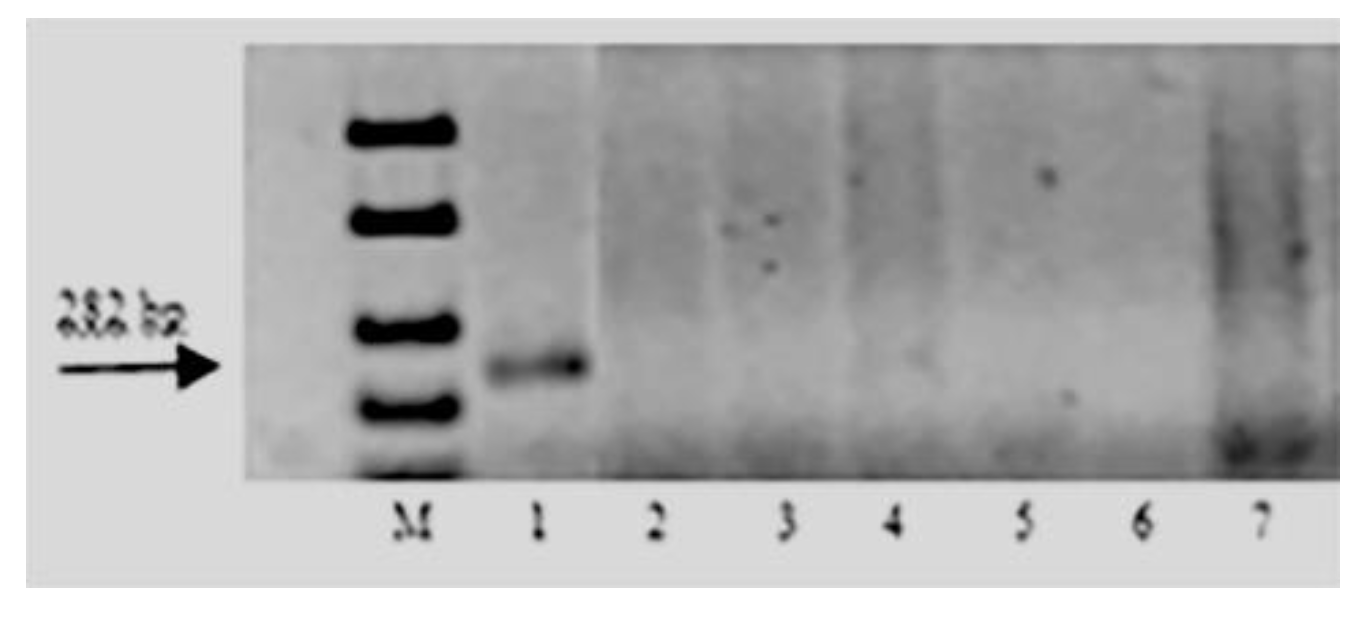{kind=link}
{kind=link}
{kind=link}
| Gene | Marker | Primer Sequence (5’-3’) | Fragment Size (bp) | Reference |
|---|---|---|---|---|
| Lr9 | SCS5550 | F: TGC GCC CTT CAA AGG AAG R: TGC GCC CTT CTG AAC TGT AT | 550 | [49] |
| Lr19 | Gb | F: CAT CCT TGG GGA CCT C R: CCA GCT CGC ATA CAT CCA | 130 | [50] |
| Lr24 | SCS73719 | F: TCG TCC AGA TCA GAA TGT G R: CTC GTC GAT TAG CAG TGA G | 719 | [51] |
| Lr41 | GDM35 | F: CCT GCT CTG CCC TAG ATA CG R: ATG TGA ATG TGA TGC ATG CA | 190 | [31] |
| Lr47 | PS10 | F: GCT GAT GAC CCT GAC CGG T R: TCT TCA TGC CCG GTC GGG T | 282 | [52] |
2.4. Adult Plant Resistance Screening
3. Results
3.1. Juvenile Resistance in Samples of Wild Wheat Species to the Diseases
3.2. Adult Resistance of Wild Wheat Species to the Diseases
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wegulo, S.N. Rust Diseases of Wheat. University of Nebraska–Lincoln Extension Neb Guide G2180, 2012. Available online: https://extensionpublications.unl.edu/assets/pdf/g2180.pdf (accessed on 6 November 2022).
- Krupnov, V.A. Strategy for genetic protection of wheat from leaf rust in the Volga region. Vestn. Russ. Agric. Sci. 1997, 6, 12–15. (In Russian) [Google Scholar]
- Germán, S.; Barcellos, A.; Chaves, M.; Kohli, M.; Campos, P.; Viedma, L. The situation of common wheat rusts in the Southern Cone of America and perspectives for control. Austr. J. Agric. Res. 2007, 58, 620–630. [Google Scholar] [CrossRef]
- Huerta-Espino, J.; Singh, R.P.; Germán, S.; McCallum, B.D.; Park, R.F.; Chen, W.Q.; Bhardwa, S.C.; Goyeau, H. Global status of wheat leaf rust caused by Puccinia triticina. Euphytica 2011, 179, 143–160. [Google Scholar] [CrossRef]
- Mehra, L.; Adhikari, U.; Cowger, C.; Ojiambo, P.S. Septoria nodorum blotch of wheat. PeerJ Prepr. 2018, 6, e27039v2. [Google Scholar]
- Chowdhury, A.K.; Singh, G.; Tyagi, B.S.; Ojha, A.; Dhar, T.; Bhattacharya, P.M. Spot blotch disease of wheat—A new thrust area for sustaining productivity. J. Wheat Res. 2013, 5, 1–11. [Google Scholar]
- Maxwell, J.J.; Lyerly, J.H.; Cowger, C.; Marshall, D.; Brown-Guedira, G.; Murphy, J.P. MlAG12: A Triticum timopheevii-derived powdery mildew resistance gene in common wheat on chromosome 7AL. Theor. Appl. Genet. 2009, 119, 1489–1495. [Google Scholar] [CrossRef]
- Vedeneeva, M.L.; Markelova, T.S.; Kirillova, T.V.; Anikeeva, N.V. Strategy of wheat disease-resistant varieties breeding in Volga region 1. Leaf rust, powdery mildew, loose smut and bunt. AgroХХI 2002, 2, 12–13. (In Russian) [Google Scholar]
- Spanic, V.; Matthew, N.R.; Kolmer, J.A.; Anderson, J.A. Leaf and stem seedling rust resistance in wheat cultivars grown in Croatia. Euphytica 2015, 203, 437–448. [Google Scholar] [CrossRef]
- Gessese, M.K. Description of wheat rusts and their virulence variations determined through annual pathotype surveys and controlled multi-pathotype tests. Adv. Agric. 2019, 2019, 2673706. [Google Scholar] [CrossRef]
- Singh, D.P.; Kumar, P. Role of spot blotch (Bipolaris sorokiniana) in deteriorating seed quality, its management in different wheat genotypes using fungicidal seed treatment. Indian Phytopathol. 2008, 61, 49–54. [Google Scholar]
- Vavilov, N.I. Selected Works in Two Volumes; Nauka: Leningrad, Russia, 1967; Volume 1, pp. 1–424. (In Russian) [Google Scholar]
- Vavilov, N.I. Origin and Geography of Cultivated Plants; Nauka: Leningrad, Russia, 1987; pp. 1–440. (In Russian) [Google Scholar]
- Tyryshkin, L.G. Genetic Diversity of Wheat and Barley for Effective Diseases Resistance and the Possibility of Its Broadening. Doctoral Dissertation, N.I. Vavilov All-Russian Institute of Plant Industry, Saint Petersburg, Russia, 2007. (In Russian). [Google Scholar]
- Tyryshkin, L.G.; Kolesova, M.A.; Kovaleva, M.M.; Lebedeva, T.V.; Zuev, E.V.; Brykova, A.N.; Gashimov, M.E. Current status of bread wheat and its relatives from VIR collection study for effective resistance to fungal diseases. In Proceedings of the 8th International Wheat Conference, Saint Petersburg, Russia, 1–4 June 2010; Dzyubenko, N.I., Vavilov, N.I., Eds.; Research Institute of Plant Industry (VIR): Saint Petersburg, Russia, 2010; pp. 134–135. (In Russian). [Google Scholar]
- Tyryshkin, L.G.; Syukov, V.V.; Zaharov, V.G.; Zuev, E.V.; Gashimov, M.E.; Kolesova, M.A.; Chikida, N.N.; Ershova, M.A.; Belousova, M.H. Sources of effective resistance to fungal diseases in wheat and its relatives—Search, creation and use in breeding. Proc. Appl. Bot. Genet. Breed. 2012, 170, 187–201. (In Russian) [Google Scholar]
- Meshkova, L.V.; Rosseeva, L.P.; Korenyuk, E.A.; Belan, I.A. Dynamics of distribution of the wheat leaf rust pathotypes virulent to the cultivars with Lr9 gene in Omsk region. Mikol. Fitopatol. 2012, 46, 397–400. (In Russian) [Google Scholar]
- Leonova, I.N.; Skolotneva, E.S.; Salina, E.A. Genome-wide association study of leaf rust resistance in Russian spring wheat varieties. BMC Plant Biol. 2020, 20, 135. [Google Scholar] [CrossRef] [PubMed]
- Skolotneva, E.S.; Leonova, I.N.; Bukatich, E.Y.; Boiko, N.I.; Piskarev, V.V.; Salina, E.A. Effectiveness of leaf rust resistance genes against Puccinia triticina populations in Western Siberia during 2008–2017. J. Plant Dis. Prot. 2018, 125, 549–555. [Google Scholar] [CrossRef]
- Markelova, T.S. Study of the structure and variability of the wheat brown rust population in the Volga region. AgroХХI 2007, 4–6, 37–40. (In Russian) [Google Scholar]
- Roelfs, A.P.; Singh, R.P.; Saari, E.E. Rust Diseases of Wheat: Concepts and Methods of Disease Management; CIMMYT: Mexico City, Mexico, 1992; pp. 1–81. [Google Scholar]
- McCallum, B.D.; Seto-Goh, P.; Xue, A. Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2008. Can. J. Plant. Pathol. 2011, 33, 541–549. [Google Scholar] [CrossRef]
- Huerta-Espino, J.; Singh, R.P.; Reyna-Martinez, J. First detection of virulence to genes Lr9 and Lr25 conferring resistance to leaf rust of wheat caused by Puccinia triticina in Mexico. Plant Dis. 2008, 92, 311. [Google Scholar] [CrossRef]
- Hanzalová, A.; Bartoš, P. The virulence spectrum of the wheat leaf rust population analyzed in the Czech Republic from 2002 to 2011. Czech J. Genet. Plant Breed. 2014, 50, 288–292. [Google Scholar] [CrossRef]
- El-Orabey, W.M. Virulence of some Puccinia triticina races to the effective wheat leaf rust resistant genes Lr9 and Lr19 under Egyptian field conditions. Physiol. Mol. Plant Pathol. 2018, 102, 163–172. [Google Scholar] [CrossRef]
- Huerta-Espino, J.; Singh, R.P. First report of virulence to wheat with leaf rust resistance gene Lr19 in Mexico. Plant Dis. 1994, 78, 640. [Google Scholar] [CrossRef]
- Bhardwaj, S.C.; Prashar, M.; Kumar, S.; Jain, S.K.; Datta, D. Lr19 resistance in wheat becomes susceptible to Puccinia triticina in India. Plant Dis. 2005, 89, 1360. [Google Scholar] [CrossRef] [PubMed]
- Boshoff, W.H.P.; Labuschagne, R.; Terefe, T.; Pretorius, Z.A.; Visser, B. New Puccinia triticina races on wheat in South Africa. Australas. Plant Pathol. 2018, 47, 325–334. [Google Scholar] [CrossRef]
- Figlan, S.; Ntushelo, K.; Mwadzingeni, L.; Terefe, T.; Tsilo, T.J.; Shimelis, H. Breeding wheat for durable leaf rust resistance in southern Africa: Variability, distribution, current control strategies, challenges, and future prospects. Front. Plant Sci. 2020, 11, 549. [Google Scholar] [CrossRef] [PubMed]
- Park, R.F. Breeding cereals for rust resistance in Australia. Plant Pathol. 2008, 57, 591–602. [Google Scholar] [CrossRef]
- Singh, S.; Franks, C.D.; Huang, L.; Brown-Guedira, G.L.; Marshall, D.S.; Gill, B.S.; Fritz, A. Lr41, Lr39, and a leaf rust resistance gene from Aegilops cylindrica may be allelic and are located on wheat chromosome 2DS. Theor. Appl. Genet. 2004, 108, 586–591. [Google Scholar] [CrossRef]
- Tyryshkin, L.G.; Gashimov, M.E.; Kolesova, M.A.; Anphilova, N.A. Juvenile resistance to diseases in samples of Triticum L. species from VIR World Collection. Cereal Res. Comm. 2006, 34, 1073–1079. [Google Scholar] [CrossRef]
- Vavilov, N.I. Selected Works in Two Volumes; Nauka: Leningrad, Russia, 1967; Volume 2, pp. 1–480. (In Russian) [Google Scholar]
- Yamaleev, A.M.; Dolotovskij, I.M.; Nuriakhmetov, D.F.; Zolotov, A.L. Resistance in various species of wheats and Aegilops to Septoria nodorum Berk. Proc. Appl. Bot. Genet. Breed. 1990, 132, 44–50. (In Russian) [Google Scholar]
- Makarova, N.A.; Lebedeva, T.V.; Radchenko, E.E. Wheat. (Immunological characteristics of rare species). In Catalogue of the VIR World Collection; N.I. Vavilov All-Russian Institute of Plant Industry: Saint Petersburg, Russia, 1993; Volume 640, pp. 1–59. (In Russian) [Google Scholar]
- Mikhailova, L.A.; Kovalenko, N.M.; Smurova, S.G.; Ternyuk, I.G.; Mitrofanova, O.P.; Lyapunova, O.A.; Zuev, E.V.; Chikida, N.N.; Loskutova, N.P.; Pyukkenen, V.P. Resistance of Triticum L. and Aegilops L. species from the VIR collection to tan and dark-brown leaf spots (catalogue). In Catalogue; All-Russian Institute of Plant Protection: Saint Petersburg, Russia, 2007; pp. 1–60. (In Russian) [Google Scholar]
- Dorofeev, V.F.; Filatenko, A.A.; Migushova, E.F.; Udachin, R.A.; Jacubziner, M.M. Cultural Flora of USSR. Wheat; Kolos: Leningrad, Russia, 1979; pp. 1–348. (In Russian) [Google Scholar]
- Dorofeev, V.F.; Udachin, R.A.; Semenova, L.V.; Novikova, M.V.; Gradchaninova, O.D.; Shitova, I.P.; Merezhko, A.F.; Filatenko, A.A. Wheats of the World, 2nd ed.; Agropromisdat: Leningrad, Russia, 1987; pp. 1–560. (In Russian) [Google Scholar]
- Kolesova, M.M.; Lysenko, N.S.; Tyryshkin, L.G. Resistance to diseases in samples of rare wheat species from the N.I. Vavilov All-Russian Institute of Plant Genetic Resources. Cereal Res. Commun. 2022, 50, 287–296. [Google Scholar] [CrossRef]
- Mains, E.В.; Jасkson, H.S. Physiological specialization in leaf rust of wheat Puccinia triticina Erikss. Phytopathology 1926, 16, 89–120. [Google Scholar]
- Mains, E.B.; Dietz, S.M. Physiologic forms of barley mildew, Erysiphe graminis hordei Marchal. Phytopathology 1930, 20, 229–239. [Google Scholar]
- Tyryshkin, L.G. Modification Variability of Virulence and Aggressiveness in Phytopathogens of Cereal Crops: Conclusions, Consequences, Possibilities of Practical Application; Saint Petersburg State Agrarian University: Saint Petersburg, Russia, 2016; pp. 1–137. (In Russian) [Google Scholar]
- McIntosh, R.A.; Yamazaki, Y.; Dubcovsky, J.; Rogers, J.; Morris, C.; Appels, R.; Xia, X.C. Catalogue of Gene Symbols for Wheat. 2013. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/macgene/2013/GeneSymbol.pdf (accessed on 3 September 2022).
- Dzhenin, S.V.; Lapochkina, I.F.; Zhemchuzhina, A.I.; Kovalenko, E.D. Donors of spring wheat resistant to the leaf rust with the genetic material of Aegilops speltoides L., Aegilops triuncialis L., Triticum kiharae Dorof. et Migusсh. Russ. Agric. Sci. 2009, 5, 3–7. (In Russian) [Google Scholar]
- Gajnullin, N.R.; Lapochkina, I.F.; Zhemchuzhina, A.I.; Kiseleva, M.I.; Kolomiets, T.M.; Kovalenko, E.D. Phytopathological and molecular genetic identification of leaf rust resistance genes in common wheat accessions with alien genetic material. Russ. J. Genet. 2007, 43, 875–881. (In Russian) [Google Scholar] [CrossRef]
- Hovhannisyan, N.A.; Dulloo, M.E.; Yesayan, A.H.; Knüpffer, H.; Amri, A. Tracking of powdery mildew and leaf rust resistance genes in Triticum boeoticum and T. urartu, wild relatives of common wheat. Czech J. Genet. Plant Breed. 2011, 47, 45–57. [Google Scholar] [CrossRef]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucl. Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef]
- Dorokhov, D.B.; Klocke, E. A rapid and economic technique for RAPD analysis of plant genomes. Russ. J. Genet. 1997, 33, 358–365. [Google Scholar]
- Gupta, S.K.; Charpe, A.; Koul, S.; Prabhu, K.V.; Haq, Q.M.R. Development and validation of molecular markers linked to an Aegilops umbellulate-derived leaf-rust-resistance gene, Lr9, for marker-assisted selection in bread wheat. Genome 2005, 48, 823–830. [Google Scholar] [CrossRef]
- Prins, R.; Groenewald, J.Z.; Marais, G.F.; Snape, J.W.; Koebner, R.M.D. AFLP and STS tagging of Lr19, a gene conferring resistance to leaf rust in wheat. Theor. Appl. Genet. 2001, 103, 618–624. [Google Scholar] [CrossRef]
- Prabhu, K.V.; Gupta, S.K.; Charpe, A.; Koul, S. SCAR marker tagged to the alien leaf rust resistance gene Lr19 uniquely marking the Agropyron elongatum-derived gene Lr24 in wheat: A revision. Plant Breed. 2004, 123, 417–420. [Google Scholar] [CrossRef]
- Helguera, M.; Khan, I.A.; Dubcovsky, J. Development of PCR markers for the wheat leaf rust resistance gene Lr47. Theor. Appl. Genet. 2000, 100, 1137–1143. [Google Scholar] [CrossRef]
- Hetzler, J. Host-Pathogen Interactions in Populations of Bipolaris sorokiniana in the Warm Non-Traditional Areas. Ph.D. Thesis, Georg August University Göttingen, Göttingen, Germany, 1992. [Google Scholar]
- Kumar, J.; Schäfer, P.; Hückelhoven, R.; Langen, G.; Baltruschat, H.; Stein, E.; Nagarajan, S.; Kogel, K.H. Bipolaris sorokiniana, a cereal pathogen of global concern: Cytological and molecular approaches towards better control. Mol. Plant Pathol. 2002, 3, 185–195. [Google Scholar] [CrossRef]
- Duveiller, E.M.; Sharma, R.C. Genetic improvement, and crop management strategies to minimize yield losses in warm non-traditional wheat growing areas due to spot blotch pathogen Cochliobolus sativus. J. Phytopathol. 2009, 157, 521–534. [Google Scholar] [CrossRef]
- Joshi, A.K.; Chand, R.; Arun, B. Relationship of plant height and days to maturity with resistance to spot blotch in wheat. Euphytica 2002, 123, 221–228. [Google Scholar] [CrossRef]
- Smurova, S.G. New Sources and Donors of Wheat Resistance to Cochliobolus sativus Drechs. ex Dastur. Candidate Dissertation (Ph.D.), All-Russian Institute of Plant Protection, Saint Petersburg, Russia, 2008. (In Russian). [Google Scholar]
- Askhadullin, D.F.; Askhadullin, D.F.; Vasilova, N.Z.; Khusainova, I.I.; Bagavieva, E.Z.; Tazutdinova, M.R. Dark-brown leaf spot on spring bread wheat in Tatarstan. Plant Prot. Quar. 2018, 9, 17–19. (In Russian) [Google Scholar]
- Alexandrov, A.E. Sources of Spring Bread Wheat Resistance to Powdery Mildew in the Lower Volga Region. Candidate Dissertation (Ph.D.), NIISH of the South-East, Saratov, Russia, 2000. (In Russian). [Google Scholar]
- Smurova, S.G.; Mikhailova, L.A. Sources of resistance to wheat spot blotch. Russ. Agric. Sci. 2007, 33, 378–380. [Google Scholar] [CrossRef]
- Kolesnikov, L.E.; Vlasova, E.A.; Vinogradov, A.A. On wheat leaf blotch agent’s development in plants of collection samples of soft wheat. Agric. Biol. 2009, 5, 90–93. (In Russian) [Google Scholar]
- Lebedeva, T.V.; Zuev, E.V. Studies of powdery mildew resistance (Blumeria graminis f. sp. tritici Golov.) in varieties of common wheat (Triticum aestivum L.). Ach. Sci. Technol. AIC 2015, 29, 17–20. (In Russian) [Google Scholar]
- Sadovaya, A.S.; Gultyaeva, E.I.; Mitrofanova, O.P.; Shaidayuk, E.L.; Hakimova, A.G.; Zuev, E.V. Leaf rust resistance in common wheat varieties and lines from the collection of the Vavilov Plant Industry Institute carrying alien genetic material. Russ. J. Genet. Appl. Res. 2015, 5, 233–241. [Google Scholar] [CrossRef]
- Kolomiets, T.M.; Pankratova, L.F.; Skatenok, O.O.; Pakholkova, E.V. Creation of genebank of wheat resistance sources to Septoriosis. Plant Prot. Quar. 2015, 7, 44–46. (In Russian) [Google Scholar]
- Podgorny, S.V.; Samofalov, A.P.; Skripka, O.V. Collection Samples of Soft Winter Wheat Tolerant to Leaf Rust and Powdery Mildew. Available online: http://ej.kubagro.ru/2015/09/pdf/105.pdf (accessed on 12 June 2022).
- Riaz, A.; Athiyannan, N.; Periyannan, S.; Afanasenko, O.; Mitrofanova, O.; Aitken, E.A.B.; Lagudah, E.; Hickey, L.T. Mining Vavilov’s treasure chest of wheat diversity for adult plant resistance to Puccinia triticina. Plant Dis. 2017, 101, 317–323. [Google Scholar] [CrossRef]
- Riaz, A.; Athiyannan, N.; Periyannan, S.K.; Afanasenko, O.; Mitrofanova, O.P.; Platz, G.J.; Aitken, E.A.B.; Snowdon, R.J.; Lagudah, E.S.; Hickey, L.T.; et al. Unlocking new alleles for leaf rust resistance in the Vavilov wheat collection. Theor. Appl. Genet. 2018, 131, 127–144. [Google Scholar] [CrossRef]
- Phan, H.T.T.; Rybak, K.; Bertazzoni, S.; Furuki, E.; Dinglasan, E.; Hickey, L.T.; Oliver, R.P.; Tan, K.C. Novel sources of resistance to Septoria nodorum blotch in the Vavilov wheat collection identified by genome-wide association studies. Theor. Appl. Genet. 2018, 131, 1223–1238. [Google Scholar] [CrossRef] [PubMed]
- Tyryshkin, L.G.; Volkova, G.V.; Kolomiets, T.M.; Brykova, A.N.; Zuev, E.V. Effective resistance to leaf rust in spring bread wheat accessions among recent additions to the VIR collection. Vavilovia 2019, 2, 35–43. (In Russian) [Google Scholar] [CrossRef]
- Lebedeva, T.V.; Zuev, E.V.; Brykova, A.N. Prospects of employing modern European cultivars of spring bread wheat in the breeding for powdery mildew resistance in the Northwestern region of Russia. Proc. Appl. Bot. Genet. Breed. 2019, 180, 170–176. (In Russian) [Google Scholar] [CrossRef][Green Version]
- Lebedeva, T.V.; Brykova, A.N.; Zuev, E.V. Powdery mildew resistance of Nordic spring bread wheat accessions from the Collection of the Vavilov Institute (VIR). Proc. Appl. Bot. Genet. Breed. 2020, 181, 146–154. (In Russian) [Google Scholar] [CrossRef]
- Tyryshkin, L.G.; Tyryshkina, N.A. Resistance to diseases in wheat collection samples and somaclonal variants. Czech. J. Genet. Plant Breed. 2003, 39, 21–23. [Google Scholar] [CrossRef]
- Tyryshkin, L.G.; Ershova, M.A. Inheritance of resistance to Septoria blotch in common wheat sample MN81330. Russ. J. Genet. 2004, 40, 454–457. [Google Scholar] [CrossRef]
- Anikster, Y.; Manisterski, J.; Long, D.L.; Leonard, K.J. Leaf rust and stem rust resistance in Triticum dicoccoides populations in Israel. Plant Dis. 2005, 89, 55–62. [Google Scholar] [CrossRef]
- Elkot, A.F.; Singh, R.; Kaur, S.; Kaur, J.; Chhuneja, P. Mapping novel sources of leaf rust and stripe rust resistance introgressed from Triticum dicoccoides in cultivated tetraploid wheat background. J. Plant Biochem. Biotechnol. 2021, 30, 336–342. [Google Scholar] [CrossRef]
- Marais, G.F.; Pretorius, Z.A.; Wellings, C.R.; McCallum, B.; Marais, A.S. Leaf rust and stripe rust resistance genes transferred to common wheat from Triticum dicoccoides. Euphytica 2005, 143, 115–123. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Bernardo, A.; Bai, G.; Hayden, M.J.; Anderson, J.A. Thatcher wheat line RL6149 carries Lr64 and a second leaf rust resistance gene on chromosome 1DS. Theor. Appl. Genet. 2019, 132, 2809–2814. [Google Scholar] [CrossRef]
- Brown-Guedira, G.L.; Gill, B.S.; Cox, T.S.; Leath, S. Transfer of disease resistance genes from Triticum araraticum to common wheat. Plant Breed. 1997, 116, 105–112. [Google Scholar] [CrossRef]
- Brown-Guedira, G.L.; Singh, S.; Fritz, A.K. Performance and mapping of leaf rust resistance transferred to wheat from Triticum timopheevii subsp. armeniacum. Phytopathology 2003, 93, 784–789. [Google Scholar] [CrossRef]
- Lebedeva, T.V.; Zuev, E.V. Inheritance of powdery mildew resistance in selected spring bread wheat accessions from the VIR collection. Vavilovia 2018, 1, 18–24. (In Russian) [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, K.; Dong, L.; Liang, Y.; Li, G.; Fang, T.; Guo, G.; Wu, Q.; Xie, J.; Chen, Y.; et al. Wheat powdery mildew resistance gene Pm64 derived from wild emmer (Triticum turgidum var. dicoccoides) is tightly linked in repulsion with stripe rust resistance gene Yr5. Crop J. 2019, 7, 761–770. [Google Scholar] [CrossRef]
- Xie, C.; Sun, Q.; Ni, Z.; Yang, T.; Nevo, E.; Fahima, T. Chromosomal location of a Triticum dicoccoides-derived powdery mildew resistance gene in common wheat by using microsatellite markers. Theor. Appl. Genet. 2003, 106, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Mergoum, M.; Ali, S.; Adhikari, T.B.; Elias, E.M.; Hughes, G.R. Identification of new sources of resistance to tan spot, Stagonospora nodorum blotch, and Septoria tritici blotch of wheat. Crop Sci. 2006, 46, 2047–2053. [Google Scholar] [CrossRef]
- Chu, C.G.; Xu, S.S.; Faris, J.D.; Nevo, E.; Friesen, T.L. Seedling resistance to tan spot and Stagonospora nodorum leaf blotch in wild emmer wheat (Triticum dicoccoides). Plant Dis. 2008, 92, 1229–1236. [Google Scholar] [CrossRef]
- Vavilov, N.I. Materials on the question of the resistance of cereals against parasitic fungi. Proc. Breed. Stn. Mosc. Agric. Inst. 1913, 1, 1–118. (In Russian) [Google Scholar]
- Vavilov, N.I. Plant immunity to infectious diseases. Proc. Petrovsk. Agric. Acad. 1918, 1–4, 1–244. (In Russian) [Google Scholar]
- Vavilov, N.I. The theory of plant immunity to infectious diseases. In Theoretical Bases of Plant Breeding; State Publishing House of State and Collective Farms Literature: Moscow/Leningrad, Russia, 1935; Volume 1, pp. 893–990. (In Russian) [Google Scholar]
- Vavilov, N.I. Laws of natural immunity of plants to infectious diseases (keys to finding immune forms). Izv. Akad. Nauk. SSSR. Ser. Biol. 1961, 1, 117–157. (In Russian) [Google Scholar]
- Tyryshkin, L.G. Vavilov’s laws of natural immunity to diseases: A modern view. In Proceedings of the Conference Scientific Support for the Development of Agriculture Industrial Complex under Conditions of Import Substitution, Saint Petersburg, Russia, 25–27 January 2018; Zhgulev, E.V., Ed.; Ministry of Agriculture of the Russian Federation, Saint Petersburg State Agrarian University: Saint Petersburg, Russia, 2018. Part I. pp. 41–45. (In Russian). [Google Scholar]
- Tyryshkin, L.G.; Gashimov, M.E.; Petrova, N.S.; Zveinek, I.A.; Kovaleva, O.N.; Chernov, V.E. Effective barley resistance to foliar fungal diseases (leaf rust, powdery mildew, dark-brown leaf spot blotch). Proc. Appl. Bot. Genet. Breed. 2013, 171, 57–60. (In Russian) [Google Scholar]





| Species | 2n | Genome | Country | VIR Catalogue No, kk- |
|---|---|---|---|---|
T. urartu Thum. ex Gandil. | 14 | Аu | Armenia Iran Jordan Lebanon Syria Turkey | 33870, 33871 58497 66361, 66362 64772–64780, 64782–64784 62458, 62464, 62479, 65752 58504, 64768–64771, 64786–64788, 64790, 66371, 67176, 67390 |
T. boeoticum Boiss. | 14 | Аb | Armenia Azerbaijan Bulgaria Greece Iran Iraq Syria Turkey Russia Ukraine | 58506, 58509, 58511, 58674, 59166, 64831, 64832, 65530, 65531, 65538, 65540 28280, 28283, 58489, 59159, 59163 62491, 62492 61660 65753 40117, 65543 65754, 65755 27141, 65952, 65970, 66369, 66370 18424 59178, 65535 |
| T. dicoccoides (Koern. ex Aschers. et Graebn.) Schweinf. | 28 | АuB | Iraq Israel Jordan Lebanon Turkey Syria | 42632, 67163 5199, 15903, 15907, 17256, 17259, 20403, 26117, 26119, 42633, 61678, 61681, 61690, 61692, 61697, 61707, 61709, 61720, 61722, 61723, 61730, 61732, 61742, 61750, 61753, 61754, 61761, 61763, 61764, 61766–61768, 61778, 61779, 61781, 61782, 61785, 61794, 61805, 61811, 61812, 61818, 61820, 61828, 61832, 61833, 62329, 62333, 62338, 62345, 62347, 67164, 67177 65748, 65550–65552, 65750, 67174, 67175 65751 66434, 66436 17157, 61712, 61714, 61717, 61719, 62362, 62364, 62366, 67161, 67162 |
T. araraticum Jakubz. | 28 | АbG | Armenia Azerbaijan Iraq Turkey | 28132, 30258, 31628, 59940, 61654, 64838, 65541, 65542, 66115 28244, 28247, 30234, 30240, 30268, 58668 40120, 64836, 64837, 64847–64850, 66103–66106, 66108, 66110 66111–66114 64846, 66372 |
| VIR Catalogue No. kk- | Species | Botanical Variety | Growth Stage | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Seedlings, Inoculated with Population Clones Virulent to Genes * | Flag Leaves, Inoculated with | |||||||||
| Lr9 | Lr19 | Lr24 | Lr41 | Lr47 | Microchambers | Spraying | ||||
| 62492 | T. boeoticum | pseudoboeoticum | 0;−1 | 0 | 0 | 0 | 0 | 0 | 0; | 0; |
| 66369 | T. boeoticum | pseudoboeoticum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 64777 | T. urartu | spontaneoalbum | 0;−1 | 0 | 0 | 0 | 0 | 0 | 0; | 0; |
| 64782 | T. urartu | albonigricans | 0;−1 | 0 | 0 | 0 | 0 | 0 | 0; | 0; |
| 64783 | T. urartu | spontaneoalbum | 0;−1 | 0 | 0 | 0 | 0 | 0 | 0; | 0; |
| 66372 | T. araraticum | kurdistanicum | 0–2 | 0 | 0 | 0 | 0 | 0 | 0–2 | 0–2 |
| VIR Catalogue No, kk- | Species | Botanical Variety | Growth Stage | ||
|---|---|---|---|---|---|
| Seedlings | Flag Leaves, Inoculation with | ||||
| Microchambers | Spraying | ||||
| 61712 | T. dicoccoides | dicoccoides | 0 | 0 | 0 |
| 61720 | T. dicoccoides | vavilovii | 0 | 0 | 0 |
| 61732 | T. dicoccoides | aaronsohnii | 0 | 0 | 0 |
| 61753 | T. dicoccoides | namuricum | 0 | 0 | 0 |
| 61764 | T. dicoccoides | vavilovii | 0 | 0 | 0 |
| 61767 | T. dicoccoides | vavilovii | 0 | 0 | 0 |
| 61768 | T. dicoccoides | arabicum | 0 | 0 | 0 |
| 61805 | T. dicoccoides | namuricum | 0 | 0 | 0 |
| 61811 | T. dicoccoides | aaronsohnii | 0 | 0 | 0 |
| 61833 | T. dicoccoides | arabicum | 0 | 0 | 0 |
| 67175 | T. dicoccoides | dicoccoides | 0 | 0 | 0 |
| 58489 | T. boeoticum | pseudoboeoticum | 0 | 0 | 0 |
| 62491 | T. boeoticum | kovarskii | 0 | 0 | 0 |
| 66370 | T. boeoticum | boeoticum | 0 | 0 | 0 |
| 64846 | T. araraticum | araxicum | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyryshkin, L.G.; Lysenko, N.S.; Kolesova, M.A. Effective Resistance to Four Fungal Foliar Diseases in Samples of Wild Triticum L. Species from the VIR (N.I. Vavilov All-Russian Institute of Plant Genetic Resources) Collection: View from Vavilov’s Concepts of Plant Immunity. Plants 2022, 11, 3467. https://doi.org/10.3390/plants11243467
Tyryshkin LG, Lysenko NS, Kolesova MA. Effective Resistance to Four Fungal Foliar Diseases in Samples of Wild Triticum L. Species from the VIR (N.I. Vavilov All-Russian Institute of Plant Genetic Resources) Collection: View from Vavilov’s Concepts of Plant Immunity. Plants. 2022; 11(24):3467. https://doi.org/10.3390/plants11243467
Chicago/Turabian StyleTyryshkin, Lev G., Natalia S. Lysenko, and Maria A. Kolesova. 2022. "Effective Resistance to Four Fungal Foliar Diseases in Samples of Wild Triticum L. Species from the VIR (N.I. Vavilov All-Russian Institute of Plant Genetic Resources) Collection: View from Vavilov’s Concepts of Plant Immunity" Plants 11, no. 24: 3467. https://doi.org/10.3390/plants11243467
APA StyleTyryshkin, L. G., Lysenko, N. S., & Kolesova, M. A. (2022). Effective Resistance to Four Fungal Foliar Diseases in Samples of Wild Triticum L. Species from the VIR (N.I. Vavilov All-Russian Institute of Plant Genetic Resources) Collection: View from Vavilov’s Concepts of Plant Immunity. Plants, 11(24), 3467. https://doi.org/10.3390/plants11243467