
Bioinformatics in Plant Breeding and Research on Disease Resistance

Abstract
1. Introduction
2. Databases Provide Abundant Gene and Pathway Information to Study Plant Biology
3. Various Algorithms Create Possibilities for Customized Analysis
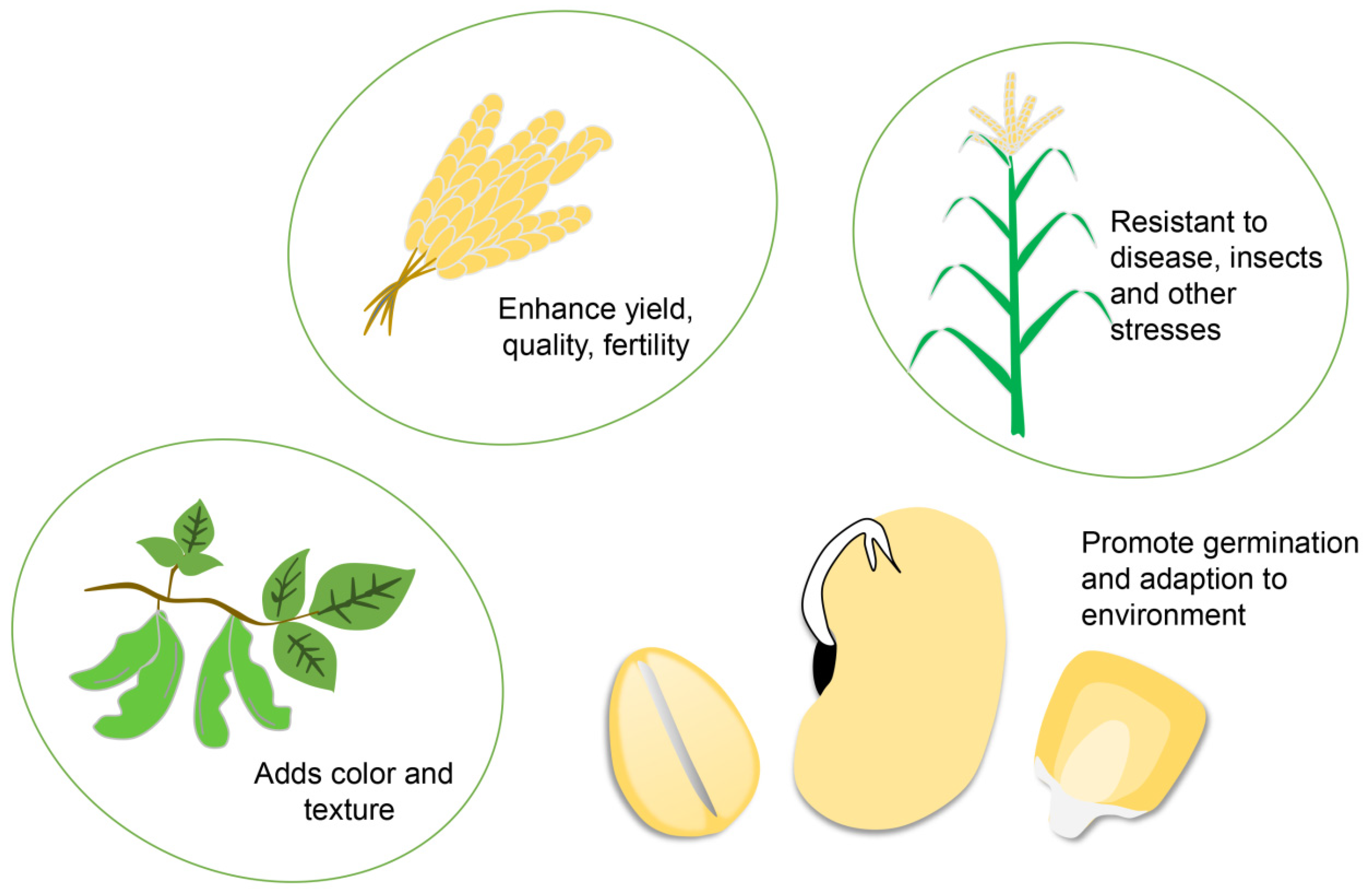
4. Application of Bioinformatics in Plant Breeding
4.1. Bioinformatics Can Be Applied to Breed Germplasm with High Yield and Quality
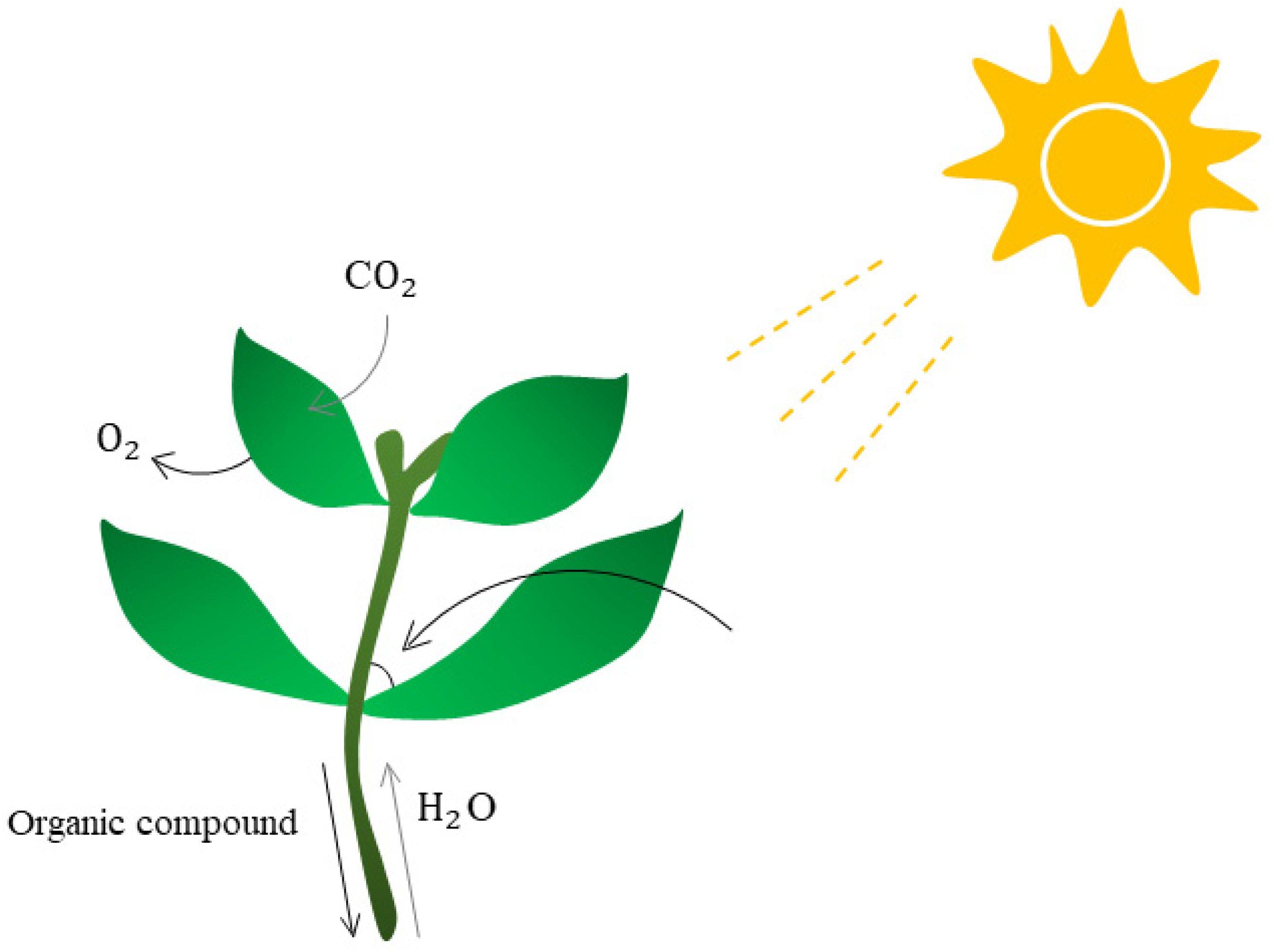
4.2. Bioinformatics Can Accurately Predict Plant Growth and Conditions
4.3. Automation for Agriculture
4.4. Accurate Prediction of Experimental Results and Transgenic Phenotypes
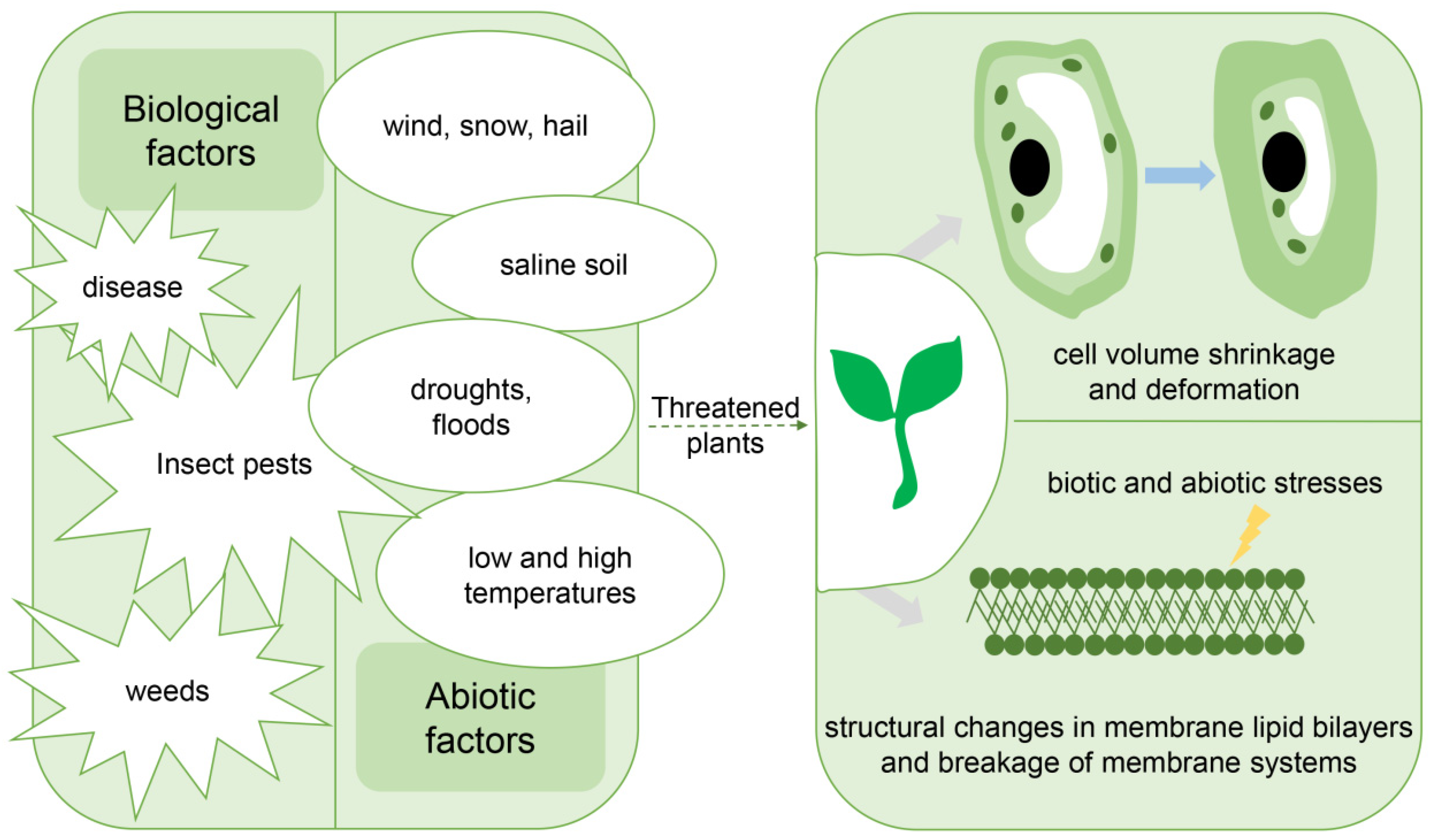
5. Application of Bioinformatics in Research on Plant Disease Resistance and Various Abiotic Stresses
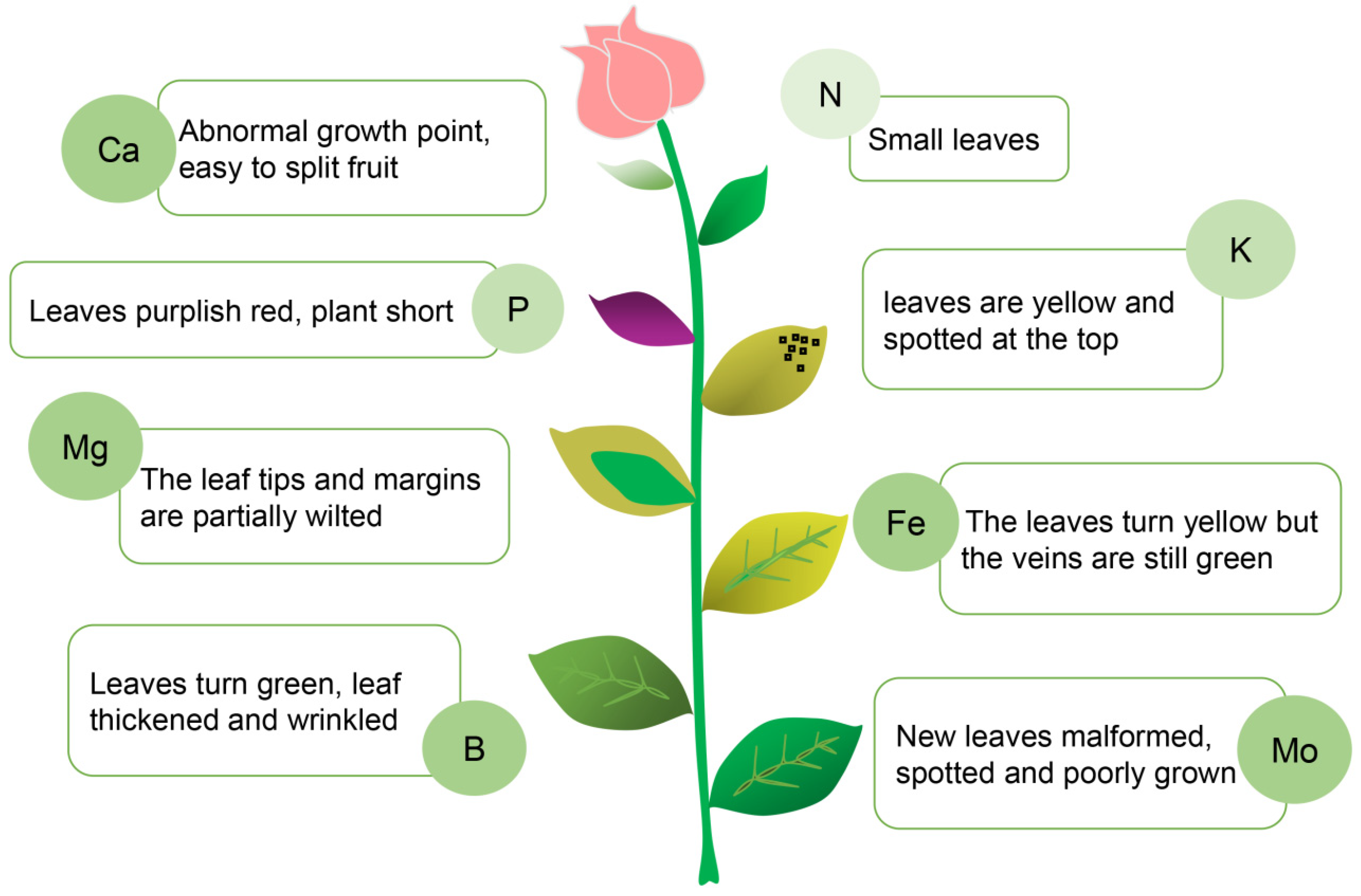
5.1. Predicting Plant Resistance Based on Key Indicators
5.2. Genome Annotation Is Available and Feasible
5.3. Multiple Genes Can Be Merged to Analyze Their Roles in Various Resistance
6. Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, C.; Huang, H.; Wu, C.H. Protein Bioinformatics Databases and Resources. Methods Mol. Biol. 2017, 1558, 3–39. [Google Scholar] [CrossRef] [PubMed]
- Małyska, A.; Jacobi, J. Plant breeding as the cornerstone of a sustainable bioeconomy. New Biotechnol. 2018, 40, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Sun, X.; Shen, J.; Gao, F.; Qiu, G.; Wang, T.; Nie, X.; Zhang, W.; Gao, Y.; Bai, Y. Molecular Evolutionary Analysis of Potato Virus Y Infecting Potato Based on the VPg Gene. Front. Microbiol. 2019, 10, 1708. [Google Scholar] [CrossRef] [PubMed]
- Blätke, M.A.; Szymanski, J.J.; Gladilin, E.; Scholz, U.; Beier, S. Advances in Applied Bioinformatics in Crops. Front. Plant Sci. 2021, 12, 640394. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, U.K.S.; Deo, I.; Jaiswal, J.P.; Prasad, B. Role of Bioinformatics in Crop Improvement. Glob. J. Sci. Front. Res. 2017, 17, 13–23. [Google Scholar]
- Alemu, K. The role and application of bioinformatics in plant disease management. Adv. Life Sci. Technol. 2015, 28, 28–33. [Google Scholar]
- Khan, A.; Singh, S.; Singh, V.K. Bioinformatics in Plant Pathology. In Emerging Trends in Plant Pathology; Singh, K.P., Jahagirdar, S., Sarma, B.K., Eds.; Springer: Singapore, 2021. [Google Scholar] [CrossRef]
- Vassilev, D.; Leunissen, J.; Atanassov, A.; Nenov, A.; Dimov, G. Application of Bioinformatics in Plant Breeding. Biotechnol. Biotechnol. Equip. 2005, 19, 139–152. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, Y.; Li, Y. Plant rejuvenation: From phenotypes to mechanisms. Plant Cell Rep. 2020, 39, 1249–1262. [Google Scholar] [CrossRef]
- Yasrab, R.; Zhang, J.; Smyth, P.; Pound, M.P. Predicting Plant Growth from Time-Series Data Using Deep Learning. Remote Sens. 2021, 13, 331. [Google Scholar] [CrossRef]
- Song, T.; Zhou, M.; Yuan, Y.; Yu, J.; Cai, H.; Li, J.; Chen, Y.; Bai, Y.; Zhou, G.; Cui, G. First high-quality reference genome of Amphicarpaea edgeworthii. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gomez-Casati, D.F.; Busi, M.V.; Barchiesi, J.; Peralta, D.A.; Hedin, N.; Bhadauria, V. Applications of Bioinformatics to Plant Biotechnology. Curr. Issues Mol. Biol. 2018, 27, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Houmard, N.M.; Mainville, J.L.; Bonin, C.P.; Huang, S.; Luethy, M.H.; Malvar, T.M. High-lysine corn generated by endosperm-specific suppression of lysine catabolism using RNAi. Plant Biotechnol. J. 2007, 5, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, R.H.; Fu, X.; Sun, X.; Tang, K. Over-expression of l-galactono-γ-lactone dehydrogenase increases vitamin C, total phenolics and antioxidant activity in lettuce through bio-fortification. Plant Cell Tissue Organ Cult. (PCTOC) 2013, 114, 225–236. [Google Scholar] [CrossRef]
- Li, J.; Scarano, A.; Gonzalez, N.M.; D’Orso, F.; Yue, Y.; Nemeth, K.; Saalbach, G.; Hill, L.; de Oliveira Martins, C.; Moran, R.; et al. Biofortified tomatoes provide a new route to vitamin D sufficiency. Nat. Plants 2022, 8, 611–616. [Google Scholar] [CrossRef]
- Ko, G.; Kim, P.-G.; Yoon, J.; Han, G.; Park, S.-J.; Song, W.; Lee, B. Closha: Bioinformatics workflow system for the analysis of massive sequencing data. BMC Bioinform. 2018, 19, 43. [Google Scholar] [CrossRef]
- Luciano, A.M.; Peluso, J.J. PGRMC1 and the faithful progression through mitosis and meiosis. Cell Cycle 2016, 15, 2239–2240. [Google Scholar] [CrossRef]
- Hoffmann, R.D.; Portes, M.T.; Olsen, L.I.; Damineli, D.S.C.; Hayashi, M.; Nunes, C.O.; Pedersen, J.T.; Lima, P.T.; Campos, C.; Feijó, J.A.; et al. Plasma membrane H+-ATPases sustain pollen tube growth and fertilization. Nat. Commun. 2020, 11, 2395. [Google Scholar] [CrossRef]
- Kim, T.; Lee, S.-H.; Kim, J.-O. A Novel Shape Based Plant Growth Prediction Algorithm Using Deep Learning and Spatial Transformation. IEEE Access 2022, 10, 37731–37742. [Google Scholar] [CrossRef]
- İncili, Ç.Y.; Arslan, B.; Çelik, E.N.Y.; Ulu, F.; Horuz, E.; Baloglu, M.C.; Çağlıyan, E.; Burcu, G.; Bayarslan, A.U.; Altunoglu, Y.C. Comparative bioinformatics analysis and abiotic stress responses of expansin proteins in Cucurbitaceae members: Watermelon and melon. Protoplasma 2022. [Google Scholar] [CrossRef]
- Aditya Shastry, K.; Sanjay, H.A. Hybrid prediction strategy to predict agricultural information. Appl. Soft Comput. 2020, 98, 106811. [Google Scholar] [CrossRef]
- Cock, P.J.A.; Grüning, B.A.; Paszkiewicz, K.; Pritchard, L. Galaxy tools and workflows for sequence analysis with applications in molecular plant pathology. PeerJ 2013, 1, e167. [Google Scholar] [CrossRef] [PubMed]
- Lucaciu, R.; Pelikan, C.; Gerner, S.M.; Zioutis, C.; Köstlbacher, S.; Marx, H.; Herbold, C.W.; Schmidt, H.; Rattei, T. A Bioinformatics Guide to Plant Microbiome Analysis. Front. Plant Sci. 2019, 10, 1313. [Google Scholar] [CrossRef] [PubMed]
- Dong, A.-Y.; Wang, Z.; Huang, J.-J.; Song, B.-A.; Hao, G.-F. Bioinformatic tools support decision-making in plant disease management. Trends Plant Sci. 2021, 26, 953–967. [Google Scholar] [CrossRef] [PubMed]
- Pundir, S.; Martin, M.J.; O’Donovan, C.; null, n. UniProt Tools. Curr. Protoc. Bioinform. 2016, 53, 1.29.1–1.29.15. [Google Scholar] [CrossRef]
- Darabi, M.; Seddigh, S. Computational study of biochemical properties of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) enzyme in C 3 plants. J. Plant Biol. 2017, 60, 35–47. [Google Scholar] [CrossRef]
- Reiser, L.; Subramaniam, S.; Li, D.; Huala, E. Using the Arabidopsis Information Resource (TAIR) to Find Information About Arabidopsis Genes. Curr. Protoc. Bioinform. 2017, 60, 1.11.1–1.11.45. [Google Scholar] [CrossRef]
- Zhang, J.; Li, C.; Wu, C.; Xiong, L.; Chen, G.; Zhang, Q.; Wang, S. RMD: A rice mutant database for functional analysis of the rice genome. Nucleic Acids Res. 2006, 34, D745–D748. [Google Scholar] [CrossRef]
- Wang, J.; Fu, W.; Wang, R.; Hu, D.; Cheng, H.; Zhao, J.; Jiang, Y.; Kang, Z. WGVD: An integrated web-database for wheat genome variation and selective signatures. Database J. Biol. Databases Curation 2020, 2020, baaa090. [Google Scholar] [CrossRef]
- Gui, S.; Yang, L.; Li, J.; Luo, J.; Xu, X.; Yuan, J.; Chen, L.; Li, W.; Yang, X.; Wu, S.; et al. ZEAMAP, a Comprehensive Database Adapted to the Maize Multi-Omics Era. iScience 2020, 23, 101241. [Google Scholar] [CrossRef]
- Liu, W.; Xie, X.; Ma, X.; Li, J.; Chen, J.; Liu, Y.-G. DSDecode: A Web-Based Tool for Decoding of Sequencing Chromatograms for Genotyping of Targeted Mutations. Mol. Plant 2015, 8, 1431–1433. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2013, 30, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-C.; Lee, T.-Y.; Huang, H.-D.; Huang, H.-Y.; Pan, R.-L. PlantPAN: Plant promoter analysis navigator, for identifying combinatorial cis-regulatory elements with distance constraint in plant gene groups. BMC Genom. 2008, 9, 561. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, Z.; Li, X.; Ni, Z.; Hu, Z.; Xin, M.; Peng, H.; Yao, Y.; Sun, Q.; Guo, W. SnpHub: An easy-to-set-up web server framework for exploring large-scale genomic variation data in the post-genomic era with applications in wheat. GigaScience 2020, 9, giaa060. [Google Scholar] [CrossRef] [PubMed]
- Ristić, M.M. R programming for bioinformatics. J. Appl. Stat. 2009, 36, 925. [Google Scholar] [CrossRef]
- Bioinformatics: Biological models in Python. Nat. Methods 2013, 10, 384. [CrossRef]
- Chen, X.-W.; Gao, J.X. Big Data Bioinformatics. Methods 2016, 111, 1–2. [Google Scholar] [CrossRef]
- Smith, A.M. FAO should focus on real not nominal food prices. Nature 2022, 602, 33. [Google Scholar] [CrossRef]
- Nordin, S.M.; Zolkepli, I.A.; Rizal, A.R.A.; Tariq, R.; Mannan, S.; Ramayah, T. Paving the way to paddy food security: A multigroup analysis of agricultural education on Circular Economy Adoption. J. Clean. Prod. 2022, 375, 134089. [Google Scholar] [CrossRef]
- Esposito, A.; Colantuono, C.; Ruggieri, V.; Chiusano, M.L. Bioinformatics for agriculture in the Next-Generation sequencing era. Chem. Biol. Technol. Agric. 2016, 3, 9. [Google Scholar] [CrossRef]
- Li, P.; Su, J.; Guan, Z.; Fang, W.; Chen, F.; Zhang, F. Association analysis of drought tolerance in cut chrysanthemum (Chrysanthemum morifolium Ramat.) at seedling stage. 3 Biotech 2018, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhang, F.; Chong, X.; Song, A.; Guan, Z.; Fang, W.; Chen, F. Genome-wide association study identifies favorable SNP alleles and candidate genes for waterlogging tolerance in chrysanthemums. Hortic. Res. 2019, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Chong, X.; Su, J.; Wang, F.; Wang, H.; Song, A.; Guan, Z.; Fang, W.; Jiang, J.; Chen, S.; Chen, F.; et al. Identification of favorable SNP alleles and candidate genes responsible for inflorescence-related traits via GWAS in chrysanthemum. Plant Mol. Biol. 2019, 99, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, K.; Arora, S.; Silva, P.; Sánchez-Martín, J.; Horsnell, R.; Gao, L.; Brar, G.S.; Widrig, V.; John Raupp, W.; Singh, N.; et al. Population genomic analysis of Aegilops tauschii identifies targets for bread wheat improvement. Nat. Biotechnol. 2021, 40, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Mascher, M.; Schreiber, M.; Scholz, U.; Graner, A.; Reif, J.C.; Stein, N. Genebank genomics bridges the gap between the conservation of crop diversity and plant breeding. Nat. Genet. 2019, 51, 1076–1081. [Google Scholar] [CrossRef]
- Hu, J.; Chen, B.; Zhao, J.; Zhang, F.; Xie, T.; Xu, K.; Gao, G.; Yan, G.; Li, H.; Li, L.; et al. Genomic selection and genetic architecture of agronomic traits during modern rapeseed breeding. Nat. Genet. 2022, 54, 694–704. [Google Scholar] [CrossRef]
- Cao, Y.; Zhong, Z.; Wang, H.; Shen, R. Leaf angle: A target of genetic improvement in cereal crops tailored for high-density planting. Plant Biotechnol. J. 2022, 20, 426–436. [Google Scholar] [CrossRef]
- An, G.; Qi, Y.; Zhang, W.; Gao, H.; Qian, J.; Larkin, R.M.; Chen, J.; Kuang, H. LsNRL4 enhances photosynthesis and decreases leaf angles in lettuce. Plant Biotechnol. J. 2022, 20, 1956–1967. [Google Scholar] [CrossRef]
- Peng, B.; Zhao, X.; Wang, Y.; Li, C.; Li, Y.; Zhang, D.; Shi, Y.; Song, Y.; Wang, L.; Li, Y.; et al. Genome-wide association studies of leaf angle in maize. Mol. Breed. 2021, 41, 50. [Google Scholar] [CrossRef]
- Kenchanmane Raju, S.K.; Adkins, M.; Enersen, A.; Santana de Carvalho, D.; Studer, A.J.; Ganapathysubramanian, B.; Schnable, P.S.; Schnable, J.C. Leaf Angle eXtractor: A high-throughput image processing framework for leaf angle measurements in maize and sorghum. Appl. Plant Sci. 2020, 8, e11385. [Google Scholar] [CrossRef]
- Maathuis, F.J.M. Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Alzarah, M.I. Macro and micro-elements concentrations in Calligonum comosum wild grazing plant through its growth period. Saudi J. Biol. Sci. 2021, 28, 6992–6999. [Google Scholar] [CrossRef] [PubMed]
- Holušová, K.; Vrána, J.; Šafář, J.; Šimková, H.; Balcárková, B.; Frenkel, Z.; Darrier, B.; Paux, E.; Cattonaro, F.; Berges, H.; et al. Physical Map of the Short Arm of Bread Wheat Chromosome 3D. Plant Genome 2017, 10, 1–11. [Google Scholar] [CrossRef]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The Impacts of Phosphorus Deficiency on the Photosynthetic Electron Transport Chain. Plant Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef]
- Aleksandrov, V. Identification of nutrient deficiency in plants by artificial intelligence. Acta Physiol. Plant. 2022, 44, 29. [Google Scholar] [CrossRef]
- Bochtis, D.D.; Sørensen, C.G.C.; Busato, P. Advances in agricultural machinery management: A review. Biosyst. Eng. 2014, 126, 69–81. [Google Scholar] [CrossRef]
- Hyde, J.; Engel, P. Investing in a robotic milking system: A Monte Carlo simulation analysis. J. Dairy Sci. 2002, 85, 2207–2214. [Google Scholar] [CrossRef]
- Baltazar, A.R.; Santos, F.N.d.; Moreira, A.P.; Valente, A.; Cunha, J.B. Smarter Robotic Sprayer System for Precision Agriculture. Electronics 2021, 10, 2061. [Google Scholar] [CrossRef]
- Suhaimi, A.F.; Yaakob, N.; Saad, S.A.; Sidek, K.A.; Elshaikh, M.E.; Dafhalla, A.K.; Lynn, O.B.; Almashor, M. IoT Based Smart Agriculture Monitoring, Automation and Intrusion Detection System. J. Phys. Conf. Ser. 2021, 1962, 012016. [Google Scholar] [CrossRef]
- Wurtzel, E.T.; Kutchan, T.M. Plant metabolism, the diverse chemistry set of the future. Science 2016, 353, 1232–1236. [Google Scholar] [CrossRef]
- Thackston, K.A.; Deheyn, D.D.; Sievenpiper, D.F. Limitations on electromagnetic communication by vibrational resonances in biological systems. Phys. Rev. E 2020, 101, 062401. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Fang, S.; Gao, J.; Zhang, Y.; Jin, G.; Wang, S.; Zhu, Y.; Xu, J. Could Vegetation Index be Derive from Synthetic Aperture Radar?—The Linear Relationship between Interferometric Coherence and NDVI. Sci. Rep. 2020, 10, 6749. [Google Scholar] [CrossRef] [PubMed]
- Guzzetta, G.; Jurman, G.; Furlanello, C. A machine learning pipeline for quantitative phenotype prediction from genotype data. BMC Bioinform. 2010, 11, S3. [Google Scholar] [CrossRef] [PubMed]
- Nigro, D.; Gadaleta, A.; Mangini, G.; Colasuonno, P.; Marcotuli, I.; Giancaspro, A.; Giove, S.L.; Simeone, R.; Blanco, A. Candidate genes and genome-wide association study of grain protein content and protein deviation in durum wheat. Planta 2019, 249, 1157–1175. [Google Scholar] [CrossRef]
- Blanco, A.; Mangini, G.; Giancaspro, A.; Giove, S.; Colasuonno, P.; Simeone, R.; Signorile, A.; De Vita, P.; Mastrangelo, A.M.; Cattivelli, L.; et al. Relationships between grain protein content and grain yield components through quantitative trait locus analyses in a recombinant inbred line population derived from two elite durum wheat cultivars. Mol. Breed. 2012, 30, 79–92. [Google Scholar] [CrossRef]
- Maccaferri, M.; Mantovani, P.; Tuberosa, R.; Deambrogio, E.; Giuliani, S.; Demontis, A.; Massi, A.; Sanguineti, M.C. A major QTL for durable leaf rust resistance widely exploited in durum wheat breeding programs maps on the distal region of chromosome arm 7BL. Theor. Appl. Genet. 2008, 117, 1225–1240. [Google Scholar] [CrossRef]
- Zhu, K.; Li, M.; Wu, H.; Zhang, D.; Dong, L.; Wu, Q.; Chen, Y.; Xie, J.; Lu, P.; Guo, G.; et al. Fine mapping of powdery mildew resistance gene MlWE74 derived from wild emmer wheat (Triticum turgidum ssp. dicoccoides) in an NBS-LRR gene cluster. Theor. Appl. Genet. 2022, 135, 1235–1245. [Google Scholar] [CrossRef]
- Blanco, A.; Colasuonno, P.; Gadaleta, A.; Mangini, G.; Schiavulli, A.; Simeone, R.; Digesù, A.M.; De Vita, P.; Mastrangelo, A.M.; Cattivelli, L. Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds in durum wheat. J. Cereal Sci. 2011, 54, 255–264. [Google Scholar] [CrossRef]
- Hara-Skrzypiec, A.; Śliwka, J.; Jakuczun, H.; Zimnoch-Guzowska, E. QTL for tuber morphology traits in diploid potato. J. Appl. Genet. 2018, 59, 123–132. [Google Scholar] [CrossRef]
- Badji, A.; Otim, M.; Machida, L.; Odong, T.; Kwemoi, D.B.; Okii, D.; Agbahoungba, S.; Mwila, N.; Kumi, F.; Ibanda, A.; et al. Maize Combined Insect Resistance Genomic Regions and Their Co-localization With Cell Wall Constituents Revealed by Tissue-Specific QTL Meta-Analyses. Front. Plant Sci. 2018, 9, 895. [Google Scholar] [CrossRef]
- Martins, L.B.; Rucker, E.; Thomason, W.; Wisser, R.J.; Holland, J.B.; Balint-Kurti, P. Validation and Characterization of Maize Multiple Disease Resistance QTL. G3 Genes Genomes Genet. 2019, 9, 2905–2912. [Google Scholar] [CrossRef] [PubMed]
- Maharijaya, A.; Vosman, B.; Steenhuis-Broers, G.; Pelgrom, K.; Purwito, A.; Visser, R.G.F.; Voorrips, R.E. QTL mapping of thrips resistance in pepper. Theor. Appl. Genet. 2015, 128, 1945–1956. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.; Izquierdo, P.; Jeffery, H.; Shaw, S.; Nchimbi-Msolla, S.; Cichy, K. QTL analysis of cooking time and quality traits in dry bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2020, 133, 2291–2305. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, A.D.J.; Kootstra, G.; Kruijer, W.; de Ridder, D. Machine learning in plant science and plant breeding. iScience 2020, 24, 101890. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Loo, E.P.-I. Plant immunity in signal integration between biotic and abiotic stress responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Savchenko, T.; Baidoo, E.E.; Chehab, W.E.; Hayden, D.M.; Tolstikov, V.; Corwin, J.A.; Kliebenstein, D.J.; Keasling, J.D.; Dehesh, K. Retrograde signaling by the plastidial metabolite MEcPP regulates expression of nuclear stress-response genes. Cell 2012, 149, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Parker, J.E. The impact of temperature on balancing immune responsiveness and growth in Arabidopsis. Trends Plant Sci. 2011, 16, 666–675. [Google Scholar] [CrossRef]
- Griffith, M.; Yaish, M.W.F. Antifreeze proteins in overwintering plants: A tale of two activities. Trends Plant Sci. 2004, 9, 399–405. [Google Scholar] [CrossRef]
- Catalá, R.; Medina, J.; Salinas, J. Integration of low temperature and light signaling during cold acclimation response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 16475–16480. [Google Scholar] [CrossRef]
- Olate, E.; Jiménez-Gómez, J.M.; Holuigue, L.; Salinas, J. NPR1 mediates a novel regulatory pathway in cold acclimation by interacting with HSFA1 factors. Nat. Plants 2018, 4, 811–823. [Google Scholar] [CrossRef]
- Feng, W.; Kita, D.; Peaucelle, A.; Cartwright, H.N.; Doan, V.; Duan, Q.; Liu, M.-C.; Maman, J.; Steinhorst, L.; Schmitz-Thom, I.; et al. The FERONIA Receptor Kinase Maintains Cell-Wall Integrity during Salt Stress through Ca2+ Signaling. Curr. Biol. 2018, 28, 666–675.e5. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, G.; Ferrari, S.; Cervone, F.; Okun, E. Extracellular DAMPs in Plants and Mammals: Immunity, Tissue Damage and Repair. Trends Immunol. 2018, 39, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, T.; Matsushita, Y.; Hosokawa, M. Chrysanthemum Stunt Viroid Resistance in Chrysanthemum. Viruses 2018, 10, 719. [Google Scholar] [CrossRef] [PubMed]
- Simón, D.; Borsani, O.; Filippi, C.V. RFPDR: A random forest approach for plant disease resistance protein prediction. PeerJ 2022, 10, e11683. [Google Scholar] [CrossRef]
- Longuespée, R.; Casadonte, R.; Schwamborn, K.; Reuss, D.; Kazdal, D.; Kriegsmann, K.; von Deimling, A.; Weichert, W.; Schirmacher, P.; Kriegsmann, J.; et al. Proteomics in Pathology. Proteomics 2018, 18, 1700361. [Google Scholar] [CrossRef]
- Knief, C. Analysis of plant microbe interactions in the era of next generation sequencing technologies. Front. Plant Sci. 2014, 5, 216. [Google Scholar] [CrossRef]
- Bashir, K.; Matsui, A.; Rasheed, S.; Seki, M. Recent advances in the characterization of plant transcriptomes in response to drought, salinity, heat, and cold stress. F1000Research 2019, 8, 658. [Google Scholar] [CrossRef]
- Garg, R.; Shankar, R.; Thakkar, B.; Kudapa, H.; Krishnamurthy, L.; Mantri, N.; Varshney, R.K.; Bhatia, S.; Jain, M. Transcriptome analyses reveal genotype- and developmental stage-specific molecular responses to drought and salinity stresses in chickpea. Sci. Rep. 2016, 6, 19228. [Google Scholar] [CrossRef]
- Chopra, R.; Burow, G.; Hayes, C.; Emendack, Y.; Xin, Z.; Burke, J. Transcriptome profiling and validation of gene based single nucleotide polymorphisms (SNPs) in sorghum genotypes with contrasting responses to cold stress. BMC Genom. 2015, 16, 1040. [Google Scholar] [CrossRef]
- Filichkin, S.A.; Hamilton, M.; Dharmawardhana, P.D.; Singh, S.K.; Sullivan, C.; Ben-Hur, A.; Reddy, A.S.N.; Jaiswal, P. Abiotic Stresses Modulate Landscape of Poplar Transcriptome via Alternative Splicing, Differential Intron Retention, and Isoform Ratio Switching. Front. Plant Sci. 2018, 9, 5. [Google Scholar] [CrossRef]
- González-Schain, N.; Dreni, L.; Lawas, L.M.F.; Galbiati, M.; Colombo, L.; Heuer, S.; Jagadish, K.S.V.; Kater, M.M. Genome-Wide Transcriptome Analysis During Anthesis Reveals New Insights into the Molecular Basis of Heat Stress Responses in Tolerant and Sensitive Rice Varieties. Plant Cell Physiol. 2015, 57, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yu, L.; Xuan, J.; Lu, Y.; Lu, S.; Zhu, W. De novo transcriptome sequencing and gene expression profiling of spinach (Spinacia oleracea L.) leaves under heat stress. Sci. Rep. 2016, 6, 19473. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, M.G.; Ishizaki, T.; Valencia, M.; Ogawa, S.; Dedicova, B.; Ogata, T.; Yoshiwara, K.; Maruyama, K.; Kusano, M.; Saito, K.; et al. Overexpression of an Arabidopsis thaliana galactinol synthase gene improves drought tolerance in transgenic rice and increased grain yield in the field. Plant Biotechnol. J. 2017, 15, 1465–1477. [Google Scholar] [CrossRef]
- Meng, H.; Sun, M.; Jiang, Z.; Liu, Y.; Sun, Y.; Liu, D.; Jiang, C.; Ren, M.; Yuan, G.; Yu, W.; et al. Comparative transcriptome analysis reveals resistant and susceptible genes in tobacco cultivars in response to infection by Phytophthora nicotianae. Sci. Rep. 2021, 11, 809. [Google Scholar] [CrossRef]
- Natarajan, P.; Ahn, E.; Reddy, U.K.; Perumal, R.; Prom, L.K.; Magill, C. RNA-Sequencing in Resistant (QL3) and Susceptible (Theis) Sorghum Cultivars Inoculated With Johnsongrass Isolates of Colletotrichum sublineola. Front. Genet. 2021, 12, 722519. [Google Scholar] [CrossRef]
- Lewis, S.E.; Searle, S.M.J.; Harris, N.; Gibson, M.; Lyer, V.; Richter, J.; Wiel, C.; Bayraktaroglu, L.; Birney, E.; Crosby, M.A.; et al. Apollo: A sequence annotation editor. Genome Biol. 2003, 3, 1–14. [Google Scholar] [CrossRef]
- Olsen, J.L.; Rouzé, P.; Verhelst, B.; Lin, Y.-C.; Bayer, T.; Collen, J.; Dattolo, E.; De Paoli, E.; Dittami, S.; Maumus, F.; et al. The genome of the seagrass Zostera marina reveals angiosperm adaptation to the sea. Nature 2016, 530, 331–335. [Google Scholar] [CrossRef]
- Yuan, F.; Wang, X.; Zhao, B.; Xu, X.; Shi, M.; Leng, B.; Dong, X.; Lu, C.; Feng, Z.; Guo, J.; et al. The genome of the recretohalophyte Limonium bicolor provides insights into salt gland development and salinity adaptation during terrestrial evolution. Mol. Plant 2022, 15, 1024–1044. [Google Scholar] [CrossRef]
- VanBuren, R.; Wai, C.M.; Ou, S.; Pardo, J.; Bryant, D.; Jiang, N.; Mockler, T.C.; Edger, P.; Michael, T.P. Extreme haplotype variation in the desiccation-tolerant clubmoss Selaginella lepidophylla. Nat. Commun. 2018, 9, 13. [Google Scholar] [CrossRef]
- Fukushima, K.; Fang, X.; Alvarez-Ponce, D.; Cai, H.; Carretero-Paulet, L.; Chen, C.; Chang, T.-H.; Farr, K.M.; Fujita, T.; Hiwatashi, Y.; et al. Genome of the pitcher plant Cephalotus reveals genetic changes associated with carnivory. Nat. Ecol. Evol. 2017, 1, 0059. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, F.; Zhang, X.; Li, Z.; Zhao, Y.; Lohaus, R.; Chang, X.; Dong, W.; Ho, S.Y.W.; Liu, X.; et al. The water lily genome and the early evolution of flowering plants. Nature 2019, 577, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Meyer, I.M.; Durbin, R. Comparative ab initio prediction of gene structures using pair HMMs. Bioinformatics 2002, 18, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Fraley, R.T.; Rogers, S.G.; Horsch, R.B.; Sanders, P.R.; Flick, J.S.; Adams, S.P.; Bittner, M.L.; Brand, L.A.; Fink, C.L.; Fry, J.S.; et al. Expression of bacterial genes in plant cells. Proc. Natl. Acad. Sci. USA 1983, 80, 4803–4807. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, A.; Arró, M.; Manzano, D.; Altabella, T. Strategies and Methodologies for the Co-expression of Multiple Proteins in Plants. Adv. Exp. Med. Biol. 2016, 896, 263–285. [Google Scholar] [CrossRef]
- Jobling, S.A.; Westcott, R.J.; Tayal, A.; Jeffcoat, R.; Schwall, G.P. Production of a freeze-thaw-stable potato starch by antisense inhibition of three starch synthase genes. Nat. Biotechnol. 2002, 20, 295–299. [Google Scholar] [CrossRef]
- Qi, B.; Fraser, T.; Mugford, S.; Dobson, G.; Sayanova, O.; Butler, J.; Napier, J.A.; Stobart, A.K.; Lazarus, C.M. Production of very long chain polyunsaturated omega-3 and omega-6 fatty acids in plants. Nat. Biotechnol. 2004, 22, 739–745. [Google Scholar] [CrossRef]
- Ma, J.K.C.; Hiatt, A.; Hein, M.; Vine, N.D.; Wang, F.; Stabila, P.; van Dolleweerd, C.; Mostov, K.; Lehner, T. Generation and assembly of secretory antibodies in plants. Science 1995, 268, 716–719. [Google Scholar] [CrossRef]
- van Erp, H.; Kelly, A.A.; Menard, G.; Eastmond, P.J. Multigene engineering of triacylglycerol metabolism boosts seed oil content in Arabidopsis. Plant Physiol. 2014, 165, 30–36. [Google Scholar] [CrossRef]
- Datta, K.; Baisakh, N.; Thet, K.M.; Tu, J.; Datta, S.K. Pyramiding transgenes for multiple resistance in rice against bacterial blight, yellow stem borer and sheath blight. Theor. Appl. Genet. 2003, 106, 1–8. [Google Scholar] [CrossRef]
- Houshyani, B.; Assareh, M.; Busquets, A.; Ferrer, A.; Bouwmeester, H.J.; Kappers, I.F. Three-step pathway engineering results in more incidence rate and higher emission of nerolidol and improved attraction of Diadegma semiclausum. Metab. Eng. 2013, 15, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Magi, A.; Benelli, M.; Gozzini, A.; Girolami, F.; Torricelli, F.; Brandi, M.L. Bioinformatics for Next Generation Sequencing Data. Genes 2010, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.K.; Kumar, S.; Jaiswal, A.; Prasad, M.; Gandomi, A.H. Towards Precision Agriculture: IoT-Enabled Intelligent Irrigation Systems Using Deep Learning Neural Network. IEEE Sens. J. 2021, 21, 17479–17491. [Google Scholar] [CrossRef]
- Deusch, S.; Tilocca, B.; Camarinha-Silva, A.; Seifert, J. News in livestock research—Use of Omics-technologies to study the microbiota in the gastrointestinal tract of farm animals. Comput. Struct. Biotechnol. J. 2015, 13, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Rott, M.; Xiang, Y.; Boyes, I.; Belton, M.; Saeed, H.; Kesanakurti, P.; Hayes, S.; Lawrence, T.; Birch, C.; Bhagwat, B.; et al. Application of Next Generation Sequencing for Diagnostic Testing of Tree Fruit Viruses and Viroids. Plant Dis. 2017, 101, 1489–1499. [Google Scholar] [CrossRef]
- Cao, Y.; Geddes, T.A.; Yang, J.Y.H.; Yang, P. Ensemble deep learning in bioinformatics. Nat. Mach. Intell. 2020, 2, 500–508. [Google Scholar] [CrossRef]
- Coleman-Derr, D.; Tringe, S.G. Building the crops of tomorrow: Advantages of symbiont-based approaches to improving abiotic stress tolerance. Front. Microbiol. 2014, 5, 283. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| URL | Note | Description |
|---|---|---|
| http://bigd.big.ac.cn/databasecommons/ Accessed on 4 May 2022. | Comprehensive publicly available data repository covering a wide range of organisms | Bigd database consolidates all the information collected about the database. Each database is classified by data type, category, subject, and location, so that people can easily find a specific collection of databases of interest. |
| https://www.expasy.org/ Accessed on 4 May 2022. | Covers a wide range of biological research databases and software tools | Expasy database is divided into several areas: DNA, RNA, protein, population, cell, etc. According to omics, it is divided into proteome, genome, transcriptome, structure analysis, population genetics, and so on. |
| https://www.agbiodata.org/ Accessed on 4 May 2022. | Integrated platform of agricultural biological databases and related resources | It is a consortium of agricultural biological databases that integrate standards and best practices for the acquisition, display, and reuse of genomic, genetic, and breeding data. |
| https://phytozome.jgi.doe.gov/pz/portal.html Accessed on 4 May 2022. | Plant Comparative Genomics Repository | Phytozome database, the Plant Comparative Genomics portal of the Department of Energy’s Joint Genome Institute, provides a hub for accessing, visualizing, and analyzing JGI-sequenced plant genomes, as well as selected genomes and datasets sequenced elsewhere. |
| http://harvest.ucr.edu/ Accessed on 4 May 2022. | Platform for Crop EST sequences and related molecular information | HarvEST database includes various functions, such as microarray content design, SNP identification, genotyping platform design, comparative genomics, and the coupling of physical and genetic profiles. |
| https://www.uniprot.org/ Accessed on 4 May 2022. | Protein sequence and functional information resource database and analysis platform | Uniprot database is the world’s leading resource for high-quality, comprehensive, and freely accessible protein sequence and functional information. |
| http://www.plantgdb.org/ Accessed on 4 May 2022. | Plant Genome Sequence Database | Plantgdb database includes software, visualization, and data access portals that implement novel prediction algorithms, as well as a network infrastructure environment implementation of development tools for distributed computing, protocol sharing, and analysis of source records. |
| https://mpss.danforthcenter.org/index.php Accessed on 4 May 2022. | NGS database, including small RNAs and genome resources for plants | Meyers Lab database focuses on many aspects of plant small RNAs, including their major roles in gene and transposable regulation, but also their biogenesis and evolution. Includes small RNA sequencing, cut target RNA sequencing, and a variety of informatics tools. |
| http://metacrop.ipk-gatersleben.de Accessed on 4 May 2022. | Crop Metabolism Pathway Database | Metacrop database summarizes various information about metabolic pathways in crops and allows the automatic export of information to create detailed metabolic models. |
| URL | Note |
|---|---|
| http://www.arabidopsis.org Accessed on 4 May 2022. | The most commonly used repository of Arabidopsis genetic and molecular biology data |
| http://rarge.gsc.riken.jp/ Accessed on 4 May 2022. | Arabidopsis cDNA, mutant, and microarray database |
| http://www.athamap.de/ Accessed on 4 May 2022. | A genome-wide database of putative transcription factor binding sites in Arabidopsis |
| http://www.plprot.ethz.ch/ Accessed on 4 May 2022. | Arabidopsis plastid protein database |
| http://seedgenes.org/ Accessed on 4 May 2022. | Database of key Arabidopsis developmental genes |
| http://suba.live/ Accessed on 4 May 2022. | Subcellular localization database for Arabidopsis proteins |
| http://atrm.cbi.pku.edu.cn/ Accessed on 4 May 2022. | Arabidopsis transcriptional regulatory mapping database |
| http://wanglab.sippe.ac.cn/rootatlas/ Accessed on 4 May 2022. | Arabidopsis root single-cell RNA-seq database |
| http://ipf.sustech.edu.cn/pub/athrna/ Accessed on 4 May 2022. | Arabidopsis RNA-seq data resources |
| http://signal.salk.edu/ Accessed on 4 May 2022. | A database showing all T-DNA insertions and methyl group data |
| URL | Note |
|---|---|
| http://www.ricedata.cn/index.htm Accessed on 4 May 2022. | National Rice Data Center |
| http://signal.salk.edu/cgi-bin/RiceGE Accessed on 4 May 2022. | Rice functional genome expression database |
| https://shigen.nig.ac.jp/rice/oryzabase/ Accessed on 4 May 2022. | Rice genetics and genomics database |
| http://www.wheatgenome.org/ Accessed on 4 May 2022. | Wheat genome information database |
| http://earth.nig.ac.jp/~dclust/cgi-bin/index.cgi Accessed on 4 May 2022. | Barley germplasm resources and genome analysis database |
| http://maize.jcvi.org/ Accessed on 4 May 2022. | Maize genome database |
| https://www.maizegdb.org/ Accessed on 4 May 2022. | Maize genome and genetic analysis platform |
| https://soybase.org/ Accessed on 4 May 2022. | Soybean genomics and molecular biology database |
| http://www.ildis.org/LegumeWeb/ Accessed on 4 May 2022. | International legume database and information service |
| https://www.cottongen.org/ Accessed on 4 May 2022. | Cotton genomics, genetics, and breeding database |
| http://ted.bti.cornell.edu/ Accessed on 4 May 2022. | Tomato functional genome database |
| http://ted.bti.cornell.edu/epigenome/ Accessed on 4 May 2022. | Tomato epigenome database |
| http://tea.solgenomics.net/ Accessed on 4 May 2022. | High-resolution mapping and search tool for tomato genes and their products |
| http://tomexpress.toulouse.inra.fr/ Accessed on 4 May 2022. | Tomato transcriptome data visualization and analysis platform |
| https://solgenomics.net/ Accessed on 4 May 2022. | Genome sequencing database of Solanaceae species |
| http://gabipd.org/projects/Pomamo/ Accessed on 4 May 2022. | Potato bioinformatics database |
| Database Name | URL | Note |
|---|---|---|
| Tbtools | https://github.com/srbehera11/stag-cns Accessed on 4 May 2022. | An integrated toolkit for interactive analysis of big biological data |
| SMART | http://smart.embl-heidelberg.de/ Accessed on 4 May 2022. | Protein conserved domain prediction tool |
| STAG-CNS | https://github.com/srbehera11/stag-cns Accessed on 4 May 2022. | A sequentially conserved non-coding sequence discovery tool for an arbitrary number of species |
| FED | http://www.hi-tom.net/FED Accessed on 4 May 2022. | Genome editing exogenous component detection platform |
| MAFFT | https://mafft.cbrc.jp/alignment/server/ Accessed on 4 May 2022. | Online sequence matching tool |
| Protter | http://wlab.ethz.ch/protter/start/ Accessed on 4 May 2022. | Online protein structure mapping tool |
| EvolView | https://www.evolgenius.info/evolview/ Accessed on 4 May 2022. | Web-based tools for visualizing, annotating, and managing system trees |
| iTOL | https://itol.embl.de/ Accessed on 4 May 2022. | Online tool for displaying, annotating, and managing system development trees |
| Database Name | URL | NOTE |
|---|---|---|
| BAR | http://www.bar.utoronto.ca/welcome.htm Accessed on 4 May 2022. | Plant biology analysis tools platform |
| CRISPR-P | http://crispr.hzau.edu.cn/CRISPR2/ Accessed on 4 May 2022. | Improved CRISPR/Cas9 toolkit for plant genome editing |
| ACT | https://www.michalopoulos.net/act/ Accessed on 4 May 2022. | Arabidopsis co-expression analysis tool |
| OryGenesDB | http://orygenesdb.cirad.fr/ Accessed on 4 May 2022. | An interactive tool for reverse genetics studies in rice |
| T-DNA Express | http://signal.salk.edu/cgi-bin/tdnaexpress Accessed on 4 May 2022. | Arabidopsis gene targeting tool |
| Plant MetGenMAP | http://bioinfo.bti.cornell.edu/cgi-bin/MetGenMAP/home.cgi Accessed on 4 May 2022. | Web-based tools for comprehensive mining and integration of gene expression and metabolite changes in the context of biochemical pathways |
| iTAK | http://itak.feilab.net/cgi-bin/itak/index.cgi Accessed on 4 May 2022. | Software packages for identifying and classifying plant transcription factors and protein kinases |
| PlantPAN | http://plantpan2.itps.ncku.edu.tw/ Accessed on 4 May 2022. | Tools for detecting transcription factor binding sites in plants |
| SnpHub | http://guoweilong.github.io/SnpHub/ Accessed on 4 May 2022. | A unified web server framework for exploring large-scale genomic variation data |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, H.; Wang, B.; Yuan, F. Bioinformatics in Plant Breeding and Research on Disease Resistance. Plants 2022, 11, 3118. https://doi.org/10.3390/plants11223118
Mu H, Wang B, Yuan F. Bioinformatics in Plant Breeding and Research on Disease Resistance. Plants. 2022; 11(22):3118. https://doi.org/10.3390/plants11223118
Chicago/Turabian StyleMu, Huiying, Baoshan Wang, and Fang Yuan. 2022. "Bioinformatics in Plant Breeding and Research on Disease Resistance" Plants 11, no. 22: 3118. https://doi.org/10.3390/plants11223118
APA StyleMu, H., Wang, B., & Yuan, F. (2022). Bioinformatics in Plant Breeding and Research on Disease Resistance. Plants, 11(22), 3118. https://doi.org/10.3390/plants11223118