
Phytomanagement of a Lead-Polluted Shooting Range Using an Aromatic Plant Species and Its Effects on the Rhizosphere Bacterial Diversity and Essential Oil Production
 , ,
, ,  and
and
Abstract
1. Introduction
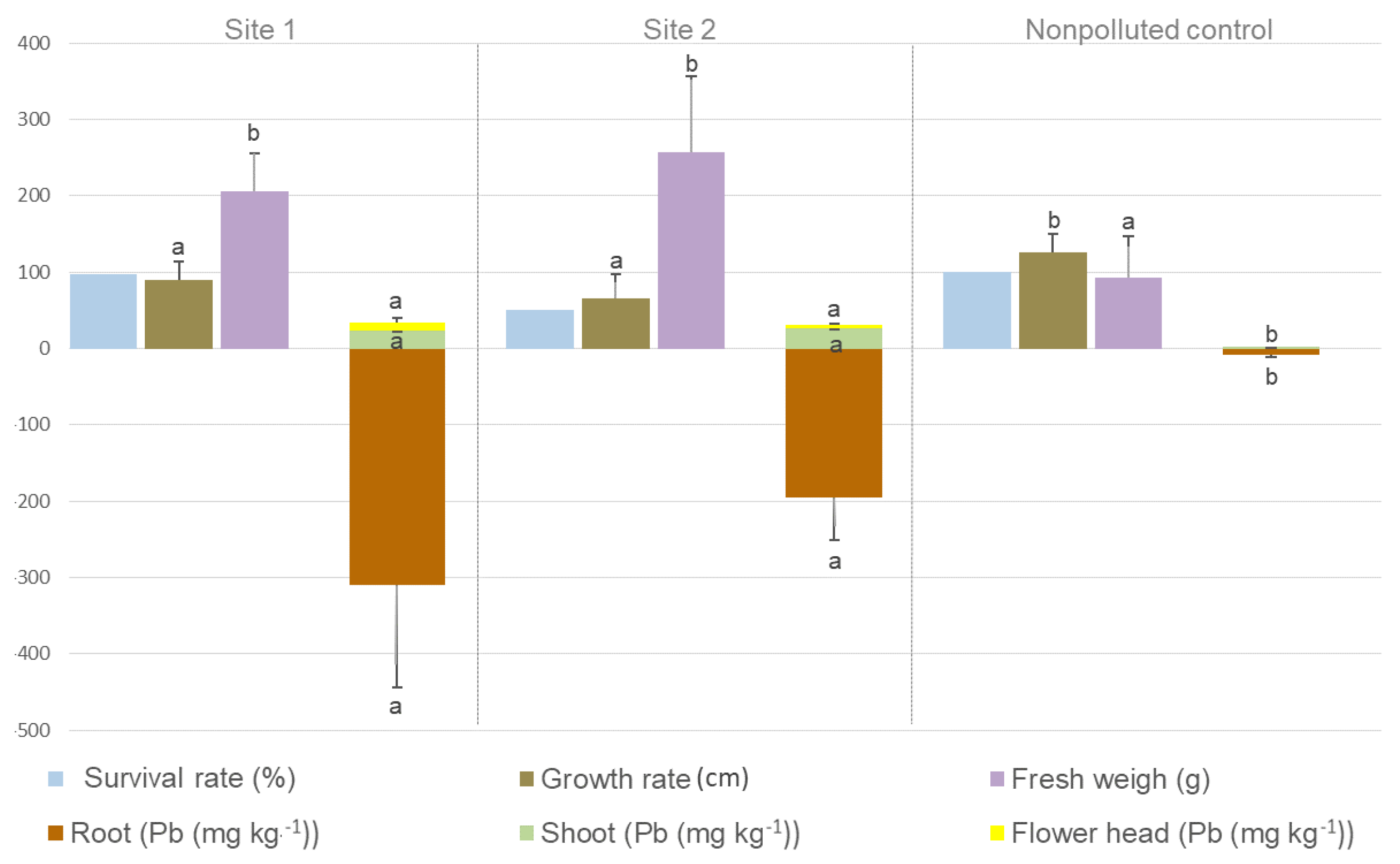
2. Results
3. Discussion
4. Materials and Methods
4.1. Soil Sampling and Site Characterization
4.2. Trial Design
4.3. Plant Survival, Biomass, and Trace Element Accumulation
4.4. H. petiolaris Rhizosphere Bacterial Community Analyses
4.5. Essential Oil Extraction, GC–MS Analysis, and Pb Concentration
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmad, M.; Lee, S.S.; Yang, J.E.; Ro, H.M.; Lee, Y.H.; Ok, Y.S. Effects of soil dilution and amendments (mussel shell, cow bone, and biochar) on Pb availability and phytotoxicity in military shooting range soil. Ecotoxicol. Environ. Saf. 2012, 79, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.Q.; Hardison, D.W.; Harris, W.G.; Cao, X.; Zhou, Q. Effects of soil property and soil amendment on weathering of abraded metallic Pb in shooting ranges. Water Air Soil Pollut. 2007, 178, 297–307. [Google Scholar] [CrossRef]
- Sorvari, J.; Antikainen, R.; Pyy, O. Environmental contamination at finish shooting ranges-the scope of the problem and management options. Sci. Total Environ. 2006, 366, 21–31. [Google Scholar] [CrossRef]
- Ahmad, M.; Lee, S.S.; Moon, D.H.; Yang, J.E.; Ok, Y.S. A review of environmental contamination and remediation strategies for heavy metals at shooting range soils. In Environmental Protection Strategies for Sustainable Development; Springer: Berlin/Heidelberg, Germany, 2012; pp. 437–452. [Google Scholar] [CrossRef]
- USEPA (United States Environmental Protection Agency). Soil Screening Guidance: User’s Guidance; Office of Solid Waste and Emergency Response, USEPA: Washington, DC, USA, 1996. Available online: https://semspub.epa.gov/work/HQ/211.pdf (accessed on 17 April 2013).
- Cao, X.; Wahbi, A.; Ma, L.; Li, B.; Yang, Y. Immobilization of Zn, Cu, and Pb in contaminated soils using phosphate rock and phosphoric acid. J. Hazard. Mater. 2009, 164, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Spuller, C.; Weigand, H.; Marb, C. Trace metal stabilization in a shooting range soil: Mobility and phytotoxicity. J. Hazard. Mater. 2007, 141, 378–387. [Google Scholar] [CrossRef]
- Roman-Ponce, B.; Ramos-Garza, J.; Vasquez-Murrieta, M.S.; Rivera-Orduna, F.N.; Chen, W.F.; Yan, J.; Estrada-de Los Santos, P.; Wang, E.T. Cultivable endophytic bacteria from heavy metal(loid)-tolerant plants. Arch. Microbiol. 2016, 198, 941–956. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Lead dietary exposure in the European population. EFSA J. 2012, 10, 2831. [Google Scholar] [CrossRef]
- Robinson, B.H.; Bischofberger, S.; Stoll, A.; Schroer, D.; Furrer, G.; Roulier, S.; Gruenwald, A.; Attinger, W.; Schulin, R. Plant uptake of trace elements on a Swiss military shooting range: Uptake pathways and land management implications. Environ. Pollut. 2008, 153, 668–676. [Google Scholar] [CrossRef]
- Kumar, P.; Gupta, S.B.; Anurag Soni, R. Bioremediation of Cadmium by Mixed Indigenous Isolates Serratia liquefaciens BSWC3 and Klebsiella pneumoniae RpSWC3 Isolated from Industrial and Mining Affected Water Samples. Pollution 2019, 5, 351–360. [Google Scholar] [CrossRef]
- Schupp, T.; Damm, G.; Foth, H.; Freyberger, A.; Gebel, T.; Gundert-Remy, U.; Hengstler, J.G.; Mangerich, A.; Partosch, F.; Röhl, C. Long-term simulation of lead concentrations in agricultural soils in relation to human adverse health effects. Arch. Toxicol. 2020, 94, 2319–2329. [Google Scholar] [CrossRef]
- Pandey, V.C.; Rai, A.; Korstad, J. Aromatic Crops in Phytoremediation: From Contaminated to Waste Dumpsites. In Phytomanag of Poll Sites, 1st ed.; Pandey, V.C., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 9, pp. 255–275. [Google Scholar]
- Gomes, H.I. Phytoremediation for bioenergy: Challenges and opportunities. Environ. Technol. 2012, 1, 59–66. [Google Scholar] [CrossRef]
- Zago, V.C.P.; Dores, N.C.; Watts, B.A. Strategy for phytomanagement in an area affected by iron ore dam rupture: A study case in Minas Gerais State, Brazil. Environ. Pollut. 2019, 249, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Jeannin, T.; Yung, L.; Evon, P.; Labonne, L.; Ouagne, P.; Lecourt, M.; Cazaux, D.; Chalot, M.; Placet, V. Native stinging nettle (Urtica dioica L.) growing spontaneously under short rotation coppice for phytomanagement of trace element contaminated soils: Fibre yield, processability and quality. Ind. Crops Prod. 2020, 145, 111997. [Google Scholar] [CrossRef]
- Perlein, A.; Bert, V.; Fernandes de Souza, M.; Gaucher, R.; Papin, A.; Geuens, J.; Wens, A.; Meers, E. Phytomanagement of a Trace Element-Contaminated Site to Produce a Natural Dye: First Screening of an Emerging Biomass Valorization Chain. Appl. Sci. 2021, 11, 10613. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Bert, V.; Perlein, A.; Tisserant, B.; Ferrant, P.; Lounès-Hadj Sahraoui, A. In situ cultivation of aromatic plant species for the phytomanagement of an aged-trace element polluted soil: Plant biomass improvement options and techno-economic assessment of the essential oil production. Sci. Total Environ. 2021, 789, 147944. [Google Scholar] [CrossRef]
- Thijs, S.; Langill, T.; Vangronsveld, J. The bacterial and fungal microbiota of hyperaccumulator plants: Small organisms, large influence. Adv. Bot. Res. 2017, 83, 43–86. [Google Scholar] [CrossRef]
- Bravin, M.N.; Garnier, C.; Lenoble, V.; Gérard, F.; Dudal, Y.; Hinsinger, P. Root-induced changes in pH and dissolved organic matter binding capacity affect copper dynamic speciation in the rhizosphere. Geochim. Cosmochim. Acta 2012, 84, 256–268. [Google Scholar] [CrossRef]
- Płociniczak, T.; Sinkkonen, A.; Romantschuk, M.; Sułowicz, S.; Piotrowska-Seget, Z. Rhizospheric Bacterial Strain Brevibacterium casei MH8a Colonizes Plant Tissues and Enhances Cd, Zn, Cu Phytoextraction by White Mustard. Front. Plant Sci. 2016, 7, 101–122. [Google Scholar] [CrossRef]
- Poverene, M.; Carrera, A.; Ureta, S.; Cantamutto, M. Wild Helianthus species and wild-sunflower hybridization in Argentina. Helia 2004, 27, 133–142. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Raymond, O.; Rosenthal, D.M.; Lai, Z.; Livingstone, K.; Nakazato, T.; Durphy, J.L.; Schwarzbach, A.E.; Donovan, L.A.; Lexer, C. Major ecological transitions in wild sunflowers facilitated by hybridization. Science 2003, 301, 1211–1216. [Google Scholar] [CrossRef]
- Saran, A.; Fernandez, L.; Cora, F.; Savio, M.; Thijs, S.; Vangronsveld, J.; Merini, L.J. Phytostabilization of Pb and Cd polluted soils using Helianthus petiolaris as pioneer aromatic plant species. Int. J. Phytoremediation 2020, 11, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Saran, A.; Fernandez, L.; Minig, M.; Bellozas Reinhard, M.; Merini, L.J. Repellent activity of essential oils from native plants and their blend for Tribolium castaneum control in store grains. Semiárida 2019, 29, 4351. [Google Scholar] [CrossRef]
- Oorts, K.; Smolders, E.; Lanno, R.; Chowdhury, M.J. Bioavailability and Ecotoxicity of Lead in Soil: Implications for Setting Ecological Soil Quality Standards. Environ. Toxicol. Chem. 2021, 40, 1948–1961. [Google Scholar] [CrossRef]
- Antoniadis, V.; Shaheen, S.M.; Levizou, E.; Shahid, M.; Niazi, N.K.; Vithanage, M.; Ok, Y.S.; Bolan, N.; Rinklebe, J. A critical prospective analysis of the potential toxicity of trace element regulation limits in soils worldwide: Are they protective concerning health risk assessment?—A review. Environ. Int. 2019, 127, 819–847. [Google Scholar] [CrossRef]
- Mariussen, E.; Vaa Johnsen, I.; Stromseng, A.E. Distribution and mobility of lead (Pb), copper (Cu), zinc (Zn), and antimony (Sb) from ammunition residues on shooting ranges for small arms located on mires. Environ. Sci. Pollut. Res. 2017, 24, 10182–10196. [Google Scholar] [CrossRef] [PubMed]
- Ghazaryan, K.; Movsesyan, H.; Ghazaryan, N.; Watts, B.A. Copper phytoremediation potential of wild plant species growing in the mine polluted areas of Armenia. Environ. Pollut. 2019, 249, 491–501. [Google Scholar] [CrossRef]
- Kiran, B.R.; Prasad, M.N.V.; Sateesh, S. Ricinus communis L. (castor bean) as a potential candidate for revegetating industrial waste contaminated sites in peri-urban greater Hyderabad: Remarks on seed oil. Environ. Sci. Pollut. 2017, 24, 19955–19964. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Chen, Y.; Yang, X.; Cui, Z. Optimization of combined phytoremediation for heavy metal contaminated mine tailings by a field-scale orthogonal experiment. Ecotoxicol. Environ. Saf. 2019, 168, 1–8. [Google Scholar] [CrossRef]
- Bickford, W.A.; Zak, D.R.; Kowalski, K.P.; Goldberg, D.E. Differences in rhizosphere microbial communities between native and non-native Phragmites australis may depend on stand density. Ecol. Evol. 2020, 10, 11739–11751. [Google Scholar] [CrossRef]
- Galitskaya, P.; Biktasheva, L.; Blagodatsky, S.; Selivanovskaya, S. Response of bacterial and fungal communities to high petroleum pollution in different soils. Sci. Rep. 2021, 11, 164. [Google Scholar] [CrossRef]
- Nwagwu, E.C.; Yilwa, V.M.; Egbe, N.E.; Onwumere, B.G. Isolation and characterization of heavy metal tolerant bacteria from Panteka stream, Kaduna, Nigeria and their potential for bioremediation. Afr. J. Biotechnol. 2017, 16, 32–40. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The Multifarious PGPR Serratia marcescens CDP-13 Augments Induced Systemic Resistance and Enhanced Salinity Tolerance of Wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef] [PubMed]
- Rotkittikhun, P.; Kruatrachue, M.; Pokethitiyook, P.; Baker, A.J. Tolerance and accumulation of lead in Vetiveria zizanioides and its effect on oil production. J. Environ. Biol. 2010, 31, 329. [Google Scholar]
- Khajanchi, L.; Yadava, R.K.; Kaurb, R.; Bundelaa, D.S.; Khana, M.I.; Chaudharya, M.; Meenaa, R.L.; Dara, S.R.; Singha, G. Productivity, essential oil yield, and heavy metal accumulation in lemon grass (Cymbopogon flexuosus) under varied wastewater-groundwater irrigation regimes. Ind. Crops Prod. 2013, 45, 270–278. [Google Scholar] [CrossRef]
- Chrastný, V.; Komárek, M.; Jrovcová, E.; Štíchová, J. A critical evaluation of the 0.05 M EDTA extraction of Pb from forest soils. Int. J. Environ. Anal. Chem. 2008, 88, 385–396. [Google Scholar] [CrossRef]
- Yang, S.X.; Liao, B.; Li, J.T.; Guo, T.; Shu, W.S. Acidification, heavy metal mobility and nutrient accumulation in the soil-plant system of a revegetated acid mine wasteland. Chemosphere 2010, 80, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Loponte, D.; Corriale, M.J. Patterns of Resource Use and Isotopic Niche Overlap Among Guanaco (Lamaguanicoe), Pampas Deer (Ozotoceros bezoarticus) and Marsh Deer (Blastocerusdichotomus) in the Pampas. Ecological, Paleoenvironmental and Archaeological Implications. Environ. Archaeol. 2019, 25, 411–444. [Google Scholar] [CrossRef]
- Method 3050B; Acid Digestion of Sediments, Sludges, and Soils, Revision 2. USEPA (United States Environmental Protection Agency): Washington, DC, USA, 1996.
- Rafati, M.; Khorasani, N.; Moattar, F.; Shirvany, A.; Moraghebi, F.; Hosseinzadeh, S. Phytoremediation potential of Populus alba and Morus alba for cadmium, chromuim and nickel absorption from polluted soil. Int. J. Environ. Res. 2011, 5, 961–970. [Google Scholar] [CrossRef]
- Padmavathiamma, P.K.; Li, L.Y. Phytoremediation technology: Hyperaccumulation metals in plants. Water Air Soil Pollut. 2007, 184, 105–126. [Google Scholar] [CrossRef]
- Bentley, D.R.; Balasubramanian, S.; Swerdlow, H.P.; Smith, G.P.; Milton, J.; Brown, C.G.; Hall, K.P.; Evers, D.J.; Barnes, C.L.; Bignell, H.R.; et al. Accurate whole human genome sequencing using reversible terminator chemistry. Nature 2008, 456, 53–59. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson AJ, A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web788 based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Organic Matter (%) | Texture | EC (mS/m) | pH | Total Pb | Ammonium Acetate EDTA Extractable Upper a Pb | Ammonium Acetate EDTA Extractable Bottom b Pb | Ammonium Acetate EDTA Extractable Pb after Harvest | |
|---|---|---|---|---|---|---|---|---|
| Site 1 | 1.8 | Loamy sand | 271 ± 5 a | 7.7 ± 0.7 a | 427.8 ± 135.5 a | 15.1 ± 5.1 b | 7.3 ± 4.0 b | 10.4 ± 6.8 a |
| Site 2 | 0.5 | Sandy | 18 ± 6 b | 7.4 ± 0.3 a | 416.5 ± 102.1 a | 31.1 ± 8.3 a | 23.6 ± 9.9 a | 24.2 ± 13.5 a |
| Nonpolluted | 1.9 | Loamy sand | 200 ± 4 a | 7.4 ± 0.5 a | 20.5 ± 3.4 b | 3.9 ± 2.1 c | 4.3 ± 1.4 b | 2.8 ± 6.8 b |
| BAF | TF | |||
|---|---|---|---|---|
| Roots | Shoots | Flower | ||
| Site 1 | 0.72 ± 0.35 a | 0.05 ± 0.03 a | 0.02 ± 0.00 a | 0.27 ± 0.63 a |
| Site 2 | 0.47 ± 0.12 a | 0.06 ± 0.05 a | 0.01 ± 0.00 a | 0.23 ± 0.19 a |
| Nonpolluted | 0.42 ± 0.22 a | 0.09 ± 0.03 a | - | 0.21 ± 0.18 b |
| Rt (min) | Compounds | CAS | Relative Area | |
|---|---|---|---|---|
| Nonpolluted | Site 1 | |||
| 7.84 | α-Pinene | 7785-70-8 | 66 | 67 |
| 8.25 | Camphene | 79-92-5 | 8 | 8 |
| 18.53 | Acetic acid,1,7,7-trimethyl-bicyclo[2,1,1]hept-2-yl ester | 92618-89-8 | 10 | 10 |
| 23.87 | D-germacrene | 23986-74-5 | 16 | 15 |
| 9.04 | Thujene | 3387-41-5 | <1 | <1 |
| 9.12 | β-Pinene | 127-91-3 | <1 | <1 |
| 10.77 | Limoneno | 5989-54-8 | <1 | <1 |
| 12.92 | α-Pinene epoxide | 1686-14-2 | <1 | <1 |
| 21.45 | β-Cubene | 13744-15-5 | <1 | <1 |
| 22.24 | Caryophyllene | 87-44-5 | <1 | <1 |
| 22.63 | α-Bergamotene | 17699-05-7 | <1 | <1 |
| 23.14 | Humulene | 6753-98-6 | <1 | <1 |
| 26.41 | Caryophyllene oxide | 1139-30-6 | <1 | <1 |
| Oil yield | 0.14 | 0.014 | ||
| mg Pb/L floral water | <dL | <dL | ||
| mg Pb/L essential oil | <dL | <dL | ||
| mg Pb/Kg plant waste | 2.18 ± 0.20 | 23.57 ± 0.7 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saran, A.; Fernandez, L.; Latini, C.Y.; Reinhard, M.B.; Minig, M.; Thijs, S.; Vangronsveld, J.; Merini, L.J. Phytomanagement of a Lead-Polluted Shooting Range Using an Aromatic Plant Species and Its Effects on the Rhizosphere Bacterial Diversity and Essential Oil Production. Plants 2022, 11, 3024. https://doi.org/10.3390/plants11223024
Saran A, Fernandez L, Latini CY, Reinhard MB, Minig M, Thijs S, Vangronsveld J, Merini LJ. Phytomanagement of a Lead-Polluted Shooting Range Using an Aromatic Plant Species and Its Effects on the Rhizosphere Bacterial Diversity and Essential Oil Production. Plants. 2022; 11(22):3024. https://doi.org/10.3390/plants11223024
Chicago/Turabian StyleSaran, Anabel, Lucia Fernandez, Cinthia Yanela Latini, Monica Bellozas Reinhard, Marisol Minig, Sofie Thijs, Jaco Vangronsveld, and Luciano Jose Merini. 2022. "Phytomanagement of a Lead-Polluted Shooting Range Using an Aromatic Plant Species and Its Effects on the Rhizosphere Bacterial Diversity and Essential Oil Production" Plants 11, no. 22: 3024. https://doi.org/10.3390/plants11223024
APA StyleSaran, A., Fernandez, L., Latini, C. Y., Reinhard, M. B., Minig, M., Thijs, S., Vangronsveld, J., & Merini, L. J. (2022). Phytomanagement of a Lead-Polluted Shooting Range Using an Aromatic Plant Species and Its Effects on the Rhizosphere Bacterial Diversity and Essential Oil Production. Plants, 11(22), 3024. https://doi.org/10.3390/plants11223024