
Synergistic Effects of Trichoderma harzianum, 1,3 Dichloropropene and Organic Matter in Controlling the Root-Knot Nematode Meloidogyne incognita on Tomato

 ,
,  ,
,  , , , and
, , , and
Abstract
1. Introduction
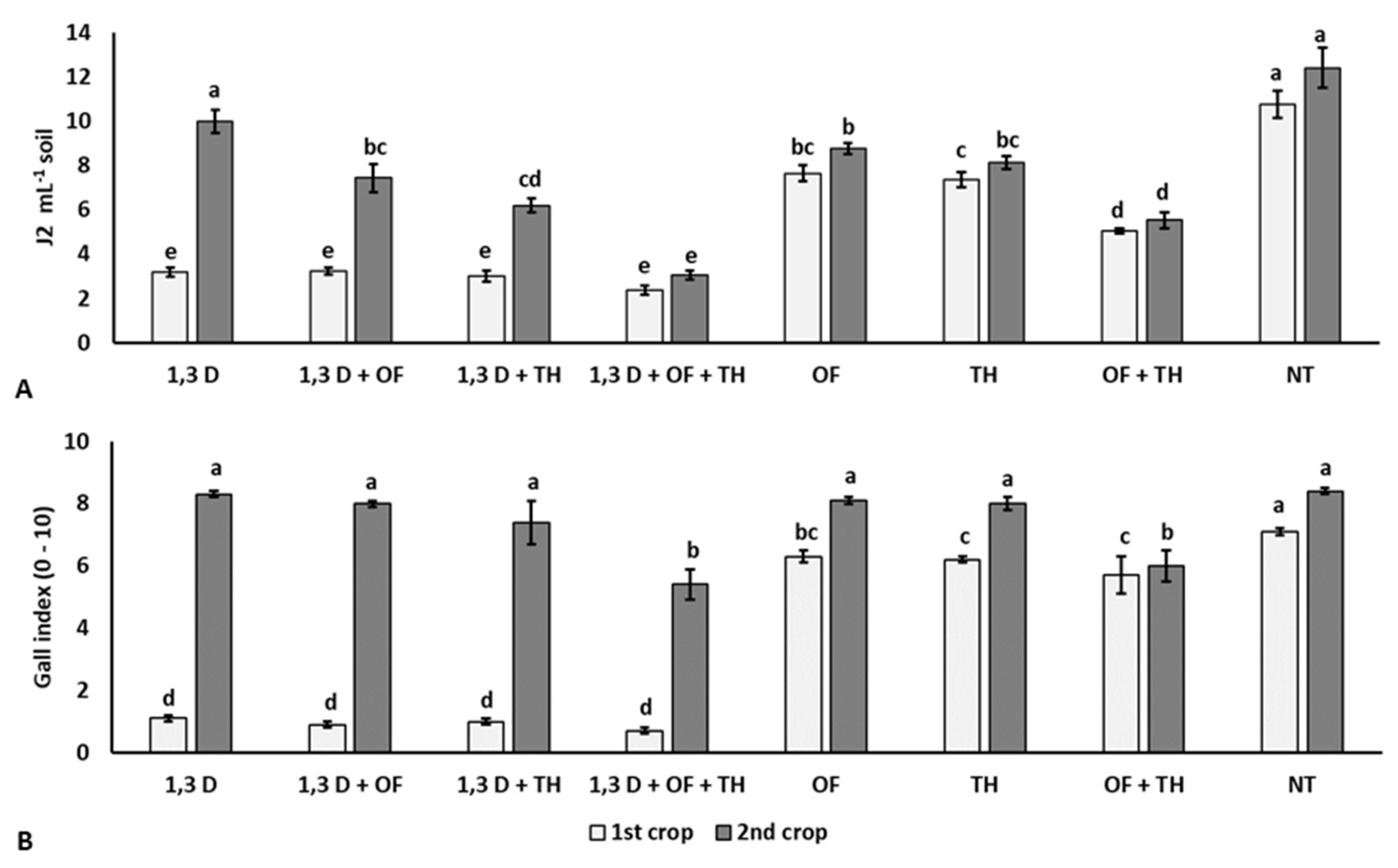
2. Results
2.1. Effects on Nematode Infestation
2.2. Effects on Tomato Yield
2.3. Effects on Tomato Plant Growth
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jones, J.T.; Haegeman, A.; Danchin, E.G.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, C.J.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Massin-Barbet, M.; Salles, J.M.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, B.; Singh, A.P. Nematodes: A threat to sustainability of agriculture. Procedia Environ. Sci. 2015, 29, 215–216. [Google Scholar] [CrossRef]
- Siddiqui, S.; Grundler, F.M. Parasitic nematodes manipulate plant development to establish feeding sites. Curr. Opin. Microbiol. 2018, 46, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Palomares-Rius, J.E.; Escobar, C.; Cabrera, J.; Vovlas, A.; Castillo, P. Anatomical alterations in plant tissues induced by plant-parasitic nematodes. Front. Plant Sci. 2017, 8, 1987. [Google Scholar] [CrossRef]
- Ragozzino, A.; d’Errico, G. Interactions between nematodes and fungi: A concise review. Redia 2011, 94, 123–125. [Google Scholar]
- Regulation (EC) of the European Parliament and of the Council of 21 October 2009 concerning the placing of plant protection products on the market and repealing Council Directives 79/117/EEC and 91/414/EEC No. 1107/2009. OJ 2009, L309, 1–50.
- Regulation (EC) of 25 May 2011 implementing Regulation (EC) No1107/2009 of the European Parliament and of the Council as regards the list of approved active substances. No. 540/2011. OJ 2011, L153, 1–186.
- Nnamdi, C.; Grey, T.L.; Hajihassani, A. Root-knot nematode management for pepper and squash rotations using plasticulture systems with fumigants and non-fumigant nematicides. Crop Prot. 2022, 152, 105844. [Google Scholar] [CrossRef]
- Minuto, A.; Gullino, M.L.; Lamberti, F.; D’Addabbo, T.; Tescari, E.; Ajwa, H.; Garibaldi, A. Application of an emulsifiable mixture of 1,3-dichloropropene and chloropicrin against root knot nematodes and soilborne fungi for greenhouse tomatoes in Italy. Crop Prot. 2006, 25, 1244–1252. [Google Scholar] [CrossRef]
- Desaeger, J.; Csinos, A.; Timper, P.; Hammes, G.; Seebold, K. Soil fumigation and oxamyl drip applications for nematode and insect control in vegetable plasticulture. Ann. Appl. Biol. 2004, 145, 59–70. [Google Scholar] [CrossRef]
- Sharon, E.; Chet, I.; Spiegel, Y. Trichoderma as a biological control agent. In Biological Control of Plant-Parasitic Nematodes: Building Coherence Between Microbial Ecology and Molecular Mechanisms, Progress in Biological Control; Davies, K., Spiegel, Y., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 183–201. [Google Scholar]
- d’Errico, G.; Mormile, P.; Malinconico, M.; Bolletti Censi, S.; Lanzuise, S.; Crasto, A.; Woo, S.L.; Marra, R.; Lorito, M.; Vinale, F. Trichoderma spp. and a carob (Ceratonia siliqua) galactomannan to control the root-knot nematode Meloidogyne incognita on tomato plants. Can. J. Plant Pathol. 2021, 43, 267–274. [Google Scholar] [CrossRef]
- d’Errico, G.; Marra, R.; Crescenzi, A.; Davino, S.W.; Fanigliulo, A.; Woo, S.L.; Lorito, M. Integrated management strategies of Meloidogyne incognita and Pseudopyrenochaeta lycopersici on tomato using a Bacillus firmus-based product and two synthetic nematicides in two consecutive crop cycles in greenhouse. Crop Prot. 2019, 122, 159–164. [Google Scholar] [CrossRef]
- Gautam, S.B.; Chand, R.; Singh, S.K.; Chandra, S.; Maurya, M.K. Efficacy of Trichoderma viride and T. harzianum culture filtrate on egg hatching and larval mortality of root-knot nematode, Meloidogyne incognita. Ann. Plant Prot. Sci. 2021, 29, 241–245. [Google Scholar] [CrossRef]
- Fan, H.; Yao, H.; Zhao, D.; Zhu, X.; Wang, Y.; Liu, X.; Duan, Y.; Chen, L. Isolation and effect of Trichoderma citrinoviride Snef 1910 for the biological control of root-knot nematode, Meloidogyne incognita. BMC Microbiol. 2020, 20, 299. [Google Scholar] [CrossRef] [PubMed]
- Sahenabani, N.; Hadavi, N. Biological control of root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Soil Biol. Biochem. 2008, 40, 2016–2020. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef]
- Rabiey, M.; Hailey, L.E.; Roy, S.R.; Grenz, K.; Al-Zadjali, M.A.; Barrett, G.A.; Jackson, R.W. Endophytes vs tree pathogens and pests: Can they be used as biological control agents to improve tree health? Eur. J. Plant Pathol. 2019, 155, 711–729. [Google Scholar] [CrossRef]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial effects of endophytic fungi colonization on plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Mikola, J.; Helander, M. Endophytic phyllosphere fungi and nutrient cycling in terrestrial ecosystems. Curr. Sci. 2015, 109, 121–126. [Google Scholar]
- Tóthné Bogdányi, F.; Boziné Pullai, K.; Doshi, P.; Erdős, E.; Gilián, L.D.; Lajos, K.; Leonetti, P.; Nagy, P.I.; Pantaleo, V.; Petrikovszki, R.; et al. Composted municipal green waste infused with biocontrol agents to control plant parasitic nematodes–A review. Microorganisms 2021, 9, 2130. [Google Scholar] [CrossRef]
- Rosskopf, E.; Di Gioia, F.; Hong, J.C.; Pisani, C.; Kokalis-Burelle, N. Organic amendments for pathogen and nematode control. Annu. Rev. Phytopathol. 2020, 58, 277–311. [Google Scholar] [CrossRef]
- Jinu, E.; Park, K.C. Effects of dazomet on soil organisms and recolonisation of fumigated soil. Pedobiologia 2014, 57, 147–154. [Google Scholar]
- Leandro, L.F.S.; Guzman, T.; Ferguson, L.M.; Fernandez, G.E.; Louws, F.J. Population dynamics of Trichoderma in fumigated and compost-amended soil and on strawberry roots. Appl. Soil Ecol. 2007, 35, 237–246. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, D.; Ren, L.; Song, Z.; Li, Q.; Wu, J.; Fang, W.; Huang, B.; Yan, D.; Li, Y.; et al. Bio-activation of soil with beneficial microbes after soil fumigation reduces soil-borne pathogens and increases tomato yield. Environ. Pollut. 2021, 283, 117160. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, D.; Huang, B.; Song, Z.; Ren, L.; Hao, B.; Liu, J.; Zhu, J.; Fang, W.; Yan, D.; et al. Organic fertilizer improves soil fertility and restores the bacterial community after 1,3-dichloropropene fumigation. Sci. Total Environ. 2020, 738, 140345. [Google Scholar] [CrossRef]
- Sharon, E.; Bareyal, M.; Chet, I.; Herreraestrella, A.; Kleifeld, O.; Spiegel, Y. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Phytopathology 2001, 91, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, T.; Tariq-Khan, M.; Aslam, M.N. Bioefficacy of Trichoderma Species Against Javanese Root-Knot Nematode, Meloidogyne javanica, in Green Gram. Gesunde Pflanz. 2021, 73, 265–272. [Google Scholar] [CrossRef]
- Sonkar, S.S.; Bhatt, J.; Meher, J.; Kashyap, P. Bio-efficacy of Trichoderma viride against the root-knot nematode (Meloidogyne incognita) in tomato plant. J. Pharmacogn. Phytochem. 2018, 7, 2010–2014. [Google Scholar]
- Affokpon, A.; Coyne, D.L.; Htay, C.C.; Agbèdè, R.D.; Lawouin, L.; Coosemans, J. Biocontrol potential of native Trichoderma isolates against root-knot nematodes in West African vegetable production systems. Soil Biol. Biochem. 2011, 43, 600–608. [Google Scholar] [CrossRef]
- Zhang, S.; Gan, Y.; Xu, B.; Xue, Y. The parasitic and lethal effects of Trichoderma longibrachiatum against Heterodera avenae. Biol. Control 2014, 72, 1–8. [Google Scholar] [CrossRef]
- Ibrahim, D.S.S.; Elderiny, M.M.; Ansari, R.A.; Rizvi, R.; Sumbul, A.; Mahmood, I. Role of Trichoderma spp. in the Management of Plant-Parasitic Nematodes Infesting Important Crops. In Management of Phytonematodes: Recent Advances and Future Challenges; Ansari, R., Rizvi, R., Mahmood, I., Eds.; Springer: Singapore, 2020; pp. 259–278. [Google Scholar]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef]
- Sharon, E.; Chet, I.; Spiegel, Y. Improved attachment and parasitism of Trichoderma on Meloidogyne javanica in vitro. Eur. J. Plant Pathol. 2009, 123, 291–299. [Google Scholar] [CrossRef]
- Spiegel, Y.; Sharon, E.; Chet, I. Mechanisms and improved biocontrol of the root-knot nematodes by Trichoderma spp. Acta Hortic. 2005, 698, 225–228. [Google Scholar] [CrossRef]
- Zhang, S.; Gan, Y.; Ji, W.; Xu, B.; Hou, B.; Liu, J. Mechanism and characterization of Trichoderma longibrachiatum T6 in suppressing nematodes (Heterodera avenae) in wheat. Front Plant Sci. 2017, 8, 1491. [Google Scholar] [CrossRef]
- Singh, U.B.; Singh, S.; Malviya, D.; Chaurasia, R.; Imrani, M.; Rai, A.; Sharma, A. Harnessing biocontrol potential of Trichoderma harzianum for control of Meloidogyne incognita in tomato. Indian Phytopathol. 2017, 70, 331–335. [Google Scholar] [CrossRef][Green Version]
- Martínez-Medina, A.; Fernandez, I.; Lok, G.B.; Pozo, M.J.; Pieterse, C.M.; Van Wees, S.C. Shifting from priming of salicylic acid-to jasmonic acid-regulated defences by Trichoderma protects tomato against the root knot nematode Meloidogyne incognita. New Phytol. 2017, 213, 1363–1377. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; del Val, E.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere interactions with plants. FEMS Microbiol. Ecol. 2016, 92, fiw036. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Yu, Z.; Lei, L.; Xia, Z.; Shao, L.; Zhang, K.; Li, G. Nematicidal effect of volatiles produced by Trichoderma sp. J. Asia-Pac. Entomol. 2012, 15, 647–650. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational research on Trichoderma: From omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef]
- Kowalska, B.; Smolińska, U.; Szczech, M.; Winciorek, J. Application of organic waste material overgrown with Trichoderma atroviride as a control strategy for Sclerotinia sclerotiorum and Chalara thielavioides in soil. J. Plant Prot. Res. 2017, 57, 205–211. [Google Scholar] [CrossRef][Green Version]
- Amir-Ahmadi, N.; Moosavi, M.R.; Moafpourian, G. Effect of soil texture and its organic content on the efficacy of Trichoderma harzianum (MIAU 145 C) in controlling Meloidogyne javanica and stimulating the growth of kidney beans. Biocontrol Sci. Technol. 2017, 27, 115–127. [Google Scholar] [CrossRef]
- Asghar, W.; Kataoka, R. Effect of co-application of Trichoderma spp. with organic composts on plant growth enhancement, soil enzymes and fungal community in soil. Arch. Microbiol. 2021, 203, 4281–4291. [Google Scholar] [CrossRef]
- Joos, L.; Herren, G.L.; Couvreur, M.; Binnemans, I.; Oni, F.E.; Höfte, M.; Debode, J.; Bert, W.; Steel, H. Compost is a carrier medium for Trichoderma harzianum. BioControl 2020, 65, 737–749. [Google Scholar] [CrossRef]
- Chavarria-Carvajal, J.A.; Rodriguez-Kabana, R.; Klopper, J.W.; Morgan-Jones, G. Changes in populations of microorganism associated with organic amendments and benzaldeyde to control plant-parasitic nematodes. Nematology 2001, 31, 165–180. [Google Scholar]
- Bird, G. Nematodes and soil ecology. In Michigan Field Crop Ecology; Bull. E-2704; Michigan State University Extension: East Lansing, MI, USA, 2000; pp. 84–94. [Google Scholar]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.; Freckman, D.W.; Georgieva, S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Zeck, W.M. A rating scheme for field evaluation of root-knot infestations. Pflanzenschutz-Nachr. 1971, 24, 141–144. [Google Scholar]
- van Bezooijen, J. Methods and Techniques for Nematology; Wageningen University: Wageningen, The Netherlands, 2006; pp. 1–112. [Google Scholar]
- Deacon, S.; Alix, A.; Knowles, S.; Wheeler, J.; Tescari, E.; Alvarez, L.; Nicolette, J.; Rockel, M.; Burston, P.; Quadri, G. Integrating ecosystem services into crop protection and pest management: Case study with the soil fumigant 1,3-dichloropropene and its use in tomato production in Italy. Integr. Environ. Assess. Manag. 2016, 12, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Small, G.; Miles, M.; Barber, I.; Tsakonas, P.; Bucchi, R. The soil ecotoxicology of 1,3-dichloropropene under commercial growing conditions. Commun. Agric. Appl. Biol. Sci. 2008, 73, 777–785. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Operation | 1st Crop | 2nd Crop |
|---|---|---|
| Soil rotavation and plot subdivision | 6 July 2020 | 10 March 2021 |
| Soil sampling | 7 July 2020 | 13 March 2021 |
| Soil fumigation with 1,3-D | 17 July 2020 | - |
| Application of the organic fertilizer | 13 August 2020 | 13 March 2021 |
| Application of T. harzianum | 18 August 2020 | 18 March 2021 |
| Tomato transplant | 20 August 2020 | 20 March 2021 |
| Application of T. harzianum | 4 September 2020 | 19 May 2021 |
| Application of T. harzianum | 19 September 2020 | 18 May 2021 |
| Tomato harvest | 6 November 2020 | 7 June 2021 |
| Tomato harvest | 20 November 2020 | 18 June 2021 |
| Tomato harvest | 5 December 2020 | 30 June 2021 |
| Tomato harvest | 29 December 2020 | 10 July 2021 |
| Tomato harvest | - | 21 July 2021 |
| Recording SPAD, Zack’ s index, plant weight, soil sampling | 29 December 2020 | 21 July 2021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
d’Errico, G.; Greco, N.; Vinale, F.; Marra, R.; Stillittano, V.; Davino, S.W.; Woo, S.L.; D’Addabbo, T. Synergistic Effects of Trichoderma harzianum, 1,3 Dichloropropene and Organic Matter in Controlling the Root-Knot Nematode Meloidogyne incognita on Tomato. Plants 2022, 11, 2890. https://doi.org/10.3390/plants11212890
d’Errico G, Greco N, Vinale F, Marra R, Stillittano V, Davino SW, Woo SL, D’Addabbo T. Synergistic Effects of Trichoderma harzianum, 1,3 Dichloropropene and Organic Matter in Controlling the Root-Knot Nematode Meloidogyne incognita on Tomato. Plants. 2022; 11(21):2890. https://doi.org/10.3390/plants11212890
Chicago/Turabian Styled’Errico, Giada, Nicola Greco, Francesco Vinale, Roberta Marra, Virgilio Stillittano, Salvatore Walter Davino, Sheridan Lois Woo, and Trifone D’Addabbo. 2022. "Synergistic Effects of Trichoderma harzianum, 1,3 Dichloropropene and Organic Matter in Controlling the Root-Knot Nematode Meloidogyne incognita on Tomato" Plants 11, no. 21: 2890. https://doi.org/10.3390/plants11212890
APA Styled’Errico, G., Greco, N., Vinale, F., Marra, R., Stillittano, V., Davino, S. W., Woo, S. L., & D’Addabbo, T. (2022). Synergistic Effects of Trichoderma harzianum, 1,3 Dichloropropene and Organic Matter in Controlling the Root-Knot Nematode Meloidogyne incognita on Tomato. Plants, 11(21), 2890. https://doi.org/10.3390/plants11212890