


Expression and Variation of the Genes Involved in Rhizobium Nodulation in Red Clover

 ,
,  and
and
Abstract
1. Introduction
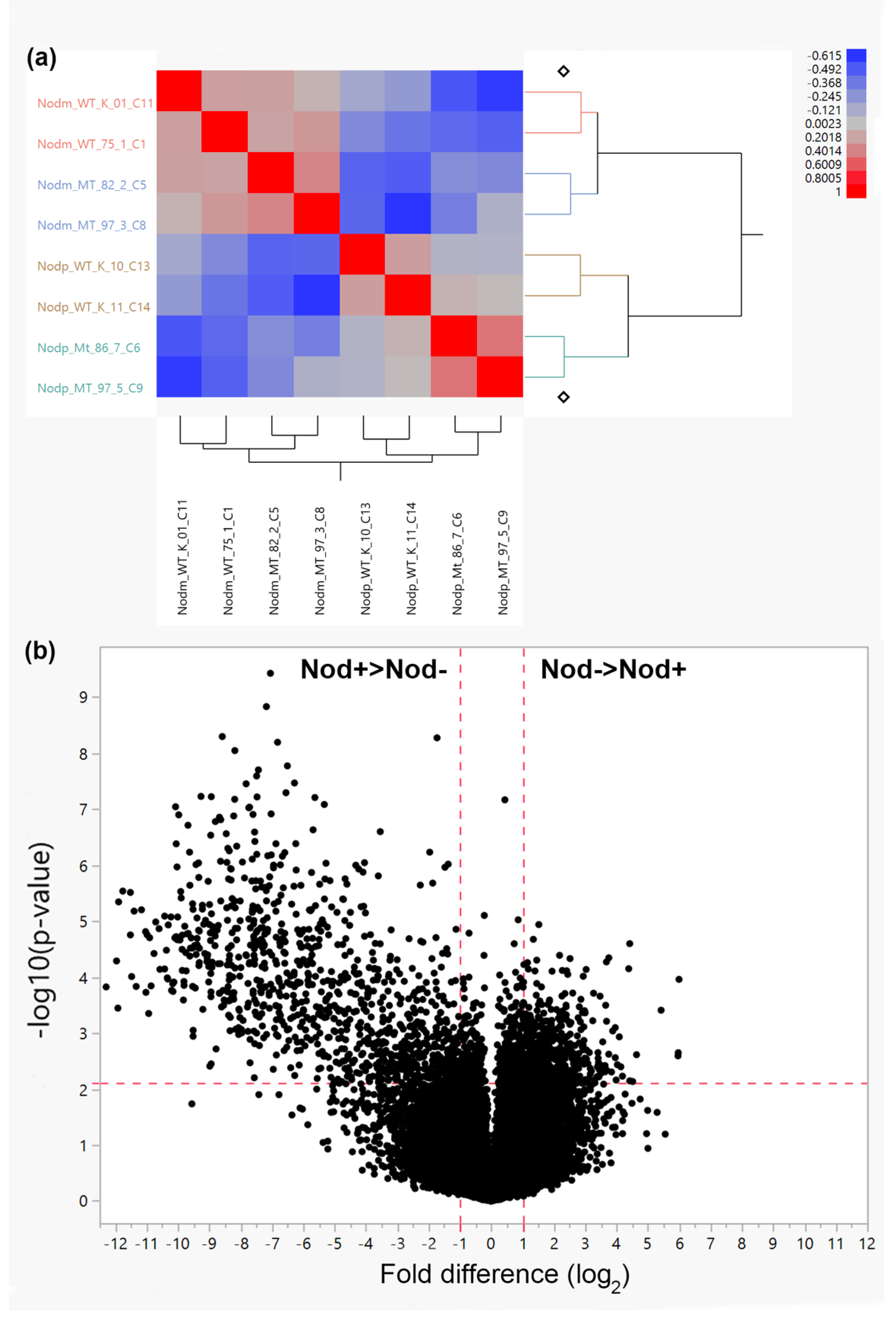
2. Results and Discussion
2.1. Up-Regulated Genes by Nodulation
2.1.1. NOD25 and N22
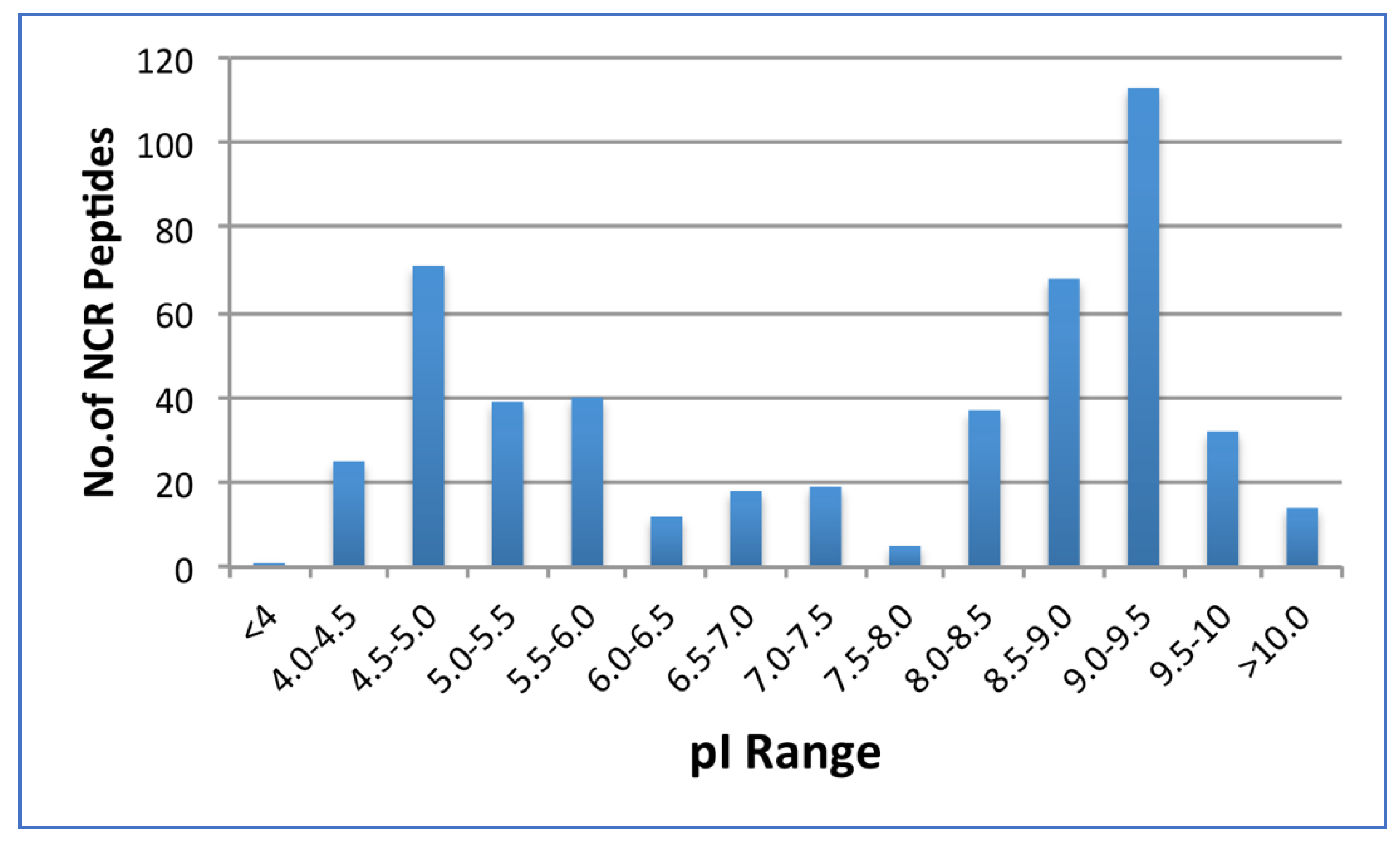
2.1.2. NCRs
2.1.3. ATS3-like/NPDs
2.1.4. NAD1/NIN
2.1.5. DNF1-like and SYP132
2.1.6. Transcription Factor-Encoding Genes
2.2. Down-Regulated Genes by Nodulation
3. Materials and Methods
3.1. Plant Growth
3.2. RNA Extraction and Library Preparation
3.3. Analysis of RNA-Seq Reads
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer Note
References
- Kereszt, A.; Mergaert, P.; Kondorosi, E. Bacteroid development in legume nodules: Evolution of mutual benefit or of sacrificial victims? Mol. Plant-Microbe Interact. 2011, 24, 1300–1309. [Google Scholar] [CrossRef] [PubMed]
- Haag, A.F.; Arnold, M.F.; Myka, K.K.; Kerscher, B.; Dall’Angelo, S.; Zanda, M.; Mergaert, P.; Ferguson, G.P. Molecular insights into bacteroid development during Rhizobium–legume symbiosis. FEMS Microbiol. Rev. 2013, 37, 364–383. [Google Scholar] [CrossRef]
- Oono, R.; Denison, R.F. Comparing Symbiotic Efficiency between Swollen versus Nonswollen Rhizobial Bacteroids. Plant Physiol. 2010, 154, 1541–1548. [Google Scholar] [CrossRef] [PubMed]
- Van de Velde, W.; Zehirov, G.; Szatmari, A.; Debreczeny, M.; Ishihara, H.; Kevei, Z.; Farkas, A.; Mikulass, K.; Nagy, A.; Tiricz, H.; et al. Plant peptides govern terminal differentiation of bacteria in symbiosis. Science 2010, 327, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.C.; Long, F.; Maruoka, E.M.; Cooper, J.B. A new proline-rich early nodulin from Medicago truncatula is highly expressed in nodule meristematic cells. Plant Cell 1994, 6, 1265–1275. [Google Scholar] [CrossRef][Green Version]
- Liu, J.; Miller, S.S.; Graham, M.; Bucciarelli, B.; Catalano, C.M.; Sherrier, D.J.; Samac, D.A.; Ivashuta, S.; Fedorova, M.; Matsumoto, P.; et al. Recruitment of Novel Calcium-Binding Proteins for Root Nodule Symbiosis in Medicago truncatula. Plant Physiol. 2006, 141, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Kevei, Z.; Vinardell, J.M.; Kiss, G.B.; Kondorosi, A.; Kondorosi, E. Glycine-Rich Proteins Encoded by a Nodule-Specific Gene Family Are Implicated in Different Stages of Symbiotic Nodule Development in Medicago spp. Mol. Plant-Microbe Interact. 2002, 15, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, D.I.; Silverstein, K.A.T.; Young, N.D. Genomic Characterization of the LEED..PEEDs, a Gene Family Unique to the Medicago Lineage. G3 Genes|Genomes|Genet. 2014, 4, 2003–2012. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, D.I.; Silverstein, K.A.T.; Young, N.D. Nodule-specific PLAT domain proteins are expanded in the Medicago lineage and required for nodulation. New Phytol. 2019, 222, 1538–1550. [Google Scholar] [CrossRef] [PubMed]
- Pislariu, C.I.; Sinharoy, S.; Torres-Jerez, I.; Nakashima, J.; Blancaflor, E.B.; Udvardi, M.K. The Nodule-Specific PLAT Domain Protein NPD1 Is Required for Nitrogen-Fixing Symbiosis. Plant Physiol. 2019, 180, 1480–1497. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yang, S.; Tang, F.; Zhu, H. Symbiosis specificity in the legume: Rhizobial mutualism. Cell. Microbiol. 2012, 14, 334–342. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Q.; Fedorova, E.; Liu, J.; Qin, Q.; Zheng, Q.; Price, P.A.; Pan, H.; Wang, D.; Griffitts, J.S.; et al. Microsymbiont discrimination mediated by a host-secreted peptide in Medicago truncatula. Proc. Natl. Acad. Sci. USA 2017, 114, 6848–6853. [Google Scholar] [CrossRef]
- Roy, P.; Achom, M.; Wilkinson, H.; Lagunas, B.; Gifford, M.L. Symbiotic Outcome Modified by the Diversification from 7 to over 700 Nodule-Specific Cysteine-Rich Peptides. Genes 2020, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Downie, J.A.; Kondorosi, E. Why Should Nodule Cysteine-Rich (NCR) Peptides Be Absent From Nodules of Some Groups of Legumes but Essential for Symbiotic N-Fixation in Others? Front. Agron. 2021, 3, 654576. [Google Scholar] [CrossRef]
- Montiel, J.; Downie, J.A.; Farkas, A.; Bihari, P.; Herczeg, R.; Bálint, B.; Mergaert, P.; Kereszt, A.; Kondorosi, É. Morphotype of bacteroids in different legumes correlates with the number and type of symbiotic NCR peptides. Proc. Natl. Acad. Sci. USA 2017, 114, 5041–5046. [Google Scholar] [CrossRef]
- Evers, G.W. Forage legumes: Forage quality, fixed nitrogen, or both. Crop Sci. 2011, 51, 403–409. [Google Scholar] [CrossRef]
- Carlsson, G.; Huss-Danell, K. Nitrogen fixation in perennial forage legumes in the field. Plant Soil 2003, 253, 353–372. [Google Scholar] [CrossRef]
- Ball, D.M.; Hoveland, C.S.; Lacefield, G.D. Southern Forages; International Plant Nutrition Group: Peachtree Corners, GA, USA, 2007; 332p. [Google Scholar]
- Taylor, N. A century of clover breeding developments in the United States. Crop Sci. 2008, 48, 1–13. [Google Scholar] [CrossRef]
- Vizintin, L.; Javornik, B.; Bohanec, B. Genetic characterization of selected Trifolium species as revealed by nuclear DNA content and ITS rDNA region analysis. Plant Sci. 2006, 170, 859–866. [Google Scholar] [CrossRef]
- Riday, H.; Krohn, A.L. Genetic map-based location of the red clover (Trifolium pratense L.) gametophytic self-incompatibility locus. Theor. Appl. Genet. 2010, 121, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, M.; Dinkins, R.D.; Hunt, A.G. De novo Transcriptome Assembly and Dynamic Spatial Gene Expression Analysis in Red Clover. Plant Genome-US 2016, 9, plantgenome2015.06.0048. [Google Scholar] [CrossRef] [PubMed]
- Istvanek, J.; Jaros, M.; Krenek, A.; Repkova, J. Genome assembly and annotation for red clover (Trifolium pratense; Fabaceae). Am. J. Bot. 2014, 101, 327–337. [Google Scholar] [CrossRef] [PubMed]
- De Vega, J.J.; Ayling, S.; Hegarty, M.; Kudrna, D.; Goicoechea, J.L.; Ergon, A.; Rognli, O.A.; Jones, C.; Swain, M.; Geurts, R.; et al. Red clover (Trifolium pratense L.) draft genome provides a platform for trait improvement. Sci. Rep. 2015, 5, 17394. [Google Scholar] [CrossRef] [PubMed]
- Bickhart, D.M.; Koch, L.M.; Smith, T.P.L.; Riday, H.; Sullivan, M.L. Chromosome-scale assembly of the highly heterozygous genome of red clover (Trifolium pratense L.), an allogamous forage crop species. Gigabyte 2022, 2022, 1–13. [Google Scholar] [CrossRef]
- Li, W.; Riday, H.; Riehle, C.; Edwards, A.; Dinkins, R. Identification of Single Nucleotide Polymorphism in Red Clover (Trifolium pratense L.) Using Targeted Genomic Amplicon Sequencing and RNA-seq. Front. Plant Sci. 2019, 10, 1257. [Google Scholar] [CrossRef]
- Osterman, J.; Hammenhag, C.; Ortiz, R.; Geleta, M. Insights Into the Genetic Diversity of Nordic Red Clover (Trifolium pratense) Revealed by SeqSNP-Based Genic Markers. Front. Plant Sci. 2021, 12, 748750. [Google Scholar] [CrossRef]
- Jones, C.; De Vega, J.; Lloyd, D.; Hegarty, M.; Ayling, S.; Powell, W.; Skøt, L. Population structure and genetic diversity in red clover (Trifolium pratense L.) germplasm. Sci. Rep. 2020, 10, 8364. [Google Scholar] [CrossRef] [PubMed]
- Istvanek, J.; Dluhosova, J.; Dluhos, P.; Patkova, L.; Nedelnik, J.; Repkova, J. Gene Classification and Mining of Molecular Markers Useful in Red Clover (Trifolium pratense) Breeding. Front. Plant Sci. 2017, 8, 367. [Google Scholar] [CrossRef]
- Benevenuto, J.; Bhakta, M.; Lohr, D.A.; Ferrão, L.F.V.; Resende, M.F.R.; Kirst, M.; Quesenberry, K.; Munoz, P. Cost-effective detection of genome-wide signatures for 2,4-D herbicide resistance adaptation in red clover. Sci. Rep. 2019, 9, 20037. [Google Scholar] [CrossRef]
- Trněný, O.; Vlk, D.; Macková, E.; Matoušková, M.; Řepková, J.; Nedělník, J.; Hofbauer, J.; Vejražka, K.; Jakešová, H.; Jansa, J.; et al. Allelic Variants for Candidate Nitrogen Fixation Genes Revealed by Sequencing in Red Clover (Trifolium pratense L.). Int. J. Mol. Sci. 2019, 20, 5470. [Google Scholar] [CrossRef]
- Dinkins, R.D.; Hancock, J.; Coe, B.L.; May, J.B.; Goodman, J.P.; Bass, W.T.; Liu, J.; Fan, Y.; Zheng, Q.; Zhu, H. Isoflavone levels, nodulation and gene expression profiles of a CRISPR/Cas9 deletion mutant in the isoflavone synthase gene of red clover. Plant Cell Rep. 2021, 40, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Hohnjec, N.; Lenz, F.; Fehlberg, V.; Vieweg, M.F.; Baier, M.C.; Hause, B.; Küster, H. The Signal Peptide of the Medicago truncatula Modular Nodulin MtNOD25 Operates as an Address Label for the Specific Targeting of Proteins to Nitrogen-Fixing Symbiosomes. Mol. Plant-Microbe Interact. 2009, 22, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, M.; Dinkins, R.D.; Hunt, A.G. Genome-wide atlas of alternative polyadenylation in the forage legume red clover. Sci. Rep. 2018, 8, 11379. [Google Scholar] [CrossRef] [PubMed]
- Jenei, S.; Tiricz, H.; Szolomájer, J.; Tímár, E.; Klement, É.; Al Bouni, M.A.; Lima, R.M.; Kata, D.; Harmati, M.; Buzás, K.; et al. Potent Chimeric Antimicrobial Derivatives of the Medicago truncatula NCR247 Symbiotic Peptide. Front. Microbiol. 2020, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.M.; Kylarová, S.; Mergaert, P.; Kondorosi, É. Unexplored Arsenals of Legume Peptides with Potential for Their Applications in Medicine and Agriculture. Front. Microbiol. 2020, 11, 1307. [Google Scholar] [CrossRef]
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2019, 88, 107–118. [Google Scholar] [CrossRef]
- Terras, F.R.; Eggermont, K.; Kovaleva, V.; Raikhel, N.V.; Osborn, R.W.; Kester, A.; Rees, S.B.; Torrekens, S.; Van Leuven, F.; Vanderleyden, J. Small cysteine-rich antifungal proteins from radish: Their role in host defense. Plant Cell 1995, 7, 573–588. [Google Scholar] [CrossRef]
- Burghardt, L.T.; Trujillo, D.I.; Epstein, B.; Tiffin, P.; Young, N.D. A Select and Resequence Approach Reveals Strain-Specific Effects of Medicago Nodule-Specific PLAT-Domain Genes. Plant Physiol. 2020, 182, 463–471. [Google Scholar] [CrossRef]
- Schauser, L.; Roussis, A.; Stiller, J.; Stougaard, J. A plant regulator controlling development of symbiotic root nodules. Nature 1999, 402, 191–195. [Google Scholar] [CrossRef]
- Soyano, T.; Kouchi, H.; Hirota, A.; Hayashi, M. Nodule Inception Directly Targets NF-Y Subunit Genes to Regulate Essential Processes of Root Nodule Development in Lotus japonicus. PLoS Genet. 2013, 9, e1003352. [Google Scholar] [CrossRef]
- Laloum, T.; Baudin, M.; Frances, L.; Lepage, A.; Billault-Penneteau, B.; Cerri, M.R.; Ariel, F.; Jardinaud, M.-F.; Gamas, P.; de Carvalho-Niebel, F.; et al. Two CCAAT-box-binding transcription factors redundantly regulate early steps of the legume-rhizobia endosymbiosis. Plant J. 2014, 79, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Combier, J.-P.; Frugier, F.; de Billy, F.; Boualem, A.; El-Yahyaoui, F.; Moreau, S.; Vernié, T.; Ott, T.; Gamas, P.; Crespi, M.; et al. MtHAP2-1 is a key transcriptional regulator of symbiotic nodule development regulated by microRNA169 in Medicago truncatula. Genes Dev. 2006, 20, 3084–3088. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.T.; Schilderink, S.; Moling, S.; Deinum, E.E.; Kondorosi, E.; Franssen, H.; Kulikova, O.; Niebel, A.; Bisseling, T. Fate map of Medicago truncatula root nodules. Development 2014, 141, 3517–3528. [Google Scholar] [CrossRef] [PubMed]
- Fournier, J.; Teillet, A.; Chabaud, M.; Ivanov, S.; Genre, A.; Limpens, E.; de Carvalho-Niebel, F.; Barker, D.G. Remodeling of the Infection Chamber before Infection Thread Formation Reveals a Two-Step Mechanism for Rhizobial Entry into the Host Legume Root Hair. Plant Physiol. 2015, 167, 1233–1242. [Google Scholar] [CrossRef]
- Xie, F.; Murray, J.D.; Kim, J.; Heckmann, A.B.; Edwards, A.; Oldroyd, G.E.D.; Downie, J.A. Legume pectate lyase required for root infection by rhizobia. Proc. Natl. Acad. Sci. USA 2011, 109, 633–638. [Google Scholar] [CrossRef]
- Marsh, J.F.; Rakocevic, A.; Mitra, R.M.; Brocard, L.; Sun, J.; Eschstruth, A.; Long, S.R.; Schultze, M.; Ratet, P.; Oldroyd, G.E.D. Medicago truncatula NIN Is Essential for Rhizobial-Independent Nodule Organogenesis Induced by Autoactive Calcium/Calmodulin-Dependent Protein Kinase. Plant Physiol. 2007, 144, 324–335. [Google Scholar] [CrossRef]
- Vernié, T.; Kim, J.; Frances, L.; Ding, Y.; Sun, J.; Guan, D.; Niebel, A.; Gifford, M.L.; de Carvalho-Niebel, F.; Oldroyd, G.E.D. The NIN Transcription Factor Coordinates Diverse Nodulation Programs in Different Tissues of the Medicago truncatula Root. Plant Cell 2015, 27, 3410–3424. [Google Scholar] [CrossRef]
- Liu, J.; Rutten, L.; Limpens, E.; van der Molen, T.; van Velzen, R.; Chen, R.; Chen, Y.; Geurts, R.; Kohlen, W.; Kulikova, O.; et al. A Remote cis-Regulatory Region Is Required for NIN Expression in the Pericycle to Initiate Nodule Primordium Formation in Medicago truncatula. Plant Cell 2019, 31, 68–83. [Google Scholar] [CrossRef]
- Combier, J.P.; de Billy, F.; Gamas, P.; Niebel, A.; Rivas, S. Trans-regulation of the expression of the transcription factor MtHAP2-1 by a uORF controls root nodule development. Genes Dev. 2008, 22, 1549–1559. [Google Scholar] [CrossRef]
- Sorin, C.; Declerck, M.; Christ, A.; Blein, T.; Ma, L.; Lelandais-Brière, C.; Njo, M.F.; Beeckman, T.; Crespi, M.; Hartmann, C. A miR169 isoform regulates specificNF-YAtargets and root architecture in Arabidopsis. New Phytol. 2014, 202, 1197–1211. [Google Scholar] [CrossRef]
- Wang, D.; Griffitts, J.; Starker, C.; Fedorova, E.; Limpens, E.; Ivanov, S.; Bisseling, T.; Long, S. A nodule-specific protein secretory pathway required for nitrogen-fixing symbiosis. Science 2010, 327, 1126–1129. [Google Scholar] [CrossRef] [PubMed]
- Huisman, R.; Hontelez, J.; Mysore, K.S.; Wen, J.; Bisseling, T.; Limpens, E. A symbiosis-dedicated SYNTAXIN OF PLANTS 13II isoform controls the formation of a stable host–microbe interface in symbiosis. New Phytol. 2016, 211, 1338–1351. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Oztas, O.; Zhang, X.; Wu, X.; Stonoha, C.; Wang, E.; Wang, B.; Wang, D. A symbiotic SNARE protein generated by alternative termination of transcription. Nat. Plants 2016, 2, 15197. [Google Scholar] [CrossRef]
- Yu, L.; Huang, T.; Qi, X.; Yu, J.; Wu, T.; Luo, Z.; Zhou, L.; Li, Y. Genome-Wide Analysis of Long Non-coding RNAs Involved in Nodule Senescence in Medicago truncatula. Front. Plant Sci. 2022, 13, 917840. [Google Scholar] [CrossRef]
- Sinharoy, S.; Torres-Jerez, I.; Bandyopadhyay, K.; Kereszt, A.; Pislariu, C.I.; Nakashima, J.; Benedito, V.A.; Kondorosi, E.; Udvardi, M.K. The C2H2 Transcription Factor REGULATOR OF SYMBIOSOME DIFFERENTIATION Represses Transcription of the Secretory Pathway Gene VAMP721a and Promotes Symbiosome Development in Medicago truncatula. Plant Cell 2013, 25, 3584–3601. [Google Scholar] [CrossRef]
- Roux, B.; Rodde, N.; Jardinaud, M.-F.; Timmers, T.; Sauviac, L.; Cottret, L.; Carrère, S.; Sallet, E.; Courcelle, E.; Moreau, S.; et al. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser-capture microdissection coupled to RNA sequencing. Plant J. 2014, 77, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Bisseling, T.; Geurts, R. Specificity in legume nodule symbiosis. Science 2020, 369, 620–621. [Google Scholar] [CrossRef]
- Bozsoki, Z.; Cheng, J.; Feng, F.; Gysel, K.; Vinther, M.; Andersen, K.R.; Oldroyd, G.; Blaise, M.; Radutoiu, S.; Stougaard, J. Receptor-mediated chitin perception in legume roots is functionally separable from Nod factor perception. Proc. Natl. Acad. Sci. USA 2017, 114, E8118–E8127. [Google Scholar] [CrossRef]
- Zhou, P.; Silverstein, K.A.T.; Gao, L.; Walton, J.D.; Nallu, S.; Guhlin, J.; Young, N.D. Detecting small plant peptides using SPADA (Small Peptide Alignment Discovery Application). BMC Bioinform. 2013, 14, 335. [Google Scholar] [CrossRef]
- Nielsen, H.; Engelbrecht, J.; Brunak, S.; von Heijne, G. Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Eng. Des. Sel. 1997, 10, 1–6. [Google Scholar] [CrossRef]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved Prediction of Signal Peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Feature ID | Normalized Reads (Means) Nod− | Normalized Reads (Means) Nod+ | Log2 Diff Express (Nod+) /(Nod−) | Product |
|---|---|---|---|---|
| LOC123882175 | 381.52 | 74,205.38 | 7.60 | leghemoglobin_Lb120-1 |
| LOC123882181 | 31.62 | 63,699.05 | 10.98 | leghemoglobin_Lb120-1 |
| LOC123916815 | 15.55 | 57,593.44 | 11.86 | nodulin-25-like isoform X10 |
| LOC123902448 | 328.64 | 38,271.05 | 6.86 | early nodulin-75-like |
| LOC123882180 | 7.62 | 28,608.07 | 11.88 | leghemoglobin_Lb120-1 |
| LOC123917408 | 97.65 | 27,409.22 | 8.13 | nodulin-26-like isoform X2 |
| LOC123882176 | 17.47 | 24,305.17 | 10.44 | leghemoglobin_Lb120-1 |
| LOC123899073 | 4.50 | 22,133.74 | 12.26 | Embryo-specific protein ATS3B-like; NPD |
| LOC123884281 | 5.08 | 20,011.87 | 11.95 | ctenidin-1-like |
| LOC123916883 | 6.72 | 17,961.34 | 11.38 | NMS32/34 protein, putative |
| LOC123915658 | 3049.67 | 13,975.22 | 2.20 | bifunctional aspartate aminotransferase |
| LOC123893140 | 4110.65 | 13,859.39 | 1.75 | legume-specific_protein |
| LOC123918940 | 4.23 | 13,807.55 | 11.67 | NMS32/34 protein, putative |
| LOC123891069 | 18.14 | 13,225.90 | 9.51 | leghemoglobin_Lb120-1 |
| LOC123888452 | 57.99 | 12,010.79 | 7.69 | early_nodulin_ENOD18 |
| LOC123913320 | 2502.68 | 11,384.90 | 2.19 | glutamine synthetase nodule isozyme |
| LOC123909356 | 6.27 | 10,611.03 | 10.72 | basic blue protein-like |
| LOC123895631 | 2916.89 | 10,575.69 | 1.86 | inactive beta-amylase 9 |
| LOC123921269 | 829.79 | 10,351.60 | 3.64 | chaperone protein dnaJ 8, chloroplastic |
| LOC123911535 | 4108.67 | 10,330.98 | 1.33 | heavy metal-associated isoprenylated plant protein 6-like |
| Feature ID | Normalized Reads (Means) Nod− | Normalized Reads (Means) Nod+ | Log2 Diff Express (Nod+) /(Nod−) | Product |
|---|---|---|---|---|
| LOC123899143 | 201.60 | 3.08 | 6.03 | wound-responsive_family_protein |
| LOC123898241 | 210.61 | 3.43 | 5.94 | wound-responsive_family_protein |
| LOC123899195 | 171.98 | 2.86 | 5.91 | structural_constituent_of_cell_wall_protein,_putative |
| LOC123900972 | 110.72 | 2.63 | 5.39 | sesquiterpene_synthase |
| LOC123923258 | 99.89 | 3.96 | 4.66 | sesquiterpene_synthase |
| LOC123908919 | 312.53 | 13.93 | 4.49 | geranylgeranyl_pyrophosphate_synthase |
| LOC123891221 | 66.37 | 3.09 | 4.42 | disease_resistance_protein_(TIR-NBS-LRR_class) |
| LOC123920096 | 45.48 | 2.20 | 4.37 | replication_factor-A_carboxy-terminal_domain_protein |
| LOC123917771 | 62.76 | 3.61 | 4.12 | translation_elongation_factor_EF1B,_gamma_chain |
| LOC123908518 | 518.67 | 30.81 | 4.07 | benzyl_alcohol_O-benzoyltransferase |
| LOC123924362 | 826.36 | 52.79 | 3.97 | polygalacturonase_plant-like_protein |
| LOC123921773 | 1639.61 | 108.42 | 3.92 | LysM_receptor_kinase_K1B |
| LOC123899196 | 98.73 | 6.92 | 3.83 | 1-aminocyclopropane-1-carboxylate_oxidase-like_protein |
| LOC123891479 | 241.74 | 18.20 | 3.73 | cysteine/histidine-rich_C1_domain_protein |
| LOC123921461 | 230.24 | 17.49 | 3.72 | cytochrome_P450_family_71_protein |
| LOC123912953 | 25.87 | 2.06 | 3.65 | cytochrome_P450_family_71_protein |
| LOC123909630 | 39.54 | 3.18 | 3.64 | amino acid transporter AVT1H |
| LOC123919510 | 15,190.12 | 1224.73 | 3.63 | alpha-copaene synthase-like |
| LOC123891304 | 59.92 | 4.84 | 3.63 | zinc finger protein 6-like |
| LOC123919667 | 210.91 | 17.06 | 3.63 | disease_resistance-responsive,_dirigent_domain_protein |
| GO Term | Ontology | Description | Number in Input List | Number in BG/Ref | p-Value | FDR |
|---|---|---|---|---|---|---|
| GO:0071555 | P | cell wall organization | 8 | 62 | 6.10 × 10−5 | 0.016 |
| GO:0045229 | P | external encapsulating structure organization | 8 | 62 | 6.10 × 10−5 | 0.016 |
| GO:0071554 | P | cell wall organization or biogenesis | 8 | 84 | 5.10 × 10−4 | 0.077 |
| GO:0042545 | P | cell wall modification | 6 | 48 | 0.0006 | 0.077 |
| GO:0030599 | F | pectinesterase activity | 6 | 48 | 0.0006 | 0.059 |
| GO:0020037 | F | heme binding | 20 | 384 | 0.00042 | 0.059 |
| GO:0046906 | F | tetrapyrrole binding | 20 | 385 | 0.00044 | 0.059 |
| GO:0052689 | F | carboxylic ester hydrolase activity | 7 | 68 | 0.00071 | 0.059 |
| GO:0016705 | F | oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen | 16 | 292 | 0.00086 | 0.059 |
| GO:0030312 | C | external encapsulating structure | 9 | 76 | 0.000043 | 0.0021 |
| GO:0005618 | C | cell wall | 9 | 76 | 0.000043 | 0.0021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinkins, R.D.; Hancock, J.A.; Bickhart, D.M.; Sullivan, M.L.; Zhu, H. Expression and Variation of the Genes Involved in Rhizobium Nodulation in Red Clover. Plants 2022, 11, 2888. https://doi.org/10.3390/plants11212888
Dinkins RD, Hancock JA, Bickhart DM, Sullivan ML, Zhu H. Expression and Variation of the Genes Involved in Rhizobium Nodulation in Red Clover. Plants. 2022; 11(21):2888. https://doi.org/10.3390/plants11212888
Chicago/Turabian StyleDinkins, Randy D., Julie A. Hancock, Derek M. Bickhart, Michael L. Sullivan, and Hongyan Zhu. 2022. "Expression and Variation of the Genes Involved in Rhizobium Nodulation in Red Clover" Plants 11, no. 21: 2888. https://doi.org/10.3390/plants11212888
APA StyleDinkins, R. D., Hancock, J. A., Bickhart, D. M., Sullivan, M. L., & Zhu, H. (2022). Expression and Variation of the Genes Involved in Rhizobium Nodulation in Red Clover. Plants, 11(21), 2888. https://doi.org/10.3390/plants11212888