
Intraspecific Leaf Trait Variation across and within Five Common Wine Grape Varieties

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Design
2.2. Functional Trait Measurements
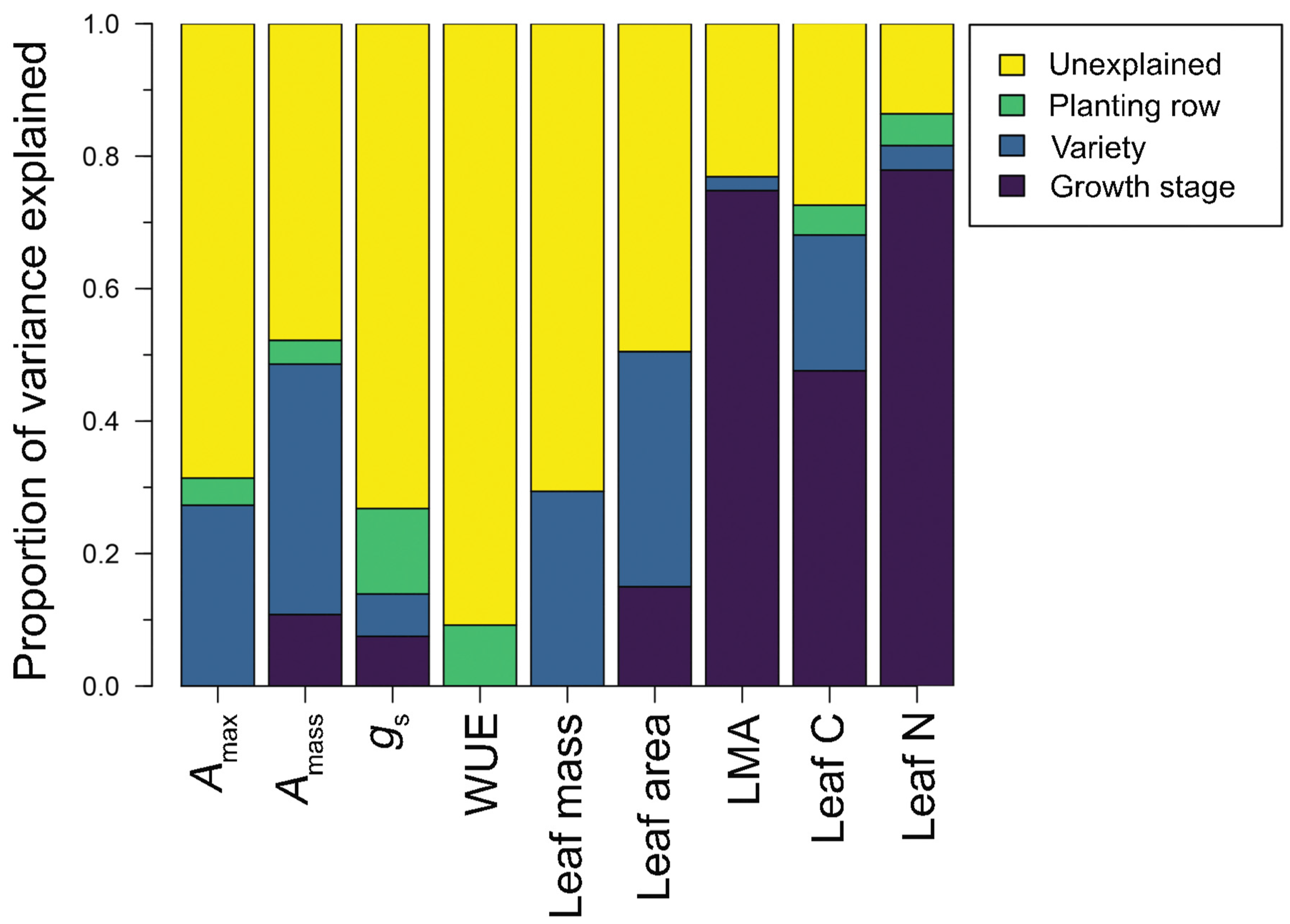
2.3. Data Analysis—Causes of Intraspecific Trait Variation in Wine Grape Varieties
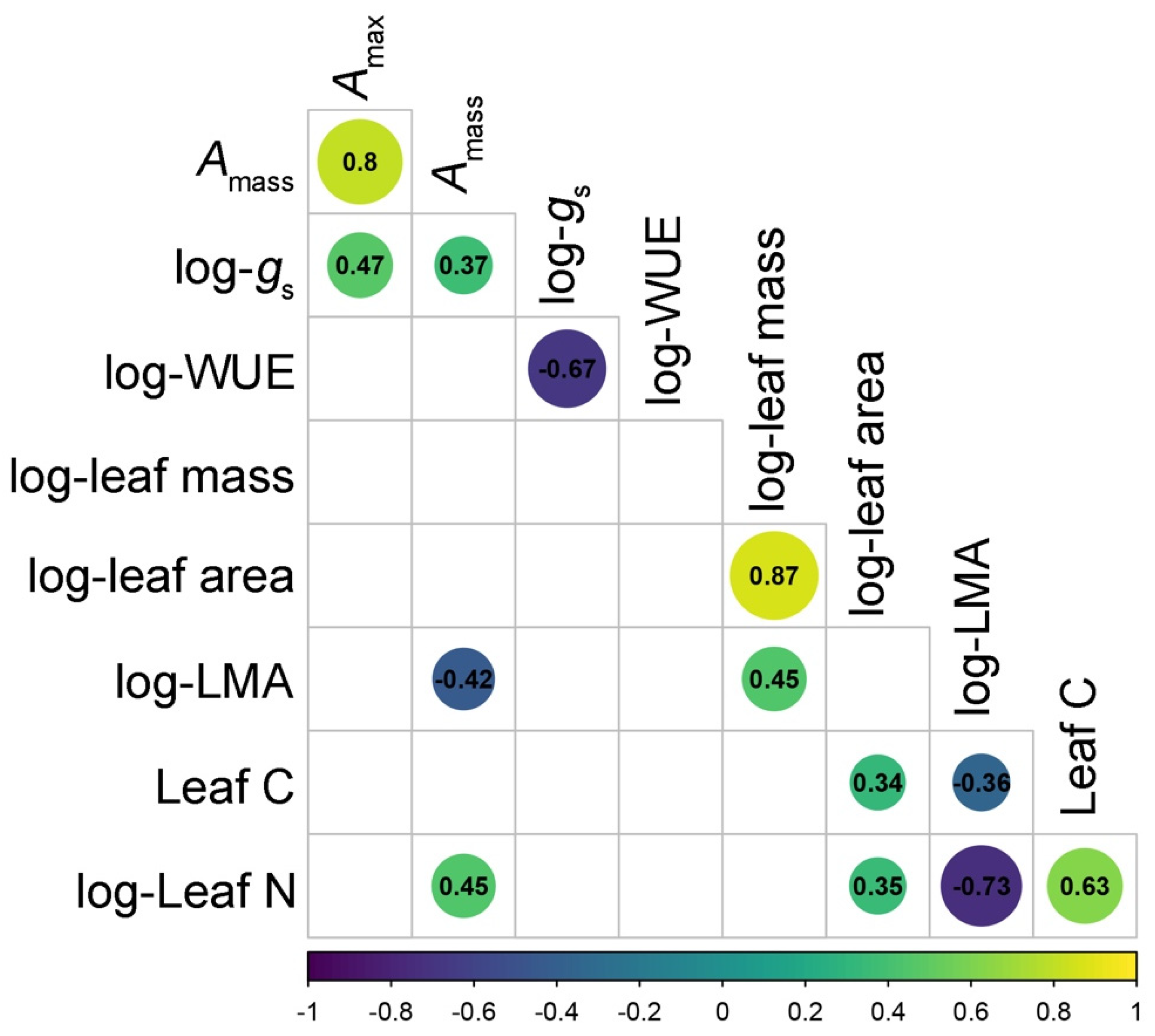
2.4. Data Analysis—Bivariate and Multivariate Trait Correlations
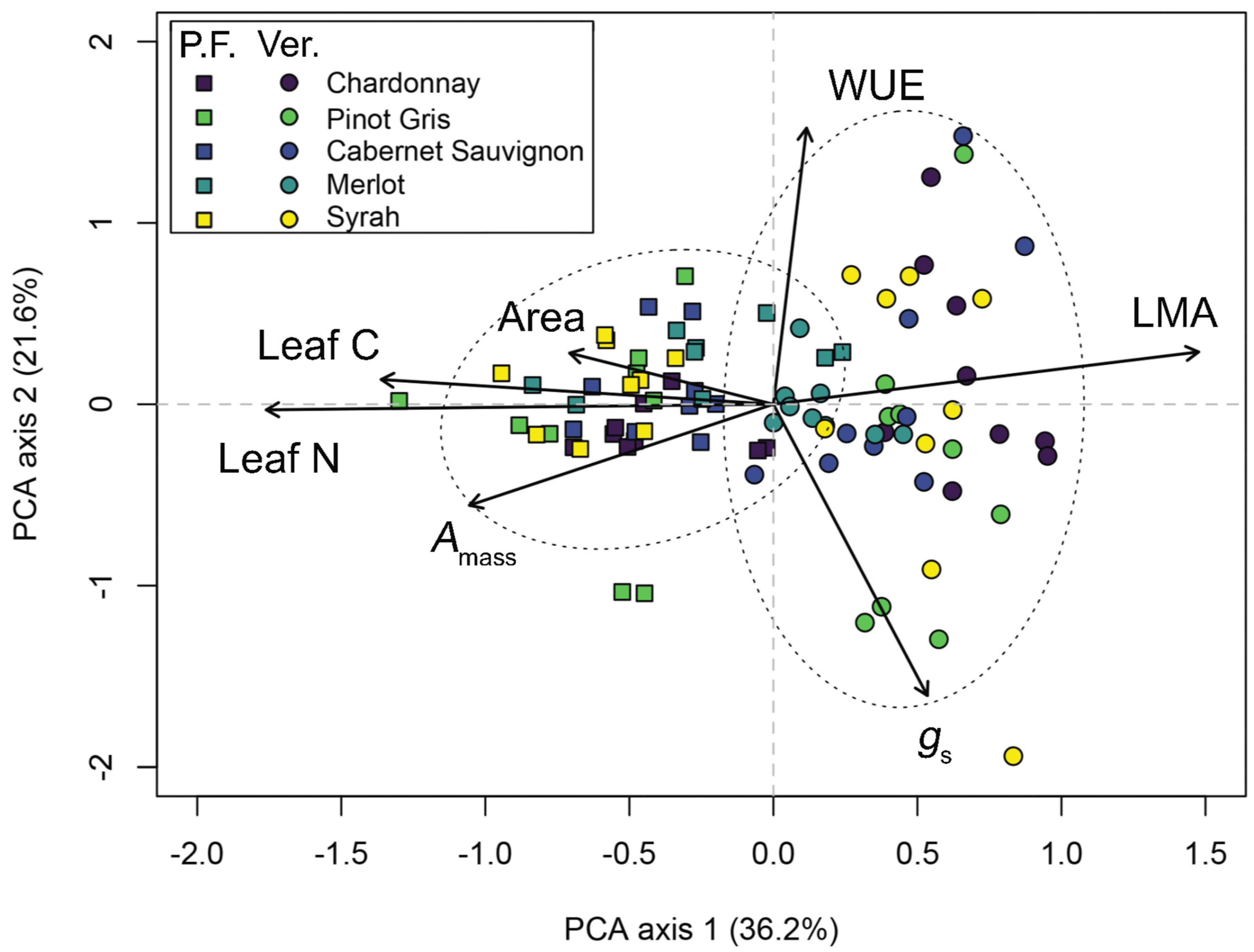
2.5. Data Analysis—An Intraspecific LES across Wine Grape Varieties
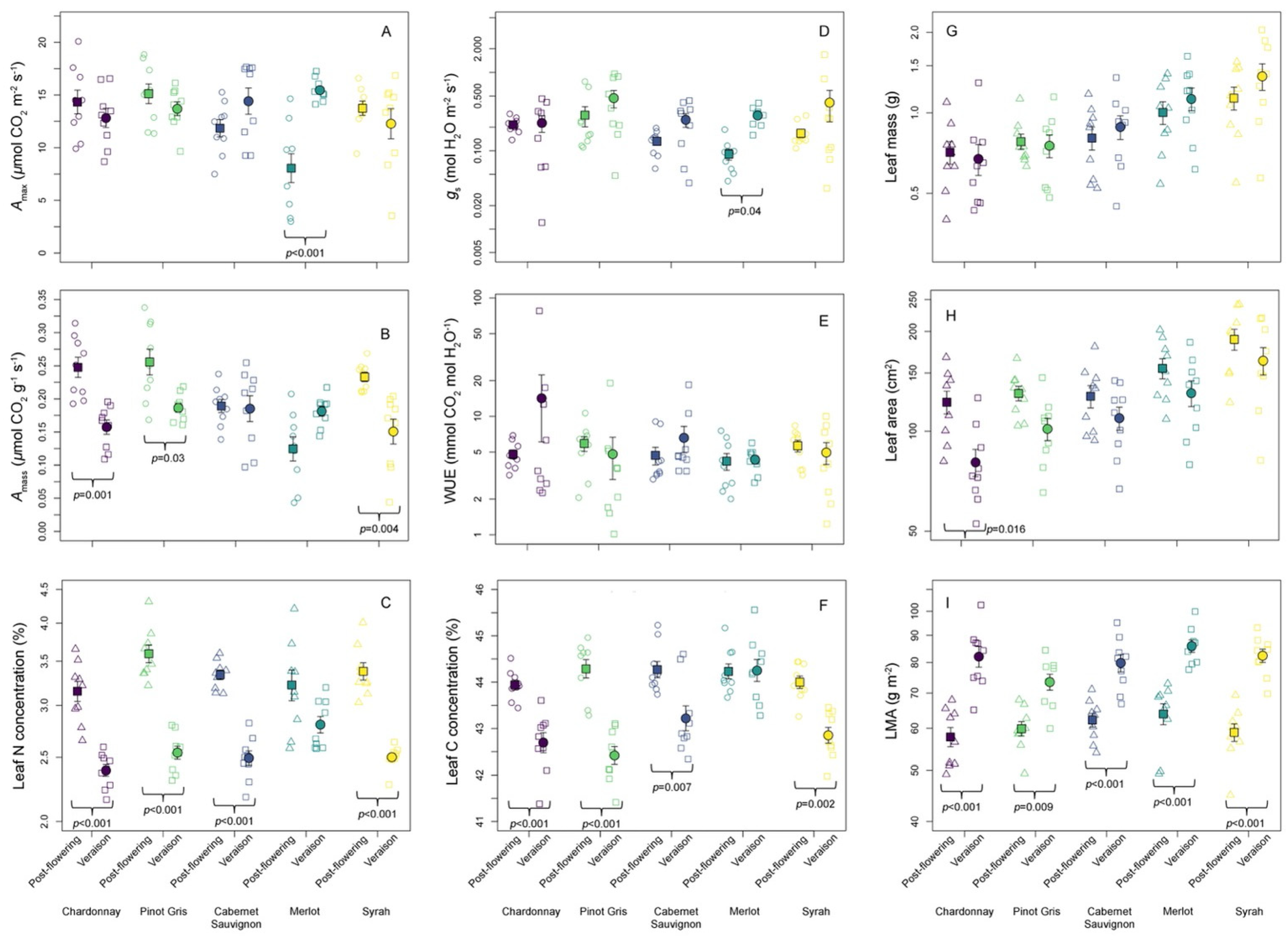
3. Results
3.1. Trait Variation across Wine Grape Varieties
3.2. Relationships among LES and Other Leaf Traits in Wine Grape Varieties
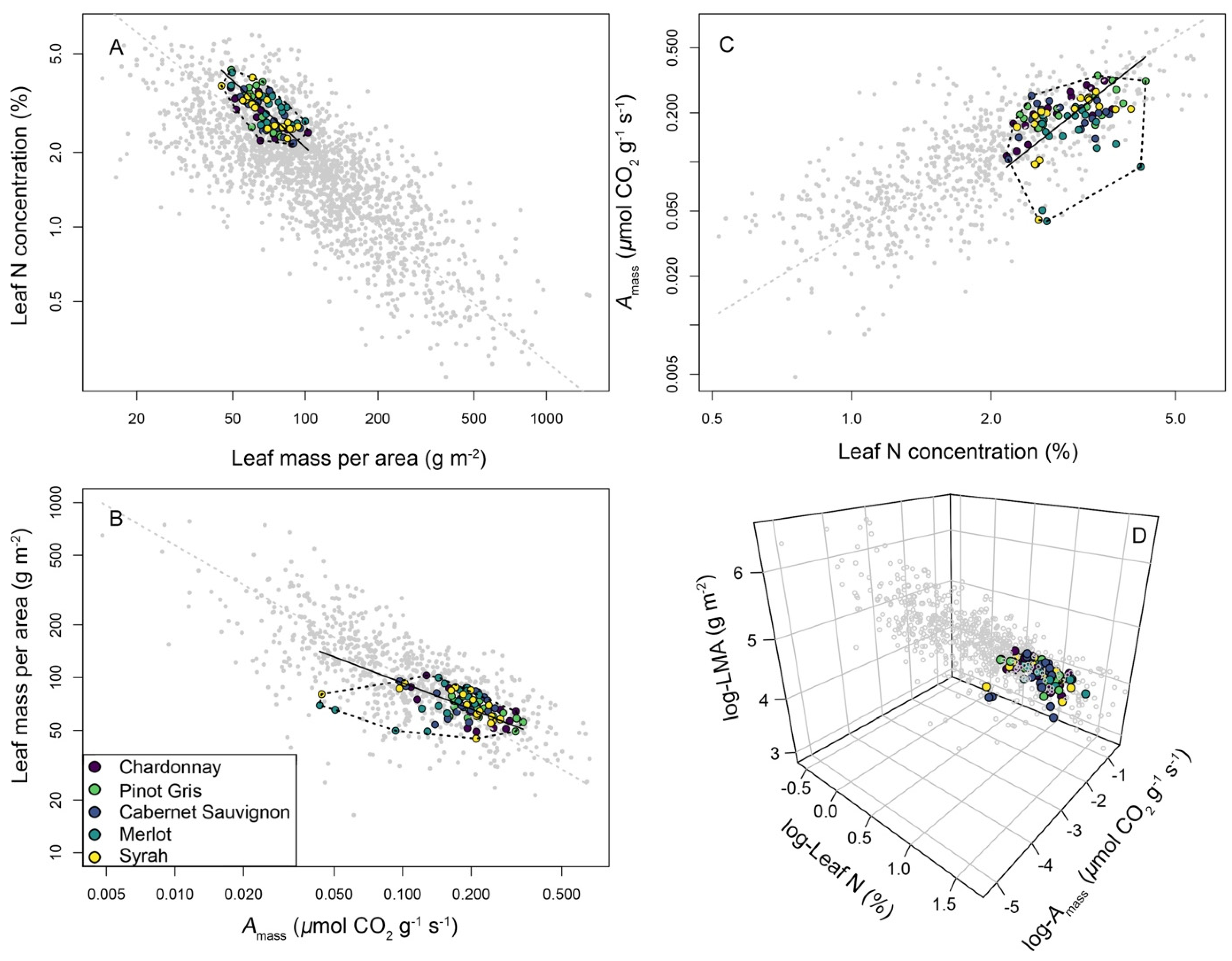
3.3. A Leaf Economics Spectrum across Wine Grape Varieties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B.; Vose, J.M.; Gresham, C.; Volin, J.C.; Bowman, W.D. Generality of leaf trait relationships: A test across six biomes. Ecology 1999, 80, 1955–1969. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Jackson, B.G.; Peltzer, D.A.; Wardle, D.A. The within-species leaf economic spectrum does not predict leaf litter decomposability at either the within-species or whole community levels. J. Ecol. 2013, 101, 1409–1419. [Google Scholar] [CrossRef]
- Niinemets, Ü. Is there a species spectrum within the world-wide leaf economics spectrum? Major variations in leaf functional traits in the Mediterranean sclerophyll Quercus ilex. New Phytol. 2015, 205, 79–96. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast–slow’plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Falster, D.S.; Duursma, R.A.; FitzJohn, R.G. How functional traits influence plant growth and shade tolerance across the life cycle. Proc. Natl. Acad. Sci. USA 2018, 115, E6789–E6798. [Google Scholar] [CrossRef]
- Givnish, T. Adaptation to sun and shade: A whole-plant perspective. Funct. Plant Biol. 1988, 15, 63–92. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, A key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Bakker, M.A.; Carreño-Rocabado, G.; Poorter, L. Leaf economics traits predict litter decomposition of tropical plants and differ among land use types. Funct. Ecol. 2011, 25, 473–483. [Google Scholar] [CrossRef]
- Sakschewski, B.; Von Bloh, W.; Boit, A.; Rammig, A.; Kattge, J.; Poorter, L.; Penuelas, J.; Thonicke, K. Leaf and stem economics spectra drive diversity of functional plant traits in a dynamic global vegetation model. Glob. Chang. Biol. 2015, 21, 2711–2725. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.R.; Isaac, M.E. Plant functional traits in agroecosystems: A blueprint for research. J. Appl. Ecol. 2015, 52, 1425–1435. [Google Scholar] [CrossRef]
- Hayes, F.J.; Buchanan, S.W.; Coleman, B.; Gordon, A.M.; Reich, P.B.; Thevathasan, N.V.; Wright, I.; Martin, A.R. Intraspecific variation in soy across the leaf economics spectrum. Ann. Bot. 2019, 123, 107–120. [Google Scholar] [CrossRef]
- Martin, A.R.; Isaac, M.E. The leaf economics spectrum’s morning coffee: Plant size-dependent changes in leaf traits and reproductive onset in a perennial tree crop. Ann. Bot. 2021, 127, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.R.; Rapidel, B.; Roupsard, O.; Van den Meersche, K.; de Melo Virginio Filho, E.; Barrios, M.; Isaac, M.E. Intraspecific trait variation across multiple scales: The leaf economics spectrum in coffee. Funct. Ecol. 2017, 31, 604–612. [Google Scholar] [CrossRef]
- Roucou, A.; Violle, C.; Fort, F.; Roumet, P.; Ecarnot, M.; Vile, D. Shifts in plant functional strategies over the course of wheat domestication. J. Appl. Ecol. 2018, 55, 25–37. [Google Scholar] [CrossRef]
- Martin, A.R.; Hale, C.E.; Cerabolini, B.E.; Cornelissen, J.H.; Craine, J.; Gough, W.A.; Kattge, J.; Tirona, C.K. Inter-and intraspecific variation in leaf economic traits in wheat and maize. AoB Plants 2018, 10, ply006. [Google Scholar] [CrossRef]
- Sauvadet, M.; Dickinson, A.K.; Somarriba, E.; Phillips-Mora, W.; Cerda, R.H.; Martin, A.R.; Isaac, M.E. Genotype–environment interactions shape leaf functional traits of cacao in agroforests. Agron. Sustain. Dev. 2021, 41, 31. [Google Scholar] [CrossRef]
- Xiong, D.; Flexas, J. Leaf economics spectrum in rice: Leaf anatomical, biochemical, and physiological trait trade-offs. J. Exp. Bot. 2018, 69, 5599–5609. [Google Scholar] [CrossRef]
- Mason, C.M.; McGaughey, S.E.; Donovan, L. Ontogeny strongly and differentially alters leaf economic and other key traits in three diverse Helianthus species. J. Exp. Bot. 2013, 64, 4089–4099. [Google Scholar] [CrossRef]
- Gagliardi, S.; Martin, A.R.; Virginio Filho, E.d.M.; Rapidel, B.; Isaac, M.E. Intraspecific leaf economic trait variation partially explains coffee performance across agroforestry management regimes. Agric. Ecosyst. Environ. 2015, 200, 151–160. [Google Scholar] [CrossRef]
- Coleman, B.R.; Martin, A.R.; Thevathasan, N.V.; Gordon, A.M.; Isaac, M.E. Leaf trait variation and decomposition in short-rotation woody biomass crops under agroforestry management. Agric. Ecosyst. Environ. 2020, 298, 106971. [Google Scholar] [CrossRef]
- Fulthorpe, R.; Martin, A.R.; Isaac, M.E. Root endophytes of coffee (Coffea arabica): Variation across climatic gradients and relationships with functional traits. Phytobiomes J. 2020, 4, 27–39. [Google Scholar] [CrossRef]
- Martin, A.R.; Hayes, F.J.; Borden, K.A.; Buchanan, S.W.; Gordon, A.M.; Isaac, M.E.; Thevathasan, N.V. Integrating nitrogen fixing structures into above- and belowground functional trait spectra in soy (Glycine max). Plant Soil 2019, 440, 53–69. [Google Scholar] [CrossRef]
- Milla, R.; Osborne, C.P.; Turcotte, M.M.; Violle, C. Plant domestication through an ecological lens. Trends Ecol. Evol. 2015, 30, 463–469. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Ranjan, A.; Martinez, C.C.; Headland, L.; Thiem, T.; Kumar, R.; Covington, M.F.; Hatcher, T.; Naylor, D.T.; Zimmerman, S.; et al. A modern ampelography: A genetic basis for leaf shape and venation patterning in grape. Plant Physiol. 2014, 164, 259–272. [Google Scholar] [CrossRef]
- Lavely, E.K.; Chen, W.; Peterson, K.A.; Klodd, A.E.; Volder, A.; Marini, R.P.; Eissenstat, D.M. On characterizing root function in perennial horticultural crops. Am. J. Bot. 2020, 107, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Wolkovich, E.M.; Burge, D.O.; Walker, M.A.; Nicholas, K. Phenological diversity provides opportunities for climate change adaptation in winegrapes. J. Ecol. 2017, 105, 905–912. [Google Scholar] [CrossRef]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine Responses to Heat Stress and Global Warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef] [PubMed]
- Wolkovich, E.; García de Cortázar-Atauri, I.; Morales-Castilla, I.; Nicholas, K.; Lacombe, T. From Pinot to Xinomavro in the world’s future wine-growing regions. Nat. Clim. Chang. 2018, 8, 29–37. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Coombe, B.G. Growth stages of the grapevine: Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. Corrigendum to: New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2016, 64, 715–716. [Google Scholar] [CrossRef]
- Delignette-Muller, M.L.; Dutang, C. Fitdistrplus: An R package for fitting distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef]
- Messier, J.; McGill, B.J.; Lechowicz, M.J. How do traits vary across ecological scales? A case for trait-based ecology. Ecol. Lett. 2010, 13, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; TEAM, C. Nlme: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1-152; Springer: New York, NY, USA, 2021. [Google Scholar]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.5-7, 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 July 2022).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Warton, D.I.; Duursma, R.A.; Falster, D.S.; Taskinen, S. smatr 3—An R package for estimation and inference about allometric lines. Methods Ecol. Evol. 2012, 3, 257–259. [Google Scholar] [CrossRef]
- Calvo-Garrido, C.; Songy, A.; Marmol, A.; Roda, R.; Clément, C.; Fontaine, F. Description of the relationship between trunk disease expression and meteorological conditions, irrigation and physiological response in Chardonnay grapevines. OENO One 2021, 55, 97–113. [Google Scholar] [CrossRef]
- Downton, W.; Grant, W.; Loveys, B. Diurnal changes in the photosynthesis of field-grown grape vines. New Phytol. 1987, 105, 71–80. [Google Scholar] [CrossRef]
- Ghaderi, N.; Talaie, A.; Ebadi, A.; Lessani, H. The physiological response of three Iranian grape cultivars to progressive drought stress. SID 2011, 13, 601–610. [Google Scholar]
- Moutinho-Pereira, J.; Gonçalves, B.; Bacelar, E.; Cunha, J.B.; Countinho, J.; Correira, C. Effects of elevated CO2 on grapevine (Vitis vinifera L.): Physiological and yield attributes. Vitis-J. Grapevine Res. 2015, 48, 159. [Google Scholar]
- Pollastrini, M.; Di Stefano, V.; Ferretti, M.; Agati, G.; Grifoni, D.; Zipoli, G.; Orlandini, S.; Bussotti, F. Influence of different light intensity regimes on leaf features of Vitis vinifera L. in ultraviolet radiation filtered condition. Environ. Exp. Bot. 2011, 73, 108–115. [Google Scholar] [CrossRef]
- Poni, S.; Casalini, L.; Bernizzoni, F.; Civardi, S.; Intrieri, C. Effects of early defoliation on shoot photosynthesis, yield components, and grape composition. Am. J. Enol. Vitic. 2006, 57, 397–407. [Google Scholar]
- Prieto, J.A.; Louarn, G.; Peña, J.P.; Ojeda, H.; Simonneau, T.; Lebon, E. A leaf gas exchange model that accounts for intra-canopy variability by considering leaf nitrogen content and local acclimation to radiation in grapevine (Vitis vinifera L.). Plant Cell Environ. 2012, 35, 1313–1328. [Google Scholar] [CrossRef]
- Roig-Oliver, M.; Nadal, M.; Clemente-Moreno, M.J.; Bota, J.; Flexas, J. Cell wall components regulate photosynthesis and leaf water relations of Vitis vinifera cv. Grenache acclimated to contrasting environmental conditions. J. Plant Physiol. 2020, 244, 153084. [Google Scholar] [CrossRef]
- Salazar-Parra, C.; Aranjuelo, I.; Pascual, I.; Aguirreolea, J.; Sánchez-Díaz, M.; Irigoyen, J.J.; Araus, J.L.; Morales, F. Is vegetative area, photosynthesis, or grape C uploading involved in the climate change-related grape sugar/anthocyanin decoupling in Tempranillo? Photosynth. Res. 2018, 138, 115–128. [Google Scholar] [CrossRef]
- Schulze-Sylvester, M.; Corronca, J.A.; Paris, C.I. Vine mealybugs disrupt biomass allocation in grapevine. OENO One 2021, 55, 93–103. [Google Scholar] [CrossRef]
- Verdenal, T.; Dienes-Nagy, Á.; Spangenberg, J.E.; Zufferey, V.; Spring, J.-L.; Viret, O.; Marin-Carbonne, J.; van Leeuwen, C. Understanding and managing nitrogen nutrition in grapevine: A review. OENO One 2021, 55, 1–43. [Google Scholar] [CrossRef]
- Vrignon-Brenas, S.; Metay, A.; Leporatti, R.; Gharibi, S.; Fraga, A.; Dauzat, M.; Rolland, G.; Pellegrino, A. Gradual responses of grapevine yield components and carbon status to nitrogen supply. OENO One 2019, 53, 289–306. [Google Scholar] [CrossRef]
- Zufferey, V.; Spring, J.-L.; Verdenal, T.; Dienes, A.; Belcher, S.; Lorenzini, F.; Koestel, C.; Rösti, J.; Gindro, K.; Spangenberg, J. The influence of water stress on plant hydraulics, gas exchange, berry composition and quality of Pinot Noir wines in Switzerland. OENO One 2017, 51. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbó, M.; Flexas, J. Variability of water use efficiency in grapevines. Environ. Exp. Bot. 2014, 103, 148–157. [Google Scholar] [CrossRef]
- Bungau, S.; Behl, T.; Aleya, L.; Bourgeade, P.; Aloui-Sossé, B.; Purza, A.L.; Abid, A.; Samuel, A.D. Expatiating the impact of anthropogenic aspects and climatic factors on long-term soil monitoring and management. Environ. Sci. Pollut. Res. 2021, 28, 30528–30550. [Google Scholar] [CrossRef]
- Pettenuzzo, S.; Cappellin, L.; Grando, M.S.; Costantini, L. Phenotyping methods to assess heat stress resilience in grapevine. J. Exp. Bot. 2022, 73, 5128–5148. [Google Scholar] [CrossRef]
- Merrill, N.K.; De Cortázar-Atauri, I.G.; Parker, A.K.; Walker, M.A.; Wolkovich, E.M. Exploring grapevine phenology and high temperatures response under controlled conditions. Front. Environ. Sci. 2020, 8, 516527. [Google Scholar] [CrossRef]
- Montazeaud, G.; Violle, C.; Roumet, P.; Rocher, A.; Ecarnot, M.; Compan, F.; Maillet, G.; Fort, F.; Fréville, H. Multifaceted functional diversity for multifaceted crop yield: Towards ecological assembly rules for varietal mixtures. J. Appl. Ecol. 2020, 57, 2285–2295. [Google Scholar] [CrossRef]
- Schultz, H.; Stoll, M. Some critical issues in environmental physiology of grapevines: Future challenges and current limitations. Aust. J. Grape Wine Res. 2010, 16, 4–24. [Google Scholar] [CrossRef]
- Martin, A.R.; Mariani, R.O.; Cathline, K.A.; Duncan, M.; Paroshy, N.J.; Robertson, G. Soil compaction drives an intra-genotype leaf economics spectrum in wine grapes. Agriculture 2022, 12, 1675. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Cornelissen, J.H.C.; Falster, D.S.; Garnier, E.; Hikosaka, K.; Lamont, B.B.; Lee, W.; Oleksyn, J.; Osada, N.; et al. Assessing the generality of global leaf trait relationships. New Phytol. 2005, 166, 485–496. [Google Scholar] [CrossRef]
- Chen, L.-S.; Cheng, L. Carbon Assimilation and Carbohydrate Metabolism ofConcord’Grape (Vitis labrusca L.) Leaves in Response to Nitrogen Supply. J. Am. Soc. Hortic. Sci. 2003, 128, 754–760. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distribution Fitting | Descriptive Statistics | Variance Partitioning | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait Group | Trait | Normal | Log-Normal | Mean/ Median | SD/ MAD | Range | CV | Time | Variety | Row | Unexplained |
| Physiological | Amax (μmol CO2 m−2 s−1) | −240.4 | −261.8 | 13.2 | 3.52 | 3.0–20.1 | 26.7 | 0.000 | 0.273 | 0.041 | 0.686 |
| Amass (μmol CO2 g−1 s−1) | 129.6 | 115.5 | 0.19 | 0.06 | 0.04–0.34 | 30.1 | 0.108 | 0.378 | 0.036 | 0.478 | |
| gs (mol H2O m−2 s−1) | −0.9 | 44.2 | 0.188 | 0.13 | 0.012–0.83 | 97.3 | 0.075 | 0.064 | 0.129 | 0.732 | |
| WUE (mmol CO2 mol H2O−1) | −318.0 | −222.5 | 4.3 | 2.2 | 1.02–19.1 | 63.9 | 0.000 | 0.000 | 0.109 | 0.891 | |
| Morphological | Leaf dry mass (g) | −32.8 | −25.1 | 0.88 | 0.36 | 0.4–2.0 | 37.9 | 0.000 | 0.294 | 0.000 | 0.706 |
| Leaf area (cm2) | −464.7 | −462.1 | 125.6 | 37.5 | 52.6–241.7 | 32.5 | 0.150 | 0.355 | 0.000 | 0.495 | |
| LMA (g m−2) | −357.9 | −357.2 | 68.8 | 14.6 | 44.9–102.8 | 18.4 | 0.748 | 0.021 | 0.000 | 0.231 | |
| Chemical | Carbon (% mass) | −116.8 | −117.1 | 43.6 | 0.9 | 41.4–45.6 | 2.0 | 0.476 | 0.205 | 0.045 | 0.275 |
| Nitrogen (% mass) | −64.6 | −61.6 | 2.9 | 0.5 | 2.2–4.3 | 17.0 | 0.779 | 0.037 | 0.048 | 0.136 | |
| Trait Group | Trait | Growth Stage | Variety | Row | Stage * Variety | Stage * Row | Variety * Row | Stage * Variety * Row |
|---|---|---|---|---|---|---|---|---|
| Physiological | Amax | 3.18 (0.08) | 1.92 (0.119) | 0.64 (0.529) | 7.99 (<0.001) | 0.08 (0.92) | 1.04 (0.417) | 1.73 (0.11) |
| Amass | 18.49 (<0.001) | 6.46 (<0.001) | 1.17 (0.317) | 10.22 (<0.001) | 0.07 (0.933) | 0.94 (0.495) | 1.82 (0.091) | |
| log-gs | 6.95 (0.011) | 1.96 (0.112) | 2.1 (0.132) | 2.66 (0.041) | 1.61 (0.208) | 1.52 (0.17) | 1.38 (0.225) | |
| log-WUE | 0.05 (0.832) | 1.795 (0.146) | 0.773 (0.466) | 0.368 (0.83) | 0.92 (0.403) | 2.78 (0.011) | 0.45 (0.889) | |
| Morphological | log-Dry mass | 0.65 (0.425) | 8.17 (<0.001) | 1.01 (0.369) | 0.54 (0.705) | 0.02 (0.984) | 0.64 (0.744) | 0.43 (0.897) |
| log-Area | 18.72 (<0.001) | 12.39 (<0.001) | 1.17 (0.318) | 0.82 (0.52) | 0.12 (0.885) | 0.61 (0.77) | 0.44 (0.895) | |
| log-LMA | 146.87 (<0.001) | 2.26 (0.074) | 0.26 (0.775) | 1.38 (0.253) | 0.12 (0.885) | 1.23 (0.297) | 1.11 (0.372) | |
| Chemical | Leaf C | 86.13 (<0.001) | 9.26 (<0.001) | 0.07 (0.937) | 7.15 (<0.001) | 0.66 (0.521) | 2.02 (0.059) | 1.53 (0.168) |
| log-Leaf N | 261.85 (<0.001) | 4.2 (0.005) | 0.82 (0.45) | 4.79 (0.002) | 1.5 (0.232) | 2.76 (0.012) | 1.83 (0.089) | |
| Multivariate | PCA 1 | 374.9 (<0.001) | 2.26 (0.073) | 0.627 (0.537) | 8.477 (<0.001) | 0.561 (0.574) | 1.56 (0.156) | 1.72 (0.112) |
| PCA 2 | 0.252 (0.617) | 1.407 (0.243) | 0.904 (0.41) | 0.953 (0.44) | 1.118 (0.334) | 1.604 (0.143) | 1.119 (0.364) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macklin, S.C.; Mariani, R.O.; Young, E.N.; Kish, R.; Cathline, K.A.; Robertson, G.; Martin, A.R. Intraspecific Leaf Trait Variation across and within Five Common Wine Grape Varieties. Plants 2022, 11, 2792. https://doi.org/10.3390/plants11202792
Macklin SC, Mariani RO, Young EN, Kish R, Cathline KA, Robertson G, Martin AR. Intraspecific Leaf Trait Variation across and within Five Common Wine Grape Varieties. Plants. 2022; 11(20):2792. https://doi.org/10.3390/plants11202792
Chicago/Turabian StyleMacklin, Samantha C., Rachel O. Mariani, Emily N. Young, Rosalyn Kish, Kimberley A. Cathline, Gavin Robertson, and Adam R. Martin. 2022. "Intraspecific Leaf Trait Variation across and within Five Common Wine Grape Varieties" Plants 11, no. 20: 2792. https://doi.org/10.3390/plants11202792
APA StyleMacklin, S. C., Mariani, R. O., Young, E. N., Kish, R., Cathline, K. A., Robertson, G., & Martin, A. R. (2022). Intraspecific Leaf Trait Variation across and within Five Common Wine Grape Varieties. Plants, 11(20), 2792. https://doi.org/10.3390/plants11202792