

Assessing Impacts of Transgenic Plants on Soil Using Functional Indicators: Twenty Years of Research and Perspectives

Abstract
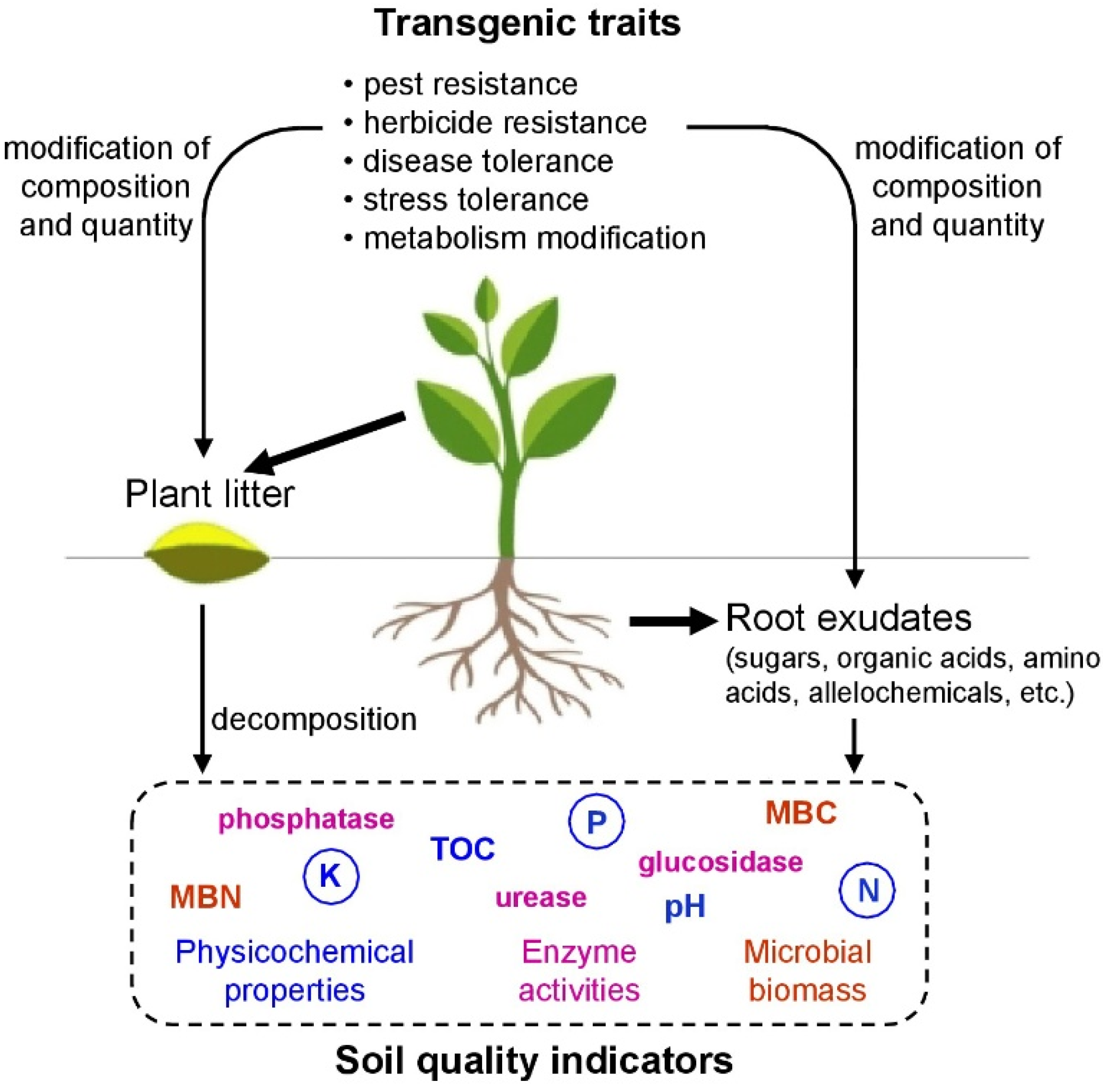
1. Introduction
2. Potential Risks of Transgenic Plants for Soil Ecosystems
3. Indicators of Soil Quality and Fertility
3.1. Soil Physicochemical Properties
3.2. Soil Enzyme Activity
3.3. Microbial Biomass
4. Effects of Transgenic Plants’ Cultivation
4.1. Insect-Resistant Transgenic Plants
4.2. Herbicide-Resistant Transgenic Plants

{kind=link}
| Species | Gene | Growth Conditions | Indicators | Additional Factors | References |
|---|---|---|---|---|---|
| oilseed rape | pat | greenhouse | MBN | growth stage | [72] |
| C, N, P, S | |||||
| wheat | epsps | field (4 years) | MBC | location | [73] |
| canola | DHA | crop rotation | |||
| maize | epsps | field (5 years) | MBC | herbicide | [74] |
| C | crop rotation | ||||
| soybean | ahas | field (3 years) | MBC, MBN | location | [75] |
| soybean | epsps | field (3 years) | MBC, MBN | location | [77] |
| C, P | herbicide | ||||
| soybean | epsps | field (8–9 years) | pH, org. matter, N, P, microelem., texture | location | [76] |
| MBC, MBN |
4.3. Disease-Tolerant Transgenic Plants
4.4. Stress-Tolerant Transgenic Plants
4.5. Transgenic Plants with Modified Metabolic Pathways
5. Factors Influencing the Interaction of Transgenic Plants with Soil
6. Effects of Transgenic Plant Residues
7. Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier; ISAAA Brief No. 55; ISAAA: Ithaca, NY, USA, 2019. [Google Scholar]
- Snow, A.A.; Palma, P.M. Commercialization of transgenic plants: Potential ecological risks. BioScience 1997, 47, 86–96. [Google Scholar] [CrossRef]
- Messeguer, J. Gene flow assessment in transgenic plants. Plant Cell Tissue Organ Cult. 2003, 73, 201–212. [Google Scholar] [CrossRef]
- Sisterson, M.S.; Carrière, Y.; Dennehy, T.J.; Tabashnik, B.E. Evolution of resistance to transgenic crops: Interactions between insect movement and field distribution. J. Econ. Entomol. 2006, 98, 1751–1762. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Meissle, M.; Bigler, F. Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nat. Biotechnol. 2006, 24, 63–71. [Google Scholar] [CrossRef]
- Bruinsma, M.; Kowalchuk, G.A.; van Veen, J.A. Effects of genetically modified plants on microbial communities and processes in soil. Biol. Fertil. Soils 2003, 37, 329–337. [Google Scholar] [CrossRef]
- Singh, A.K.; Dubey, S.K. 8—Transgenic Plants and Soil Microbes. In Current Developments in Biotechnology and Bioengineering; Dubey, S.K., Pandey, A., Sangwan, R.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 163–185. [Google Scholar] [CrossRef]
- Guan, Z.; Lu, S.; Huo, Y.; Guan, Z.-P.; Liu, B.; Wei, W. Do genetically modified plants affect adversely on soil microbial communities? Agric Ecosyst. Environ. 2016, 235, 289–305. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, L.; Wu, Z. Relationships among persistence of Bacillus thuringiensis and Cowpea trypsin inhibitor proteins, microbial properties and enzymatic activities in rhizosphere soil after repeated cultivation with transgenic cotton. Appl. Soil Ecol. 2012, 53, 23–30. [Google Scholar] [CrossRef]
- Chen, Z.; Wei, K.; Chen, L.; Wu, Z.; Luo, J.; Cui, J. Effects of the consecutive cultivation and periodic residue incorporation of Bacillus thuringiensis (Bt) cotton on soil microbe-mediated enzymatic properties. Agric Ecosyst. Environ. 2017, 239, 154–160. [Google Scholar] [CrossRef]
- Wei, H.W.; Movahedi, A.; Liu, G.; Kiani-Pouya, A.; Rasouli, F.; Yu, C.; Chen, Y.; Zhong, F.; Zhang, J. Effects of field-grown transgenic Cry1Ah1 poplar on the rhizosphere microbiome. Res. Sq. 2022, preprint. [Google Scholar] [CrossRef]
- Luo, J.-Y.; Zhang, S.; Zhu, X.-Z.; Lu, L.-M.; Wang, C.-Y.; Li, C.-H.; Cui, J.-J.; Zhou, Z.-G. Effects of soil salinity on rhizosphere soil microbes in transgenic Bt cotton fields. J. Integr. Agric. 2017, 16, 1624–1633. [Google Scholar] [CrossRef]
- Haichar, F.Z.; Marol, C.; Berge, O.; Rangel-Castro, J.I.; Prosser, J.I.; Balesdent, J.; Achouak, W. Plant host habitat and root exudates shape soil bacterial community structure. ISME J. 2008, 2, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Guyonnet, J.P.; Cantarel, A.A.M.; Simon, L.; Haichar, F.Z. Root exudation rate as functional trait involved in plant nutrient-use strategy classification. Ecol. Evol. 2018, 8, 8573–8581. [Google Scholar] [CrossRef] [PubMed]
- Hannula, S.E.; de Boer, W.; van Veen, J.A. Do genetic modifications in crops affect soil fungi? A review. Biol. Fertil. Soils 2014, 50, 433–446. [Google Scholar] [CrossRef]
- Mandal, A.; Sarkar, B.; Owens, G.; Thakur, J.K.; Manna, M.C.; Niazi, N.K.; Jayaraman, S.; Patra, A.K. Impact of genetically modified crops on rhizosphere microorganisms and processes: A review focusing on Bt cotton. Appl. Soil Ecol. 2020, 148, 103492. [Google Scholar] [CrossRef]
- Devare, M.; Londono-R, L.M.; Thies, J.E. Neither transgenic Bt maize (MON863) nor tefluthrin insecticide adversely affect soil microbial activity or biomass: A 3-year field analysis. Soil Biol. Biochem. 2007, 39, 2038–2047. [Google Scholar] [CrossRef]
- Dunfield, K.E.; Germida, J.J. Impact of genetically modified crops on soil- and plant-associated microbial communities. J. Environ. Qual. 2004, 33, 806–815. [Google Scholar] [CrossRef]
- Larson, W.E.; Pierce, F.J. Conservation and enhancement of soil quality. In Evaluation for Sustainable Land Management in the Developing World; Vol. 2: Technical Papers; International Board for Research and Management: Bangkok, Thailand, 1991; IBSRAM Proceedings No. 12; pp. 175–203. [Google Scholar]
- Beule, L.; Vaupel, A.; Moran-Rodas, V.E. Abundance, diversity, and function of soil microorganisms in temperate alley-cropping agroforestry systems: A review. Microorganisms 2022, 10, 616. [Google Scholar] [CrossRef]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Singh, B.K.; Quince, C.; Macdonald, C.A.; Khachane, A.; Thomas, N.; Al-Soud, W.A.; Sørensen, S.J.; He, Z.; White, D.; Sinclair, A.; et al. Loss of microbial diversity in soils is coincident with reductions in some specialized functions. Environ. Microbiol. 2014, 16, 2408–2420. [Google Scholar] [CrossRef]
- Escalas, A.; Hale, L.; Voordeckers, J.W.; Yang, Y.; Firestone, M.K.; Alvarez-Cohen, L.; Zhou, J. Microbial functional diversity: From concepts to applications. Ecol. Evol. 2019, 9, 12000–12016. [Google Scholar] [CrossRef]
- Peter, H.; Beier, S.; Bertilsson, S.; Lindstrom, E.S.; Langenheder, S.; Tranvik, L.J. Function-specific response to depletion of microbial diversity. ISME J. 2011, 5, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Plante, C.J. Defining disturbance for microbial ecology. Microb. Ecol. 2017, 74, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ma, K.; Huang, Y.; Yao, Z.; Chu, C. Stable soil microbial functional structure responding to biodiversity loss based on metagenomic evidences. Front. Microbiol. 2021, 12, 716764. [Google Scholar] [CrossRef]
- Louca, S.; Jacques, S.M.S.; Pires, A.P.F.; Leal, J.S.; Srivastava, D.S.; Parfrey, L.W.; Farjalla, V.F.; Doebeli, M. High taxonomic variability despite stable functional structure across microbial communities. Nat. Ecol. Evol. 2016, 1, 15. [Google Scholar] [CrossRef]
- Krause, S.M.B.; Näther, A.; Cortes, V.O.; Mullins, E.; Kessel, G.J.T.; Lotz, L.A.P.; Tebbe, C.C. No tangible effects of field-grown cisgenic potatoes on soil microbial communities. Front. Bioeng. Biotechnol. 2020, 8, 603145. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef]
- Lee, Y.H.; Ahn, B.K.; Sonn, Y.K. Effects of electrical conductivity on the soil microbial community in a controlled horticultural land for strawberry cultivation. Korean J. Soil Sci. Fert. 2011, 44, 830–835. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.; Xue, S.; Song, Z. Rhizosphere soil microbial activity under different vegetation types on the Loess Plateau, China. Geoderma 2011, 161, 115–125. [Google Scholar] [CrossRef]
- Ahamd, M.; Abbasi, W.M.; Jamil, M.; Iqbal, M.; Hussain, A.; Akhtar, M.F.; Nazli, F. Comparison of rhizosphere properties as affected by different Bt- and non-Bt-cotton (Gossypium hirsutum L.) genotypes and fertilization. Environ. Monit. Assess. 2017, 189, 278. [Google Scholar] [CrossRef]
- Song, Y.; Song, C.; Yang, G.; Miao, Y.; Wang, J.; Guo, Y. Changes in labile organic carbon fractions and soil enzyme activities after marshland reclamation and restoration in the Sanjiang Plain in Northeast China. Environ. Manag. 2012, 50, 418–426. [Google Scholar] [CrossRef]
- Hinojosa, M.B.; Carreira, J.A.; Garcıa-Ruız, R.; Dick, R.P. Soil moisture pre-treatment effects on enzyme activities as indicators of heavy metal-contaminated and reclaimed soils. Soil Biol. Biochem. 2004, 36, 1559–1568. [Google Scholar] [CrossRef]
- Lino, I.A.N.; Santos, V.M.; Escobar, I.E.C.; Silva, D.K.A.; Maia, L.C. Soil enzymatic activity in Eucalyptus grandis plantations of different ages. Land Degrad. Dev. 2016, 27, 77–82. [Google Scholar] [CrossRef]
- Brzezińska, M.; Lipiec, J.; Frąc, M.; Oszust, K.; Szarlip, P.; Turski, M. Quantitative interactions between total and specific enzyme activities and C and N contents in earthworm-affected pear orchard soil. Land Degrad. Devel. 2018, 29, 3379–3389. [Google Scholar] [CrossRef]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leiros, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Garcia, C.; Hernandez, T.; Costa, F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plant Anal. 1997, 28, 123–134. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Velmourougane, K.; Sahu, A. Impact of transgenic cottons expressing cry1Ac on soil biological attributes. Plant Soil Environ. 2013, 3, 108–114. [Google Scholar] [CrossRef]
- Jan, M.T.; Roberts, P.; Tonheim, S.K.; Jones, D.L. Protein breakdown represents a major bottleneck in nitrogen cycling in grassland soils. Soil Biol. Biochem. 2009, 41, 2272–2282. [Google Scholar] [CrossRef]
- Kramer, S.; Green, D.M. Acid and alkaline phosphatase dynamics and their relationship to soil microclimate in a semiarid woodland. Soil Biol. Biochem. 2000, 32, 179–188. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Claassen, N. Organic phosphorus compounds as a phosphorus source for higher plants through the activity of phosphatase produced by plant roots and microorganisms. Biol. Fertil. Soils 1988, 5, 308–312. [Google Scholar] [CrossRef]
- Kertesz, M.A.; Mirleau, P. The role of soil microbes in plant sulphur nutrition. J. Exp. Bot. 2004, 55, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Frankenberger, W.T., Jr.; Johanson, J.B. Method of measuring invertase activity in soils. PIant Soil 1983, 74, 301–311. [Google Scholar] [CrossRef]
- Gander, L.K.; Hendricks, C.W.; Doyle, J.D. Interferences, limitations and an improvement in the extraction and assessment of cellulase activity in soil. Soil Biol. Biochem. 1994, 26, 65–73. [Google Scholar] [CrossRef]
- Floch, C.; Alarcon-Gutiérrez, E.; Criquet, S. ABTS assay of phenol oxidase activity in soil. J. Microbiol. Methods 2007, 71, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Trasar-Cepeda, C.; Camina, F.; Leirós, C.; Gil-Sotres, F. An improved method to measure catalase activity in soils. Soil Biol. Biochem. 1999, 31, 483–485. [Google Scholar] [CrossRef]
- Singh, J.S.; Raghubanshi, A.S.; Singh, R.S.; Srivastava, S.C. Microbial biomass acts as a source of plant nutrients in dry tropical forest and savanna. Nature 1989, 338, 499–500. [Google Scholar] [CrossRef]
- Sparling, G.P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Aust. J. Soil Res. 1992, 30, 195–207. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, X.; Chen, X.; Lin, Q.; Li, G. Seasonal changes of soil microbial C, N, P and associated nutrient dynamics in a semiarid grassland of north China. Appl. Soil Ecol. 2018, 128, 89–97. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R. Short-term dynamics of soil carbon, microbial biomass, and soil enzyme activities as compared to longer-term effects of tillage in irrigated row crops. Biol. Fertil. Soils 2009, 46, 65–72. [Google Scholar] [CrossRef]
- Lee, Z.L.; Bu, N.S.; Cui, J.; Chen, X.P.; Xiao, M.Q.; Wang, F.; Song, Z.P.; Fang, C.M. Effects of long-term cultivation of transgenic Bt rice (Kefeng-6) on soil microbial functioning and C cycling. Sci. Rep. 2017, 7, 4647. [Google Scholar] [CrossRef]
- Devare, M.H.; Jones, C.M.; Thies, J.E. Effect of Cry3Bb transgenic corn and tefluthrin on the soil microbial community: Biomass, activity, and diversity. J. Environ. Qual. 2004, 33, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Icoz, I.; Saxena, D.; Andow, D.A.; Zwahlen, C.; Stotzky, G. Microbial populations and enzyme activities in soil in situ under transgenic corn expressing cry proteins from Bacillus thuringiensis. J. Environ. Qual. 2008, 37, 647–662. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lu, H.H.; Wu, W.X.; Wei, Q.K.; Chen, Y.X.; Thies, J.E. Transgenic Bt rice does not affect enzyme activities and microbial composition in the rhizosphere during crop development. Soil Biol. Biochem. 2008, 40, 475–486. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Pampulha, M.E.; Bennett, J.P. A two-year field study with transgenic Bacillus thuringiensis maize: Effects on soil microorganisms. Sci. Total Environ. 2008, 405, 351–357. [Google Scholar] [CrossRef]
- Sarkar, B.; Patra, A.K.; Purakayastha, T.J. Transgenic Bt-cotton affects enzyme activity and nutrient availability in a sub-tropical inceptisol. J. Agron. Crop Sci. 2008, 194, 289–296. [Google Scholar] [CrossRef]
- Sarkar, B.; Patra, A.K.; Purakayastha, T.; Megharaj, M. Assessment of biological and biochemical indicators in soil under transgenic Bt and non-Bt cotton crop in a sub-tropical environment. Environ. Monit. Assess. 2009, 156, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Kravchenko, A.N.; Hao, X.; Robertson, G.P. Seven years of continuously planted Bt corn did not affect mineralizable and total soil C and total N in surface soil. Plant Soil 2009, 318, 269–274. [Google Scholar] [CrossRef]
- Mina, U.; Chaudhary, A.; Kamra, A. Effect of Bt cotton on enzymes activity and microorganisms in rhizosphere. J. Agric. Sci. 2011, 3, 96–104. [Google Scholar] [CrossRef]
- Wei, M.; Tan, F.; Zhu, H.; Cheng, K.; Wu, X.; Wang, J.; Zhao, K.; Tang, X. Impact of Bt-transgenic rice (SHK601) on soil ecosystems in the rhizosphere during crop development. Plant Soil Environ. 2012, 58, 217–223. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, M.; Ding, G. Effect of transgenic Bt cotton on bioactivities and nutrients in rhizosphere soil. Commun. Soil Sci. Plant Anal. 2012, 43, 689–700. [Google Scholar] [CrossRef]
- Fließbach, A.; Messmer, M.; Nietlispach, B.; Infante, V.; Mäder, P. Effects of conventionally bred and Bacillus thuringiensis (Bt) maize varieties on soil microbial biomass and activity. Biol. Fertil. Soils 2012, 48, 315–324. [Google Scholar] [CrossRef]
- Fließbach, A.; Nietlispach, B.; Messmer, M.; Rodriguez-Romero, A.-S.; Maeder, P. Microbial response of soils with organic and conventional management history to the cultivation of Bacillus thuringiensis (Bt)-maize under climate chamber conditions. Biol. Fertil. Soils 2013, 49, 829–837. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Blackshaw, R.E. Soil microbial properties in Bt (Bacillus thuringiensis) corn cropping systems. Appl. Soil Ecol. 2013, 63, 127–133. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Xie, M.; Li, C.Y.; Wu, G.; Peng, D.L. Impacts of the transgenic CrylAc and CpTI insect-resistant cotton SGK321 on selected soil enzyme activities in the rhizosphere. Plant Soil Environ. 2014, 60, 401–406. [Google Scholar] [CrossRef]
- Zhou, D.; Xu, L.; Gao, S.; Guo, J.; Luo, J.; You, Q.; Que, Y. Cry1Ac transgenic sugarcane does not affect the diversity of microbial communities and has no significant effect on enzyme activities in rhizosphere soil within one crop season. Front. Plant Sci. 2016, 7, 265. [Google Scholar] [CrossRef]
- van Wyk, D.A.B.; Adeleke, R.; Rhode, O.H.J.; Bezuidenhout, C.C.; Mienie, C. Ecological guild and enzyme activities of rhizosphere soil microbial communities associated with Bt-maize cultivation under field conditions in North West Province of South Africa. J. Basic Microbiol. 2017, 57, 781–792. [Google Scholar] [CrossRef]
- Zuo, L.H.; Yang, R.L.; Zhen, Z.X.; Liu, J.X.; Huang, L.S.; Yang, M.S. A 5-year field study showed no apparent effect of the Bt transgenic 741 poplar on the arthropod community and soil bacterial diversity. Sci. Rep. 2018, 8, 1956. [Google Scholar] [CrossRef]
- Mandal, A.; Thakur, J.K.; Sahu, A.; Manna, M.C.; Rao, A.S.; Sarkar, B.; Patra, A.K. Effects of Bt-cotton on biological properties of Vertisols in central India. Arch. Agron. Soil Sci. 2019, 65, 670–685. [Google Scholar] [CrossRef]
- Sessitsch, A.; Gyamfi, S.; Tscherko, D.; Gerzabek, M.H.; Kandeler, E. Activity of microorganisms in the rhizosphere of herbicide treated and untreater transgenic glufosinate-tolerant and wildtype oilseed rape grown in containment. Plant Soil 2005, 266, 105–116. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Hanson, K.G.; Harker, K.N.; Clayton, G.W.; Blackshaw, R.E.; Donovan, J.T.; Johnson, E.N.; Gan, Y.; Irvine, R.B.; Monreal, M.A. Soil microbial biomass, functional diversity and enzyme activity in glyphosate-resistant wheat–canola rotations under low-disturbance direct seeding and conventional tillage. Soil Biol. Biochem. 2007, 39, 1418–1427. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Blackshaw, R.E. Soil microbiology in glyphosate-resistant corn cropping systems. Agron. J. 2012, 104, 1041–1048. [Google Scholar] [CrossRef]
- Souza, R.A.; Babujia, L.C.; Silva, A.P.; Guimarães, M.F.; Arias, C.A.; Hungria, M. Impact of the ahas transgene and of herbicides associated with the soybean crop on soil microbial community. Transgenic Res. 2013, 22, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Babujia, L.C.; Silva, A.P.; Nakatani, A.S.; Cantao, M.E.; Vasconcelos, A.T.R.; Visentainer, J.V.; Hungria, M. Impact of long-term cropping of glyphosate-resistant transgenic soybean [Glycine max (L.) Merr.] on soil microbiome. Transgenic Res. 2016, 25, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, A.S.; Fernandes, M.F.; Souza, R.A.; Silva, A.P.; Reis-Junior, F.B.; Mendes, I.C.; Hungria, M. Effects of the glyphosate-resistance gene and of herbicides applied to the soybean crop on soil microbial biomass and enzymes. Field Crops Res. 2014, 162, 20–29. [Google Scholar] [CrossRef]
- Wei, X.D.; Zou, H.L.; Chu, L.M.; Liao, B.; Ye, C.M.; Lan, C.Y. Field released transgenic papaya affects microbial communities and enzyme activities in soil. Plant Soil 2006, 285, 347–358. [Google Scholar] [CrossRef]
- Hsieh, Y.-T.; Pan, T.-M. Influence of planting papaya ringspot virus resistant transgenic papaya on soil microbial biodiversity. J. Agric. Food Chem. 2006, 54, 130–137. [Google Scholar] [CrossRef]
- Stefani, F.O.P.; Tanguay, P.; Pelletier, G.; Piche, Y.; Hamelin, R.C. Impact of endochitinase-transformed white spruce on soil fungal biomass and ectendomycorrhizal symbiosis. Appl. Environ. Microbiol. 2010, 76, 2607–2614. [Google Scholar] [CrossRef]
- Wang, B.; Shen, H.; Yang, X.; Guo, T.; Zhang, B.; Yan, W. Effects of chitinase-transgenic (McChit1) tobacco on the rhizospheric microflora and enzyme activities of the purple soil. Plant Soil Environ. 2013, 59, 241–246. [Google Scholar] [CrossRef]
- Wu, J.; Yu, M.; Xu, J.; Du, J.; Ji, F.; Dong, F.; Li, X.; Shi, J. Impact of transgenic wheat with wheat yellow mosaic virus resistance on microbial community diversity and enzyme activity in rhizosphere soil. PLoS ONE 2014, 9, e98394. [Google Scholar] [CrossRef]
- Bezirganoglu, I.; Uysal, P. Impact of transgenic AFPCHI (Cucumis melo L. Silver Light) fungal resistance melon on soil microbial communities and enzyme activities. J. Plant Biotechnol. 2017, 44, 156–163. [Google Scholar] [CrossRef]
- Khan, M.S.; Sadat, S.U.; Jan, A.; Munir, I. Impact of transgenic Brassica napus harboring the antifungal synthetic chitinase (NiC) gene on rhizosphere microbial diversity and enzyme activities. Front. Plant Sci. 2017, 8, 1307. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.-I.; Oh, Y.-J.; Ahn, J.-H.; Kang, H.-J.; Cho, W.-S.; Cho, Y.; Lee, B.K. Effects of disease resistant genetically modified rice on soil microbial community structure according to growth stage. Korean J. Environ. Agric. 2019, 38, 185–196. [Google Scholar] [CrossRef]
- Tesfaye, M.; Temple, S.J.; Allan, D.L.; Vance, C.P.; Samac, D.A. Over-expression of malate dehydrogenase in transgenic alfalfa enhances organic acid synthesis and confers tolerance to aluminum. Plant Physiol. 2001, 127, 1836–1844. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, M.; Dufault, N.S.; Dornbusch, M.R.; Deborah, L.; Allan, D.L.; Vance, C.P.; Samac, D.A. Influence of enhanced malate dehydrogenase expression by alfalfa on diversity of rhizobacteria and soil nutrient availability. Soil Biol. Biochem. 2003, 35, 1103–1113. [Google Scholar] [CrossRef]
- Mimura, M.; Lelmen, K.E.; Shimazaki, T.; Kikuchi, A.; Watanabe, K.N. Impact of environmental stress-tolerant transgenic potato on genotypic diversity of microbial communities and soil enzyme activities under stress conditions. Microbes Environ. 2008, 23, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, V.; Dang, H.Q.; Tran, N.Q.; Mishra, A.; Chauhan, P.S.; Gill, S.S.; Nautiyal, C.S.; Tuteja, N. Impact of salinity-tolerant MCM6 transgenic tobacco on soil enzymatic activities and the functional diversity of rhizosphere microbial communities. Res. Microbiol. 2012, 163, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, R.K.; Tuteja, N. Effect of salinity tolerant PDH45 transgenic rice on physicochemical properties, enzymatic activities and microbial communities of rhizosphere soils. Plant Signal. Behav. 2013, 8, e24950. [Google Scholar] [CrossRef][Green Version]
- Bai, X.; Zeng, X.; Huang, S.; Liang, J.; Dong, L.; Wei, Y.; Li, Y.; Qu, J.; Wang, Z. Marginal impact of cropping BADH transgenic maize BZ-136 on chemical property, enzyme activity, and bacterial community diversity of rhizosphere soil. Plant Soil 2019, 436, 527–541. [Google Scholar] [CrossRef]
- Tian, W.-H.; Yi, X.-L.; Liu, S.-S.; Zhou, C.; Wang, A.-Y. Effect of transgenic cotton continuous cropping on soil bacterial community. Ann. Microbiol. 2020, 70, 61. [Google Scholar] [CrossRef]
- Zeng, X.; Pei, T.; Song, Y.; Guo, P.; Zhang, H.; Li, X.; Li, H.; Di, H.; Wang, Z.A. Three-year plant study of salt-tolerant transgenic maize showed no effects on soil enzyme activity and nematode community. Life 2022, 12, 412. [Google Scholar] [CrossRef]
- Li, Q.; Song, J.; Peng, S.; Wang, J.P.; Qu, G.Z.; Sederoff, R.R.; Chiang, V.L. Plant biotechnology for lignocellulosic biofuel production. Plant Biotechnol. J. 2014, 12, 1174–1192. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, V.G.; Shestibratov, K.A. Genetic engineering of lignin biosynthesis in trees: Compromise between wood properties and plant viability. Russ. J. Plant Physiol. 2021, 68, 596–612. [Google Scholar] [CrossRef]
- Motavalli, P.P.; Kremer, R.J.; Fang, M.; Means, N.E. Impact of genetically modified crops and their management on soil microbially mediated plant nutrient transformations. J. Environ. Qual. 2004, 33, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Kolseth, A.-K.; D’Hertefeldt, T.; Emmerich, M.; Forabosco, F.; Marklund, S.; Cheeke, T.E.; Hallin, S.; Weih, M. Influence of genetically modified organisms on agro-ecosystem processes. Agric. Ecosyst. Environ. 2015, 214, 96–106. [Google Scholar] [CrossRef]
- Pilate, G.; Guiney, E.; Holt, K.; Petit-Conil, M.; Lapierre, C.; Leplé, J.-C.; Pollet, B.; Mila, I.; Webster., E.A.; Marstorp, H.G.; et al. Field and pulping performances of transgenic trees with altered lignification. Nat. Biotechnol. 2002, 20, 607–612. [Google Scholar] [CrossRef]
- Tilston, E.L.; Halpin, C.; Hopkins, D.W. Simultaneous down-regulation of enzymes in the phenylpropanoid pathway of plants has aggregated effects on rhizosphere microbial communities. Biol. Fertil. Soils 2014, 50, 455–463. [Google Scholar] [CrossRef]
- Hannula, S.E.; de Boer, W.; van Veen, J. A 3-Year study reveals that plant growth stage, season and field site affect soil fungal communities while cultivar and GM-trait have minor effects. PLoS ONE 2012, 7, e33819. [Google Scholar] [CrossRef]
- Lahl, K.; Unger, C.; Emmerling, C.; Broer, I.; Thiele-Bruhn, S. Response of soil microorganisms and enzyme activities on the decomposition of transgenic cyanophycin-producing potatoes during overwintering in soil. Eur. J. Soil Biol. 2012, 53, 1–10. [Google Scholar] [CrossRef]
- Li, P.; Dong, J.; Yang, S.; Bai, L.; Wang, J.; Wu, G.; Wu, X.; Yao, Q.; Tang, X. Impact of β-carotene transgenic rice with four synthetic genes on rhizosphere enzyme activities and bacterial communities at different growth stages. Eur. J. Soil Biol. 2014, 65, 40–46. [Google Scholar] [CrossRef]
- Liang, J.; Sun, S.; Ji, J.; Wu, H.; Meng, F.; Zhang, M.; Zheng, X.; Wu, C.; Zhang, Z. Comparison of the rhizosphere bacterial communities of zigongdongdou soybean and a high-methionine transgenic line of this cultivar. PLoS ONE 2014, 9, e103343. [Google Scholar] [CrossRef]
- Brolsma, K.M.; Vonk, J.A.; Hoffland, E.; Mulder, C.; de Goede, R.G.M. Effects of GM potato Modena on soil microbial activity and litter decomposition fall within the range of effects found for two conventional cultivars. Biol. Fertil. Soils 2015, 51, 913–922. [Google Scholar] [CrossRef]
- Sohn, S.-I.; Oh, Y.-J.; Kim, B.-Y.; Kweon, S.-J.; Cho, H.-S.; Ryu, T.-H. Effect of genetically modified rice producing resveratrol on the soil microbial communities. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 795–805. [Google Scholar] [CrossRef]
- Debruyn, J.M.; Bevard, D.A.; Essington, M.E.; Mcknight, J.Y.; Schaeffer, S.M.; Baxter, H.L.; Mazarei, M.; Mann, D.G.J.; Dixon, R.A.; Chen, F.; et al. Field-grown transgenic switchgrass (Panicum virgatum L.) with altered lignin does not affect soil chemistry, microbiology, and carbon storage potential. GCB Bioenergy 2017, 9, 1100–1109. [Google Scholar] [CrossRef]
- Xu, S.; Ottinger, S.L.; Schaeffer, S.M.; DeBruyn, J.M.; Stewart, C.N., Jr.; Mazarei, M.; Jagadamma, S. Effects of field-grown transgenic switchgrass carbon inputs on soil organic carbon cycling. PeerJ. 2019, 16, e7887. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, L.; Ren, M.; Li, J.; Guan, X.; Tao, J. Comparison of genetically modified insect-resistant maize and non-transgenic maize revealed changes in soil metabolomes but not in rhizosphere bacterial community. GM Crops Food 2022, 13, 1–14. [Google Scholar] [CrossRef]
- Lakshmanan, V. Root microbiome assemblage is modulated by plant host factors. In Plant Microbe Interactions; Bais, H., Sherrier, J., Eds.; Academic Press Ltd-Elsevier Science Ltd.: London, UK, 2015; pp. 57–79. [Google Scholar]
- Ng, E.L.; Patti, A.F.; Rose, M.T.; Schefe, C.R.; Wilkinson, K.; Cavagnaro, T.R. Functional stoichiometry of soil microbial communities after amendment with stabilized organic matter. Soil Biol. Biochem. 2014, 7, 170–178. [Google Scholar] [CrossRef]
- Zi, H.; Hu, L.; Wang, C. Differentiate responses of soil microbial community and enzyme activities to nitrogen and phosphorus addition rates in an alpine meadow. Front. Plant Sci. 2022, 13, 829381. [Google Scholar] [CrossRef]
- Choi, Y.I.; Noh, E.W.; Choi, K.S. Low level expression of prokaryotic tzs gene enhances growth performance of transgenic poplars. Trees 2009, 23, 741–750. [Google Scholar] [CrossRef]
- Nam, K.J.; Kim, D.Y.; Nam, K.-H.; Pack, I.S.; Park, J.H.; Jeong, S.-C.; Choi, Y.I.; Noh, E.W.; Kim, C.-G. Effects of transgenic poplars expressing increased levels of cellular cytokinin on rhizosphere microbial communities. Eur. J. Soil Biol. 2014, 63, 78–82. [Google Scholar] [CrossRef]
- Saxena, D.; Stotzky, G. Bt corn has a higher lignin content than non-Bt corn. Am. J. Bot. 2001, 88, 1704–1706. [Google Scholar] [CrossRef]
- Wu, W.-X.; Ye, Q.-F.; Min, H.; Duan, X.-J.; Jin, W.-M. Bt-transgenic rice straw affects the culturable microbiota and dehydrogenase and phosphatase activities in a flooded paddy soil. Soil Biol. Biochem. 2004, 36, 289–295. [Google Scholar] [CrossRef]
- Huang, S.; Zeng, X.; Wei, Y.; Bai, X.; Jin, Z.; Zhang, M.; Wang, Z.; Wang, H.; Qu, J.; Di, H. Decomposition of betaine aldehyde dehydrogenase transgenic maize straw and its effects on soil microbial biomass and microbiota diversity. Appl. Soil Ecol. 2020, 153, 103582. [Google Scholar] [CrossRef]
- Hannula, S.E.; de Boer, W.; Baldrian, P.; van Veen, J.A. Effect of genetic modification of potato starch on decomposition of leaves and tubers and on fungal decomposer communities. Soil Biol. Biochem. 2013, 58, 88–98. [Google Scholar] [CrossRef]
- Hopkins, D.W.; Webster, E.A.; Chudek, J.A.; Halpin, C. Decomposition of stems from tobacco plants with genetic modifications to lignin biosynthesis. Soil Biol. Biochem. 2001, 33, 1455–1462. [Google Scholar] [CrossRef]
- Tilston, E.L.; Halpin, C.; Hopkins, D.W. Genetic modifications to lignin biosynthesis in field-grown poplar trees have inconsistent effects on the rate of woody trunk decomposition. Soil Biol. Biochem. 2004, 36, 1903–1906. [Google Scholar] [CrossRef]
- Deacon, L.J.; Pryce-Miller, E.J.; Frankland, J.C.; Bainbridge, B.W.; Moore, P.D.; Robinson, C.H. Diversity and function of decomposer fungi from a grassland soil. Soil Biol. Biochem. 2006, 38, 7–20. [Google Scholar] [CrossRef]
- Vauramo, S.; Pasonen, H.-L.; Pappinen, A.; Setala, H. Decomposition of leaf litter from chitinase transgenic silver birch (Betula pendula) and effects on decomposer populations in a field trial. Appl. Soil Ecol. 2006, 32, 338–349. [Google Scholar] [CrossRef]
- Qin, W.; Hu, C.; Oenema, O. Soil mulching significantly enhances yields and water and nitrogen use efficiencies of maize and wheat: A meta-analysis. Sci. Rep. 2015, 5, 16210. [Google Scholar] [CrossRef]
- Marral, M.W.R.; Khan, M.B.; Ahmad, F.; Farooq, S.; Hussain, M. The influence of transgenic (Bt) and nontransgenic (non-Bt) cotton mulches on weed dynamics, soil properties and productivity of different winter crops. PLoS ONE 2020, 15, e0238716. [Google Scholar] [CrossRef]
- Dellero, Y. Manipulating amino acid metabolism to improve crop nitrogen use efficiency for a sustainable agriculture. Front. Plant Sci. 2020, 11, 602548. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Popova, A.A.; Shestibratov, K.A. Genetic engineering and genome editing for improving nitrogen use efficiency in plants. Cells 2021, 10, 3303. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.; Ceasar, S.A.; Palmer, A.J.; Paterson, J.B.; Qi, W.; Muench, S.P.; Baldwin, S.A. Replace, reuse, recycle: Improving the sustainable use of phosphorus by plants. J. Exp. Bot. 2015, 66, 3523–3540. [Google Scholar] [CrossRef] [PubMed]
- Teng, W.; He, X.; Tong, Y.-P. Transgenic approaches for improving use efficiency of nitrogen, phosphorus and potassium in crops. J. Integr. Agric. 2017, 16, 2657–2673. [Google Scholar] [CrossRef]
- Iqrar, S.; Ashrafi, K.; Kiran, U.; Fatima, S.; Kamaluddin; Abdin, M.Z. Chapter seven—Transgenic plants with improved nutrient use efficiency. In Transgenic Technology Based Value Addition in Plant Biotechnology; Kiran, U., Abdin, M.Z., Kamaluddin, Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 157–185. [Google Scholar] [CrossRef]
- Singer, S.D.; Soolanayakanahally, R.Y.; Foroud, N.A.; Kroeber, R. Biotechnological strategies for improved photosynthesis in a future of elevated atmospheric CO2. Planta 2020, 251, 24. [Google Scholar] [CrossRef]
- Araus, J.L.; Sanchez-Bragado, R.; Vicente, R. Improving crop yield and resilience through optimization of photosynthesis: Panacea or pipe dream? J. Exp. Bot. 2021, 72, 3936–3955. [Google Scholar] [CrossRef]
- Weir, T.L.; Park, S.W.; Vivanco, J.M. Biochemical and physiological mechanisms mediated by allelochemicals. Curr. Opin. Plant Biol. 2004, 7, 472–479. [Google Scholar] [CrossRef]
- Schmutterer, H.; Singh, R.P. List of insect pests susceptible to neem products. In The Neem Tree: Azadirachta Indica A. Juss and Other Meliacious Plants—Sources of Unique Natural Products for Integrated Pest Management, Medicine, Industry and Other Purposes, 2nd ed.; Schmutterer, H., Ed.; Neem Foundation: Mumbai, India, 2002; pp. 411–456. [Google Scholar]
- Gopal, M.; Gupta, A.V.; Arunachalam, V.; Magu, S.P. Impact of azadirachtin, an insecticidal allelochemical from neem on soil microflora, enzyme and respiratory activities. Bioresour. Technol. 2007, 98, 3154–3158. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, P.; Kong, C.H. Urease, invertase, dehydrogenase and polyphenoloxidase activities in paddy soil influenced by allelopathic rice variety. Eur. J. Soil Biol. 2009, 45, 436–441. [Google Scholar] [CrossRef]
- Reigosa, M.S.; Gonzalesy, L.; Souto, X.C.; Pastoriza, J.E. Allelopathy in forest ecosystem. In Allelopathy in Ecological Agriculture and Forestry; Narwal, S.S., Hoagland, R.E., Dilday, R.H., Reigosa, M.J., Eds.; Kluwer Academic Pubishersl: Dordrecht, The Netherlands, 2000; pp. 183–193. [Google Scholar]
- Lebedev, V.G.; Krutovsky, K.V.; Shestibratov, K.A. Fell Upas sits, the hydra-tree of death, or the Phytotoxicity of trees. Molecules 2019, 24, 1636. [Google Scholar] [CrossRef]
- ISAAA. ISAAA’s GM Approval Database. 2022. Available online: http://www.isaaa.org/gmapprovaldatabase/ (accessed on 4 July 2022).
- Kikuchi, A.; Yu, X.; Shimazaki, T.; Kawaoka, A.; Ebinuma, H.; Watanabe, K.N. Allelopathy assessments for the environmental biosafety of the salt-tolerant transgenic Eucalyptus camaldulensis, genotypes codA12-5B, codA12-5C, and codA20C. J. Wood Sci. 2009, 55, 149–153. [Google Scholar] [CrossRef]
- Yu, X.; Kikuchi, A.; Shimazaki, T.; Yamada, A.; Ozeki, Y.; Matsunaga, E.; Ebinuma, H.; Watanabe, K.N. Assessment of the salt tolerance and environmental biosafety of Eucalyptus camaldulensis harboring a mangrin transgene. J. Plant Res. 2013, 126, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.-H.T.; Oguchi, T.; Matsunaga, E.; Kawaoka, A.; Watanabe, K.N.; Kikuchi, A. Evaluation of potential impacts on biodiversity of the salt-tolerant transgenic Eucalyptus camaldulensis harboring an RNA chaperonic RNA-Binding-Protein gene derived from common ice plan. Transgenic Res. 2021, 30, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, T.; Kikuchi, A.; Matsunaga, E.; Nanto, K.; Shimada, T.; Watanabe, K.N. Establishment of a homogenized method for environmental biosafety assessments of transgenic plants. Plant Biotechnol. 2009, 26, 143–148. [Google Scholar] [CrossRef]
- Lebedev, V.; Larionova, A.; Bykhovets, S.; Shanin, V.; Komarov, A.; Shestibratov, K. Model assessment of transgenic trees impact on nitrogen and carbon cycles in forest plantations. In Proceedings of the IUFRO Tree Biotechnology Conference 2015 “Forests: The importance to the planet and society”, Florence, Italy, 8–12 June 2015; pp. 186–188. [Google Scholar] [CrossRef]

| Species | Gene | Growth Conditions | Indicators | Additional Factors | References |
|---|---|---|---|---|---|
| maize | Cry3Bb | field (2 years) | MBC 1 | growth stage | [54] |
| insecticide | |||||
| maize | Cry3Bb | field (3 years) | MBC | growth stage | [17] |
| insecticide | |||||
| maize | Cry1Ab, Cry3Bb1 | field (4 years) | N, P(2), S, DHA 2 | variety | [55] |
| rice | Cry1Ab | field (2 years) | P, DHA | growth stage | [56] |
| insecticide | |||||
| maize | Cry1Ab | field | N, DHA | growth stage | [57] |
| cotton | Cry1Ac | net house | NH4, NO3, N, P 3 | growth stage | [58] |
| DHA | |||||
| cotton | Cry1Ac | net house | organic C | growth stage | [59] |
| N, P(2) | |||||
| MBC, MBN, MBP | |||||
| maize | Cry1Ab | field (7 years) | C, N, texture | [60] | |
| cotton | Cry1Ac | field (3 years) | N(2), P, DHA | [61] | |
| cotton | Cry1Ac | greenhouse (4 years) | C, N(3), P(2), S, DHA, CAT | [9] | |
| Cry1Ac + CpTI | MBC | ||||
| rice | Cry1Ac | open air (pots) | C(2), N(2), P, DHA | growth stage | [62] |
| cotton | Cry1Ac | pots | org. matter, N, P, K | growth stage | [63] |
| C, N(2), P, DHA | |||||
| maize | Cry1Ab | climate chamber | DHA | variety | [64] |
| MBC, MBN | |||||
| maize | Cry1Ab | climate chamber | DHA | soil type | [65] |
| MBC, MBN | |||||
| maize | Cry1Ab | field (5 years) | C | insecticide | [66] |
| MBC | crop rotation | ||||
| cotton | cry1Ac | field | N, DHA | [40] | |
| MBC | |||||
| cotton | CrylAc + CpTI | field (2 years) | N, P, DHA | growth stage | [67] |
| sugarcane | Cry1Ac | field | C, N(2), P | growth stage | [68] |
| cotton | Cry1Ac | field | pH, EC, org. C, NO3, NH4, P, K | fertilization | [32] |
| MBC | |||||
| cotton | Cry1Ac | field (6 years) | C, N(3), P(2), S, DHA | [10] | |
| MBC | |||||
| rice | Cry1Ac | field (8 years) | org. C, N, P, C/N | growth stage | [53] |
| P, N, DHA, CAT | |||||
| MBC, MBN | |||||
| cotton | Cry1Ac | field (2 years) | N, P(2), CAT | growth stage | [12] |
| MBC | salinity | ||||
| maize | Cry1Ab | field | pH, org. C, NO3, NH4, P | irrigation | [69] |
| C, N, P | |||||
| poplar | Cry1Ac, Cry3A | field (5 years) | pH, org. matter, N, P, K | growth stage | [70] |
| cotton | Bt | field | N, P, K | crop rotation | [71] |
| P, DHA | |||||
| MBC | |||||
| poplar | Cry1Ah1 | field (4 years) | pH, N, P | [11] | |
| MBC, MBN, MBP |
| Species | Gene | Growth Conditions | Indicators | Additional Factors | References |
|---|---|---|---|---|---|
| papaya | PRSV CP | field | pH, org. C, N | [79] | |
| papaya | PRSV RP | open air (pots) | pH, EC, org. matter, N, P, K, microel. | [78] | |
| C(3), N(2), P(3), S, DHA, CAT | |||||
| spruce | ech42 | greenh. (5 years) | fungal biomass | [80] | |
| tobacco | McChit1 | chamber house | pH | growth stage | [81] |
| N(2), CAT | |||||
| wheat | WYMV-Nib8 | field (2 years) | C, N, DHA | growth stage | [82] |
| location | |||||
| melon | AFP + CHI | greenhouse | pH, EC, org. matter, P, K | [83] | |
| C(2), N(2), P(2), S, DHA, CAT | |||||
| oilseed rape | NiC | greenhouse | C (2), N, P, S | variety | [84] |
| rice | OsCK1 | field | pH, EC, org. matter, N, P, K, microel. | growth stage | [85] |
| potato | Rpi-vnt1.1 | field (2 years) | pH, org. C, N, C/N | variety | [28] |
| fungicide | |||||
| location |
| Species | Gene | Growth Conditions | Indicators | Additional Factors | References |
|---|---|---|---|---|---|
| alfalfa | MDH | field | pH, P, K, micro | [87] | |
| potato | DREB1A | greenhouse | C, N, P, S | salinity | [88] |
| tobacco | MCM6 | greenhouse | P, DHA | salinity | [89] |
| rice | PDH45 | greenhouse | pH, EC, org. C, N, P, K, microel., texture | salinity | [90] |
| N(2), P, DHA | soil type | ||||
| maize | BADH | greenhouse | pH, EC, org. C, N | growth stage | [91] |
| C, N, CAT | soil type | ||||
| cotton | CBF1 | field (3 years) | pH, EC, org. matter, N, P, K | [92] | |
| maize | BADH | field (3 years) | C, N, DHA | growth stage | [93] |
| Species | Gene | Growth Conditions | Indicators | Additional Factors | References |
|---|---|---|---|---|---|
| poplar | CAD, COMT (AS) 1 | field (4 years) | C, N | location | [98] |
| MBC | |||||
| potato | GBSS (RNAi) | field (3 years) | C(2), PER | growth stage | [100] |
| variety | |||||
| location | |||||
| potato | cphA | field (3 years) | C(4), N(2), P | growth stage | [101] |
| rice | pacrtB + pacrtE + | field | C, N(2), CAT | growth stage | [102] |
| pacrtY + pacrtI | |||||
| soybean | AtD-CGS | field | C, N | [103] | |
| tobacco | CAD, COMT, CCR | greenhouse | MBN | [99] | |
| C(3) | |||||
| potato | GBSS (RNAi) 2 | field | org. matter | growth stage | [104] |
| location | |||||
| rice | AhSTS1 | field | pH, EC, org. matter, N, P, K, microel. | growth stage | [105] |
| switchgrass | COMT (RNAi) | field (5 years) | pH, org. matter, P, K, microel. | [106] | |
| switchgrass | COMT (RNAi) | field (5–6 years) | organic C | [107] | |
| MYB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebedev, V.; Lebedeva, T.; Tikhonova, E.; Shestibratov, K. Assessing Impacts of Transgenic Plants on Soil Using Functional Indicators: Twenty Years of Research and Perspectives. Plants 2022, 11, 2439. https://doi.org/10.3390/plants11182439
Lebedev V, Lebedeva T, Tikhonova E, Shestibratov K. Assessing Impacts of Transgenic Plants on Soil Using Functional Indicators: Twenty Years of Research and Perspectives. Plants. 2022; 11(18):2439. https://doi.org/10.3390/plants11182439
Chicago/Turabian StyleLebedev, Vadim, Tatyana Lebedeva, Elena Tikhonova, and Konstantin Shestibratov. 2022. "Assessing Impacts of Transgenic Plants on Soil Using Functional Indicators: Twenty Years of Research and Perspectives" Plants 11, no. 18: 2439. https://doi.org/10.3390/plants11182439
APA StyleLebedev, V., Lebedeva, T., Tikhonova, E., & Shestibratov, K. (2022). Assessing Impacts of Transgenic Plants on Soil Using Functional Indicators: Twenty Years of Research and Perspectives. Plants, 11(18), 2439. https://doi.org/10.3390/plants11182439