
Effects of Salt Stress on the Morphology, Growth and Physiological Parameters of Juglansmicrocarpa L. Seedlings

Abstract
:1. Introduction
2. Results
2.1. Effects of Salt Stress on External Morphological Characteristics of Seedlings
2.1.1. Symptom Classification of Salt Injury
2.1.2. Change in Morphological Characteristics
2.2. Effects of Salt Stress on Plant Growth
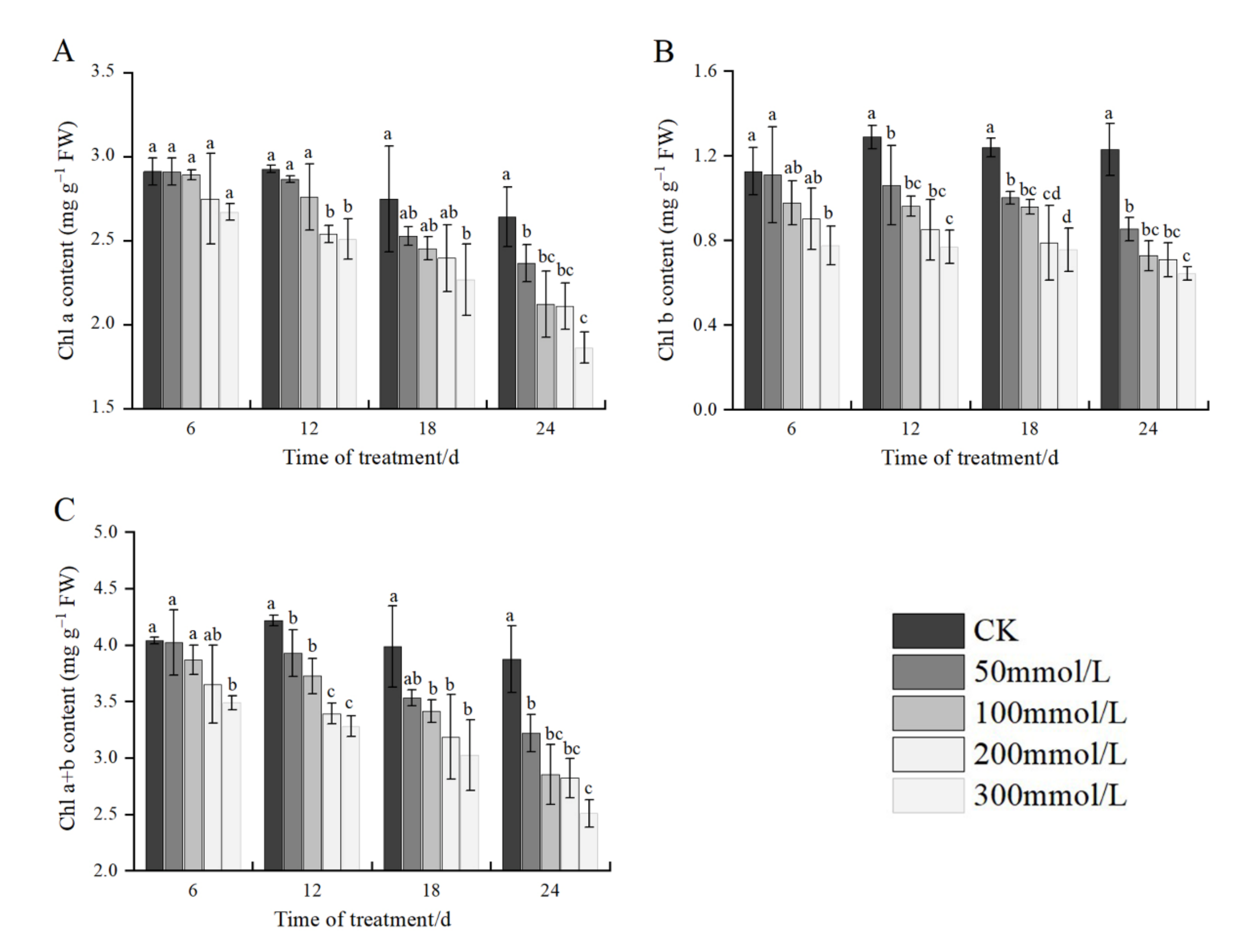
2.3. Effects of Salt Stress on Chlorophyll Content
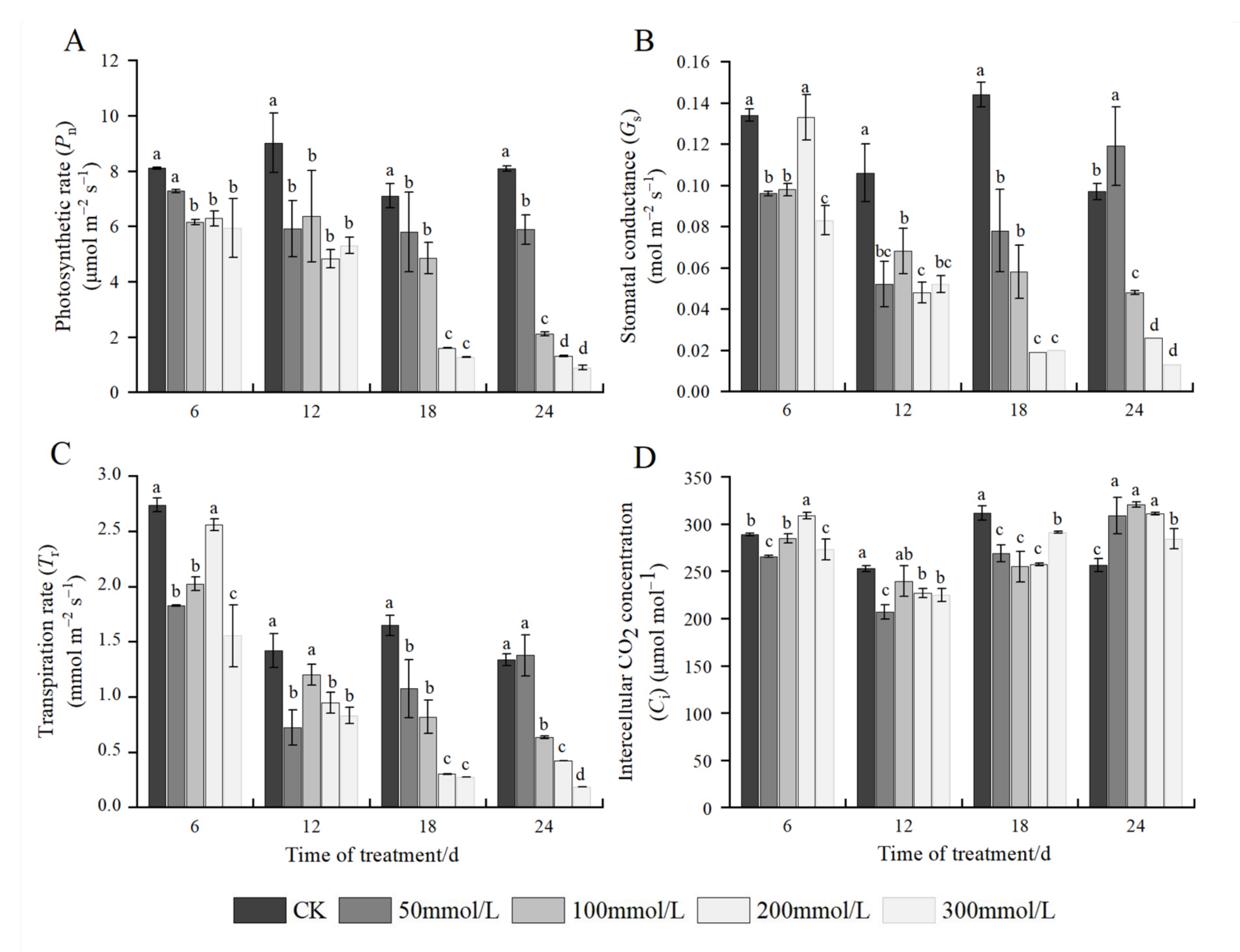
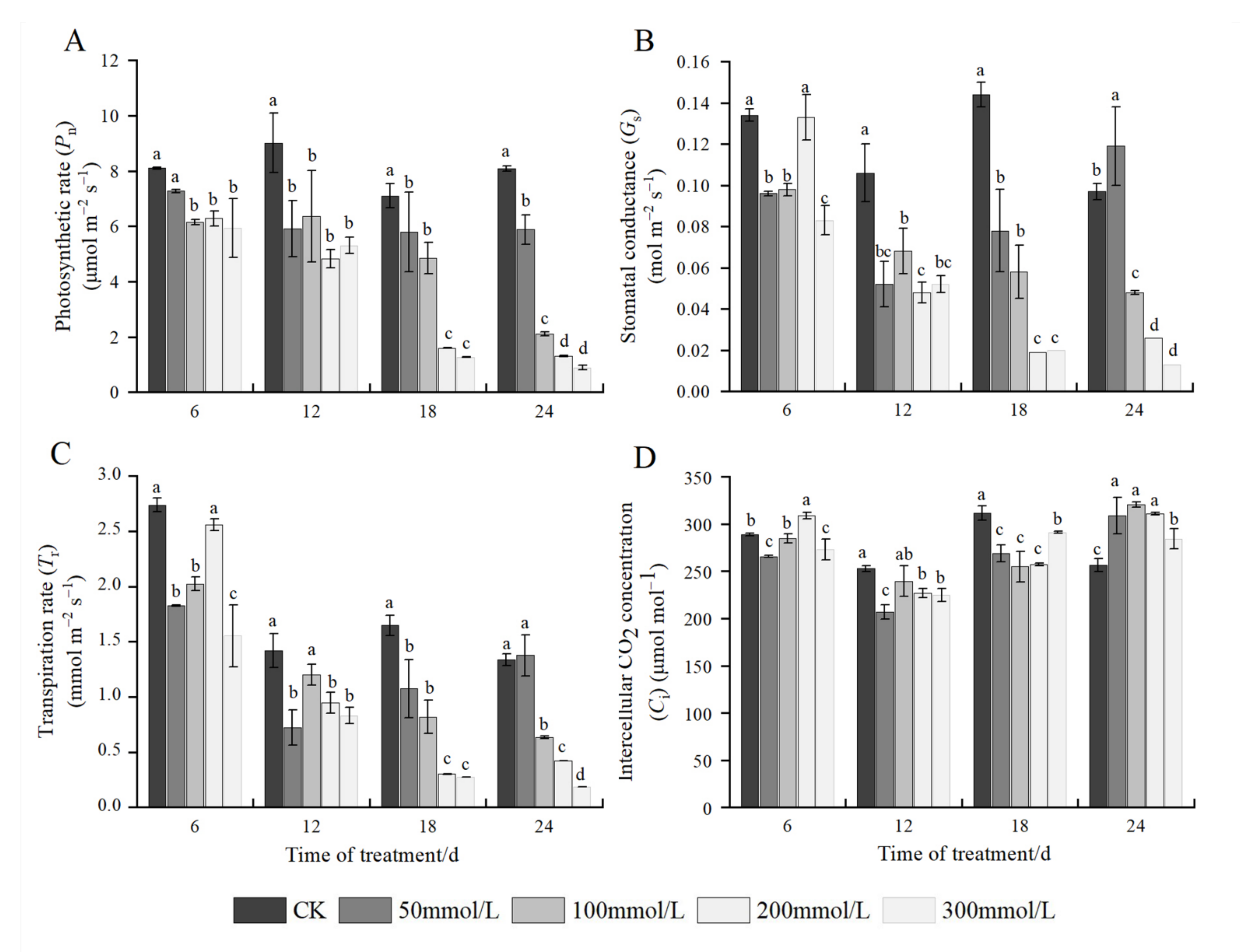
2.4. Effects of Salt Stress on Photosynthetic Parameters
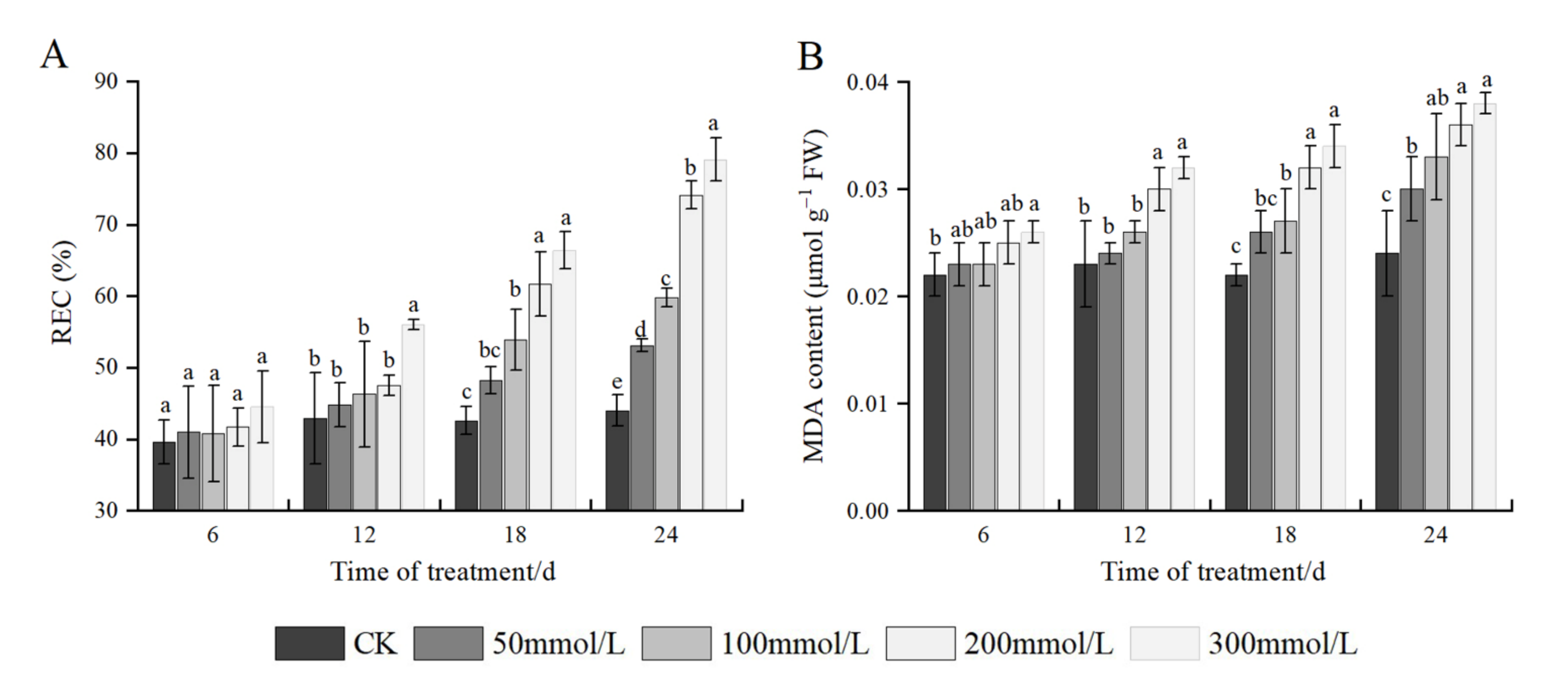
2.5. Effects of Salt Stress on REC and MDA Content
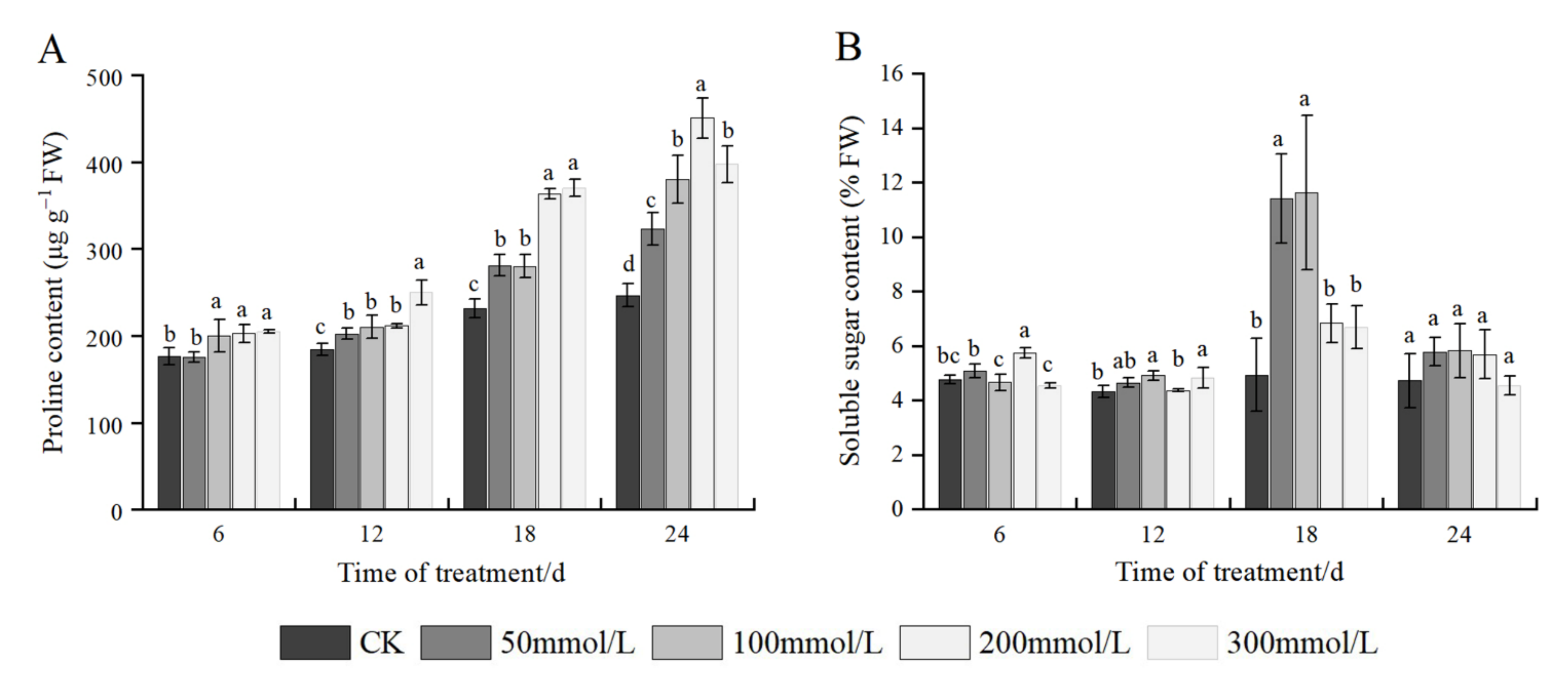
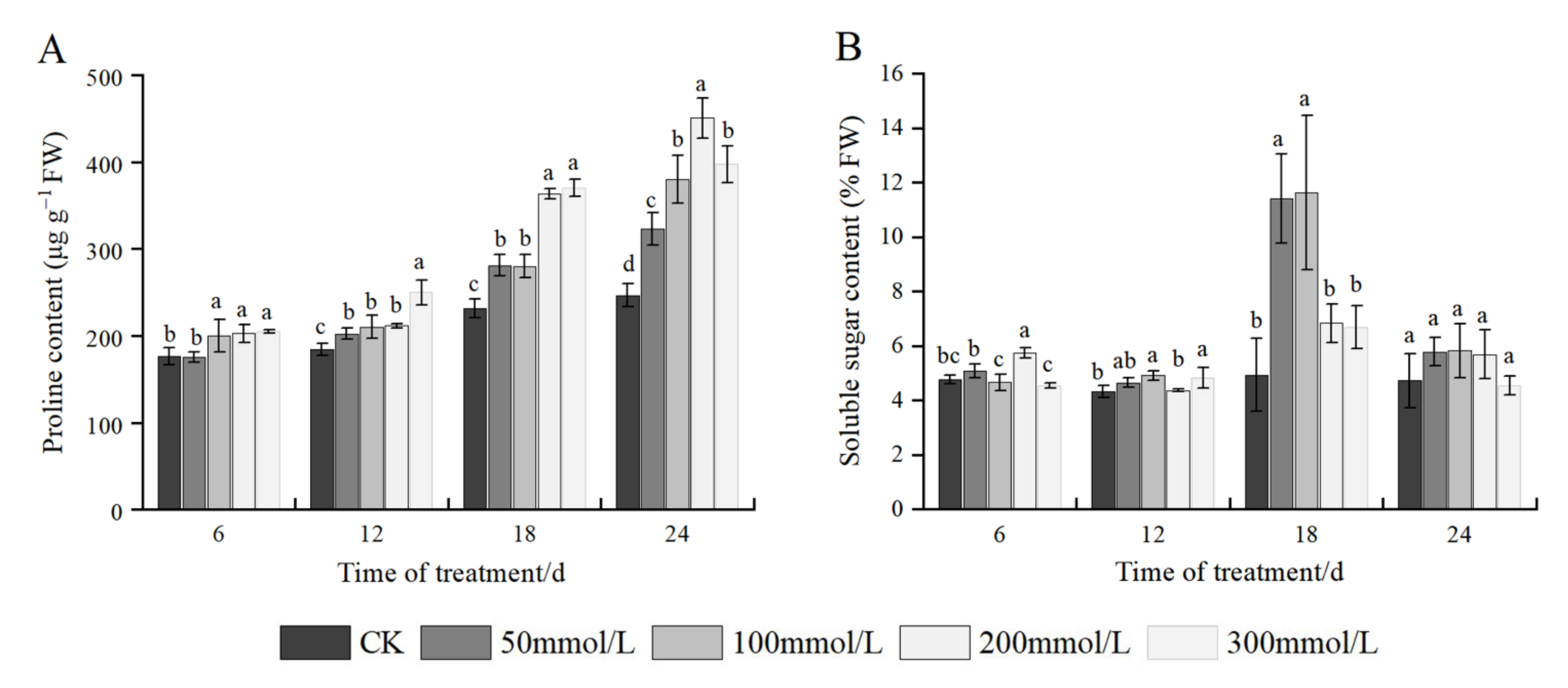
2.6. Effects of Salt Stress on Osmotic Adjustment Substances
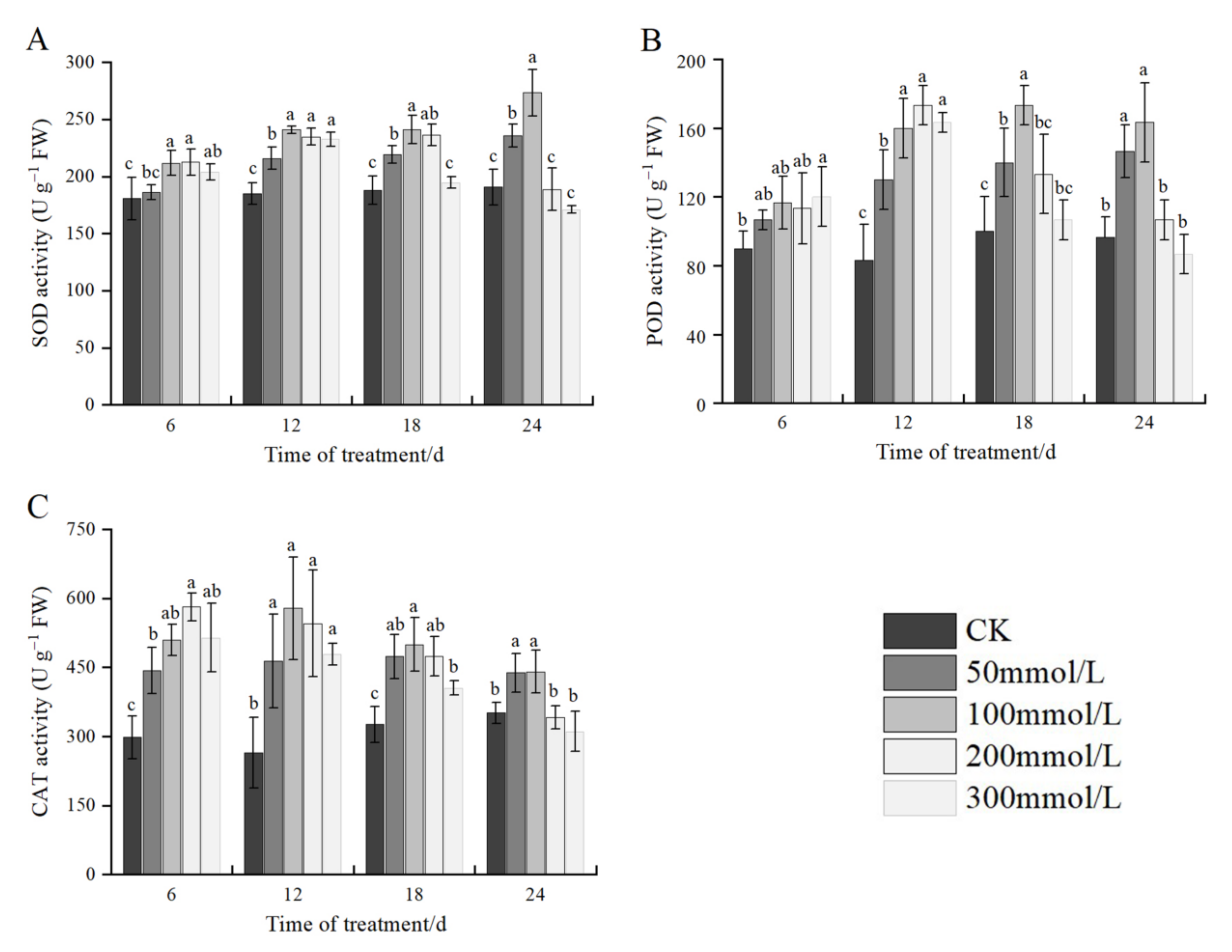
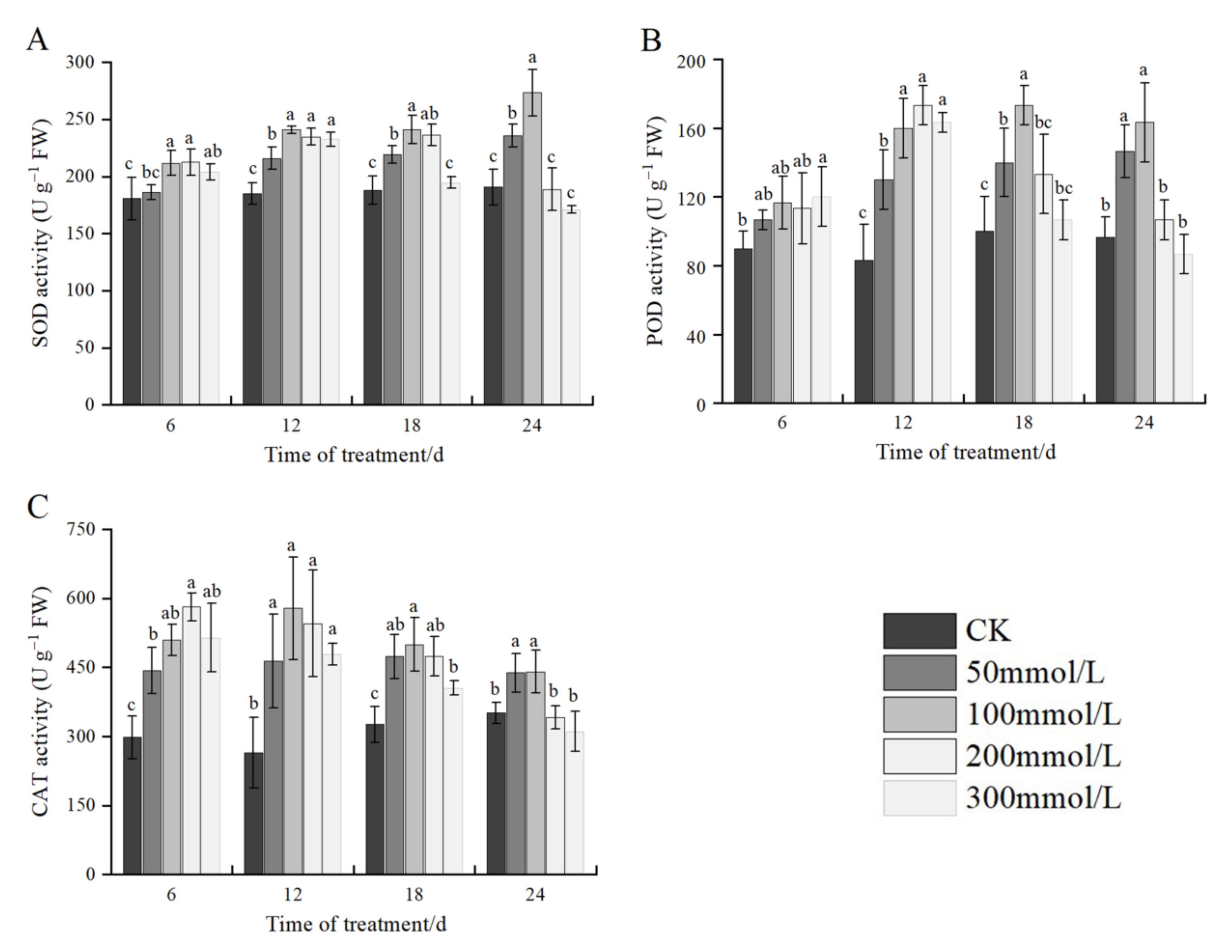
2.7. Effects of Salt Stress on Antioxidant Enzyme Activities
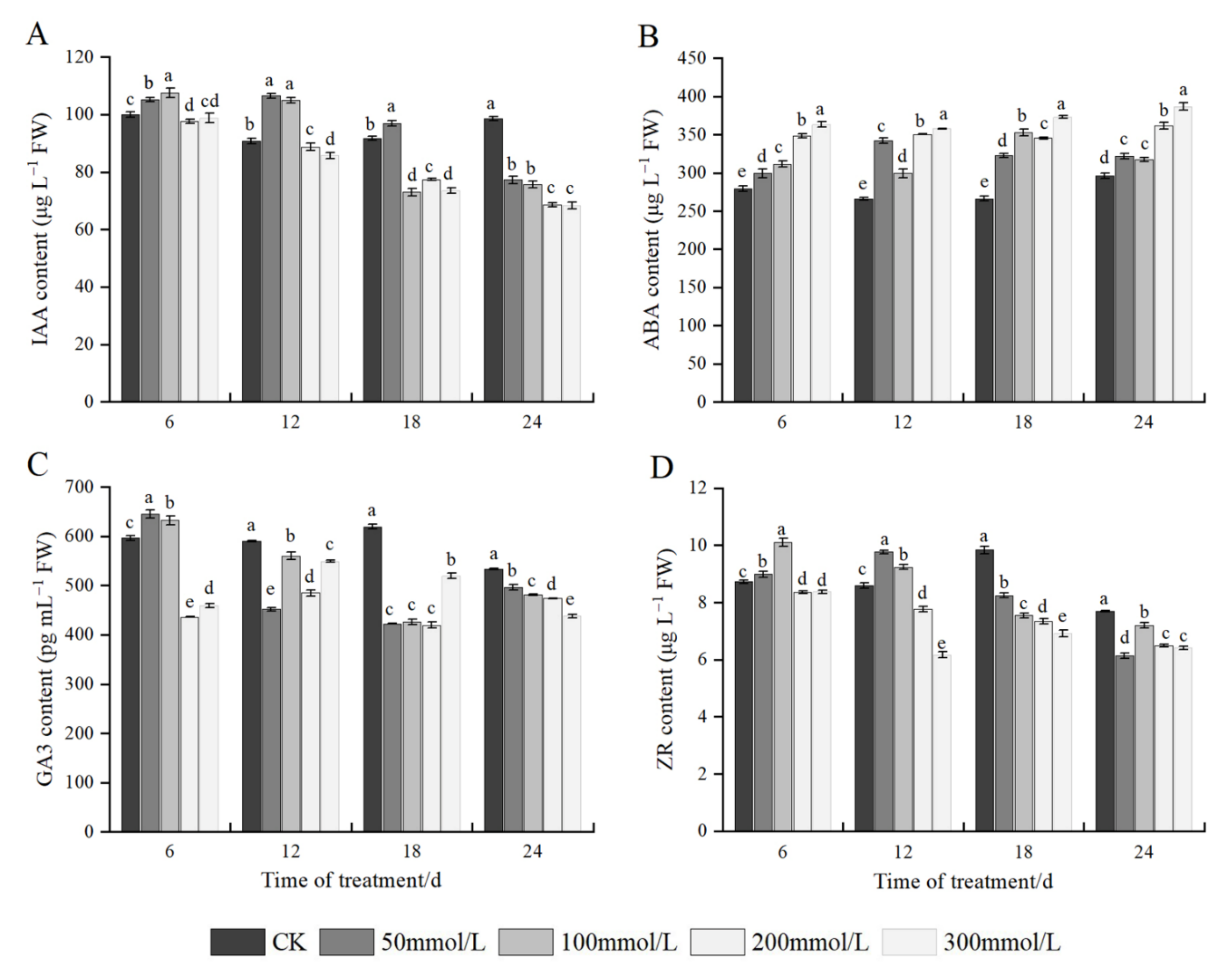
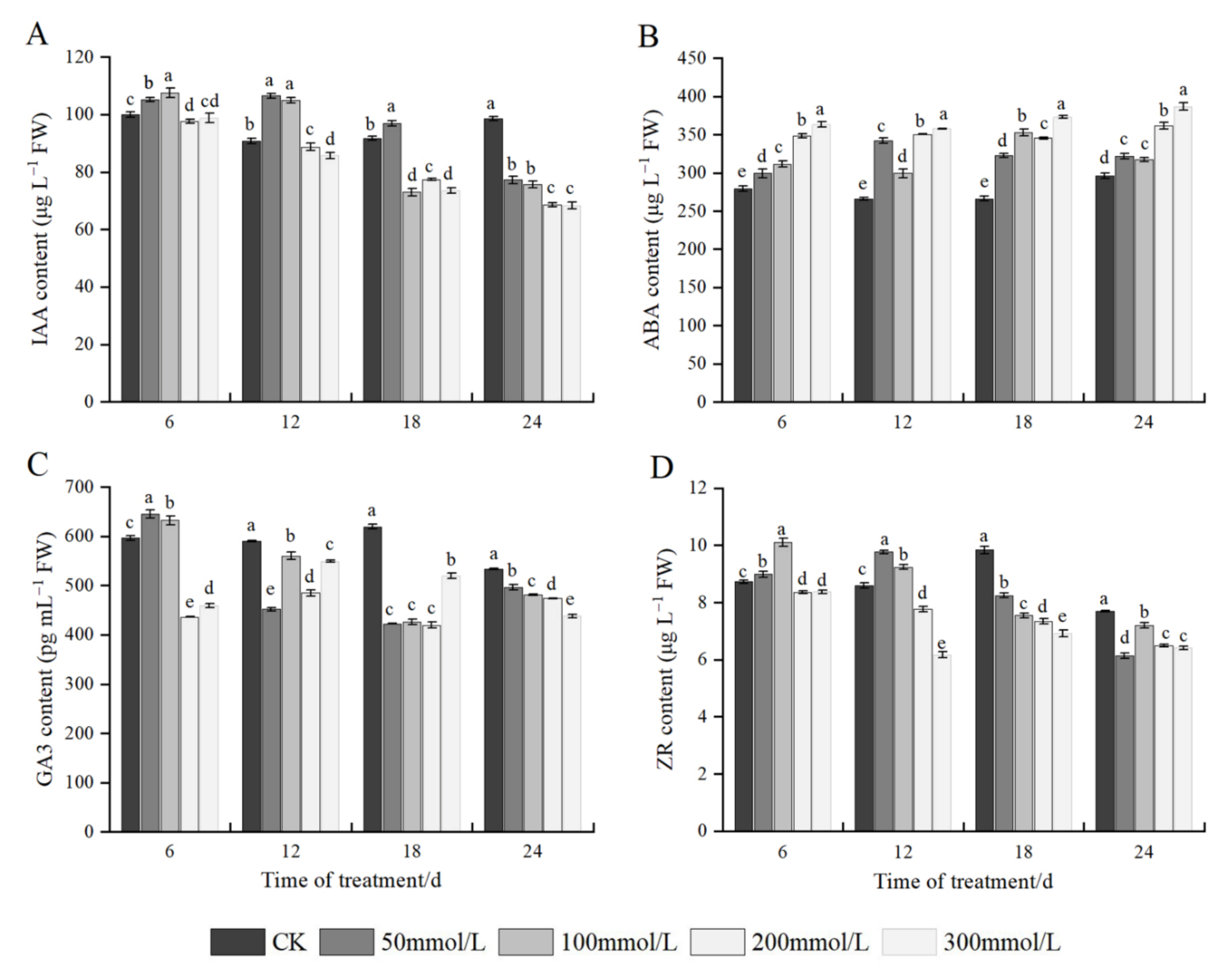
2.8. Effects of Salt Stress on Endogenous Hormones
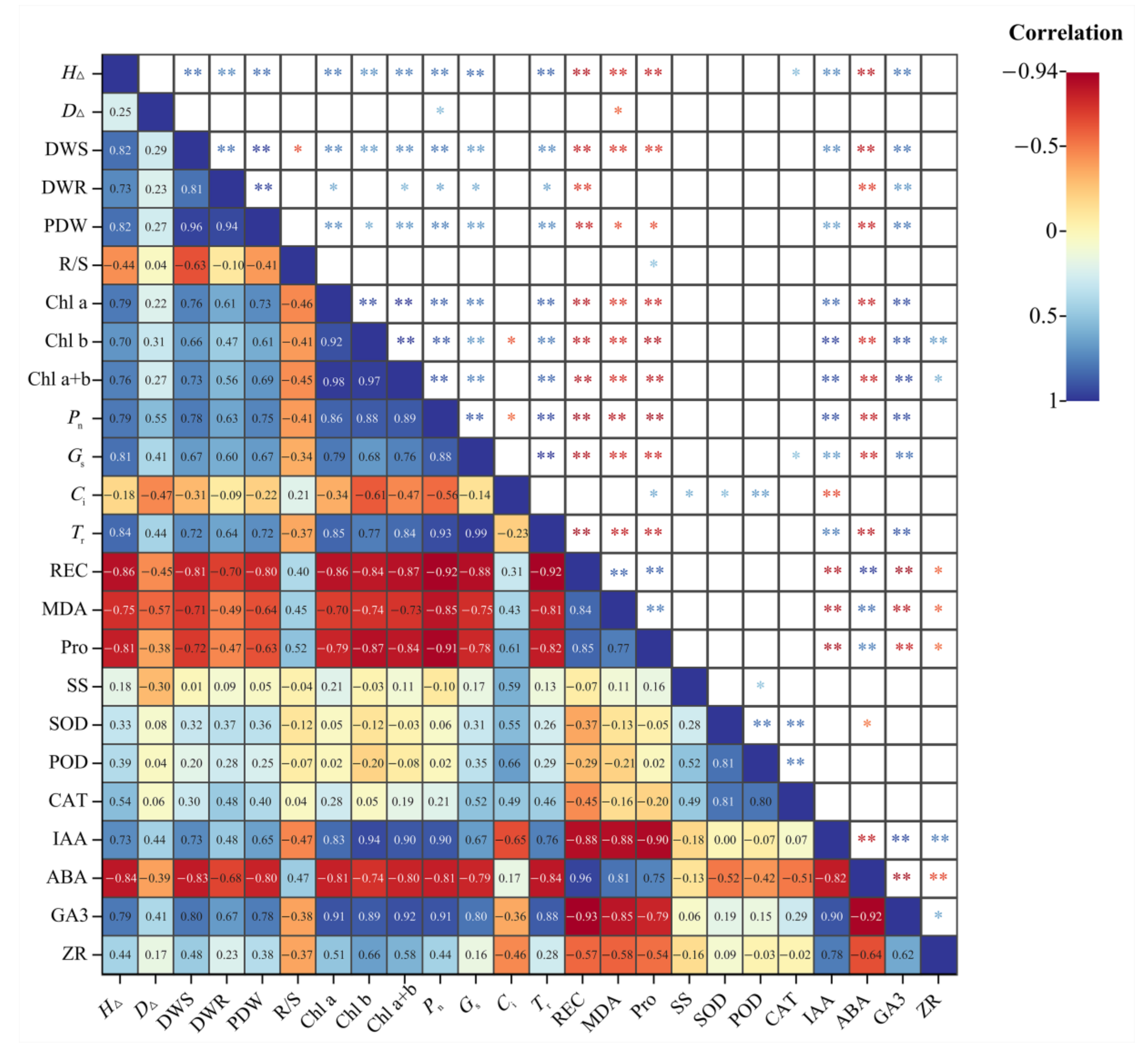
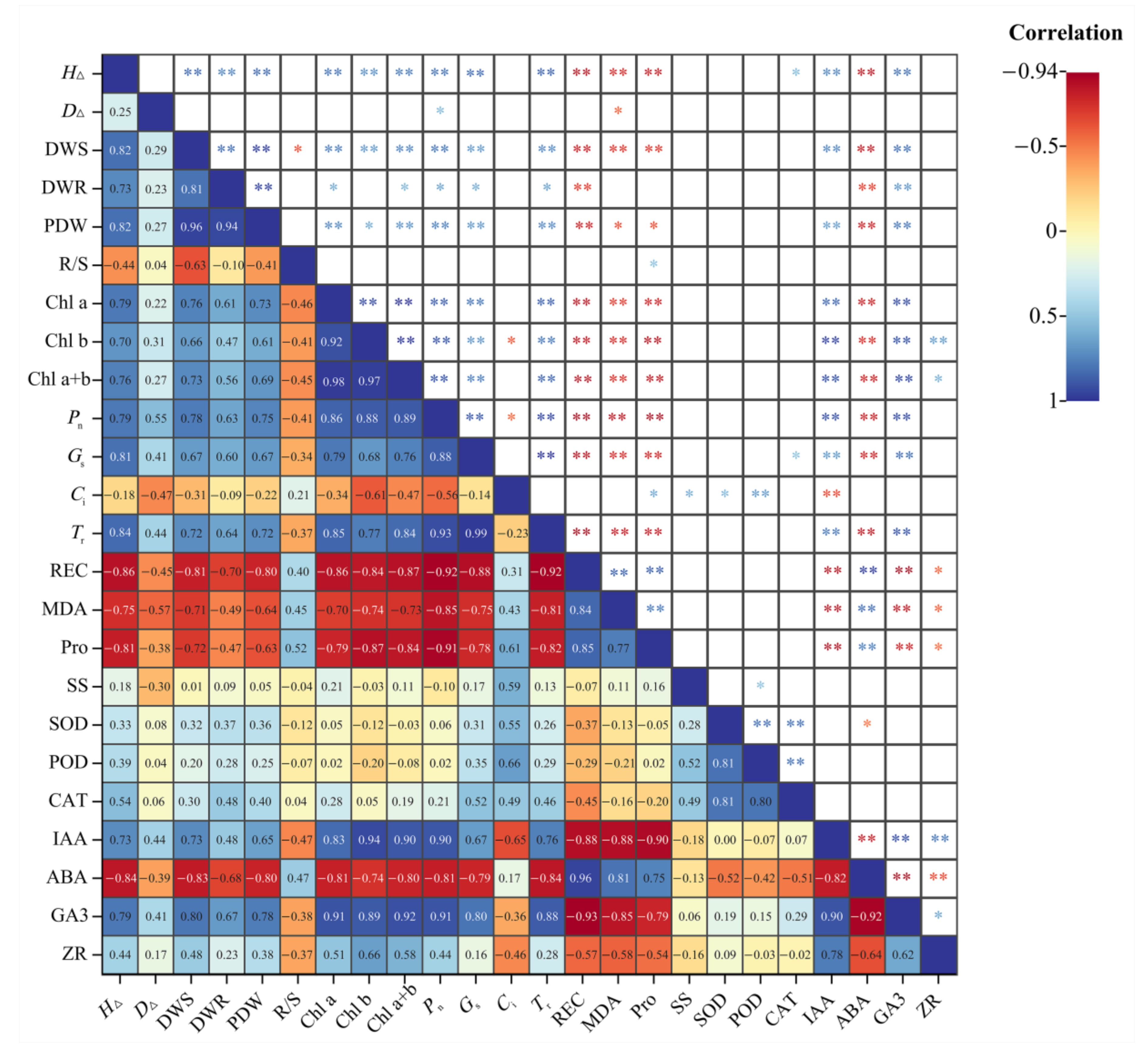
2.9. Correlation Analysis
3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Experimental Design
3.3. Determination of Morphology and Growth Parameters
3.4. Determination of Physiological and Biochemical Indicators
3.4.1. Determination of the Chlorophyll Content
3.4.2. Determination of Photosynthetic Parameters
3.4.3. Determination of Relative Electrical Conductivity
3.4.4. Determination of Malondialdehyde Content
3.4.5. Determination of Proline Content
3.4.6. Determination of Soluble Sugar Content
3.4.7. Determination of Activity of Antioxidant Enzymes
3.4.8. Determination of Endogenous Hormones
3.5. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- He, X.; Yang, Y.; Wei, H.; Yuan, Z. Soil microbiome-mediated salinity tolerance in poplar plantlets is source-dependent. Chemosphere 2021, 272, 129600. [Google Scholar] [CrossRef] [PubMed]
- Dichala, O.; Therios, I.; Papadopoulos, A.; Chatzistathis, T.; Chatzisavvidis, C.; Antonopoulou, C. Effects of varying concentrations of different salts on mineral composition of leaves and roots of three pomegranate (Punica granatum L.) cultivars. Sci. Hortic. 2021, 275, 109718. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Q.; Xu, H.M.; He, J.Q.; Li, A.; Gao, Y. Effects of stress on osmotic regulator and antioxidase activities in Juglans microcarpa seedings. J. Tianjin Agric. Univ. 2021, 28, 1–5. [Google Scholar] [CrossRef]
- Luo, Y.J.; Zhou, P.; Peng, Z.; Zhang, W.X. Effects of salt stress on growth and endogenous hormones of two species of crabapple. Non-Wood For. Res. 2021, 39, 201–210. [Google Scholar]
- Zhong, Y.-P.; Qi, X.-J.; Chen, J.-Y.; Li, Z.; Bai, D.-F.; Wei, C.-G.; Fang, J.-B. Growth and physiological responses of four kiwifruit genotypes to salt stress and resistance evaluation. J. Integr. Agric. 2019, 18, 83–95. [Google Scholar] [CrossRef]
- Pathania, S.; Singh, H. Evaluation and prediction of salinity tolerance behavior of citrus rootstocks. Sci. Hortic. 2021, 289, 110422. [Google Scholar] [CrossRef]
- Ma, Y.; Wei, Z.; Liu, J.; Liu, X.; Liu, F. Growth and physiological responses of cotton plants to salt stress. J. Agron. Crop Sci. 2021, 207, 565–576. [Google Scholar] [CrossRef]
- Abid, M.; Zhang, Y.J.; Li, Z.; Bai, D.F.; Zhong, Y.P.; Fang, J.B. Effect of Salt stress on growth, physiological and biochemical characters of Four kiwifruit genotypes. Sci. Hortic. 2020, 271, 109473. [Google Scholar] [CrossRef]
- Feng, X.; Hussain, T.; Guo, K.; An, P.; Liu, X. Physiological, morphological and anatomical responses of Hibiscus moscheutos to non-uniform salinity stress. Environ. Exp. Bot. 2021, 182, 104301. [Google Scholar] [CrossRef]
- Shahid, M.A.; Balal, R.M.; Khan, N.; Simón-Grao, S.; Alfosea-Simón, M.; Cámara-Zapata, J.M.; Mattson, N.S.; Garcia-Sanchez, F. Rootstocks influence the salt tolerance of Kinnow mandarin trees by altering the antioxidant defense system, osmolyte concentration, and toxic ion accumulation. Sci. Hortic. 2019, 250, 1–11. [Google Scholar] [CrossRef]
- Regni, L.; Del Pino, A.M.; Mousavi, S.; Palmerini, C.A.; Baldoni, L.; Mariotti, R.; Mairech, H.; Gardi, T.; D’Amato, R.; Proietti, P. Behavior of Four Olive Cultivars During Salt Stress. Front. Plant Sci 2019, 10, 867. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.; Karami, H.; Vahdati, K.; Mokhtassi-Bidgoli, A. Antioxidative responses to short-term salinity stress induce drought tolerance in walnut. Sci. Hortic. 2020, 267, 109322. [Google Scholar] [CrossRef]
- Nassar, R.M.A.; Kamel, H.A.; Ghoniem, A.E.; Alarcon, J.J.; Sekara, A.; Ulrichs, C.; Abdelhamid, M.T. Physiological and Anatomical Mechanisms in Wheat to Cope with Salt Stress Induced by Seawater. Plants 2020, 9, 237. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.R.S.; Batista, B.L.; Lobato, A.K.S. Anatomical changes in stem and root of soybean plants submitted to salt stress. Plant Biol. 2021, 23, 57–65. [Google Scholar] [CrossRef]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How Plant Hormones Mediate Salt Stress Responses. Trends Plant Sci 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd Allah, E.F. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef]
- He, L.; Xiaoxue, D.; Jialin, Z.; Jinna, L.; Sixue, C.; Huizi, D.; Haiying, L. Quantitative redox proteomics revealed molecular mechanisms of salt tolerance in the roots of sugar beet monomeric addition line M14. Bot. Stud. 2022, 63, 5. [Google Scholar]
- Da-Ru, W.; Kuo, Y.; Xun, W.; Chun-Xiang, Y. A C2H2-type zinc finger transcription factor, MdZAT17, acts as a positive regulator in response to salt stress. J. Plant Physiol. 2022, 275, 153737. [Google Scholar]
- Zhang, J.X.; Wei, X.P.; Mao, Z.Q.; Ji, Q.L. The germplasm of american black walnut and their exploitation and utilization. J. Shangdon Agric. Univ. 2001, 32, 555–560. [Google Scholar]
- Wang, X.L. Physiological and Biochemical responses of the new cultivar of Zelkova schneideriana ‘Hentiangao’ under Saline-alkalinity Stress; Central South University of Forestry and Technology: Changsha, China, 2016. [Google Scholar]
- Zhu, S.; Nong, J.; Luo, G.; Li, Q.; Wang, F.; Jiang, D.; Zhao, X. Varied tolerance and different responses of five citrus rootstocks to acid stress by principle component analysis and orthogonal analysis. Sci. Hortic. 2021, 278, 109853. [Google Scholar] [CrossRef]
- Esfandiari Ghalati, R.; Shamili, M.; Homaei, A. Effect of putrescine on biochemical and physiological characteristics of guava (Psidium guajava L.) seedlings under salt stress. Sci. Hortic. 2020, 261, 108961. [Google Scholar] [CrossRef]
- Wang, J.; Qiao, Q.; Tao, J. The Physiological Response of Three Narcissus pseudonarcissus under NaCl Stress. Am. J. Plant Sci. 2019, 10, 447–461. [Google Scholar] [CrossRef]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef]
- Kong, X.; Pan, J.; Zhang, M.; Xing, X.; Zhou, Y.; Liu, Y.; Li, D.; Li, D. ZmMKK4, a novel group C mitogen-activated protein kinase kinase in maize (Zea mays), confers salt and cold tolerance in transgenic Arabidopsis. Plant Cell Environ. 2011, 34, 1291–1303. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yan, M.; Huang, X.; Yuan, Z.J.P.J.o.B. Effcts of salt stress on growth and physiological characteristics of pomegranate (Punica granatum L.) cuttings. Pak. J. Bot. 2018, 50, 457–464. [Google Scholar]
- Khalid, M.F.; Hussain, S.; Anjum, M.A.; Ahmad, S.; Ali, M.A.; Ejaz, S.; Morillon, R. Better salinity tolerance in tetraploid vs diploid volkamer lemon seedlings is associated with robust antioxidant and osmotic adjustment mechanisms. J. Plant Physiol. 2020, 244, 153071. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Muñoz, J.L.; García-Molina, F.; García-Ruiz, P.A.; Arribas, E.; Tudela, J.; García-Cánovas, F.; Rodríguez-López, J.N. Enzymatic and chemical oxidation of trihydroxylated phenols. Food Chem. 2009, 113, 435–444. [Google Scholar] [CrossRef]
- Urbinati, G.; Nota, P.; Frattarelli, A.; Lucioli, S.; Forni, C.; Caboni, E. Morpho-physiological responses of sea buckthorn (Hippophae rhamnoides) to NaCl stress. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2019, 154, 827–834. [Google Scholar] [CrossRef]
- Mao, P.; Zhang, Y.; Cao, B.; Guo, L.; Shao, H.; Cao, Z.; Jiang, Q.; Wang, X. Effects of salt stress on eco-physiological characteristics in Robinia pseudoacacia based on salt-soil rhizosphere. Sci. Total Environ. 2016, 568, 118–123. [Google Scholar] [CrossRef]
- Lidia, A.; Aurea, H.; Rocío, C.; Arjona-López, J.M.; Arenas-Arenas, F.J. Salinity Effect on Plant Physiological and Nutritional Parameters of New Huanglongbing Disease-Tolerant Citrus Rootstocks. Agronomy 2021, 11, 653. [Google Scholar]
- Banakar, M.H.; Amiri, H.; Sarafraz Ardakani, M.R.; Ranjbar, G.H. Susceptibility and tolerance of fenugreek (Trigonella foenum-graceum L.) to salt stress: Physiological and biochemical inspections. Environ. Exp. Bot. 2022, 194, 104748. [Google Scholar] [CrossRef]
- Karimi, R.; Ebrahimi, M.; Amerian, M. Abscisic acid mitigates NaCl toxicity in grapevine by influencing phytochemical compounds and mineral nutrients in leaves. Sci. Hortic. 2021, 288, 110336. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How Does Proline Treatment Promote Salt Stress Tolerance During Crop Plant Development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Crizel, R.L.; Perin, E.C.; Siebeneichler, T.J.; Borowski, J.M.; Messias, R.S.; Rombaldi, C.V.; Galli, V. Abscisic acid and stress induced by salt: Effect on the phenylpropanoid, L-ascorbic acid and abscisic acid metabolism of strawberry fruits. Plant Physiol. Biochem. 2020, 152, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Albacete, A.; Ghanem, M.E.; Martinez-Andujar, C.; Acosta, M.; Sanchez-Bravo, J.; Martinez, V.; Lutts, S.; Dodd, I.C.; Perez-Alfocea, F. Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2008, 59, 4119–4131. [Google Scholar] [CrossRef] [PubMed]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. Evol. Syst. 2004, 6, 207–215. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Diaz-Vivancos, P.; Alvarez, S.; Fernandez-Garcia, N.; Sanchez-Blanco, M.J.; Hernandez, J.A. NaCl-induced physiological and biochemical adaptative mechanisms in the ornamental Myrtus communis L. plants. J. Plant Physiol. 2015, 183, 41–51. [Google Scholar] [CrossRef]
- Sarabi, B.; Fresneau, C.; Ghaderi, N.; Bolandnazar, S.; Streb, P.; Badeck, F.W.; Citerne, S.; Tangama, M.; David, A.; Ghashghaie, J. Stomatal and non-stomatal limitations are responsible in down-regulation of photosynthesis in melon plants grown under the saline condition: Application of carbon isotope discrimination as a reliable proxy. Plant Physiol. Biochem. 2019, 141, 1–19. [Google Scholar] [CrossRef]
- Yuan, J.Q.; Sun, D.W.; Lu, Q.; Yang, L.; Wang, H.W.; Fu, X.X. Responses of Physiology, Photosynthesis, and Related Genes to Saline Stress in Cornus hongkongensis subsp. tonkinensis (W. P. Fang) Q. Y. Xiang. Plants 2022, 11, 940. [Google Scholar] [CrossRef]
- Lu, Q.; Yang, L.; Wang, H.; Yuan, J.; Fu, X.; Fang, Y. Responses of photosynthetic characteristics and chloroplast ultrastructure to salt stress in seedlings of Cornus hongkongensis subsp. Elegans. J. Nanjing For. Univ. 2020, 44, 29–36. [Google Scholar]
- Khoshbakht, D.; Ramin, A.A.; Baninasab, B. Effects of sodium chloride stress on gas exchange, chlorophyll content and nutrient concentrations of nine citrus rootstocks. Photosynthetica 2015, 53, 241–249. [Google Scholar] [CrossRef]
- Upham, B.L.; Jahnke, L.S.J.P.r. Photooxidative reactions in chloroplast thylakoids. Evidence for a Fenton-type reaction promoted by superoxide or ascorbate. Photosynth. Res. 1986, 8, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xu, Z.; Peng, S.; Sun, Y.; Jia, C.; Zhai, M.J.P.c.r. In planta characterization of a tau class glutathione S-transferase gene from Juglans regia (JrGSTTau1) involved in chilling tolerance. Plant Cell Rep. 2016, 35, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, W.; Liu, Z.; Yi-Maer, A.Y.; Zhai, M.; Xu, Z. Both JrWRKY2 and JrWRKY7 of Juglans regia mediate responses to abiotic stresses and abscisic acid through formation of homodimers and interaction. Plant Biol. 2017, 19, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Env. 2014, 37, 300–311. [Google Scholar] [CrossRef]
- Alam, A.; Ullah, H.; Attia, A.; Datta, A. Effects of Salinity Stress on Growth, Mineral Nutrient Accumulation and Biochemical Parameters of Seedlings of Three Citrus Rootstocks. Int. J. Fruit Sci. 2019, 20, 786–804. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, J.; Pei, D.; Yu, L. Combined effects of water stress and salinity on growth, physiological, and biochemical traits in two walnut genotypes. Physiol. Plant 2021, 172, 176–187. [Google Scholar] [CrossRef]
- De Baerdemaeker, N.J.F.; Salomon, R.L.; De Roo, L.; Steppe, K. Sugars from woody tissue photosynthesis reduce xylem vulnerability to cavitation. New Phytol. 2017, 216, 720–727. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, J.; Rui, M.; Yang, L.; Shen, J.; Chu, H.; Song, S.; Chen, Y. OsFTIP7 determines metallic oxide nanoparticles response and tolerance by regulating auxin biosynthesis in rice. J. Hazard Mater. 2021, 403, 123946. [Google Scholar] [CrossRef]
- Sachs, T. Auxin’s role as an example of the mechanisms of shoot/root relations. Plant Soil. 2005, 268, 13–19. [Google Scholar] [CrossRef]
- Lovelli, S.; Scopa, A.; Perniola, M.; Di Tommaso, T.; Sofo, A. Abscisic acid root and leaf concentration in relation to biomass partitioning in salinized tomato plants. J. Plant Physiol. 2012, 169, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhang, Q.; Ervin, E.H.; Yang, Z.; Zhang, X. Physiological Mechanism of Enhancing Salt Stress Tolerance of Perennial Ryegrass by 24-Epibrassinolide. Front. Plant Sci. 2017, 8, 1017. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F. The role of salicylic acid and gibberellin signaling in plant responses to abiotic stress with an emphasis on heavy metals. Plant Signal. Behav. 2020, 15, 1777372. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Wang, X.; Dong, C.; Zhang, Z.; Shang, Q.J.R.J.o.P.P. Salicylic acid induces stomatal closure by modulating endogenous hormone levels in cucumber cotyledons. J. Plant Physiol. 2011, 58, 906–913. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Salt Stress Symptoms |
|---|---|
| 0 | No symptoms of salt stress damage. |
| 1 | A few tips and margins of the lower old leaves turn yellow and wither. |
| 2 | Some tips and margins of the lower old leaves turn yellow and wither. |
| 3 | Some tips and margins of the lower old leaves were withered, and the withered and curled number of the middle and upper leaves increased. |
| 4 | Most leaves are withered and curled, and some compound leaves have withered petioles. |
| 5 | Most leaves are withered and curled, accompanied by falling; the lower compound leaves and total petioles are withered, accompanied by a few fallen off. |
| 6 | Most leaves are withered, curled, and fallen off, and most compound leaves and total petioles are withered and fallen off. |
| NaCl /(mmol·L−1) | Stress Symptom Grade | |||
|---|---|---|---|---|
| 6 d | 12 d | 18 d | 24 d | |
| 50 | 0 | 0 | 1 | 3 |
| 100 | 0 | 0 | 2 | 3 |
| 200 | 0 | 2 | 3 | 5 |
| 300 | 0 | 2 | 5 | 6 |
| NaCl /(mmol·L−1) | HΔ/cm | DΔ/mm | DWS/g | DWR/g | PDW/g | R/S |
|---|---|---|---|---|---|---|
| 0 | 1.133 ± 0.058 a | 0.907 ± 0.291 a | 8.187 ± 0.973 a | 7.890 ± 1.068 a | 16.077 ± 1.842 a | 0.966 ± 0.102 a |
| 50 | 1.067 ± 0.153 a | 0.808 ± 0.384 a | 7.083 ± 1.320 ab | 7.950 ± 1.541 a | 15.033 ± 2.838 ab | 1.122 ± 0.049 a |
| 100 | 0.933 ± 0.058 a | 0.598 ± 0.140 a | 6.437 ± 0.883 ab | 7.010 ± 0.879 ab | 13.477 ± 1.746 ab | 1.091 ± 0.044 a |
| 200 | 0.633 ± 0.208 b | 0.588 ± 0.122 a | 4.970 ± 1.537 bc | 6.510 ± 0.518 ab | 11.480 ± 1.686 bc | 1.410 ± 0.487 a |
| 300 | 0.600 ± 0.173 b | 0.582 ± 0.304 a | 4.053 ± 1.220 c | 4.883 ± 1.727 b | 8.937 ± 2.600 c | 1.247 ± 0.449 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, X.; Tang, J.; Zhang, J. Effects of Salt Stress on the Morphology, Growth and Physiological Parameters of Juglansmicrocarpa L. Seedlings. Plants 2022, 11, 2381. https://doi.org/10.3390/plants11182381
Ji X, Tang J, Zhang J. Effects of Salt Stress on the Morphology, Growth and Physiological Parameters of Juglansmicrocarpa L. Seedlings. Plants. 2022; 11(18):2381. https://doi.org/10.3390/plants11182381
Chicago/Turabian StyleJi, Xinying, Jiali Tang, and Junpei Zhang. 2022. "Effects of Salt Stress on the Morphology, Growth and Physiological Parameters of Juglansmicrocarpa L. Seedlings" Plants 11, no. 18: 2381. https://doi.org/10.3390/plants11182381
APA StyleJi, X., Tang, J., & Zhang, J. (2022). Effects of Salt Stress on the Morphology, Growth and Physiological Parameters of Juglansmicrocarpa L. Seedlings. Plants, 11(18), 2381. https://doi.org/10.3390/plants11182381