
Diversity of Molluscan Assemblage in Relation to Biotic and Abiotic Variables in Brown Algal Forests




Abstract
:1. Introduction
2. Materials and Methods
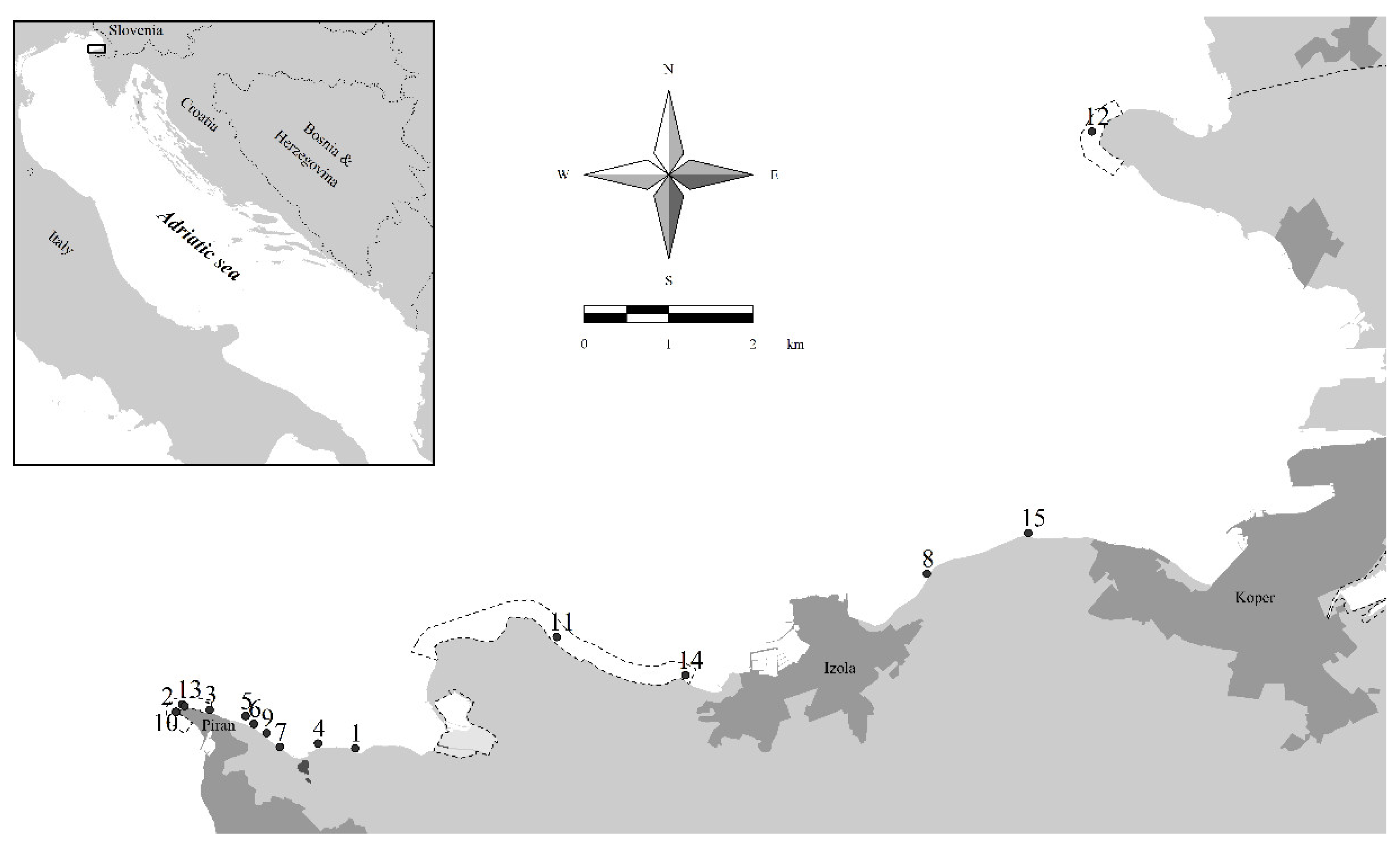
2.1. Study Area and Sampling Sites
2.2. Fieldwork and Laboratory Work
2.3. Physicochemical Parameters
2.4. Data Analysis
3. Results
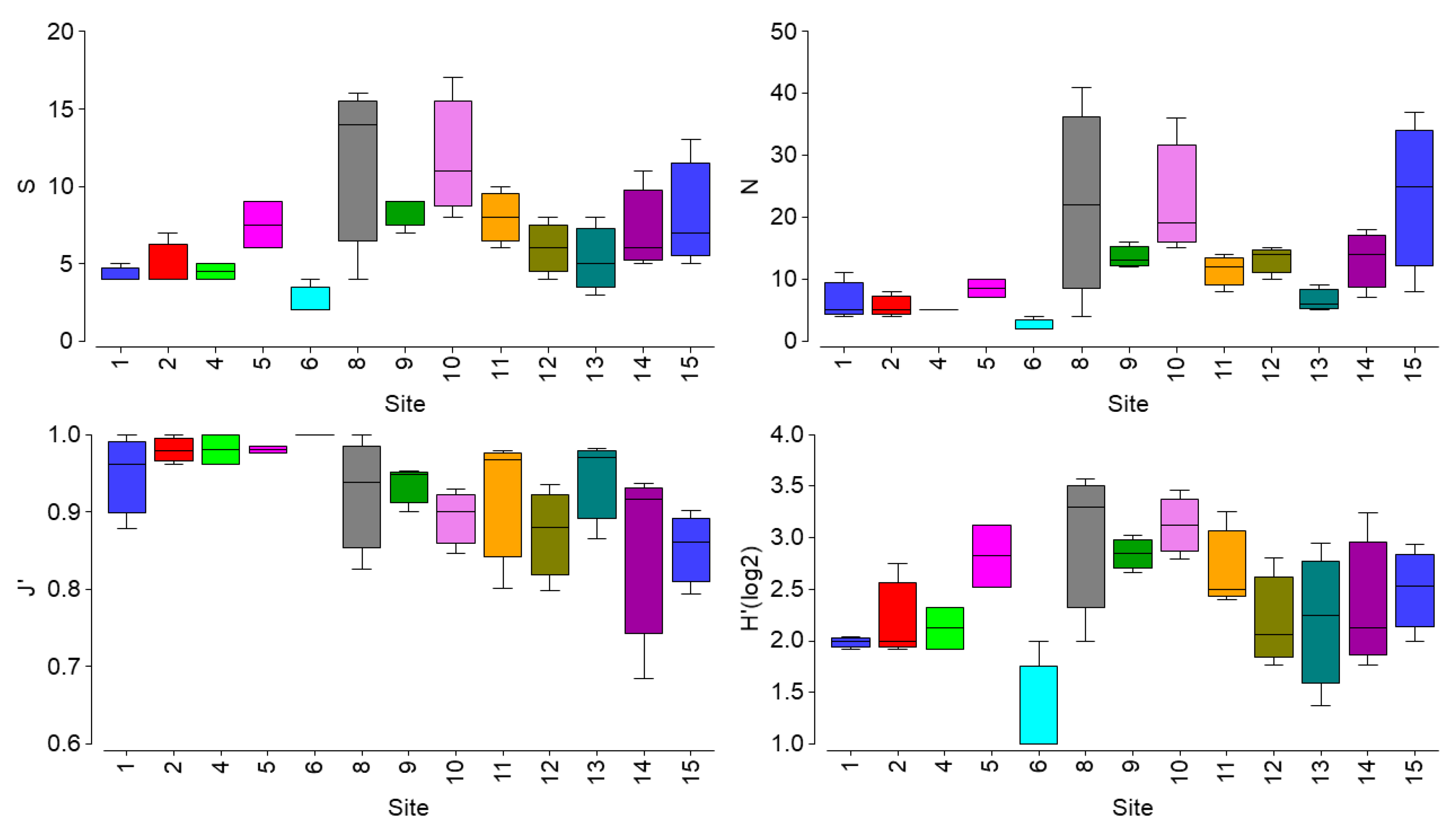
3.1. Ecological Status Evaluation of Macroalgal Communities
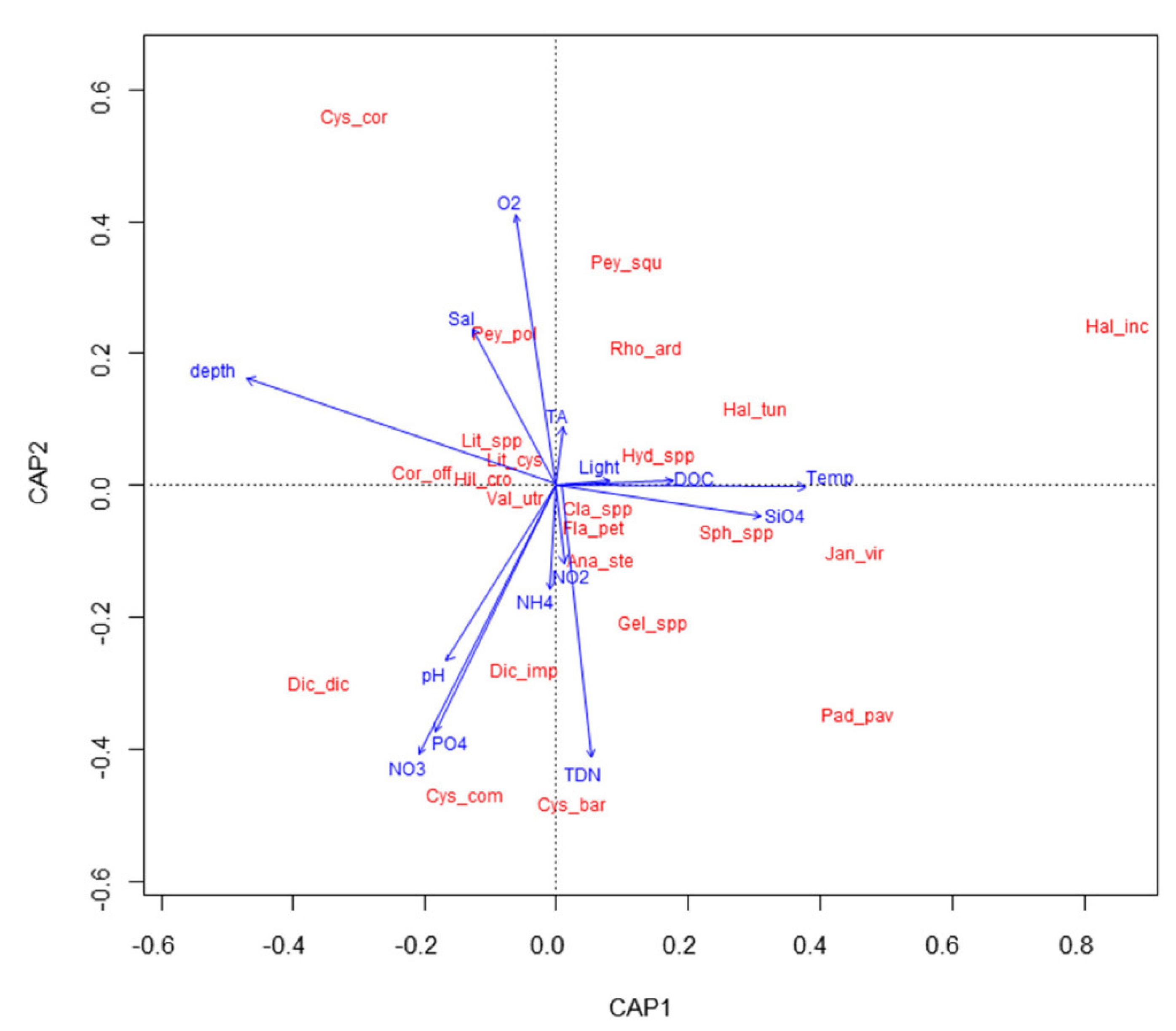
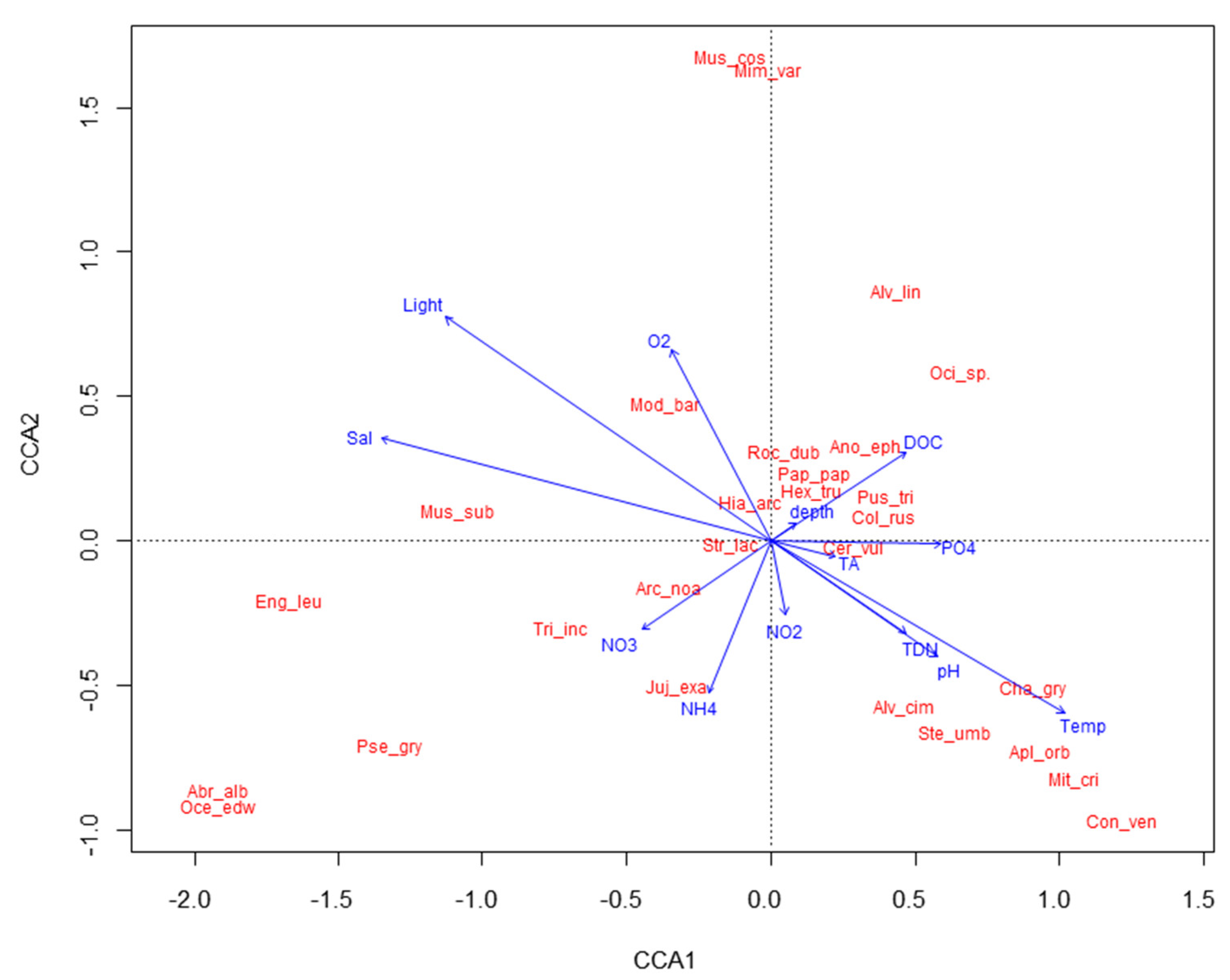
3.2. Molluscan and Algal Assemblages in Relation to Abiotic Factors
4. Discussion
4.1. Characterization of Algal and Molluscan Assemblages
4.2. Biotic and Abiotic Factors Affecting Molluscan Assemblages
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schiel, D.R.; Foster, M.S. The population biology of large brown seaweeds: Ecological consequences of multiphase life histories in dynamic coastal environments. Annu. Rev. Ecol. Syst. 2006, 37, 343–372. [Google Scholar] [CrossRef]
- Bulleri, F.; Benedetti-Cecchi, L.; Cusson, M.; Maggi, E.; Arenas, F.; Aspden, R.; Bertocci, I.; Crowe, T.P.; Davoult, D.; Eriksson, B.K.; et al. Temporal stability of European rocky shore assemblages: Variation across a latitudinal gradient and the role of habitat-formers. Oikos 2012, 121, 1801–1809. [Google Scholar] [CrossRef]
- Derrien-Courtel, S.; Le Gal, A.; Grall, J. Regional-scale analysis of subtidal rocky shore community. Helgol. Mar. Res. 2013, 67, 697–712. [Google Scholar] [CrossRef]
- Bringloe, T.T.; Starko, S.; Wade, R.M.; Vieira, C.; Kawai, H.; De Clerck, O.; Cock, J.M.; Coelho, S.M.; Destombe, C.; Valero, M.; et al. Phylogeny and Evolution of the Brown Algae. Crit. Rev. Plant Sci. 2020, 39, 281–321. [Google Scholar] [CrossRef]
- Orfanidis, S.; Rindi, F.; Cebrian, E.; Fraschetti, S.; Nasto, I.; Taskin, E.; Bianchelli, S.; Papathanasiou, V.; Kosmidou, M.; Caragnano, A.; et al. Effects of Natural and Anthropogenic Stressors on Fucalean Brown Seaweeds Across Different Spatial Scales in the Mediterranean Sea. Front. Mar. Sci. 2021, 8, 658417. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Bermejo, R.; Chefaoui, R.M.; Engelen, A.H.; Buonomo, R.; Neiva, J.; Ferreira-Costa, J.; Pearson, G.A.; Marbà, N.; Duarte, C.M.; Airoldi, L.; et al. Marine forests of the Mediterranean-Atlantic Cystoseira tamariscifolia complex show a southern Iberian genetic hotspot and no reproductive isolation in parapatry. Sci. Rep. 2018, 8, 10427. [Google Scholar] [CrossRef]
- Assis, J.; Fragkopoulou, E.; Frade, D.; Neiva, J.; Oliveira, A.; Abecasis, D.; Faugeron, S.; Serrão, E.A. A fine-tuned global distribution dataset of marine forests. Sci. Data 2020, 7, 119. [Google Scholar] [CrossRef]
- Airoldi, L.; Balata, D.; Beck, M.W. The Gray zone: Relationships between habitat loss and marine diversity and their applications in conservation. J. Exp. Mar. Biol. Ecol. 2008, 366, 8–15. [Google Scholar] [CrossRef]
- Airoldi, L.; Beck, M.W.; Firth, L.B.; Bugnot, A.B.; Steinberg, P.D.; Dafforn, K.A. Emerging Solutions to Return Nature to the Urban Ocean. Ann. Rev. Mar. Sci. 2021, 13, 445–477. [Google Scholar] [CrossRef]
- Hawkins, S.J.; Moore, P.J.; Burrows, M.T.; Poloczanska, E.; Mieszkowska, N.; Herbert, R.J.; Jenkins, S.R.; Thompson, R.C.; Genner, M.J.; Southward, A.J. Complex interactions in a rapidly changing world: Responses of rocky shore communities to recent climate change. Clim. Res. 2008, 37, 123–133. [Google Scholar] [CrossRef]
- Lamela-Silvarrey, C.; Fernández, C.; Anadón, R.; Arrontes, J. Fucoid assemblages on the north coast of Spain: Past and present (1977–2007). Bot. Mar. 2012, 55, 199–207. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Verlaque, M. Decline and local extinction of Fucales in French Riviera: The harbinger of future extinctions? Mediterr. Mar. Sci. 2015, 16, 206–224. [Google Scholar] [CrossRef]
- Catra, M.; Alongi, G.; Leonardi, R.; Negri, M.; Sanfilippo, R.; Sciuto, F.; Serio, D.; Viola, A.; Rosso, A. Degradation of a photophilic algal community and its associated fauna from eastern Sicily (Mediterranean Sea). Mediterr. Mar. Sci. 2019, 20, 74–89. [Google Scholar] [CrossRef]
- Bevilacqua, S.; Savonitto, G.; Lipizer, M.; Mancuso, P.; Ciriaco, S.; Srijemsi, M.; Falace, A. Climatic anomalies may create a long-lasting ecological phase shift by altering the reproduction of a foundation species. Ecology 2019, 100, e02838. [Google Scholar] [CrossRef]
- Smale, D.A. Impacts of ocean warming on kelp forest ecosystems. N. Phytol. 2020, 225, 1447–1454. [Google Scholar] [CrossRef]
- Perkol-Finkel, S.; Airoldi, L. Loss and Recovery Potential of Marine Habitats: An Experimental Study of Factors Maintaining Resilience in Subtidal Algal Forests at the Adriatic Sea. PLoS ONE 2010, 5, e10791. [Google Scholar] [CrossRef]
- Sala, E.; Ballesteros, E.; Dendrinos, P.; Di Franco, A.; Ferretti, F.; Foley, D.; Fraschetti, S.; Friedlander, A.; Garrabou, J.; Güçlüsoy, H.; et al. The structure of Mediterranean rocky reef ecosystems across environmental and human gradients, and conservation implications. PLoS ONE 2012, 7, e32742. [Google Scholar] [CrossRef]
- Vergés, A.; Steinberg, P.D.; Hay, M.E.; Poore, A.G.B.; Campbell, A.H.; Ballesteros, E.; Heck, K.L., Jr.; Booth, D.J.; Coleman, M.A.; Feary, D.A.; et al. The tropicalization of the temperate marine ecosystems: Climate-mediated changes in herbivory and community phase shifts. Proc. R. Soc. B Biol. Sci. 2012, 281, 20140846. [Google Scholar] [CrossRef]
- Strain, E.M.; Thomson, R.J.; Micheli, F.; Mancuso, F.P.; Airoldi, L. Identifying the interacting roles of stressors in driving the global loss of canopy-forming to mat-forming algae in marine ecosystems. Glob. Change Biol. 2014, 20, 3300–3312. [Google Scholar] [CrossRef]
- Nikolić, V.; Žuljević, A.; Mangialajo, L.; Antolić, B.; Kušpilić, G.; Ballesteros, E. Cartography of littoral rocky-shore communities (CARLIT) as a tool for ecological quality assessment of coastal waters in the Eastern Adriatic Sea. Ecol. Indic. 2013, 34, 87–93. [Google Scholar] [CrossRef]
- Agnetta, D.; Badalamenti, F.; Ceccherelli, G.; Di Trapani, F.; Bonaviri, C.; Gianguzza, P. Role of two co-occurring Mediterranean Sea urchins in the formation of barren from Cystoseira canopy. Estuar. Coast. Shelf Sci. 2015, 152, 73–77. [Google Scholar] [CrossRef]
- Medrano, A.; Linares, C.; Aspillaga, E.; Capdevila, P.; Montero-Serra, I.; Pagès-Escolà, M.; Hereu, B. No-take marine reserves control the recovery of sea urchin populations after mass mortality events. Mar. Environ. Res. 2019, 145, 147–154. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Blanfuné, A.; Pergent, G.; Pergent-Martini, C.; Perret-Boudouresque, M.; Thibaut, T. Impacts of Marine and Lagoon Aquaculture on Macrophytes in Mediterranean Benthic Ecosystems. Front. Mar. Sci. 2020, 7, 218. [Google Scholar] [CrossRef]
- Gianni, F.; Bartolini, F.; Airoldi, L.; Mangialajo, L. Reduction of herbivorous fish pressure can facilitate focal algal species forestation on artificial structures. Mar. Environ. Res. 2018, 138, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Gianni, F.; Mačić, V.; Bartolini, F.; Pey, A.; Laurent, M.; Mangialajo, L. Optimizing canopy-forming algae conservation and restoration with a new herbivorous Fish Deterrent device (DeFish). Restor. Ecol. 2020, 28, 750–756. [Google Scholar] [CrossRef]
- Iveša, L.; Djakovac, T.; Devescovi, M. Long-term fluctuations in Cystoseira populations along the west Istrian Coast (Croatia) related to eutrophication patterns in the northern Adriatic Sea. Mar. Pollut. Bull. 2016, 106, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Personnic, S.; Ruitton, S.; Ballesteros, E.; Bellan-Santini, D.; Bianchi, C.N.; Bussotti, S.; Cebrian, E.; et al. An ecosystem-based approach to assess the status of Mediterranean algae-dominated shallow rocky reefs. Mar. Pollut. Bull. 2017, 117, 311–329. [Google Scholar] [CrossRef]
- Rindi, L.; Dal Bello, M.; Dai, L.; Benedetti-Cecchi, L. Direct observation of increasing recovery length before collapse of a marine benthic ecosystem. Nat. Ecol. Evol. 2017, 1, 0153. [Google Scholar] [CrossRef]
- Rindi, L.; Dal Bello, M.; Benedetti-Cecchi, L. Experimental evidence of spatial signatures of approaching regime shifts in macroalgal canopies. Ecology 2018, 99, 1709–1715. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Pitacco, V.; Lipej, L. Loss of canopy-forming algal richness and coverage in the northern Adriatic Sea. Ecol. Ind. 2021, 125, 107501. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Pitacco, V.; Slavinec, P.; Šiško, M.; Makovec, T.; Falace, A. First Restoration Experiment for Gongolaria barbata in Slovenian Coastal Waters. What Can Go Wrong? Plants 2021, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Savonitto, G.; De La Fuente, G.; Tordoni, E.; Ciriaco, S.; Srijemsi, M.; Bacaro, G.; Chiantore, M.; Falace, A. Addressing reproductive stochasticity and grazing impacts in the restoration of a canopy-forming brown alga by implementing mitigation solutions. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 1611–1623. [Google Scholar] [CrossRef]
- Bernal-Ibáñez, A.; Gestoso, I.; Wirtz, P.; Kaufmann, M.; Serrão, E.A.; Canning-Clode, J.; Cacabelos, E. The collapse of marine forests: Drastic reduction in populations of the family Sargassaceae in Madeira Island (NE Atlantic). Reg. Environ. Change 2021, 21, 71. [Google Scholar] [CrossRef]
- Molinari-Novoa, E.; Guiry, M. Reinstatement of the genera Gongolaria Boehmer and Ericaria Stackhouse (Sargassaceae, Phaeophyceae). Not. Algarum 2020, 172, 1–10. [Google Scholar]
- Ballesteros, E. Production of seaweeds in Northwestern Mediterranean marine communities: Its relation with environmental factors. Sci. Mar. 1989, 53, 357–364. [Google Scholar]
- Sales, M.; Ballesteros, E.; Anderson, M.J.; Iveša, L.; Cardona, E. Biogeographical patterns of algal communities from the Mediterranean Sea: Cystoseira crinita-dominated assemblages as a case study. J. Biogeogr. 2012, 39, 140–152. [Google Scholar] [CrossRef]
- European Union. European Red List of Habitats. Part 1. Marine Habitats; Publications Office of the European Union: Luxembourg, 2016; 52p. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Corsini-Foka, M.; Morri, C.; Zenetos, A. Thirty years after: Dramatic change in the coastal marine ecosystems of Kos Island (Greece), 1981–2013. Mediterr. Mar. Sci. 2014, 15, 482–497. [Google Scholar] [CrossRef]
- Bevilacqua, S.; Airoldi, L.; Ballesteros, E.; Benedetti-Cecchi, L.; Boero, F.; Bulleri, F.; Cebrian, E.; Cerrano, C.; Claudet, J.; Colloca, F.; et al. Chapter one—Mediterranean rocky reefs in the Anthropocene: Present status and future concerns. In Advances of Marine Biology; Sheppard, C., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 89, pp. 1–51. [Google Scholar] [CrossRef]
- Benedetti-Cecchi, L.; Tamburello, L.; Bulleri, F.; Maggi, E.; Gennusa, V.; Miller, M. Linking patterns and processes across scales: The application of scale-transition theory to algal dynamics on rocky shores. J. Exp. Biol. 2012, 215, 977–985. [Google Scholar] [CrossRef]
- Tait, L.W.; Schiel, D.R. Legacy Effects of Canopy Disturbance on Ecosystem Functioning in Macroalgal Assemblages. PLoS ONE 2011, 6, e26986. [Google Scholar] [CrossRef]
- Mačić, V.; Svirčev, Z. Macroepiphytes on Cystoseira species (Phaeophyceae) on the coast of Montenegro. Fresenius Environ. Bull. 2014, 23, 29–34. [Google Scholar]
- Pitacco, V.; Orlando-Bonaca, M.; Mavrič, B.; Popovič, A.; Lipej, L. Mollusc fauna associated with the Cystoseira algal associations in the Gulf of Trieste (northern Adriatic Sea). Mediterr. Mar. Sci. 2014, 15, 225–238. [Google Scholar] [CrossRef]
- Bedini, R.; Bonechi, L.; Piazzi, L. Mobile epifaunal assemblages associated with Cystoseira beds: Comparison between areas invaded and not invaded by Lophocladia lallemandii. Sci. Mar. 2014, 78, 425–432. [Google Scholar] [CrossRef]
- Bianchelli, S.; Buschi, E.; Danovaro, R.; Pusceddu, A. Biodiversity loss and turnover in alternative states in the Mediterranean Sea: A case study on meiofauna. Sci. Rep. 2016, 6, 34544. [Google Scholar] [CrossRef] [PubMed]
- Cheminée, A.; Sala, E.; Pastor, J.; Bodilis, P.; Thiriet, P.; Mangialajo, L.; Cottalorda, J.-M.; Francour, P. Nursery value of Cystoseira forests for Mediterranean rocky reef fishes. J. Exp. Mar. Biol. Ecol. 2013, 442, 70–79. [Google Scholar] [CrossRef]
- Morse, A.N.C. Role of algae in the recruitment of marine invertebrate larvae. In Plant-Animal Interaction in the Marine Benthos; John, D.M., Hawkins, S.J., Price, J.H., Eds.; Clarendon Press: Oxford, UK, 1992; pp. 385–403. [Google Scholar]
- Orlando-Bonaca, M.; Lipej, L. Factors affecting habitat occupancy of fish assemblage in the Gulf of Trieste (Northern Adriatic Sea). Mar. Ecol. 2005, 26, 42–53. [Google Scholar] [CrossRef]
- Thiriet, P.D.; Di Franco, A.; Cheminée, A.; Guidetti, P.; Bianchimani, O.; Basthard-Bogain, S.; Cottalorda, J.-M.; Arceo, H.; Moranta, J.; Lejeusne, P.; et al. Abundance and diversity of crypto- and necto-benthic coastal fish are higher in marine forests than in structurally less complex macroalgal assemblages. PLoS ONE 2016, 11, e0164121. [Google Scholar] [CrossRef]
- Chemello, R.; Milazzo, M. Effect of algal architecture on associated fauna: Some evidence from phytal molluscs. Mar. Biol. 2002, 140, 981–990. [Google Scholar] [CrossRef]
- Poursanidis, D.; Koutsoubas, D.; Arvanitidis, C.; Chatzigeorgiou, G. ReefMedMol: Mollusca from the infralittoral rocky shores—The biocoenosis of photophilic algae—In the Mediterranean Sea. Biodivers. Data J. 2016, 4, e7516. [Google Scholar] [CrossRef]
- Lolas, A.; Antoniadou, C.; Vafidis, D. Spatial variation of molluscan fauna associated with Cystoseira assemblages from a semienclosed gulf in the Aegean Sea. Reg. Stud. Mar. Sci. 2018, 19, 17–24. [Google Scholar] [CrossRef]
- Chiarore, A.; Bertocci, I.; Fioretti, S.; Meccariello, A.; Saccone, G.; Crocetta, F.; Patti, F.P. Syntopic Cystoseira taxa support different molluscan assemblages in the Gulf of Naples (southern Tyrrhenian Sea). Mar. Freshw. Res. 2019, 70, 1561–1575. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Lipej, L.; Orfanidis, S. Benthic macrophytes as a tool for delineating, monitoring and assessing ecological status: The case of Slovenian coastal waters. Mar. Pollut. Bull. 2008, 56, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Water Framework Directive. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 Establishing a Framework for Community Action in the Field of Water Policy; European Parliament: Strasbourg, France, 2000. [Google Scholar]
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; de Torres, M. A new methodology based on littoral community cartography dominated by macroalgae for the implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Orfanidis, S.; Panayotidis, P.; Stamatis, N. Ecological evaluation of transitional and coastal waters: A marine benthic macrophytes-based model. Mediterr. Mar. Sci. 2001, 2, 45–65. [Google Scholar] [CrossRef]
- Orfanidis, S.; Panayotidis, P.; Ugland, K.I. Ecological Evaluation Index continuous formula (EEI-c) application: A step forward for functional groups, the formula and reference condition values. Mediterr. Mar. Sci. 2011, 12, 199–231. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Francé, J.; Mavrič, B.; Grego, M.; Lipej, L.; Flander-Putrle, V.; Šiško, M.; Falace, A. A new index (MediSkew) for the assessment of the Cymodocea nodosa (Ucria) Ascherson meadow’s status. Mar. Environ. Res. 2015, 110, 132–141. [Google Scholar] [CrossRef]
- Section 4: Macroalgae. In WFD Intercalibration Final Report. Part 3—Coastal and Transitional Waters; MED-GIG; European Commission: Luxembourg, 2007; p. 23.
- Marine Strategy Framework Directive. Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 Establishing a Framework for Community Action in the Field of Marine Environmental Policy; European Parliament: Strasbourg, France, 2008.
- Orlando-Bonaca, M.; Lipej, L.; Malej, A.; Francé, J.; Čermelj, B.; Bajt, O.; Kovač, N.; Mavrič, B.; Turk, V.; Mozetič, P.; et al. Selection of Elements to Establish the Monitoring Program of the Marine Environment (Article 11 MSFD); National report in Slovenian; Reports MBP 144; Marine Biology Station Piran, National Institute of Biology: Piran, Slovenia, 2013; 29p. [Google Scholar]
- Orlando-Bonaca, M.; Rotter, A. Any signs of replacement of canopy-forming algae by turf-forming algae in the northern Adriatic Sea? Ecol. Ind. 2018, 87, 272–284. [Google Scholar] [CrossRef]
- Boicourt, W.C.; Ličer, M.; Li, M.; Vodopivec, M.; Malačič, V. Sea state: Recent progress in the context of climate change. In Coastal Ecosystems in Transition: A Comparative Analysis of the Northern Adriatic and Chesapeake Bay, 1st ed.; Malone, T., Malej, A., Faganeli, J., Eds.; Geophysical Monograph Series; American Geophysical Union: Washington, DC, USA; Wiley: Hoboken, NJ, USA, 2021; pp. 21–48. [Google Scholar] [CrossRef]
- Mozetič, P.; Fonda Umani, S.; Cataletto, B.; Malej, A. Seasonal and inter-annual plankton variability in the Gulf of Trieste (northern Adriatic). ICES J. Mar. Sci. 1998, 55, 711–722. [Google Scholar] [CrossRef]
- Stravisi, F. The vertical structure annual cycle of the mass field parameters in the Gulf of Trieste. Boll. Oceanol. Teor. Appl. 1983, 1, 239–250. [Google Scholar]
- Ogorelec, B.; Faganeli, J.; Mišič, M.; Čermelj, B. Reconstruction of paleoenvironment in the bay of Koper (Gulf of Trieste, Northern Adriatic). Ann. Ser. Hist. Nat. 1997, 11, 187–200. [Google Scholar]
- Giovanardi, F.; Francé, J.; Mozetič, P.; Precali, R. Development of ecological classification criteria for the Biological Quality Element phytoplankton for Adriatic and Tyrrhenian coastal waters by means of chlorophyll a (2000/60/EC WFD). Ecol. Ind. 2018, 93, 316–332. [Google Scholar] [CrossRef]
- Mozetič, P.; Francé, J.; Kogovšek, T.; Talaber, I.; Malej, A. Plankton trends and community changes in a coastal sea (northern Adriatic): Bottom-up vs. top-down control in relation to environmental drivers. Estuar. Coast. Shelf. Sci. 2012, 115, 138–148. [Google Scholar] [CrossRef]
- Cozzi, S.; Cabrini, M.; Kralj, M.; De Vittor, C.; Celio, M.; Giani, M. Climatic and Anthropogenic Impacts on Environmental Conditions and Phytoplankton Community in the Gulf of Trieste (Northern Adriatic Sea). Water 2020, 12, 2652. [Google Scholar] [CrossRef]
- Urbini, L.; Ingrosso, G.; Djakovac, T.; Piacentino, S.; Giani, M. Temporal and Spatial Variability of the CO2 System in a Riverine Influenced Area of the Mediterranean Sea, the Northern Adriatic. Front. Mar. Sci. 2020, 7, 679. [Google Scholar] [CrossRef]
- Lipej, L.; Turk, R.; Makovec, T. Endangered Species and Habitat Types in the Slovenian Sea; Institute of the Republic of Slovenia for Nature Conservation: Ljubljana, Slovenia, 2006; 264p. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2022; Available online: https://www.algaebase.org (accessed on 25 June 2022).
- WoRMS Editorial Board. World Register of Marine Species. 2022. Available online: http://www.marinespecies.org (accessed on 12 May 2022).
- Iveša, L.; Djakovac, T.; Bilajac, A.; Gljušćić, E.; Devescovi, M. Increased ammonium levels occurring during benthic algal blooms are potentially toxic to fucalean algae. Bot. Mar. 2021, 64, 267–274. [Google Scholar] [CrossRef]
- Hansen, H.P.; Koroleff, F. Determination of nutrients. In Methods of Seawater Analysis; Wiley: Hoboken, NJ, USA, 1999; pp. 159–228. [Google Scholar]
- Hansen, H.P. In situ determination of pH and oxygen. In Methods of Seawater Analysis; Wiley: Hoboken, NJ, USA, 1999; pp. 399–405. [Google Scholar]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis. Bull. Fish Res. Board. Can. 1968, 167, 1–311. [Google Scholar]
- Clarke, K.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Spearman, C. Demonstration of formulae for true measurement of correlation. Am. J. Psychol. 1907, 18, 161–169. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2015. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Smith, G.M. Analyzing Ecological Data; Springer Science & Business Media: Berlin, Germany, 2007. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Simpson, G.L.; Solymos, P.; Stevens, H.; Legendre, P.; Wagner, H.; Guillaume Blanchet, F. The Vegan Package. Community Ecology Package. 2008. Available online: http://r-forge.r-project.org/projects/vegan/ (accessed on 21 March 2022).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008; Available online: https://www.R-project.org/ (accessed on 20 March 2022).
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The fate of Cystoseira crinita, a forest-forming Fucale (Phaeophyceae, Stramenopiles), in France (North Western Mediterranean Sea). Estuar. Coast. Shelf Sci. 2016, 181, 196–208. [Google Scholar] [CrossRef]
- Cormaci, M.; Furnari, G. Changes of the benthic alga flora of the Tremiti Island (southern Adriatic) Italy. Hydrobiologia 2003, 398/399, 75–79. [Google Scholar] [CrossRef]
- Vukovič, A. Associations of marine benthic algae in the Gulf of Piran. Biol. Vestn. 1980, 28, 103–124. (In Slovenian) [Google Scholar]
- Devescovi, M. Effects of bottom topography and anthropogenic pressure on northern Adriatic Cystoseira spp. (Phaeophyceae, Fucales). Aquat. Bot. 2015, 121, 26–32. [Google Scholar] [CrossRef]
- Orlando-Bonaca, M.; Trkov, D. After more than forty-five years a new finding of Cystoseira foeniculacea f. latiramosa in the coastal sea of Slovenia. Ann. Ser. Hist. Nat. 2020, 30, 233–238. [Google Scholar]
- Avčin, A.; Keržan, I.; Kubik, L.; Meith-Avčin, N.; Štirn, J.; Tušnik, P.; Valentinčič, T.; Vrišer, B.; Vukovič, A. Akvatični ekosistemi v Strunjanskem zalivu I.: Preliminarno poročilo [Aquatic ecosystems of the Strunjan Bay, I.: Preliminary report]. In Akvatični Sistemi v Strunjanskem Zalivu I: Skupno Delo [Aquatic Systems in the Strunjan Bay, I: Collaborative Work]; Prispevki k Znanosti o Morju [Contributions to Marine Science]; Zgodovinsko društvo za južno Primorsko—Koper: Koper, Slovenia, 1973; Volume 5, pp. 168–216. [Google Scholar]
- Falace, A.; Kaleb, S.; De La Fuente, G.; Asnaghi, V.; Chiantore, M. Ex situ cultivation protocol for Cystoseira amentacea var. stricta (Fucales, Phaeophyceae) from a restoration perspective. PLoS ONE 2018, 13, e0193011. [Google Scholar] [CrossRef]
- Mozetič, P.; Solidoro, C.; Cossarini, G.; Socal, G.; Precali, R.; Francé, J.; Bianchi, F.; De Vittor, C.; Smodlaka, N.; Fonda Umani, S. Recent trends towards oligotrophication of the northern Adriatic: Evidence from chlorophyll a time series. Estuar. Coast. 2010, 33, 362–375. [Google Scholar] [CrossRef]
- Solidoro, C.; Bastianini, M.; Bandelj, V.; Codermatz, R.; Cossarini, G.; Melaku Canu, D.; Ravagnan, E.; Salon, S.; Trevisani, S. Current state, scales of variability, and trends of biogeochemical properties in the northern Adriatic Sea. J. Geophys. Res. 2009, 114, C07S91. [Google Scholar] [CrossRef]
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The ups and downs of a canopy-forming seaweed over a span of more than one century. Sci. Rep. 2019, 9, 5250. [Google Scholar] [CrossRef]
- Buñuel, X.; Alcoverro, T.; Pagès, J.F.; Romero, J.; Ruiz, J.M.; Arthur, R. The dominant seagrass herbivore Sarpa salpa shifts its shoaling and feeding strategies as they grow. Sci. Rep. 2020, 10, 10622. [Google Scholar] [CrossRef]
- Iveša, L. Effects of increased seawater temperature and benthic mucilage formation on shallow Cystoseira forests of the West Istrian coast (northern Adriatic Sea). Seventh Eur. Phycol. Congr. Eur. J. Phycol. 2019, 54, 96. [Google Scholar] [CrossRef]
- Bulleri, F.; Eriksson, B.K.; Queirós, A.; Airoldi, L.; Arenas, F.; Arvanitidis, C.; Bouma, T.J.; Crowe, T.P.; Davoult, D.; Guizien, K.; et al. Harnessing positive species interactions as a tool against climate-driven loss of coastal biodiversity. PLoS. Biol. 2018, 16, e2006852. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-H.; Kim, H.; Watson, W.; Di Lorenzo, E.; Sugihara, G. Climate-driven changes in abundance and distribution of larvae of oceanic fishes in the southern California region. Glob. Chang. Biol. 2009, 15, 2137–2152. [Google Scholar] [CrossRef]
- Poulicek, M. Les mollusques des biocénoses à algues photophiles en Méditerranée 2. Analyse du peuplement. (Molluscs of photophilic algal biocenoses in the Mediterranean 2. Analysis of the population). Cah. Biol. Mar. 1985, 26, 127–136. [Google Scholar]
- Vio, E.; De Min, R. Contributo alla conoscenza dei molluschi marini del Golfo di Trieste. (Contribution to the knowledge of marine molluscs in the Gulf of Trieste). Atti Mus. Civ. Stor. Nat. Trieste 1996, 47, 173–233. [Google Scholar]
- Russo, A.R. Epifauna living on sublittoral seaweeds around Cyprus. Hydrobiologia 1997, 344, 169–179. [Google Scholar] [CrossRef]
- Sánchez-Moyano, J.; Estacio, F.; Garcia-Adiego, E.; Garcia-Gómez, J. The molluscan epifauna of the alga Halopteris scoparia in Southern Spain as a bioindicator of coastal environmental conditions. J. Molluscan Stud. 2000, 66, 431–448. [Google Scholar] [CrossRef]
- Antoniadou, C.; Chintiroglou, C. Biodiversity of zoobenthic hard-substrate sublittoral communities in the Eastern Mediterranean (North Aegean Sea). Estuar. Coast. Shelf Sci. 2005, 62, 637–653. [Google Scholar] [CrossRef]
- Chiarore, A.; Fioretti, S.; Meccariello, A.; Saccone, G.; Patti, F.P. Molluscs community associated with the brown algae of the genus Cystoseira in the Gulf of Naples (south Tyrrhenian Sea). BioRxiv 2017, 160200. [Google Scholar] [CrossRef]
- Bitlis, B. Mollusk fauna associated with Cystoseira barbata (Stockhouse) C. Agardh, 1820 in the sea of Marmara (Turkey). Oceanol. Hydrobiol. Stud. 2019, 48, 174–183. [Google Scholar] [CrossRef]
- Poursanidis, D.; Chatzigeorgiou, G.; Charalampos, D.; Koutsoubas, D.; Arvanitidis, C. Testing the robustness of a coastal biodiversity data protocol in the Mediterranean: Insights from the molluskan assemblages from the sublittoral macroalgae communities. Hydrobiologia 2019, 826, 159–172. [Google Scholar] [CrossRef]
- Antoniadou, C.; Koutsoubas, D.; Chintiroglou, C. Mollusca fauna from infralittoral hard substrate assemblages in the North Aegean Sea. Belg. J. Zool. 2005, 135, 119–126. [Google Scholar]
- Myers, J.H. Population cycles: Generalities, exceptions and remaining mysteries. Proc. R. Soc. B 2018, 285, 20172841. [Google Scholar] [CrossRef] [PubMed]
- Hine, P.M. The ecology of Bonamia and decline of bivalve molluscs. N. Z. J. Ecol. 1996, 20, 109–116. [Google Scholar]
- Panarese, R.; Tedesco, P.; Chimienti, G.; Latrofa, M.S.; Quaglio, F.; Passantino, G.; Buonavoglia, C.; Gustinelli, A.; Tursi, A.; Otrant, D. Haplosporidium pinnae associated with mass mortality in endangered Pinna nobilis (Linnaeus 1758) fan mussels. J. Invertebr. Pathol. 2019, 164, 32–37. [Google Scholar] [CrossRef]
- Scarpa, F.; Sanna, D.; Azzena, I.; Cossu, P.; Casu, M. From dark to light and back again: Is Pinna nobilis, the largest Mediterranean shellfish, on the brink of extinction? Veterinaria 2021, 70, 1–14. [Google Scholar]
- Paine, R.T. Ecological determinism in the competition for space: The Robert, H. MacArthur Award Lecture. Ecology 1984, 65, 1339–1348. [Google Scholar] [CrossRef]
- Ávila, S. The littoral molluscs (Gastropoda, Bivalvia and Polyplacophora) of Sao Vicente, Capelas (Sao Miguel Island, Azores): Ecology and biological associations to algae. Soc. Española Malacol. 2003, 21, 11–33. [Google Scholar]
- Gee, J.J.; Warwick, R.M. Metazoan community structure in relation to the fractal dimension of marine macroalgae. Mar. Ecol. Prog. Ser. 1994, 103, 141–150. [Google Scholar] [CrossRef]
- Moore, P.G. The kelp fauna of northeast Britain. I. Introduction and the physical environment. J. Exp. Mar. Biol. Ecol. 1972, 13, 97–125. [Google Scholar] [CrossRef]
- Ogorelec, B.; Mišič, M.; Faganeli, J. Marine geology of the Gulf of Trieste (northern Adriatic): Sedimentological aspects. Mar. Geol. 1991, 99, 79–92. [Google Scholar] [CrossRef]
- Wigham, G.D. The biology of Rissoa parva (da Costa) (Gastropoda: Prosobranchia). J. Mar. Biol. Ass. 1975, 55, 45–68. [Google Scholar] [CrossRef]
- Myers, A.A.; Southgate, T. Artificial substrates as a means of monitoring rocky shore cryptofauna. J. Mar. Biolog. Assoc. 1980, 60, 963–975. [Google Scholar] [CrossRef]
- Southgate, T. Studies on an intertidal population of Rissoa parva (Gastropoda: Prosobranchia) in south-west Ireland. J. Nat. Hist. 1982, 16, 183–194. [Google Scholar] [CrossRef]
- Southgate, T. The biology of Barleeia unifasciata (Gastropoda: Prosobranchia) in red algal turfs in southwest Ireland. J. Mar. Biol. Assoc. 1982, 62, 461–468. [Google Scholar] [CrossRef]
- Sánchez Moyano, J.E.; García Gómez, J.C. The arthropod community, especially Crustacea, as a bioindicator in Algeciras Bay (Southern Spain) based on a spatial distribution. J. Coast. Res. 1998, 14, 1119–1133. [Google Scholar]
- Airoldi, L. The effects of sedimentation on rocky coast assemblages. Oceanogr. Mar. Biol. Annu. Rev. 2003, 41, 161–236. [Google Scholar]
- Schneider, F.I.; Mann, K.H. Species specific relationships of invertebrates to vegetation in a seagrass bed. II. Experiments on the importance of macrophyte shape, epiphyte cover and predation. J. Exp. Mar. Biol. Ecol. 1991, 145, 119–139. [Google Scholar] [CrossRef]
- Borja, A. Biologia y ecologia de tres especies de moluscos gasteropodos intermareales: Rissoa parva, Barleeia unifasciata y Bittium reticulatum. I. Estructura y dinamica de la poblaciones. (Biology and ecology of three species of intertidal gastropod molluscs:: Rissoa parva, Barleeia unifasciata y Bittium reticulatum. I. Population structure and dynamics). Cah. Biol. Mar. 1986, 27, 491–495. [Google Scholar]
- Borja, A. La alimentacion y distribucion del espacio en tres moluscos gasterópodos: Rissoa parva (Da Costa), Barleeia unifasciata (Montagu) y Bittium reticulatum (Da Costa). (Feeding and space allocation in three gastropod molluscs: Rissoa parva (Da Costa), Barleeia unifasciata (Montagu) and Bittium reticulatum (Da Costa)). Cah. Biol. Mar. 1986, 27, 69–75. [Google Scholar]
- Eckman, J.E.; Duggins, D.O. Life and death beneath macrophyte canopies: Effects of understory kelps on growth rates and survival of marine, benthic suspension feeders. Oecologia 1991, 87, 473–487. [Google Scholar] [CrossRef]
- Hicks, G.R.F. Structure of phytal harpacticoid copepod assemblages and the influence of habitat complexity and turbidity. J. Exp. Mar. Biol. Ecol. 1980, 44, 157–192. [Google Scholar] [CrossRef]
- Young, C.M.; Chia, F.S. Microhabitat-associated variability in survival and growth of subtidal solitary ascidians during the first 21 days after settlement. Mar. Biol. 1984, 81, 61–68. [Google Scholar] [CrossRef]
- Hagerman, L. The macro- and micro-fauna associated with Fucus serratus L., with some ecological remarks. Ophelia 1966, 3, 1–43. [Google Scholar] [CrossRef]
- Gunnill, F.C. Effects of plant size and distribution on the numbers of invertebrate species and individuals inhabiting the brown alga Pelvetia fastigiata. Mar. Biol. 1982, 69, 263–280. [Google Scholar] [CrossRef]
- Gibbons, M.J. The impact of sediment accumulation, relative habitat complexity and elevation on rocky shore meiofauna. J. Exp. Mar. Biol. Ecol. 1988, 122, 225–241. [Google Scholar] [CrossRef]
- Ros, J. Distibución batimétrica, abundancia y diversidad de las poblaciones de moluscos bentónicos del litoral catalán. (Bathymetric distribution, abundance and diversity of benthic mollusc populations on the Catalan coastline). Misc. Zool. 1985, 9, 109–126. [Google Scholar]
- Menéndez, M.; Comín, F.A. Spring and summer proliferation of floating macroalgae in a Mediterranean coastal lagoon (Tancadalagoon, Ebro Delta, NE Spain). Estuar. Coast. Shelf Sci. 2000, 51, 215–226. [Google Scholar] [CrossRef]
- Zaabar, W.; Zakhama-Sraieb, R.; Charfi-Cheikhrouha, F.; Achouri, M.S. Composition of a molluscan assemblage associated with macrophytes in Menzel Jemil (Bizerte lagoon, SW Mediterranean Sea). Afr. J. Ecol. 2018, 56, 537–547. [Google Scholar] [CrossRef]
- Berov, D.; Deyanova, D.; Georgieva, I.; Gyosheva, B.; Hiebaum, G. Cystoseira sp. dominated macroalgal communities in the SW Black Sea (Burgas Bay, Bulgaria). Current state and possible long-term effects of eutrophication. Biologie 2012, 65, 821–830. [Google Scholar]
- Vascotto, I.; Mozetič, P.; Francé, J. Phytoplankton Time-Series in a LTER Site of the Adriatic Sea: Methodological Approach to Decipher Community Structure and Indicative Taxa. Water 2021, 13, 2045. [Google Scholar] [CrossRef]
- Mukai, H. The phytal animals on the thalli of Sargassum serratifolium in the Sargassum region, with reference to their seasonal fluctuations. Mar. Biol. 1971, 8, 170–182. [Google Scholar] [CrossRef]
- Rueda, J.L.; Salas, C. Molluscs associated with a subtidal Zostera marina L. bed in southern Spain: Linking seasonal changes of fauna and environmental variables. Estuar. Coast. Shelf Sci. 2008, 79, 157–167. [Google Scholar] [CrossRef]
- Rueda, J.L.; Salas, C. Seasonal variation of a molluscan assemblage living in a Caulerpa prolifera meadow within the inner Bay of Cádiz (SW Spain). Estuar. Coast. Shelf Sci. 2003, 57, 909–918. [Google Scholar] [CrossRef]
- Albrecht, A.; Reise, K. Effects of Fucus vesiculosus covering intertidal mussel beds in the Wadden Sea. Helgoländer Meeresunters. 1994, 48, 243256. [Google Scholar] [CrossRef]
- Antit, M.; Daoulatli, A.; Rueda, J.L.; Salas, C. Temporal variation of the algae-associated molluscan assemblage of artificial substrata in Bay of Tunis (Tunisia). Medit. Mar. Sci. 2013, 14, 390–402. [Google Scholar] [CrossRef]
- Nakaoka, M.; Toyohara, T.; Matsumasa, M. Seasonal and between-substrate variation in mobile epifaunal community in a multispecific seagrass bed of Otsuchi Bay, Japan. Mar. Ecol. 2001, 22, 379–395. [Google Scholar] [CrossRef]
- Urra, J.; Rueda, J.L.; Ramírez, Á.M.; Marina, P.; Tirado, C.; Salas, C.; Gofas, S. Seasonal variation of molluscan assemblages in different strata of photophilous algae in the Alboran Sea (western Mediterranean). J. Sea Res. 2013, 83, 83–93. [Google Scholar] [CrossRef]
- Orabi, O.H.; El-Badru, A.E.-M.A.; Alhejoj, I. Response of mollusk assemblages to environmental conditions: A case study from the Burullus Lagoon, Egypt. J. Geol. Soc. India 2018, 91, 583–588. [Google Scholar] [CrossRef]
- Andrade, R.M.; Santos, I.N.; Oliveira, R. Contribuição para o conhecimento dos criadouros de planorbídeos na área do Distrito Federal: I. Variação de diferentes fatores químicos de suas águas. (Contribution to the knowledge of planorbid breeding sites in the Federal District area: I. Variation of different chemical factors of its waters). Rev. Bras. Malar. 1955, 7, 103–130. [Google Scholar]
- Grisolia, M.L.M.; Freitas, J.F. Características físicas e químicas do habitat da Biomphalaria tenagophila (Mollusca, Planorbidae). (Physical and chemical characteristics of the habitat of Biomphalaria tenagophila Mollusca, Planorbidae)). Mem. Inst. Oswaldo. Cruz. 1985, 80, 237–244. [Google Scholar] [CrossRef]
- Makela, T.P.; Oikari, A.O.J. The effects of low water pH on the ionic balance in the freshwater mussel Anodonta anatina L. Annal. Zool. Fennici. 1992, 29, 169–175. [Google Scholar]
- Pennak, R.W. Freshwater Invertebrates of the United States; John Wiley and Sons Inc.: Hoboken, NJ, USA, 1989; 628p. [Google Scholar]
- Francé, J.; Varkitzi, I.; Stanca, E.; Cozzoli, F.; Skejić, S.; Ungaro, N.; Vascotto, I.; Mozetič, P.; Ninčević Gladan, Ž.; Assimakopoulou, G.; et al. Large-scale testing of phytoplankton diversity indices for environmental assessment in Mediterranean sub-regions (Adriatic, Ionian and Aegean Seas). Ecol. Ind. 2021, 126, 107630. [Google Scholar] [CrossRef]
- Lacoue-Labarthe, T.; Nunes, P.A.L.D.; Ziveri, P.; Cinar, M.; Gazeau, F.; Hall-Spencer, J.M.; Hilmi, N.; Moschella, P.; Safa, A.; Sauzade, D.; et al. Impacts of ocean acidification in a warming Mediterranean Sea: An overview. Reg. Stud. Mar. Sci. 2016, 5, 1–11. [Google Scholar] [CrossRef]
- Dunson, W.A.; Travis, J. The role of abiotic factors in community organization. Am. Nat. 1991, 138, 1067–1091. [Google Scholar] [CrossRef]
- Menge, B.A.; Sutherland, J.P. Community regulation: Variation in disturbance, competition, and predation in relation to gradients of environmental stress and recruitment. Am. Nat. 1987, 130, 730–757. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Lat N | Long E | Date | Sampling Depths (m) |
|---|---|---|---|---|
| 1 | 45°31.571′ | 13°35.385′ | 19 July 2019 | 1.6; 3.7; 5.2 |
| 2 | 45°31.859′ | 13°33.816′ | 23 July 2019 | 2.0; 4.0; 8.8 |
| 3 | 45°31.824′ | 13°34.065′ | 26 July 2019 | 2.2; 4.6; 9.8 |
| 4 | 45°31.604′ | 13°35.049′ | 20 August 2019 | 2.4; 3.8 |
| 5 | 45°31.782′ | 13°34.391′ | 21 August 2019 | 2.8; 5.7; 12.0 |
| 6 | 45°31.732′ | 13°34.466′ | 29 August 2019 | 2.9; 5.3; 10.2 |
| 7 | 45°31.583′ | 13°34.700′ | 17 September 2019 | 3.0; 4.0; 8.0 |
| 8 | 45°32.658′ | 13°40.595′ | 1 July 2020 | 1.9; 2.2; 3.5 |
| 9 | 45°31.672′ | 13°34.580′ | 2 July 2020 | 2.5; 4.5; 8.7 |
| 10 | 45°31.812′ | 13°33.759′ | 6 July 2020 | 2.2; 4.4; 4.8 |
| 11 | 45°32.272′ | 13°37.226′ | 8 July 2020 | 2.2; 4.2; 8.0 |
| 12 | 45°35.478′ | 13°42.133′ | 9 July 2020 | 1.3; 3.0; 5.0 |
| 13 | 45°31.849′ | 13°33.837′ | 16 July 2020 | 2.5; 4.1; 8.2 |
| 14 | 45°32.022′ | 13°38.392′ | 20 July 2020 | 2.1; 4.2; 5.2 |
| 15 | 45°32.913′ | 13°41.523′ | 27 July 2020 | 2.0; 3.5; 4.3 |
| Sampling Site | Sampling Depth (m) | EEI-c | N of CFS | Names of CFS | N of Thalli of CFS | Coverage (%) of CFS |
|---|---|---|---|---|---|---|
| 1 | 1.6 | 9.97 | 1 | G. barbata | 7 | 53 |
| 3.7 | 6.62 | 2 | G. barbata, C. compressa | 3 | 16 | |
| 5.2 | 6.42 | 1 | C. compressa | 2 | 7 | |
| 2 | 2.0 | 10.00 | 2 | H. incurva, C. compressa | 12 | 53 |
| 4.0 | 7.17 | 1 | C. corniculata | 1 | 8 | |
| 8.8 | 6.34 | 1 | C. compressa | 1 | 4 | |
| 3 | 2.2 | 10.00 | 1 | G. barbata | 5 | 60 |
| 4.6 | 9.45 | 3 | G. barbata, C. compressa, C. corniculata | 4 | 19 | |
| 9.8 | 9.55 | 1 | C. corniculata | 2 | 70 | |
| 4 | 2.4 | 4.81 | 1 | H. incurva | 1 | 2 |
| 3.8 | 5.93 | 0 | 0 | 0 | ||
| 5 | 2.8 | 9.89 | 2 | G. barbata, C. corniculata | 3 | 14 |
| 5.7 | 8.63 | 3 | G. barbata, C. compressa, C. corniculata | 4 | 18 | |
| 12.0 | 5.28 | 1 | C. corniculata | 2 | 48 | |
| 6 | 2.9 | 10.00 | 1 | G. barbata | 3 | 38 |
| 5.3 | 10.00 | 2 | G. barbata, C. corniculata | 7 | 62 | |
| 10.2 | 9.12 | 2 | H. incurva, C. corniculata | 7 | 56 | |
| 7 | 3.0 | 8.59 | 1 | G. barbata | 3 | 35 |
| 4.0 | 6.21 | 1 | C. compressa | 5 | 5 | |
| 8.0 | 4.17 | 1 | C. compressa | 1 | 3 | |
| 8 | 1.9 | 10.00 | 2 | H. incurva, C. compressa | 8 | 103 |
| 2.2 | 10.00 | 3 | H. incurva, C. compressa, G. barbata | 11 | 101 | |
| 3.5 | 7.26 | 1 | C. compressa | 1 | 3 | |
| 9 | 2.5 | 10.00 | 2 | G. barbata, C. compressa | 7 | 78 |
| 4.5 | 9.80 | 2 | G. barbata, C. compressa | 8 | 52 | |
| 8.7 | 10.00 | 1 | C. corniculata | 3 | 75 | |
| 10 | 2.2 | 10.00 | 2 | C. compressa, E. crinita | 17 | 116 |
| 4.4 | 5.49 | 1 | C. compressa | 1 | 4 | |
| 4.8 | 2.00 | 1 | C. compressa | 2 | 6 | |
| 11 | 2.2 | 10.00 | 1 | H. incurva | 4 | 70 |
| 4.2 | 8.15 | 1 | C. compressa | 6 | 29 | |
| 8.0 | 8.17 | 2 | C. compressa, C. corniculata | 3 | 9 | |
| 12 | 1.3 | 8.51 | 1 | H. incurva | 6 | 52 |
| 3.0 | 8.72 | 1 | H. incurva | 1 | 4 | |
| 5.0 | 4.19 | 1 | C. compressa | 1 | 2 | |
| 13 | 2.5 | 10.00 | 1 | C. compressa | 18 | 66 |
| 4.1 | 10.00 | 2 | C. compressa, C. corniculata | 9 | 32 | |
| 8.2 | 7.52 | 1 | C. corniculata | 3 | 21 | |
| 14 | 2.1 | 9.49 | 3 | G. barbata, C. compressa, E. crinita | 6 | 40 |
| 4.2 | 10.00 | 1 | H. incurva | 6 | 75 | |
| 5.2 | 6.34 | 0 | 0 | 0 | ||
| 15 | 2.0 | 10.00 | 2 | H. incurva, C. compressa | 12 | 80 |
| 3.5 | 7.55 | 1 | Cystoseira foeniculacea f. latiramosa | 1 | 2 | |
| 4.3 | 7.39 | 1 | Cystoseira foeniculacea f. latiramosa | 2 | 25 |
| ESG I | ESG II | EEI-c | Algal cov | Algal vol | N Cysto | N Canopy | Smol | Nmol | J’ | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ESG II | rs | −0.51 | |||||||||
| p | <0.05 | ||||||||||
| EEI-c | rs | 0.94 | −0.73 | ||||||||
| p | <0.05 | <0.05 | |||||||||
| Algal cov | rs | 0.83 | −0.99 | 0.67 | |||||||
| p | <0.05 | NS | <0.05 | ||||||||
| Algal vol | rs | 0.55 | −0.25 | 0.51 | 0.66 | ||||||
| p | <0.05 | NS | <0.05 | <0.05 | |||||||
| N Cysto | rs | 0.45 | −0.36 | 0.43 | 0.28 | 0.38 | |||||
| p | <0.05 | <0.05 | <0.05 | NS | <0.05 | ||||||
| N canopy | rs | 0.66 | −0.72 | 0.67 | 0.41 | 0.36 | 0.77 | ||||
| p | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | |||||
| Smol | rs | −0.34 | 0.023 | −0.064 | 0.023 | 0.051 | −0.33 | −0.33 | |||
| p | NS | NS | NS | NS | NS | NS | NS | ||||
| Nmol | rs | −0.051 | −0.077 | −0.097 | 0.02 | 0.032 | −0.33 | 0.29 | 0.89 | ||
| p | NS | NS | NS | NS | NS | NS | NS | <0.05 | |||
| J’ | rs | −0.018 | −0.089 | −0.059 | 0.56 | −0.69 | 0.015 | −0.055 | −0.41 | −0.74 | |
| p | NS | NS | NS | NS | NS | NS | NS | <0.05 | <0.05 | ||
| H’ | rs | −0.027 | −0.005 | −0.51 | 0.031 | 0.008 | −0.38 | −0.38 | 0.95 | 0.76 | −0.16 |
| p | NS | NS | NS | NS | NS | NS | NS | <0.05 | <0.05 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlando-Bonaca, M.; Trkov, D.; Klun, K.; Pitacco, V. Diversity of Molluscan Assemblage in Relation to Biotic and Abiotic Variables in Brown Algal Forests. Plants 2022, 11, 2131. https://doi.org/10.3390/plants11162131
Orlando-Bonaca M, Trkov D, Klun K, Pitacco V. Diversity of Molluscan Assemblage in Relation to Biotic and Abiotic Variables in Brown Algal Forests. Plants. 2022; 11(16):2131. https://doi.org/10.3390/plants11162131
Chicago/Turabian StyleOrlando-Bonaca, Martina, Domen Trkov, Katja Klun, and Valentina Pitacco. 2022. "Diversity of Molluscan Assemblage in Relation to Biotic and Abiotic Variables in Brown Algal Forests" Plants 11, no. 16: 2131. https://doi.org/10.3390/plants11162131
APA StyleOrlando-Bonaca, M., Trkov, D., Klun, K., & Pitacco, V. (2022). Diversity of Molluscan Assemblage in Relation to Biotic and Abiotic Variables in Brown Algal Forests. Plants, 11(16), 2131. https://doi.org/10.3390/plants11162131