
New Monocyclic Terpenoid Lactones from a Brown Algae Sargassum macrocarpum as Monoamine Oxidase Inhibitors

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Bioassay-Guided Isolation of Compounds
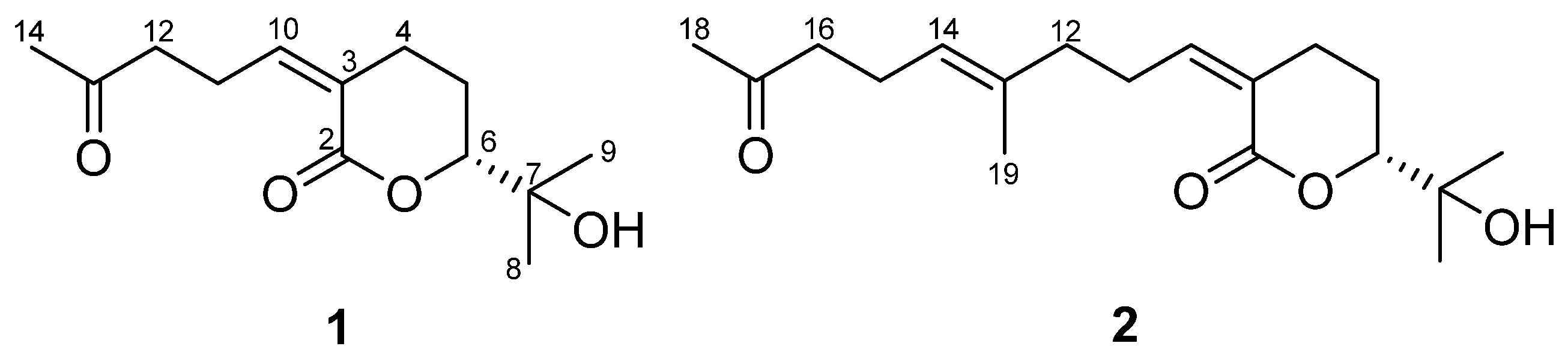
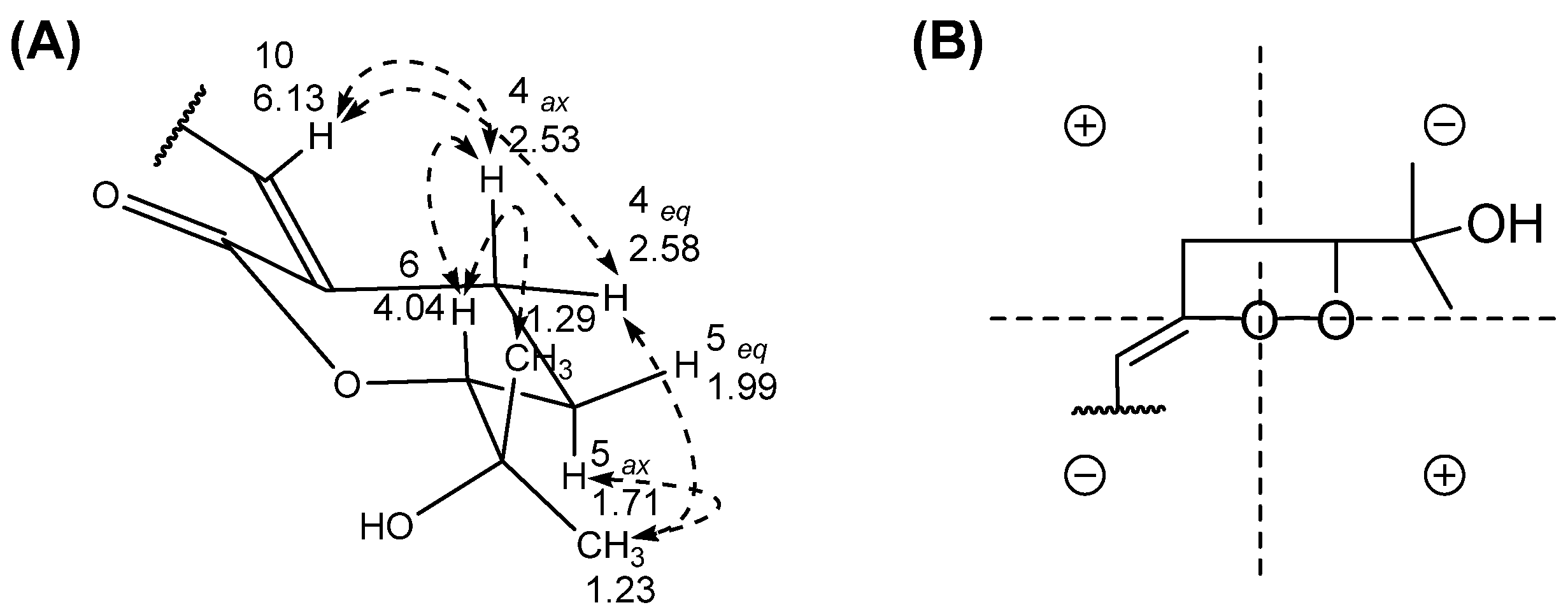
2.2. Structural Characterization of Compounds
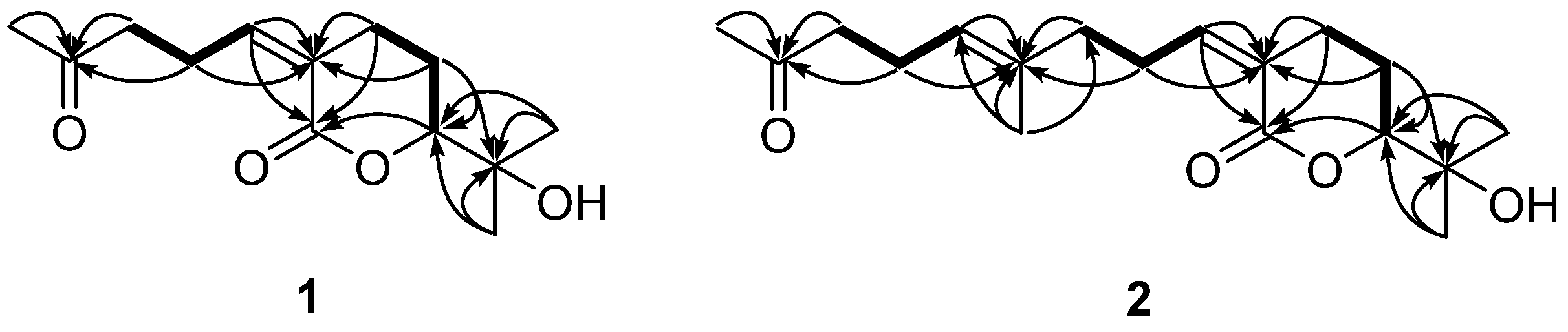
2.3. Absolute Configuration Determination of Compounds
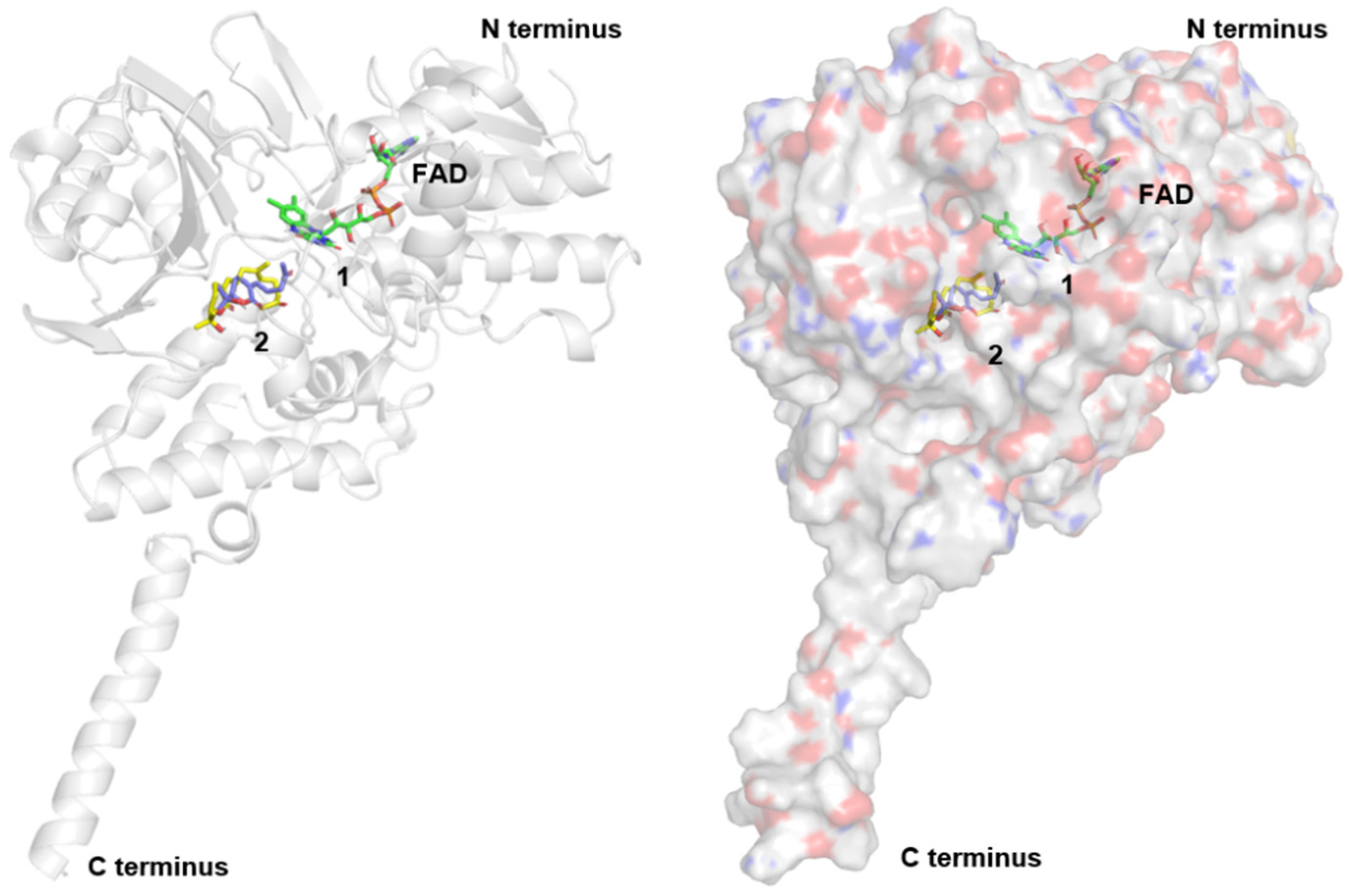
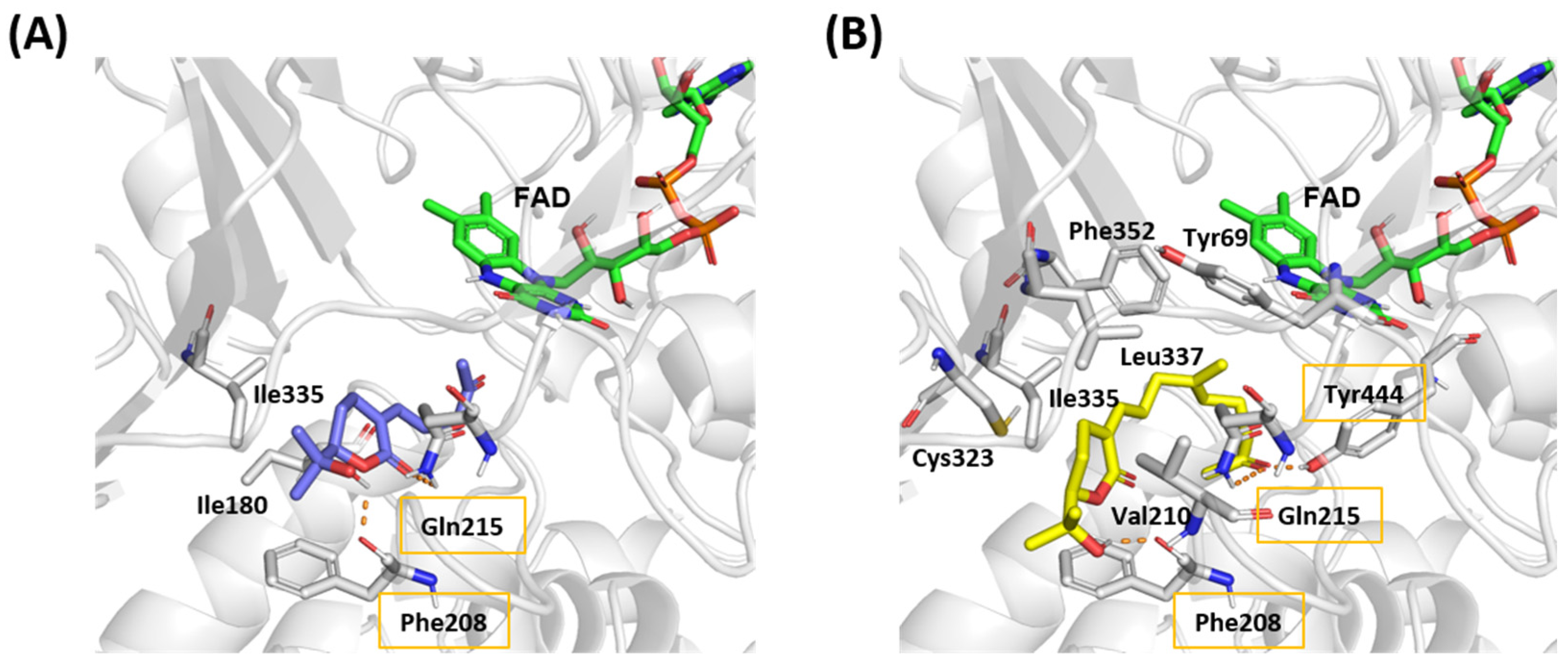
2.4. MAO Activities and Molecular Docking of the Isolated Compounds
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction
4.2. General Experimental Procedures
4.3. Sepbox Application for Subfraction of S. macrocarpum
4.4. Purification of Compounds 1 and 2
4.4.1. Sargassumin A (1)
4.4.2. Sargassumin B (2)
4.5. hMAO Assay of Compounds 1 and 2
4.6. Structural Preparation and Molecular Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Ko, S.J.; Kim, Y.K.; Hong, S.W.; Kang, M.S.; Park, C.S.; Hwang, E.K.; Lee, Y.D. Artificial seed production and cultivation of Sargassum macrocarpum (Fucales, Phaeophyta). Algae 2020, 35, 123–131. [Google Scholar] [CrossRef]
- Burke, W.J.; Li, S.W.; Chung, H.D.; Ruggiero, D.A.; Kristal, B.S.; Johnson, E.M.; Lampe, P.; Kumar, V.B.; Franko, M.; Williams, E.A. Neurotoxicity of MAO metabolites of catecholamine neurotransmitters: Role in neurodegenerative diseases. Neurotoxicology 2004, 25, 101–115. [Google Scholar] [CrossRef]
- Wood, P.L.; Khan, M.A.; Moskal, J.R. The concept of “aldehyde load” in neurodegenerative mechanisms: Cytotoxicity of the polyamine degradation products hydrogen peroxide, acrolein, 3-aminopropanal, 3-acetamidopropanal and 4-aminobutanal in a retinal ganglion cell line. Brain Res. 2007, 1145, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.-M.; Chen, K.; Shih, J.C. Monoamine oxidase A and repressor R1 are involved in apoptotic signaling pathway. Proc. Natl. Acad. Sci. USA 2006, 103, 10923–10928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shulman, K.I.; Herrmann, N.; Walker, S.E. Current place of monoamine oxidase inhibitors in the treatment of depression. CNS Drugs 2013, 27, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Tzvetkov, N.T.; Stammler, H.-G.; Neumann, B.; Hristova, S.; Antonov, L.; Gastreich, M. Crystal structures, binding interactions, and ADME evaluation of brain penetrant N-substituted indazole-5-carboxamides as subnanomolar, selective monoamine oxidase B and dual MAO-A/B inhibitors. Eur. J. Med. Chem. 2017, 127, 470–492. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Roy, A.; Jung, J.H.; Choi, J.S. Evaluation of the inhibitory effects of eckol and dieckol isolated from edible brown alga Eisenia bicyclis on human monoamine oxidases A and B. Arch. Pharm. Res. 2017, 40, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.H.; Lee, B.H.; Choi, B.W.; Lee, H.-S.; Shin, J. Chromenes from the brown alga Sargassum siliquastrum. J. Nat. Prod. 2005, 68, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.-R.; Huang, S.-T.; Liaw, C.-C.; Yen, M.-H.; Hwang, T.-L.; Chen, C.-Y.; Hou, M.-F.; Yuan, S.-S.; Cheng, Y.-B.; Wu, Y.-C. Diterpenes from Grangea maderaspatana. Phytochemistry 2016, 131, 124–129. [Google Scholar] [CrossRef]
- Musa, M.A.; Badisa, V.L.; Aghimien, M.O.; Eyunni, S.V.; Latinwo, L.M. Identification of 7, 8-dihydroxy-3-phenylcoumarin as a reversible monoamine oxidase enzyme inhibitor. J. Biochem. Mol. Toxicol. 2021, 35, e22651. [Google Scholar] [CrossRef] [PubMed]
- Pflüger, P.; Pereira, P.; Loza, M.I.; Brea, J.; Viña, D.; Kumar, A.; Fontenla, J.A. Gamma-decanolactone: Preliminary evaluation as potential antiparkinsonian drug. Eur. J. Pharmacol. 2021, 906, 174276. [Google Scholar] [CrossRef]
- Son, S.-Y.; Ma, J.; Kondou, Y.; Yoshimura, M.; Yamashita, E.; Tsukihara, T. Structure of human monoamine oxidase A at 2.2-Å resolution: The control of opening the entry for substrates/inhibitors. Proc. Natl. Acad. Sci. USA 2008, 105, 5739–5744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Colibus, L.; Li, M.; Binda, C.; Lustig, A.; Edmondson, D.E.; Mattevi, A. Three-dimensional structure of human monoamine oxidase A (MAO A): Relation to the structures of rat MAO A and human MAO B. Proc. Natl. Acad. Sci. USA 2005, 102, 12684–12689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repicč, M.; Purg, M.; Vianello, R.; Mavri, J. Examining electrostatic preorganization in monoamine oxidases A and B by structural comparison and p K a calculations. J. Phys. Chem. B 2014, 118, 4326–4332. [Google Scholar] [CrossRef] [PubMed]
- Dang, T.T.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Technol. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Kolanjinathan, K.; Ganesh, P.; Saranraj, P. Pharmacological importance of seaweeds: A review. World J. Fish Mar. Sci. 2014, 6, 1–15. [Google Scholar]
- Osório, C.; Machado, S.; Peixoto, J.; Bessada, S.; Pimentel, F.B.; Alves, R.C.; Oliveira, M. Pigments content (chlorophylls, fucoxanthin and phycobiliproteins) of different commercial dried algae. Separations 2020, 7, 33. [Google Scholar] [CrossRef]
- Hakim, M.M.; Patel, I.C. A review on phytoconstituents of marine brown algae. Futur. J. Pharm. Sci. 2020, 6, 1–11. [Google Scholar] [CrossRef]
- La Barre, S.; Potin, P.; Leblanc, C.; Delage, L. The halogenated metabolism of brown algae (Phaeophyta), its biological importance and its environmental significance. Mar. Drugs 2010, 8, 988–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, C.K.; Kamei, Y. Sargaquinoic acid supports the survival of neuronal PC12D cells in a nerve growth factor-independent manner. Eur. J. Pharmacol. 2004, 488, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Joung, E.-J.; Cao, L.; Lee, B.; Gwon, W.-G.; Park, S.-H.; Kim, H.-R. Sargahydroquinoic acid, a cyclooxygenase-2 inhibitor, attenuates inflammatory responses by regulating NF-κB Inactivation and Nrf2 activation in lipopolysaccharide-stimulated cells. Inflammation 2021, 44, 2120–2131. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Kim, J.; Lee, K.M.; Choi, Y.-J.; Ye, B.-R.; Kim, M.-S.; Ko, S.-G.; Lee, S.-H.; Kang, D.-H.; Heo, S.-J. Tuberatolide B suppresses cancer progression by promoting ROS-mediated inhibition of STAT3 signaling. Mar. Drugs 2017, 15, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Kinghorn, A.D. Development of potential antitumor agents from the scaffolds of plant-derived terpenoid lactones. J. Med. Chem. 2020, 63, 15410–15448. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.H.; Paudel, P.; Choi, J.-W.; Ahn, D.H.; Nam, T.-J.; Jung, H.A.; Choi, J.S. Probing multi-target action of phlorotannins as new monoamine oxidase inhibitors and dopaminergic receptor modulators with the potential for treatment of neuronal disorders. Mar. Drugs 2019, 17, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 (Chloroform-d) | 2 (Acetone-d6) | ||
|---|---|---|---|---|
| δC | δH | δC | δH | |
| 1 | ||||
| 2 | 165.2 | 165.7 | ||
| 3 | 125.3 | 126.8 | ||
| 4 | 28.6 | 2.58, m | 29.4 | 2.56, m |
| 2.53, m | ||||
| 5 | 23.4 | 1.99, ddt (13.5, 5.5, 2.5) | 24.2 | 2.08, m |
| 1.71, dtd (13.5, 12.0, 6.0) | 1.68, dtd (13.5, 11.5, 7.0) | |||
| 6 | 85.8 | 4.04, dd (12.0, 3.0) | 86.3 | 4.01, dd (12.0, 3.0) |
| 7 | 71.4 | 71.2 | ||
| 8 | 24.2 | 1.23, s | 25.4 | 1.20 |
| 9 | 25.7 | 1.29, s | 26.5 | 1.20 |
| 10 | 146.0 | 6.13, br tt (7.5, 2.0) | 146.1 | 6.03, br tt (7.0, 2.0) |
| 11 | 24.0 | 2.83, br dtt (7.0, 7.0, 1.0) | 28.5 | 2.66, dtt (7.0, 7.0, 1.0) |
| 12 | 42.8 | 2.63, br t (7.0) | 39.6 | 2.08, m |
| 13 | 208.1 | 135.9 | ||
| 14 | 29.7 | 2.16, s | 124.8 | 5.13, br tq (7.0, 1.0) |
| 15 | 23.1 | 2.22, dt (7.0, 7.0) | ||
| 16 | 43.8 | 2.47, t (7.0) | ||
| 17 | 207.8 | |||
| 18 | 30.7 | 2.09, s | ||
| 19 | 16.0 | 1.61, s | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, J.; Lee, K.; Hwang, H.; Kim, S.-H.; Park, S.E.; Durai, P.; Park, K.; Kim, H.-S.; Jang, D.S.; Choi, J.S.; et al. New Monocyclic Terpenoid Lactones from a Brown Algae Sargassum macrocarpum as Monoamine Oxidase Inhibitors. Plants 2022, 11, 1998. https://doi.org/10.3390/plants11151998
Kwon J, Lee K, Hwang H, Kim S-H, Park SE, Durai P, Park K, Kim H-S, Jang DS, Choi JS, et al. New Monocyclic Terpenoid Lactones from a Brown Algae Sargassum macrocarpum as Monoamine Oxidase Inhibitors. Plants. 2022; 11(15):1998. https://doi.org/10.3390/plants11151998
Chicago/Turabian StyleKwon, Jaeyoung, Kyerim Lee, Hoseong Hwang, Seong-Hwan Kim, Se Eun Park, Prasannavenkatesh Durai, Keunwan Park, Hyung-Seop Kim, Dae Sik Jang, Jae Sue Choi, and et al. 2022. "New Monocyclic Terpenoid Lactones from a Brown Algae Sargassum macrocarpum as Monoamine Oxidase Inhibitors" Plants 11, no. 15: 1998. https://doi.org/10.3390/plants11151998
APA StyleKwon, J., Lee, K., Hwang, H., Kim, S.-H., Park, S. E., Durai, P., Park, K., Kim, H.-S., Jang, D. S., Choi, J. S., & Kwon, H. C. (2022). New Monocyclic Terpenoid Lactones from a Brown Algae Sargassum macrocarpum as Monoamine Oxidase Inhibitors. Plants, 11(15), 1998. https://doi.org/10.3390/plants11151998