
Studies Regarding the Pharmaceutical Potential of Derivative Products from Plantain


 , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Bioproducts
- (a)
- A quantity of 500 g of plantain powder firstly was extracted with 5000 mL of distilled water, by boiling under reflux. The aqueous extract was concentrated at 250 mL at 50 °C, using a rotary evaporator (Heidolph, Schwabach, Germany); then, the concentrate was treated with 1250 mL of 96% methanol solvent. The resulting precipitate was then dissolved into 250 mL distilled water and then treated with 1250 mL methanol (the operation was repeated twice). The final precipitate was dried in the oven (Gallenkamp, UK) at 45 °C and resulted in a fine gray powder, which was considered as the polysaccharidic fraction (PP).
- (b)
- The solid vegetal material from the first extraction was re-extracted with the me-thanolic solution resulting from the polysaccharides precipitation (approx. 4500 mL total volume) for 1 h, at boiling temperature under reflux, in a continuous stirring system operated at 300 rpm. The resulting methanolic extract was concentrated at solid residue at 50 °C using a rotary evaporator after that, the residue was dissolved into 250 mL of distilled water. The aqueous solution was further extracted (three times) with 250 mL ethyl acetate, at boiling, under reflux, and the combined ethyl acetate fractions were concentrated in a rotary evaporator at 50 °C. The residue obtained was dissolved into ethanol and precipitated with ethyl ether. The final precipitate was considered the flavonoidic fraction (PF).
- (c)
- The three aqueous fractions resulting from (a) and (b) were combined and then filtrated on the active charcoal. The resulting aqueous extract was concentrated in a rotary evaporator at 50 °C and the obtained solid residue was considered the iridoidic fraction (PI).
2.2. Regents Used in HPTLC Analysis
2.3. Qualitative Determination
2.4. Antioxidant Activity Estimation
2.5. Antifungal Activities
2.5.1. Strains Used in Antifungal Studies
- (a)
- micromycetes: Aspergillus niger ATTC 1015, (A. niger), Aspergillus terreus ATTC 1012 (A. terreus), Penicillium citrinum ATTC 10105 (P. citrinum), Penicillium digitatum ATTC 9849 (P. digitatum), Penicillium sp. 1, and Penicillium sp. 2 (the last two species were isolated from infected wood).
- (b)
- dermatophytes: Microsporum canis ATTC 10214 (M. canis), Trichophyton mentagrophytes ATTC 18748 (T. mentagrophytes), Microsporum gypseum ATTC 24102 (M. gypseum), and Scopulariopsis brevicaulis ATTC 1102 (S. brevicaulis).
2.5.2. Methodology Used in Antifungal Studies
2.6. Antitumor Activities
2.6.1. Cell Viability Assay by Flow Cytometry Analysis
2.6.2. Cell Cultures
2.6.3. Apoptosis Assay—Annexin V-FITC/PI Double Staining
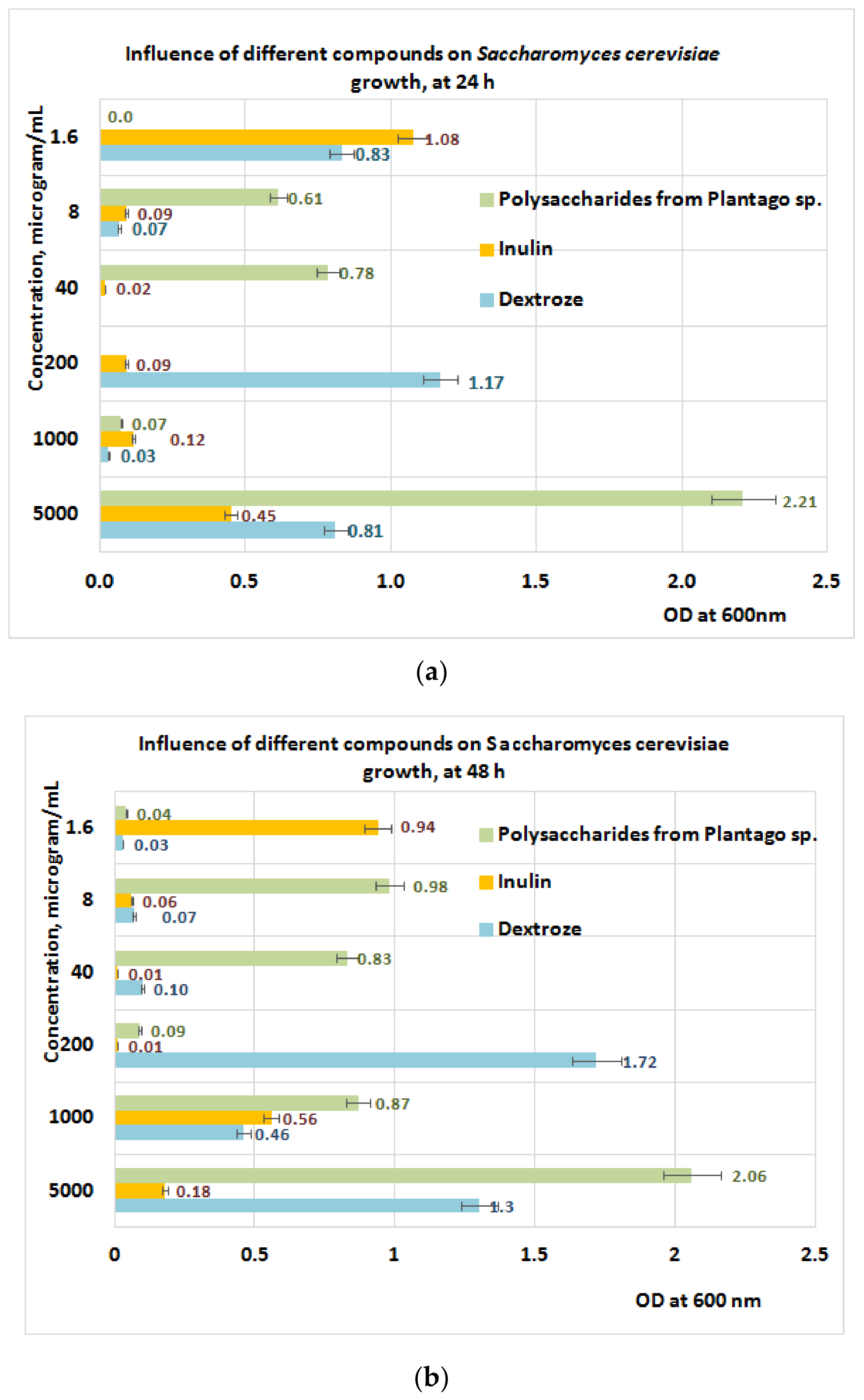
2.7. Prebiotic Activities
2.7.1. Microorganisms
2.7.2. Methodology Used in Prebiotic Activities
3. Results
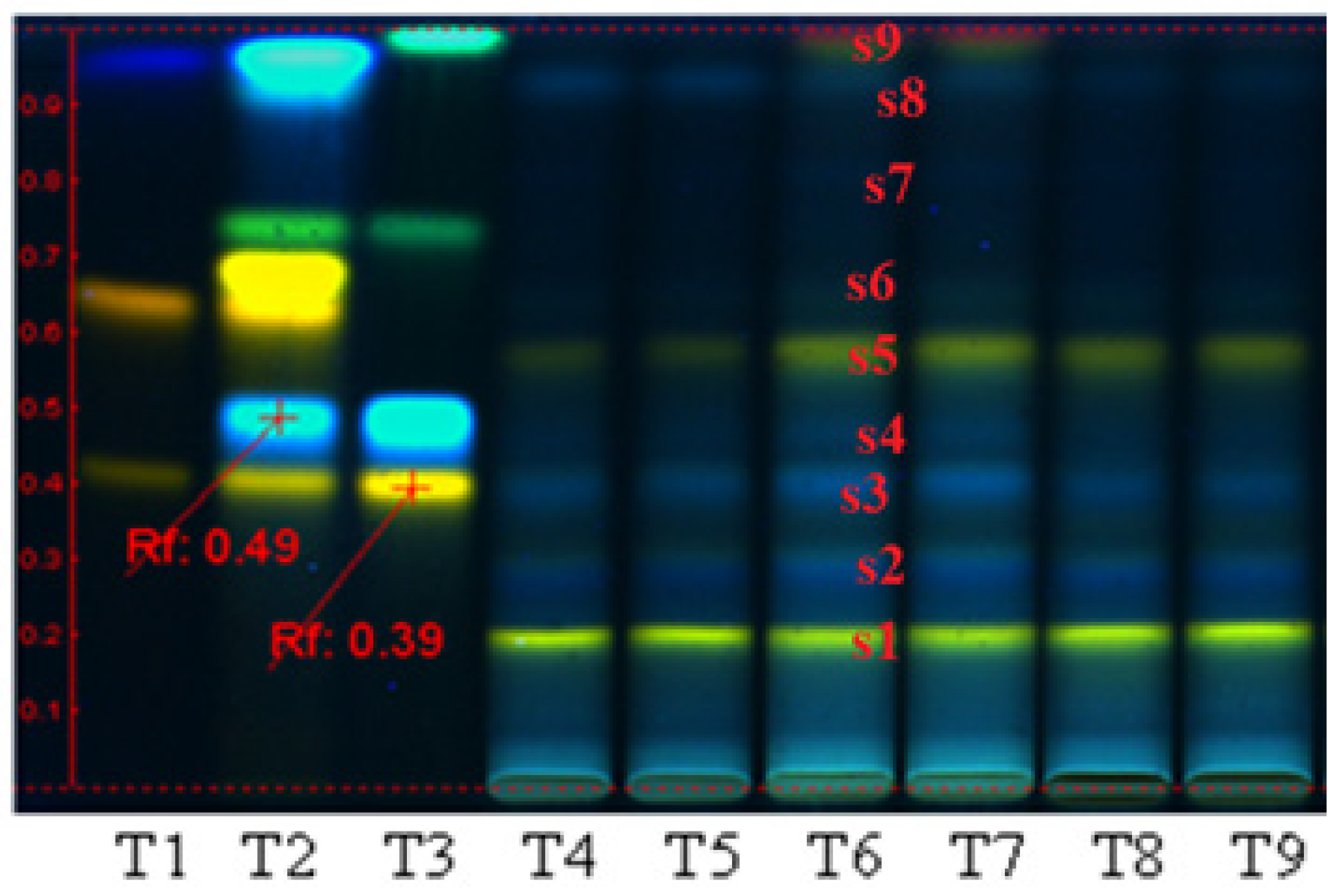
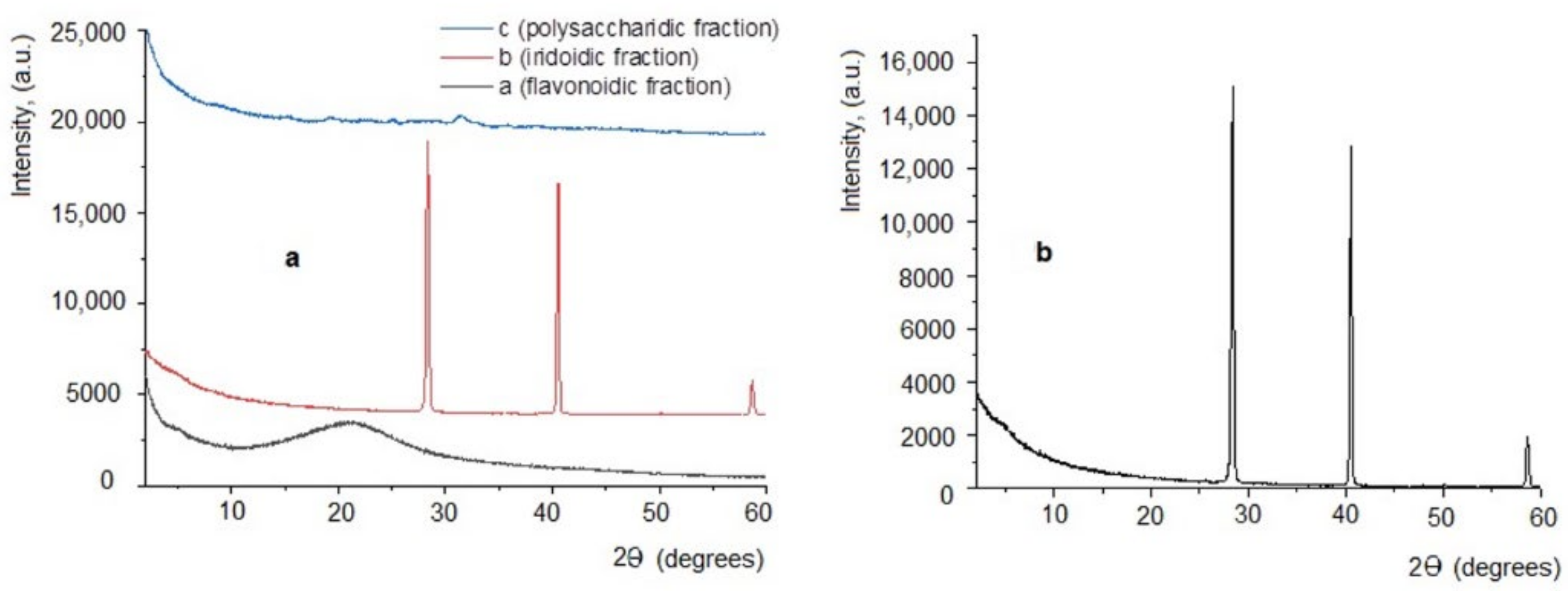
3.1. Plant Bioproduct Characterization
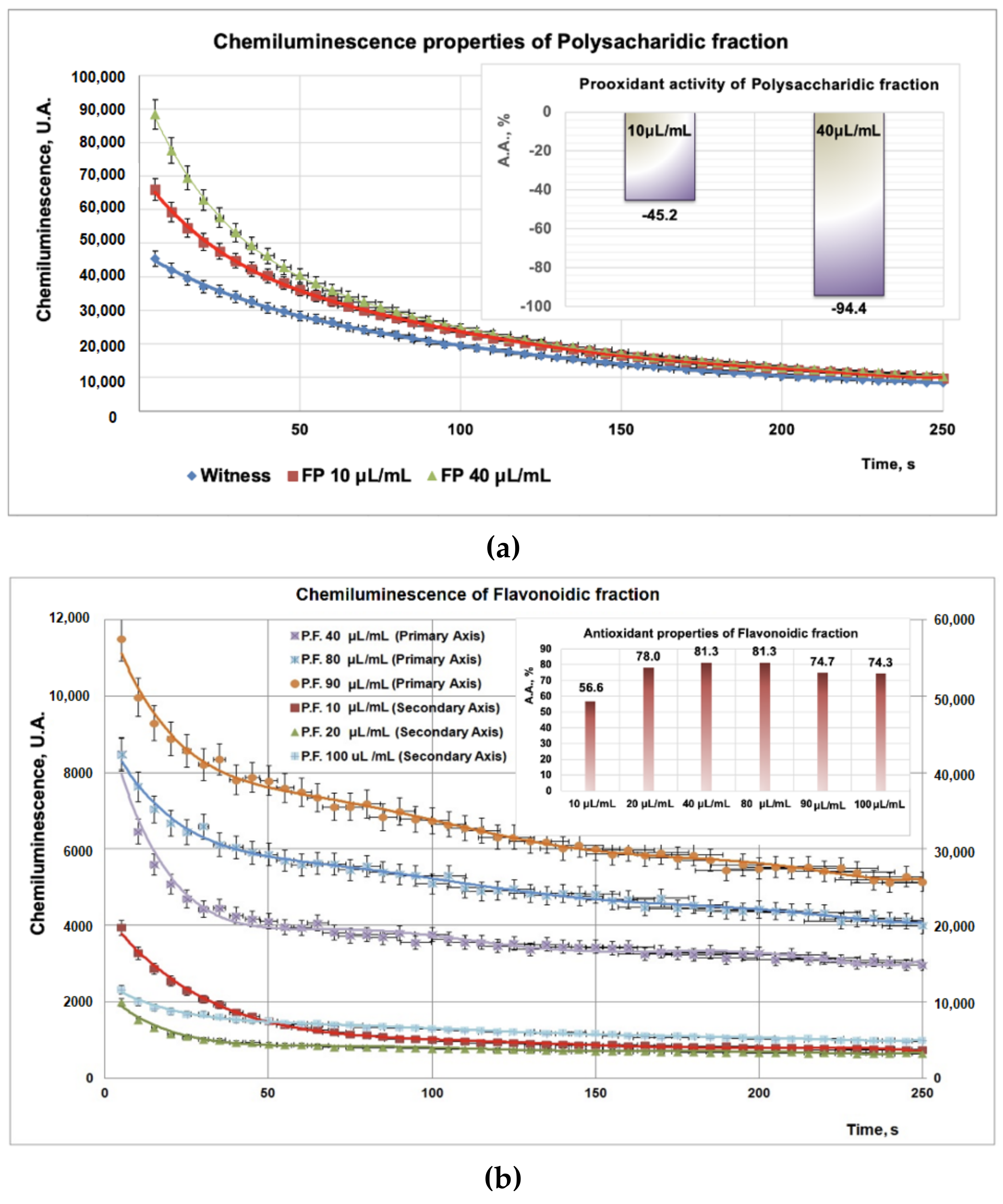
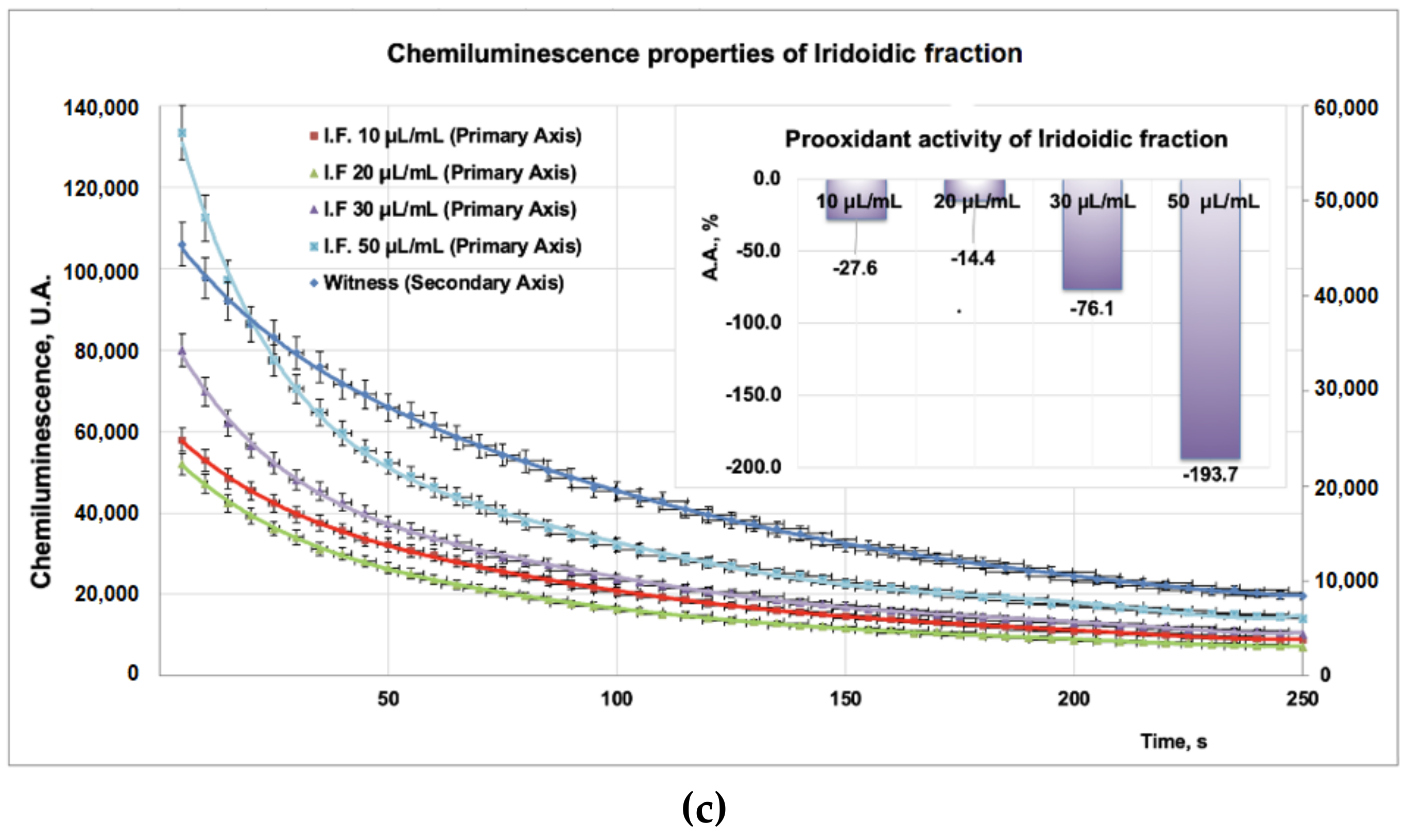
3.2. Antioxidant Properties
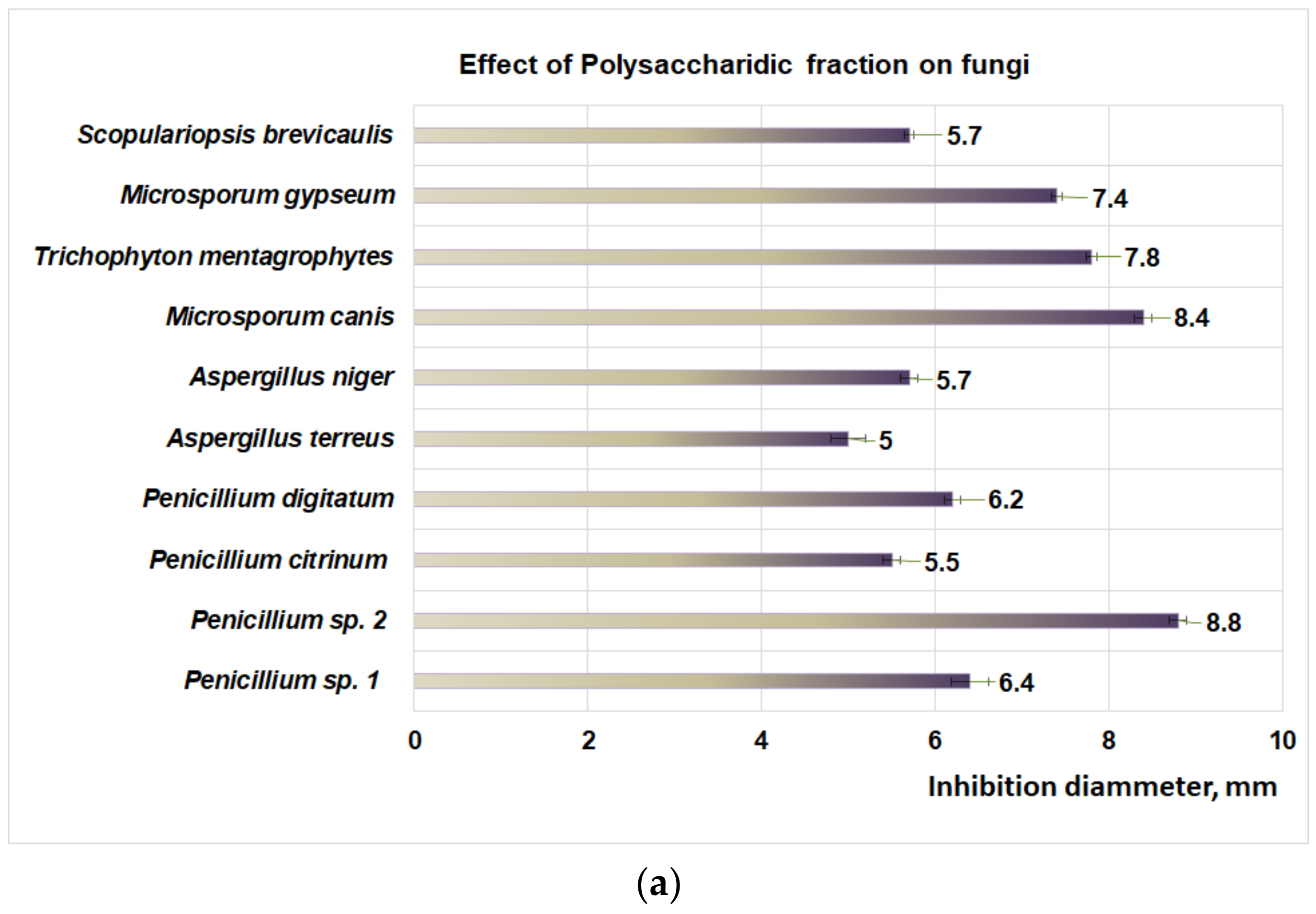
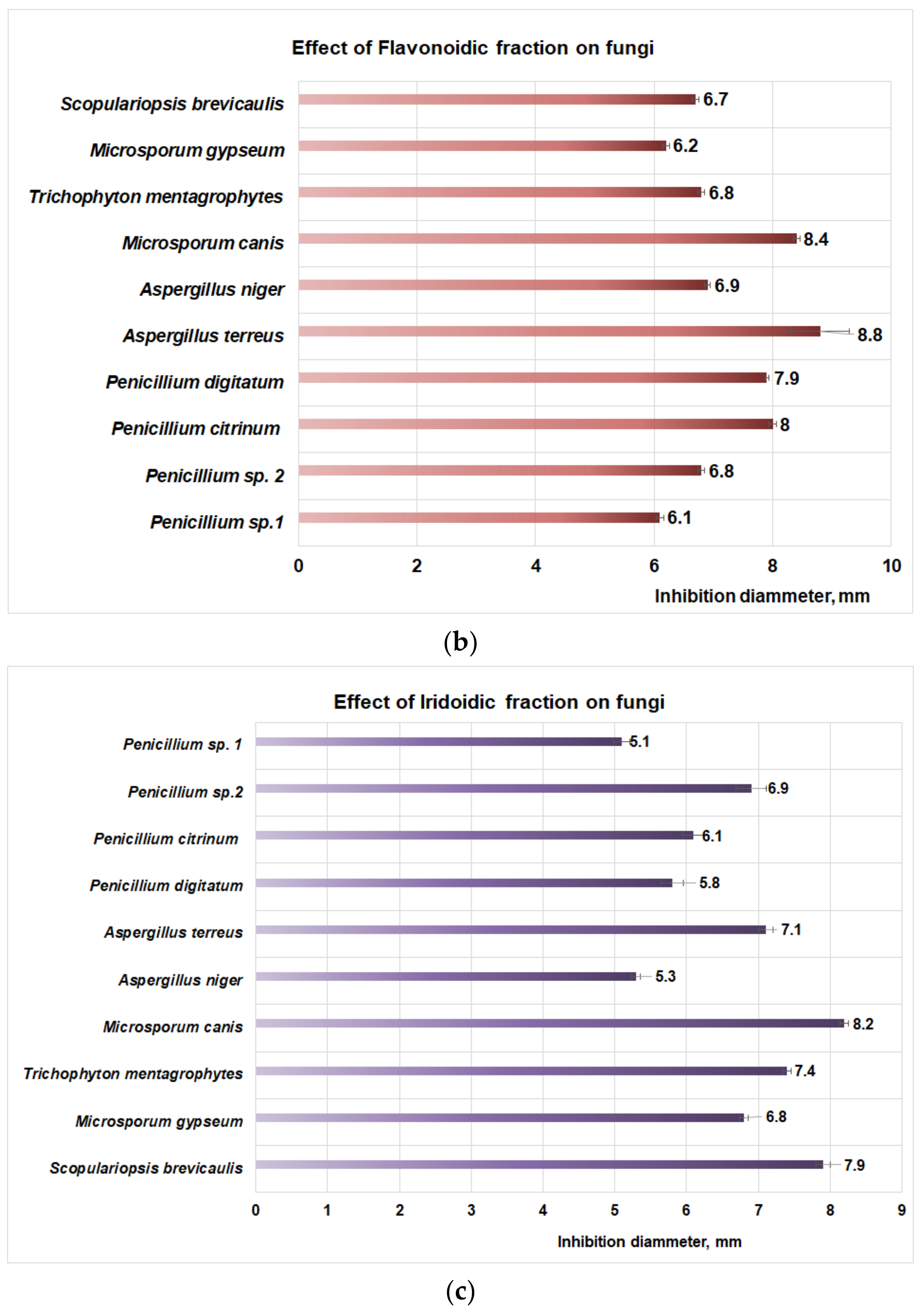
3.3. Antifungal Properties of Plantago Bioproducts
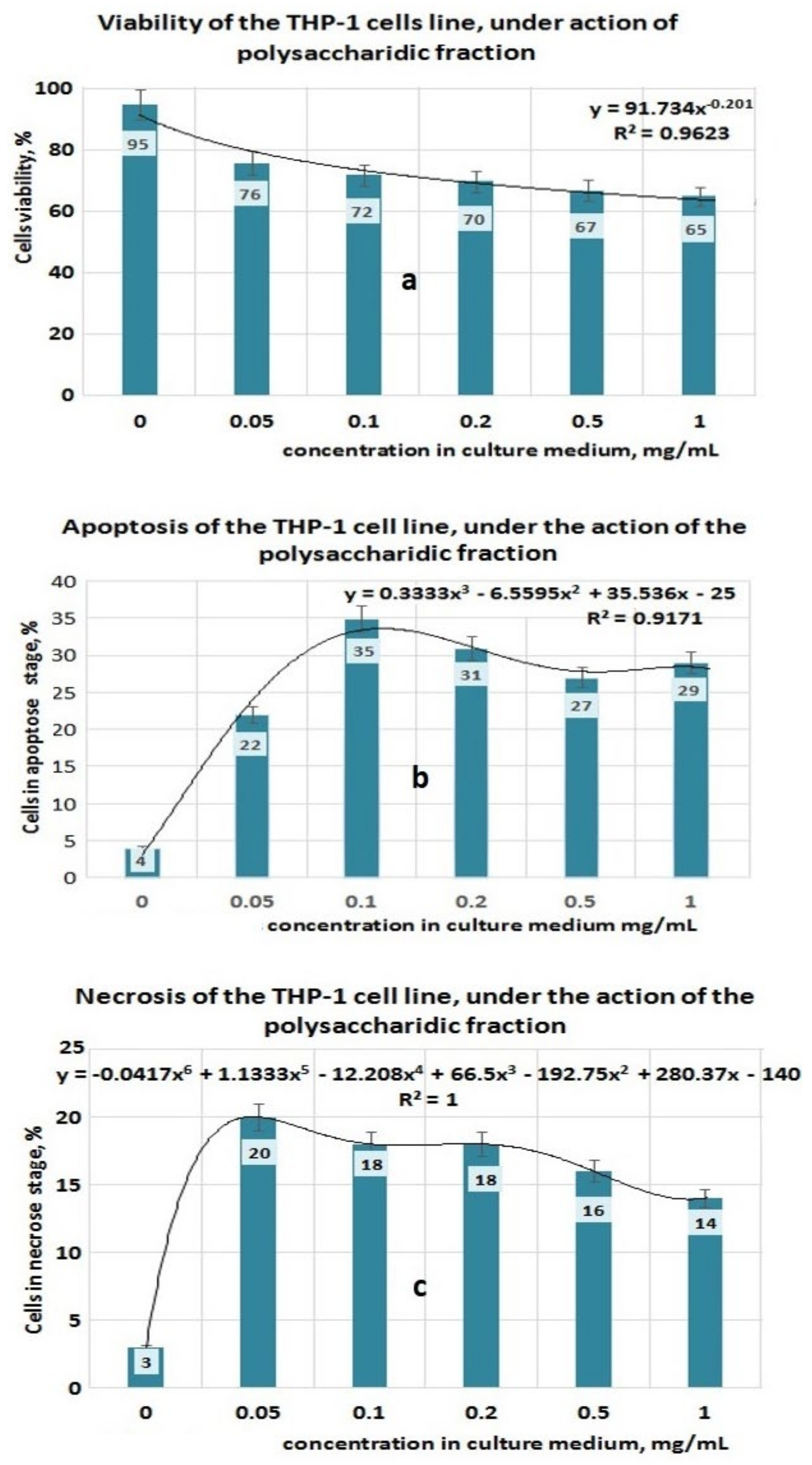
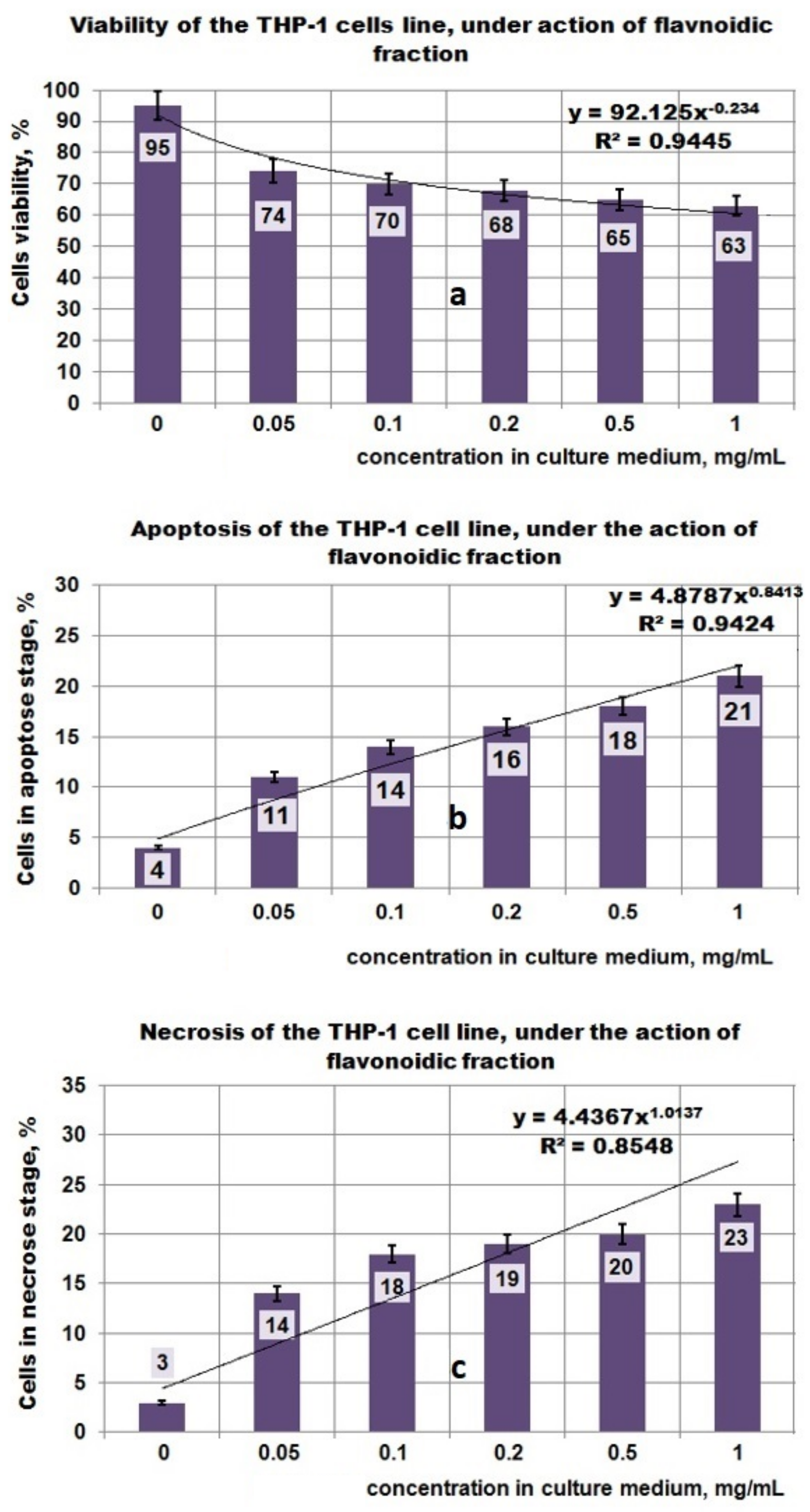
3.4. Antitumor Properties of Plantago Bioproducts
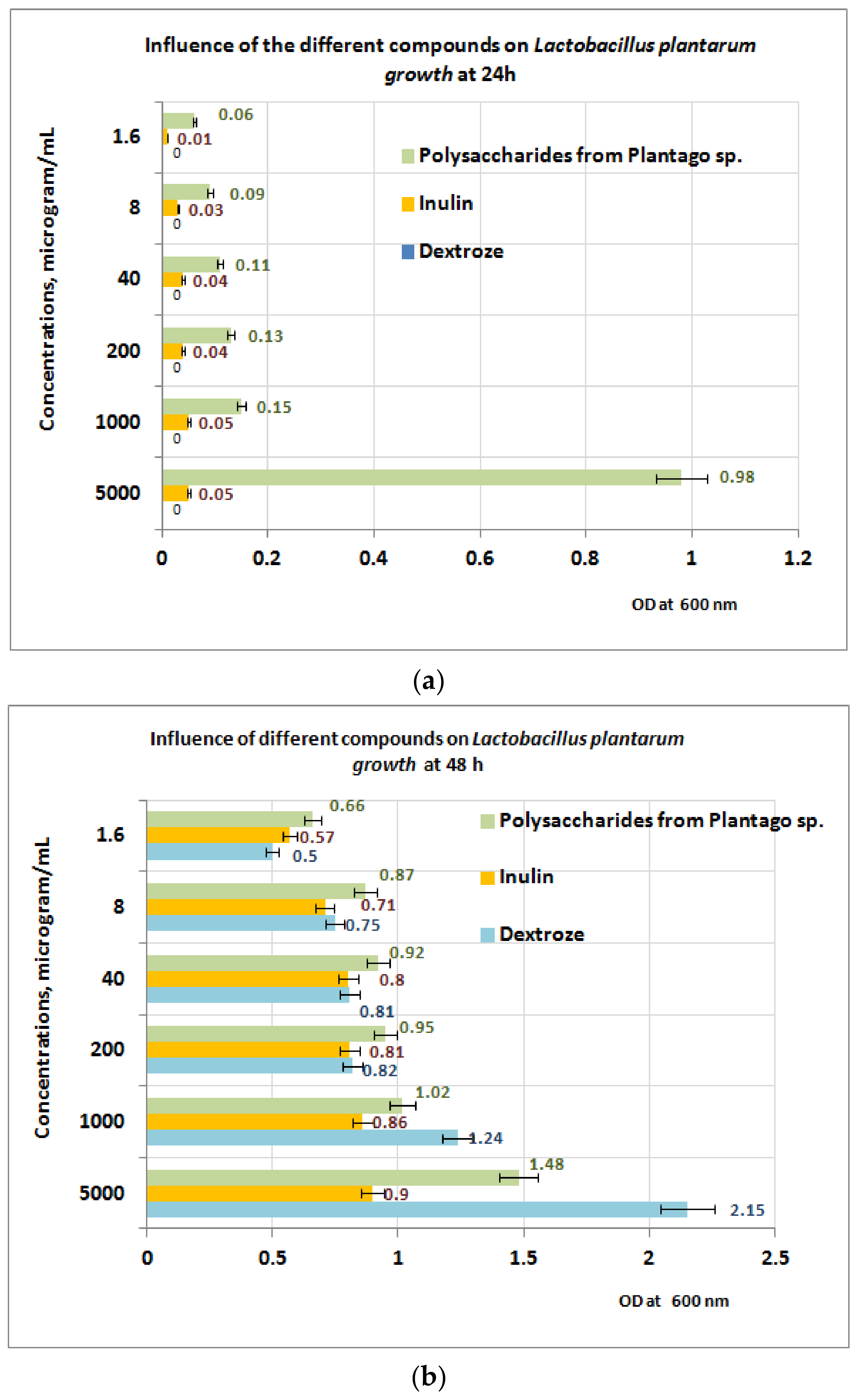
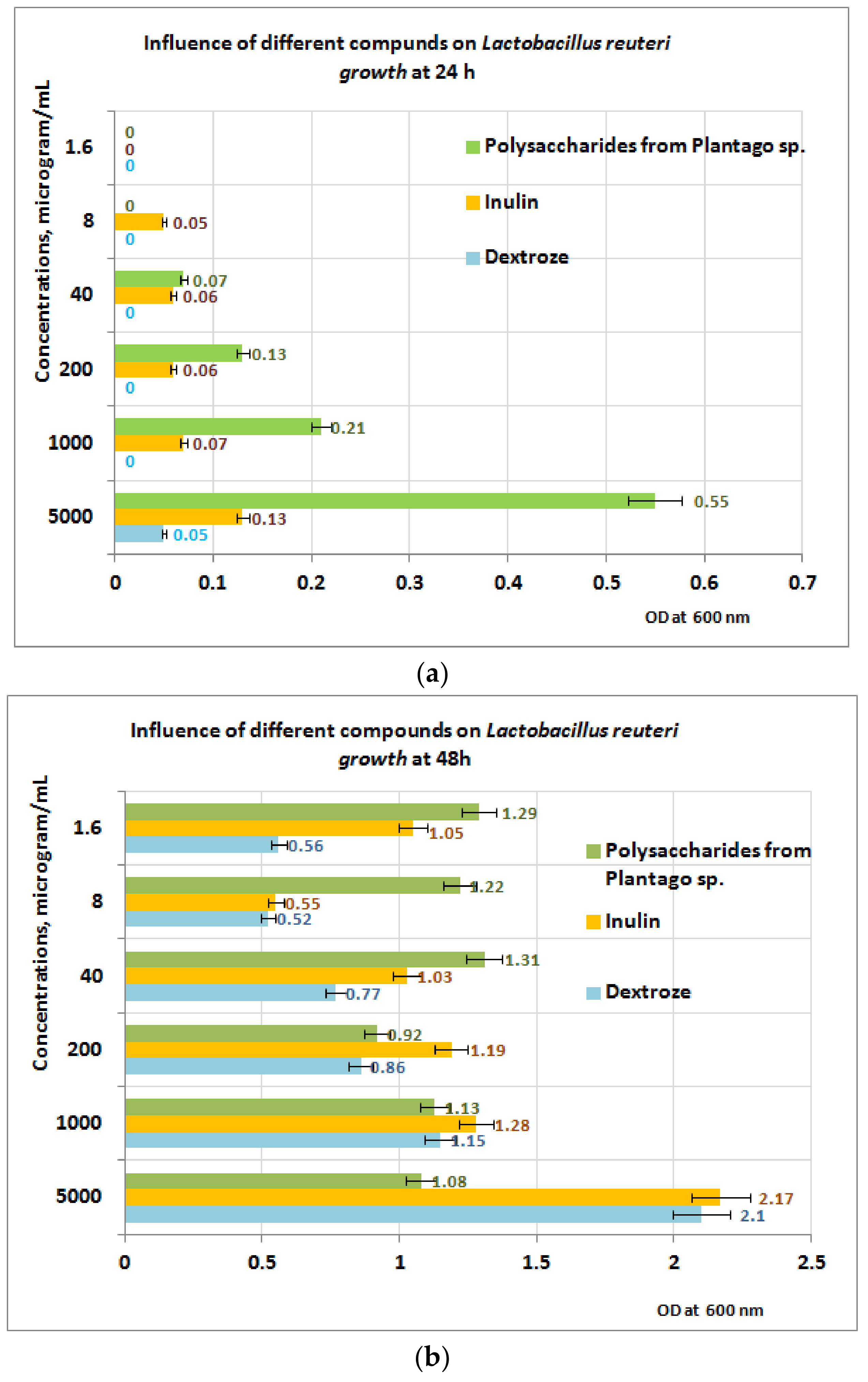
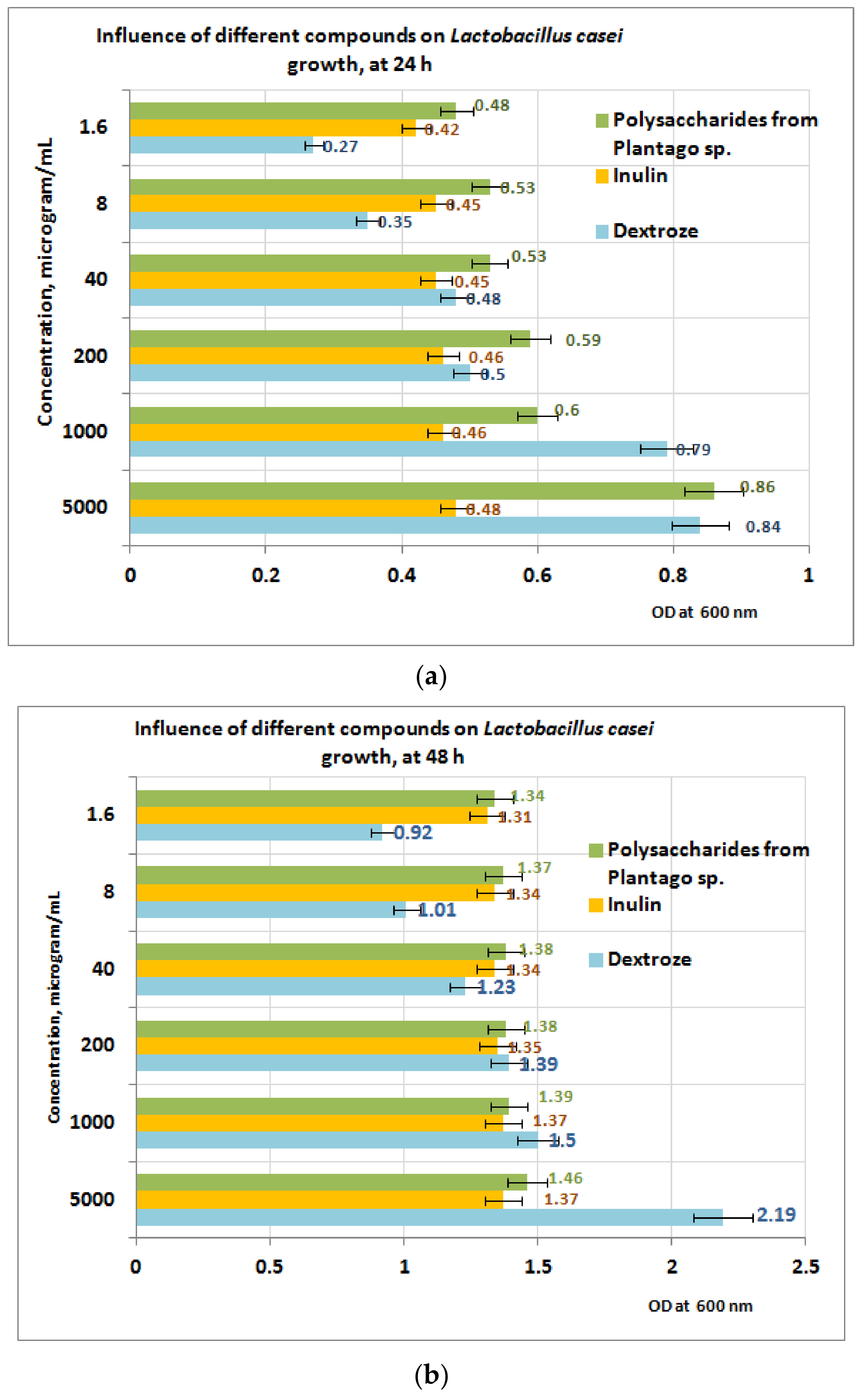
3.5. Prebiotic Activity of Plantain Bioproducts
4. Discussion
- -
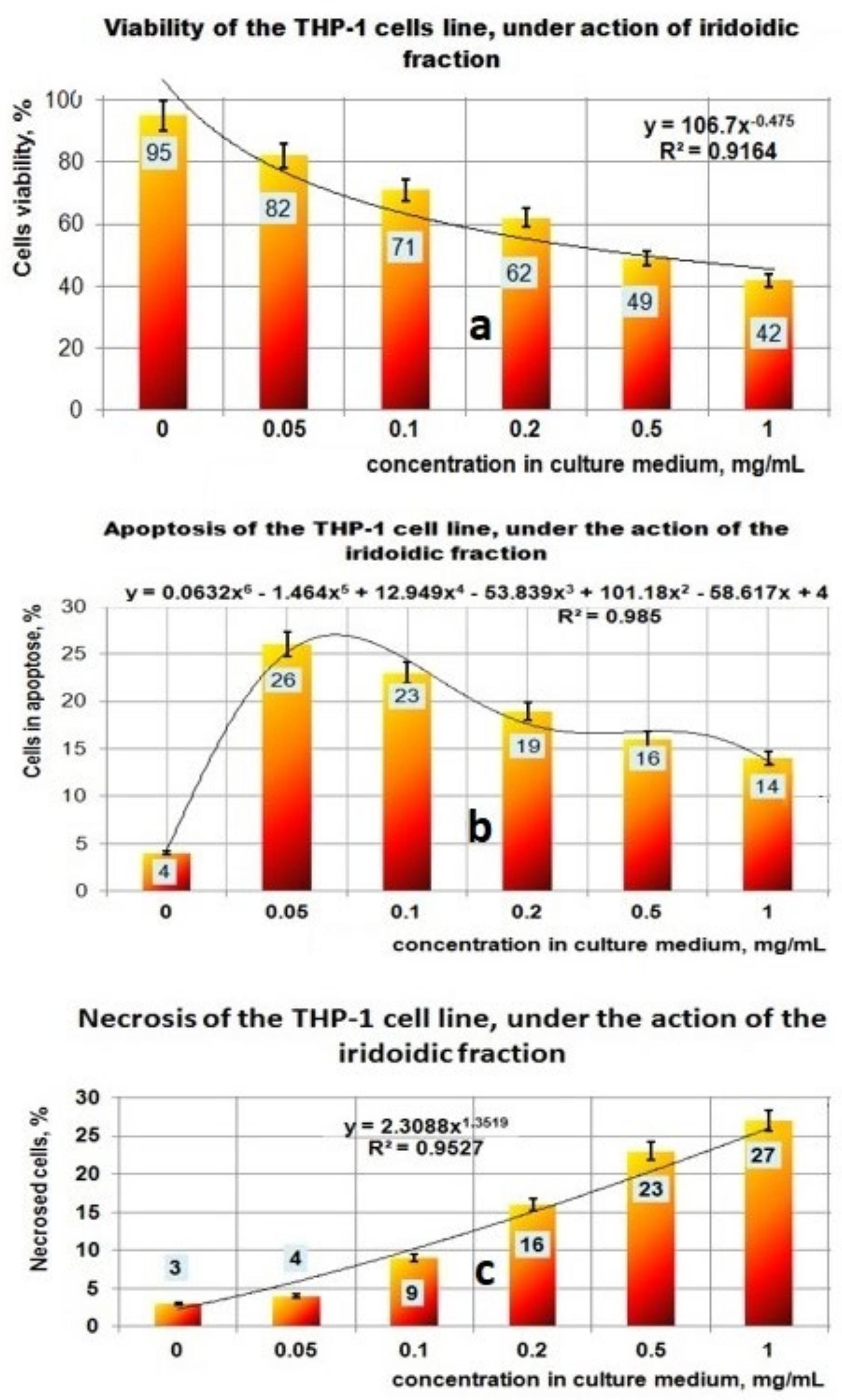
- the PP fraction reduced the cell viability of the THP-1 tumor cell line in a concentration-dependent manner. This PP fraction has antitumor properties which are sustained by its ability to induce apoptosis or necrosis of THP-1 tumor cells;
- -
- in the case of the PF fraction, the antitumor effect seems to be higher, this being supported by the mathematical analysis of the results which shows a significant increase in both the apoptotic process and the necrosis that correlates with the decrease in viability;
- -
- the fraction containing iridoidic compounds (PI) has a strong antitumor effect on THP-1 cells, which is demonstrated by the significant decrease of cell viability depending on the increase in concentration and by the amplification of apoptosis and necrosis processes leading to tumor cell death.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| MRS | cultivation media for Lactobacillus species |
| YPG | cultivation media that contain yeast extract, peptone, and glucose |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl (i.e., 2,2-Diphenyl-1-(2,4,6-trinitrophenyl)hydrazin-1-yl) |
| GAE | Galic Acid equivalents |
| MRC-5 | diploid cell culture line composed of fibroblasts |
| HeLa | cervical cancer cell line |
| MCF-7 | human breast cancer cell line |
| HT-9 | colorectal tumor cell line |
| HL-60 | promyelocytic leukemia cell line |
| K562 | human chronic myeloid leukemia cell line |
| CCRF-CEM | acute lymphoblastic leukemia cell line |
| P3HRI | Burkitt lymphoma cells |
| P338 | murine leukemia cell line |
| Bax | proapoptotic protein; apoptosis regulator |
| Bcl-2 | proapoptotic protein apoptosis regulator |
| p53 | tumor suppressor gene |
| p21 | protein which regulates cell proliferation by inhibiting the cell cycle through the cyclin kinase pathway |
| G1 | cell phase in which the cell grows physically larger copies of organelles and makes the molecular building blocks it will need in later steps |
| G0 | cell phase also known as the resting phase is the phase of the cell cycle during which a cell is neither dividing nor preparing to divide |
References
- Adom, M.B.; Taher, M.; Mutalabisin, M.F.; Amri, M.S.; Kudos, M.B.A.; Sulaiman, M.W.A.W.; Sengupta, P.; Susanti, D. Chemical constituents and medical benefits of Plantago major. Biomed. Pharmacother. 2017, 96, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Bajer, T.; Janda, V.; Bajerová, P.; Kremr, D.; Eisner, A.; Ventura, K. Chemical composition of essential oils from Plantago lanceolata L. leaves extracted by hydrodistillation. J. Food Sci. Technol. 2016, 53, 1576–1586. [Google Scholar] [CrossRef] [Green Version]
- Varban, R.; Varban, D. Comparative study of the active ingredient content Plantago lanceolata L. ProEnvironment 2012, 5, 248–250. [Google Scholar]
- Pirvu, L.C.; Nita, S.; Rusu, N.; Bazdoaca, C.; Neagu, G.; Bubueanu, C.; Udrea, M.; Udrea, R.; Enache, A. Effects of Laser Irradiation at 488, 514, 532, 552, 660, and 785 nm on the Aqueous Extracts of Plantago lanceolata L.: A Comparison on Chemical Content, Antioxidant Activity and Caco2 Viability. Appl. Sci. 2022, 12, 5517. [Google Scholar] [CrossRef]
- Beara, I.N.; Lesjak, M.M.; Orčić, D.Z.; Simin, N.Đ.; Četojević-Simin, D.D.; Božin, B.N.; Mimica-Dukić, N.M. Comparative analysis of phenolic profile, antioxidant, anti-inflammatory, and cytotoxic activity of two closely related Plantago species: Plantago altissima L. and Plantago lanceolata L. LWF Food Sci. Technol. 2012, 47, 64–70. [Google Scholar] [CrossRef]
- Nichita, C.; Neagu, G.; Cucu, A.; Virginia Vulturescu, V.; Berteşteanu, S.V.G. Antioxidative Properties of Plantago Lanceolata L. Extracts Evaluated by Chemiluminescence Method. AgroLife Sci. J. 2016, 5, 95–102. [Google Scholar]
- Bahadori, M.B.; Sarikurkcu, C.; Kocak, M.S.; Calapoglu, M.; Uren, M.C.; Ceylan, O. Plantago lanceolata as a source of health beneficial phytochemicals: Phenolic profile and antioxidant capacity. Food Biosci. 2020, 34, 100536. [Google Scholar] [CrossRef]
- Sanna, F.; Piluzza, G.; Campesi, G.; Molinu, M.G.; Re, G.A.; Sulas, L. Antioxidant Contents in a Mediterranean Population of Plantago lanceolata L. Exploited for Quarry Reclamation Interventions. Plants 2022, 11, 791. [Google Scholar] [CrossRef]
- Karadeniz, A.; Alexie, G.; Greten, H.J.; Andersch, K.; Efferth, T. Cytotoxicity of medicinal plants of the west Canadian which in Native Americans towards sensitive and multidrug-resistant cancer cells. J. Ethnopharmacol. 2015, 168, 191–200. [Google Scholar] [CrossRef]
- Kardosova, A. Polysaccharides from the Leaves of Plantago lanceolata L., var. LIBOR: An a-D-Glucan. Chem. Pap. 1992, 46, 127–130. [Google Scholar]
- Grigore, A.; Bubueanu, C.; Pirvu, L.; Ionita, L.; Toba, G. Plantago lanceolata L. Crops—Source of valuable raw materials for various industrial applications. Sci. Pap. Ser. A Agron. 2015, 57, 207–214. [Google Scholar]
- Lukova, P.K.; Karcheva-Barkevanka, D.P.; Nikolova, M.M.; Iliev, I.I.; Mladenov, R.D. Comparison of structure and antioxidant activity of polysaccharides from the leaves of Plantago major L., P. media and P. lanceolata. Bulg. Chem. Commun. 2017, 49, 282–288. [Google Scholar]
- Irmgard, U.; John-Deesbach, W. Essential and trace element content of medicinal plants and their infusions. Botanica 1984, 58, 255–266. [Google Scholar]
- Gurib-Fakim, A. Plantago lanceolata L. Record from Protabase; Schmelzer, G.H., Gurib-Fakim, A., Eds.; PROTA (Plant Resources of Tropical Africa/Ressources Végétales de l’Afrique Tropicale): Wageningen, The Netherlands, 2006; Available online: http://www.prota4u.org (accessed on 18 May 2022).
- Szucs, I.; Escobar, M.; Grodzinski, B. Emerging Roles for Plant Terpenoids. Agriculture and Related Biotechnologies; Elsevier: Amsterdam, The Netherlands, 2011; Volume 4, pp. 273–286. [Google Scholar]
- Pankoke, H.; René Gehring, R.; Müller, C. Impact of the dual defence system of Plantago lanceolata (Plantaginaceae) on performance, nutrient utilisation, and feeding choice behavior of Amata mogadorensis larvae (Lepidoptera, Erebidae). J. Insect Physiol. 2015, 82, 99–108. [Google Scholar] [CrossRef]
- Orians, C.M.; Schweiger, R.; Dukes, J.S. Combined impacts of prolonged drought and warming on plant size and foliar chemistry. Ann. Bot. 2019, 124, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Radu, N.; Ghita, I.; Rau, I. Therapeutic Effect of Polysaccharides from Plantago Species. Mol. Cryst. Liq. Cryst. 2010, 523, 236–246. [Google Scholar]
- Radu, N.; Ghita, I.; Coman, O.; Rau, I. Therapeutic Effect of Flavonoids Derived from Plantago Species. Mol. Cryst. Liq. Cryst. 2010, 523, 273–281. [Google Scholar]
- Radu, N.; Ghita, I.; Rau, I. Therapeutic Effect of Irridoidic Compounds from Plantago Species. Mol. Cryst. Liq. Cryst. 2010, 523, 289–296. [Google Scholar]
- Sahakyan, N.Z.; Ginovyan, M.M.; Petrosyan, M.T.; Trchounian, A.H. Antibacterial and anti-phage activity of Plantago major L. Proc. Yerevian State Univ. 2019, 53, 59–64. [Google Scholar]
- Yang, J.; Lee, H.; Sung, J.; Kim, Y.; Jeong, H.S.; Lee, J. Conversion of Rutin to Quercetin by Acid Treatment in Relation to Biological Activities. Prev. Nutr. Food Sci. 2019, 24, 313–320. [Google Scholar] [CrossRef]
- Farcaș, A.D.; Zăgrean-Tuza, C.; Vlase, L.; Gheldiu, A.-M.; Pârvu, M.; Moț, A.C. EPR fingerprinting and antioxidant response of four selected Plantago species. Stud. Ubb Chem. 2020, 65, 209–220. [Google Scholar] [CrossRef]
- Yang, C.; Shi, Z.; You, L.; Du, Y.; Ni, J.; Yan, D. Neuroprotective Effect of Catalpol via Anti-Oxidative, Anti-Inflammatory, and Anti-Apoptotic Mechanisms. Front. Pharmacol. 2020, 11, 690. [Google Scholar] [CrossRef]
- Bhattamisra, S.K.; Yap, K.H.; Rao, V.; Choudhury, H. Multiple Biological Effects of an Iridoid Glucoside, Catalpol, and Its Underlying Molecular Mechanisms. Biomolecules 2020, 10, 32. [Google Scholar] [CrossRef] [Green Version]
- Mirza-Aghazadeh-Attari, M.; Ekrami, E.M.; Aghdas, S.A.M.; Mihanfar, A.; Hallaj, S.; Yousefi, B.; Safa, A.; Majidinia, M. Targeting PI3K/Akt/mTOR signaling pathway by polyphenols: Implication for cancer therapy. Life Sci. 2020, 255, 117481. [Google Scholar] [CrossRef]
- Caparica, R.; Júlio, A.; Araújo, M.E.M. Anticancer Activity of Rutin and Its Combination with Ionic Liquids on Renal Cells. Biomolecules 2020, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Vandana, J.; Gupta, A.K.; Mukerjee, A. Pharmacological Activities of Miraculous Plant Plantago Major L.: A Review. IJCPS 2017, 6, 26–37. [Google Scholar]
- Sanchez-Maldonado, A.F.; Schieber, A.; Ganzle, M.G. Antifungal activity of secondary plant metabolites from potatoes (Solanum tuberosum L.): Glycoalkaloids and phenolic acids show synergistic effects. J. Appl. Microbiol. 2016, 120, 955–965. [Google Scholar] [CrossRef]
- Martínez, G.; Regente, M.; Jacobi, S.; Del Rio, M.; Pinedo, M.; de la Canal, L. Chlorogenic acid is a fungicide active against phytopathogenic fungi. Pestic. Biochem. Physiol. 2017, 140, 30–35. [Google Scholar] [CrossRef]
- Santiago, R.; de Armas, R.; Blanch, M.; Vicente, C.; Legaz, M.E. In vitro effects of caffeic acid upon the growth of the fungi Sporisorium scitamineum. J. Plant Interact. 2010, 5, 233–240. [Google Scholar] [CrossRef]
- Bone, K.; Mills, S. (Eds.) Principles and Practice of Phytotherapy. Modern Herbal Medicine; Elsevier: Amsterdam, The Netherlands, 2013; pp. 17–82. [Google Scholar]
- De Lira Mota, K.S.; Dias, G.E.N.; Pinto, M.E.F.; Luiz-Ferreira, Â.; Monteiro Souza-Brito, A.R.; Hiruma-Lima, C.A.; Barbosa-Filho, J.M.; Batista, L.M. Flavonoids with gastroprotective activity. Molecules 2009, 14, 979–1012. [Google Scholar] [CrossRef] [Green Version]
- Pirvu, L.; Panteli, M.; Bubueanu, C.; Coprean, D. Centaurea cyanus L. Polysaccharides and Polyphenols Cooperation in Achieving Strong Rat Gastric Ulcer Protection. Cent. Eur. J. Chem. 2015, 13, 910–921. [Google Scholar] [CrossRef]
- Pintilie, G.; Paraschiv, I.; Manaila, N.; Ocnaru, D.; Armatu, A.; Colceru, S.; Pirvu, L.; Rughinis, D.; Nita, S. Cascade of bioactive compounds from Plantago lanceolata L. cultivated in Romania. Planta Med. 2007, 73, 265. [Google Scholar] [CrossRef]
- Bostan, M.; Mihaila, G.; Petrica-Matei, G.; Radu, N.; Hainarosie, R.; Stefanescu, C.-D.; Roman, V.; Diaconu, C.C. Resveratrol Modulation of Apoptosis and Cell Cycle Response to Cisplatin in Head and Neck Cancer Cell Lines. Int. J. Mol. Sci. 2021, 22, 6322. [Google Scholar] [CrossRef] [PubMed]
- Bostan, M.; Petrica-Matei, G.; Radu, N.; Hainarosie, R.; Stefansescu, C.D.; Diaconu, C.; Roman, V. The Effect of Resveratrol or Curcumin on Head and Neck Cancer Cells Sensitivity to the Cytotoxic Effects of Cisplatin. Nutrients 2020, 12, 2596. [Google Scholar] [CrossRef]
- Neagu, M.; Constantin, C.; Bostan, M.; Caruntu, C.; Ignat, S.R.; Costache, M. Proteomics technologies “lens” for epithelial-mesenchymal transition process identification in oncology. Anal. Cell. Pathol. 2019, 2019, 3565970. [Google Scholar] [CrossRef] [Green Version]
- Lukova, P.; Karcheva-Bahchevanska, D.; Dimitrova-Dyulgerova, I.; Katsarov, P.; Mladenov, R.; Iliev, I. A Comparative Pharmacognostic Study and Assessment of Antioxidant Capacity of Three Species From Plantago Genus. Farmacia 2018, 66, 609–614. [Google Scholar] [CrossRef]
- Abate, L.; Bachheti, R.K.; Tadesse, M.G.; Bachheti, A. Ethnobotanical Uses, Chemical Constituents, and Application of Plantago lanceolata L. J. Chem. 2022, 2022, 1532031. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Cantile, T.; Roberto, L.; Ingenito, A.; Catania, M.R.; Roscetto, E.; Palumbo, G.; Zarrelli, A.; Pollio, A. Determination of the in vitro and in vivo antimicrobial activity on salivary Streptococci and Lactobacilli and chemical characterization of the phenolic content of a Plantago lanceolata infusion. BioMed Res. Int. 2015, 2015, 286817. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Hu, J.; Sun, Y.; Tan, H.; Yin, J.; Geng, F.; Nie, S. Review of structure and bioactivity of Plantago (Plantaginaceae) polysaccharides. Food Chem. 2021, 12, 100158. [Google Scholar] [CrossRef]
- Lukova, P.; Nikolova, M.; Petit, E.; Elboutachfaiti, R.; Vasileva, T.; Katsarov, P.; Manev, H.; Gardarin, C.; Pierre, G.; Michaud, P.; et al. Prebiotic Activity of Poly- and Oligosaccharides Obtained from Plantago major L. Leaves. Appl. Sci. 2020, 10, 2648. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.W.; Choi, K.C. Potential Roles of Iridoid Glycosides and Their Underlying Mechanisms against Diverse Cancer Growth and Metastasis: Do They Have an Inhibitory Effect on Cancer Progression? Nutrients 2021, 13, 2974. [Google Scholar] [CrossRef]
- Janković, T.; Zdunić, G.; Beara, I.; Balog, K.; Pljevljakušić, D.; Stešević, D.; Šavikin, K. Comparative study of some polyphenols in Plantago species. Biochem. Syst. Ecol. 2012, 42, 69–74. [Google Scholar] [CrossRef]
- Kahraman, Ç.; Tatli, İ.İ.; Kart, D.; Ekizoğlu, M.; Akdemir, Z.Ş. Structure Elucidation and Antimicrobial Activities of Secondary Metabolites from the Flowery Parts of Verbascum mucronatum Lam. Turk. J. Pharm. Sci. 2018, 15, 231–237. [Google Scholar] [CrossRef]
- Ziarno, M.; Kozłowska, M.; Scibisz, I.; Kowalczyk, M.; Pawelec, S.; Stochmal, A.; Szleszynski, B. The Effect of Selected Herbal Extracts on Lactic Acid Bacteria Activity. Appl. Sci. 2021, 11, 3898. [Google Scholar] [CrossRef]
- Chiang, L.C.; Chiang, W.; Chang, M.Y.; Ng, L.T.; Lin, C.C. Antileukemic activity of selected natural products in Taiwan. Am. J. Chin. Med. 2003, 31, 37–46. [Google Scholar] [CrossRef]
- Weber, R.H.; Mondolot, L.; Kelly, M.T.; Lykke, A.M. Plantamajoside—A current review. Phytochem. Lett. 2015, 12, 42–53. [Google Scholar]
- Yang, X.; Yuan, J.; Wan, J. Cytotoxic phenolic glycosides from Boschniakia himalaica. Chem. Nat. Compd. 2012, 48, 555–558. [Google Scholar] [CrossRef]
- Saraei, R.; Marofi, F.; Naimi, A.; Talebi, M.; Ghaebi, M.; Javan, N.; Salimi, O.; Hassanzadeh, A. Leukemia therapy by flavonoids: Future and involved mechanisms. J. Cell. Physiol. 2019, 234, 8203–8220. [Google Scholar] [CrossRef]
- Cotoraci, C.; Ciceu, A.; Sasu, A.; Miutescu, E.; Hermenean, A. The Anti-Leukemic Activity of Natural Compounds. Molecules 2021, 26, 2709. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Brautigam, M.; Franz, G.G. Structural Features of Plantago lanceolata Mucilage. Planta Med. 1985, 51, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.A.; Ibrahem, M.M.; Chloob, Z.M. Identification and estimation of carbohydrate and amino acids contents of mucilage from callus cultures of Plantago lanceolata L. Plant Arch. 2020, 20, 1875–1879. [Google Scholar]
- Endalur Gopinarayanan, V.; Nair, N.U. Pentose metabolism in Saccharomyces cerevisiae: The Need to Engineer Global Regulatory Systems. Biotechnol. J. 2019, 14, e1800364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.S.; Hong, M.E.; Jung, S.C.; Ha, S.J.; Yu, B.J.; Koo, H.M.; Park, S.M.; Seo, J.-H.; Kweon, D.H.; Park, J.C.; et al. Improved galactose fermentation of Saccharomyces cerevisiae through inverse metabolic engineering. Biotechnol. Bioeng. 2011, 108, 621–631. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | 2θ (°) | θ (°) | d, Å | Peak Intensity |
|---|---|---|---|---|
| 1 | 4.8883 | 2.4442 | 18.0626 | 456.9 |
| 2 | 25.5078 | 12.7539 | 3.4892 | 51.02 |
| 3 | 27.2186 | 13.6093 | 3.2736 | 79.41 |
| 4 | 27.9222 | 13.9611 | 3.1927 | 321.18 |
| 5 | 28.3559 | 14.1780 | 3.1449 | 21501.72 |
| 6 | 40.5385 | 20.2693 | 2.2235 | 14142.32 |
| 7 | 47.7775 | 23.8888 | 1.9021 | 40.15 |
| 8 | 50.1652 | 25.0826 | 1.8170 | 162.86 |
| 9 | 58.5917 | 29.2959 | 1.5742 | 2821.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaharie, M.-G.O.; Radu, N.; Pirvu, L.; Bostan, M.; Voicescu, M.; Begea, M.; Constantin, M.; Voaides, C.; Babeanu, N.; Roman, V. Studies Regarding the Pharmaceutical Potential of Derivative Products from Plantain. Plants 2022, 11, 1827. https://doi.org/10.3390/plants11141827
Zaharie M-GO, Radu N, Pirvu L, Bostan M, Voicescu M, Begea M, Constantin M, Voaides C, Babeanu N, Roman V. Studies Regarding the Pharmaceutical Potential of Derivative Products from Plantain. Plants. 2022; 11(14):1827. https://doi.org/10.3390/plants11141827
Chicago/Turabian StyleZaharie, Marilena-Gabriela Olteanu, Nicoleta Radu, Lucia Pirvu, Marinela Bostan, Mariana Voicescu, Mihaela Begea, Mariana Constantin, Catalina Voaides, Narcisa Babeanu, and Viviana Roman. 2022. "Studies Regarding the Pharmaceutical Potential of Derivative Products from Plantain" Plants 11, no. 14: 1827. https://doi.org/10.3390/plants11141827
APA StyleZaharie, M.-G. O., Radu, N., Pirvu, L., Bostan, M., Voicescu, M., Begea, M., Constantin, M., Voaides, C., Babeanu, N., & Roman, V. (2022). Studies Regarding the Pharmaceutical Potential of Derivative Products from Plantain. Plants, 11(14), 1827. https://doi.org/10.3390/plants11141827