
Expression Analysis of Phenylpropanoid Pathway Genes and Metabolomic Analysis of Phenylpropanoid Compounds in Adventitious, Hairy, and Seedling Roots of Tartary Buckwheat

 ,
, 
 and
and
Abstract
:1. Introduction
2. Results and Discussion
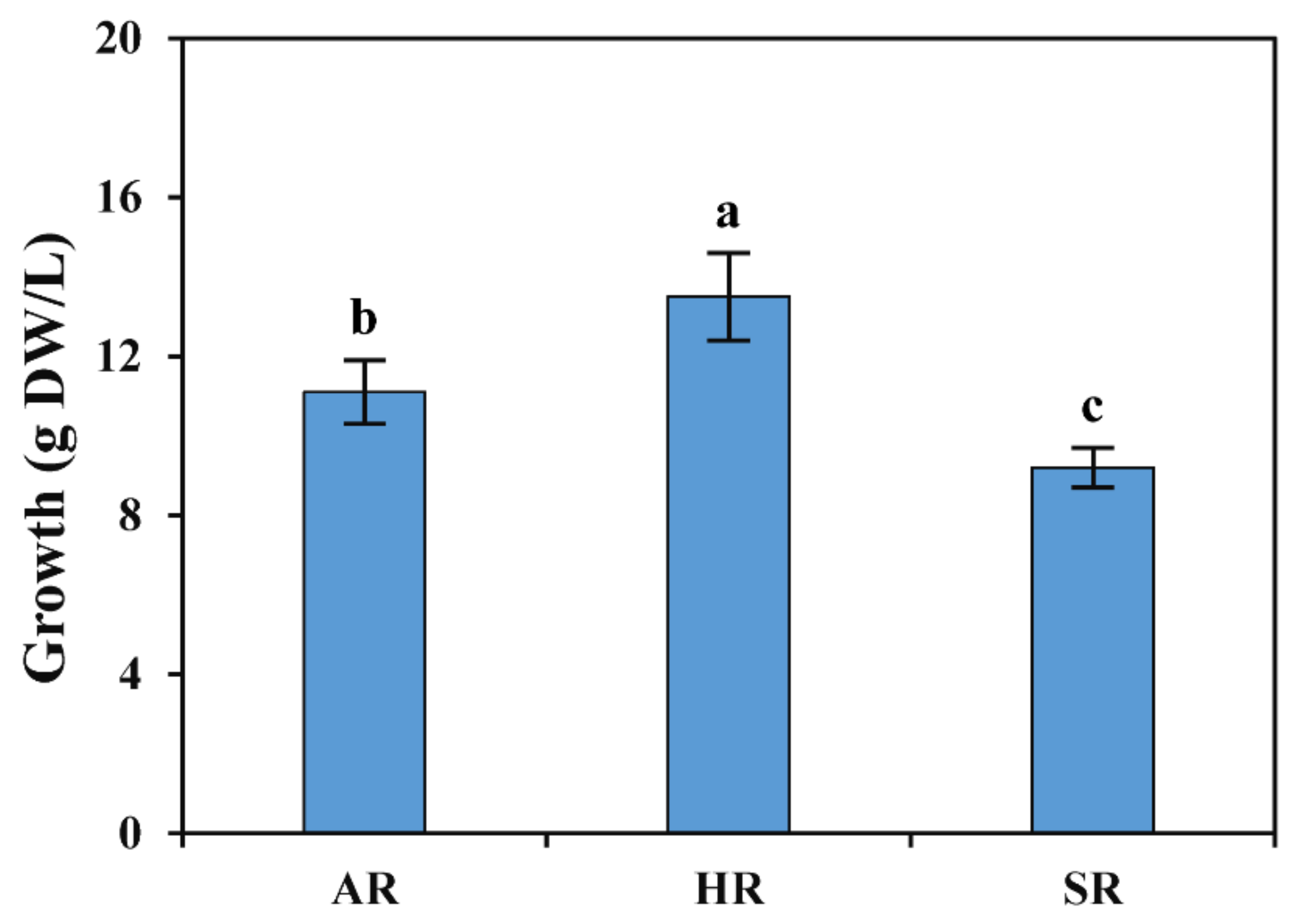
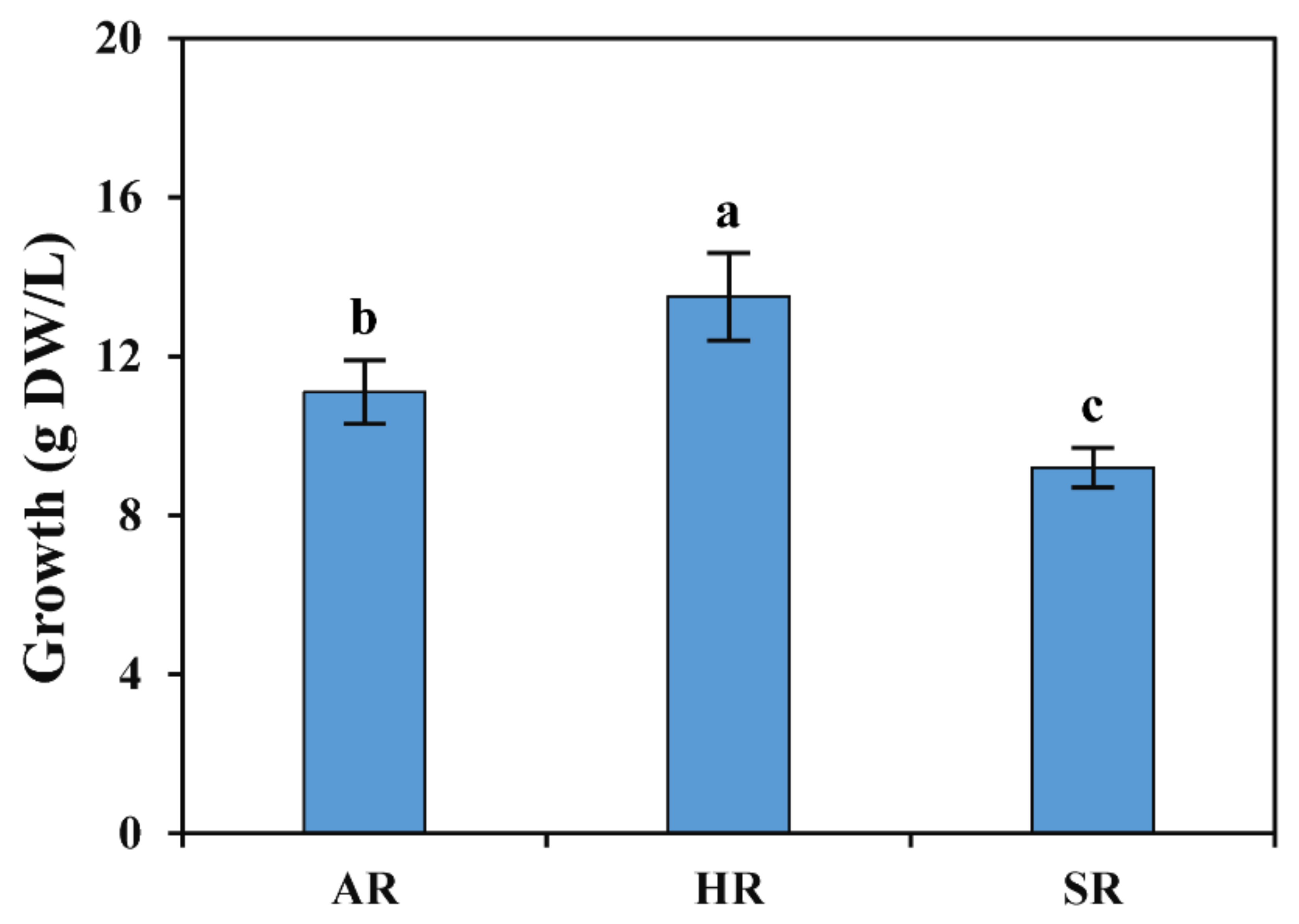
2.1. Optimization of Culture Medium for ARs Induction
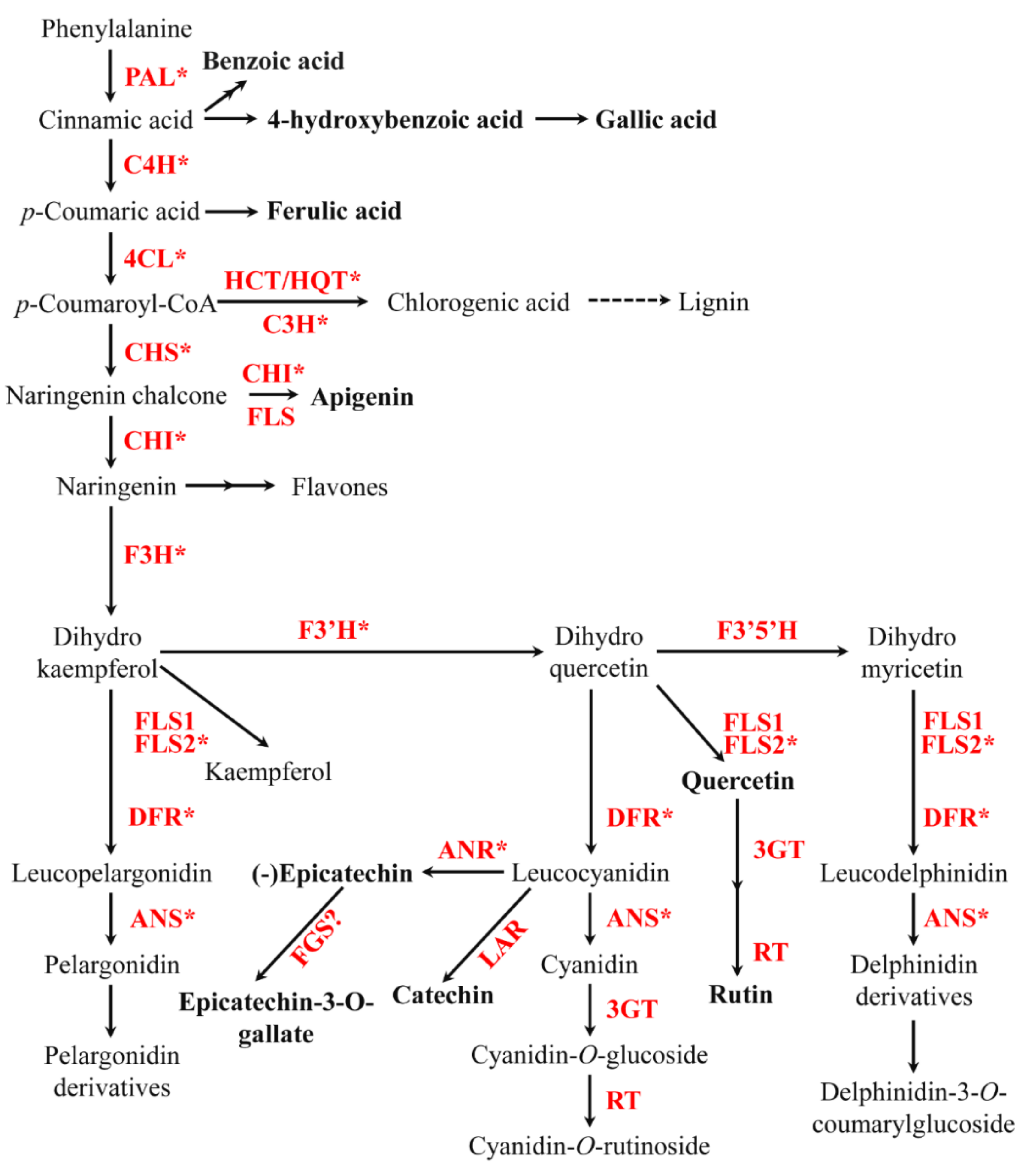
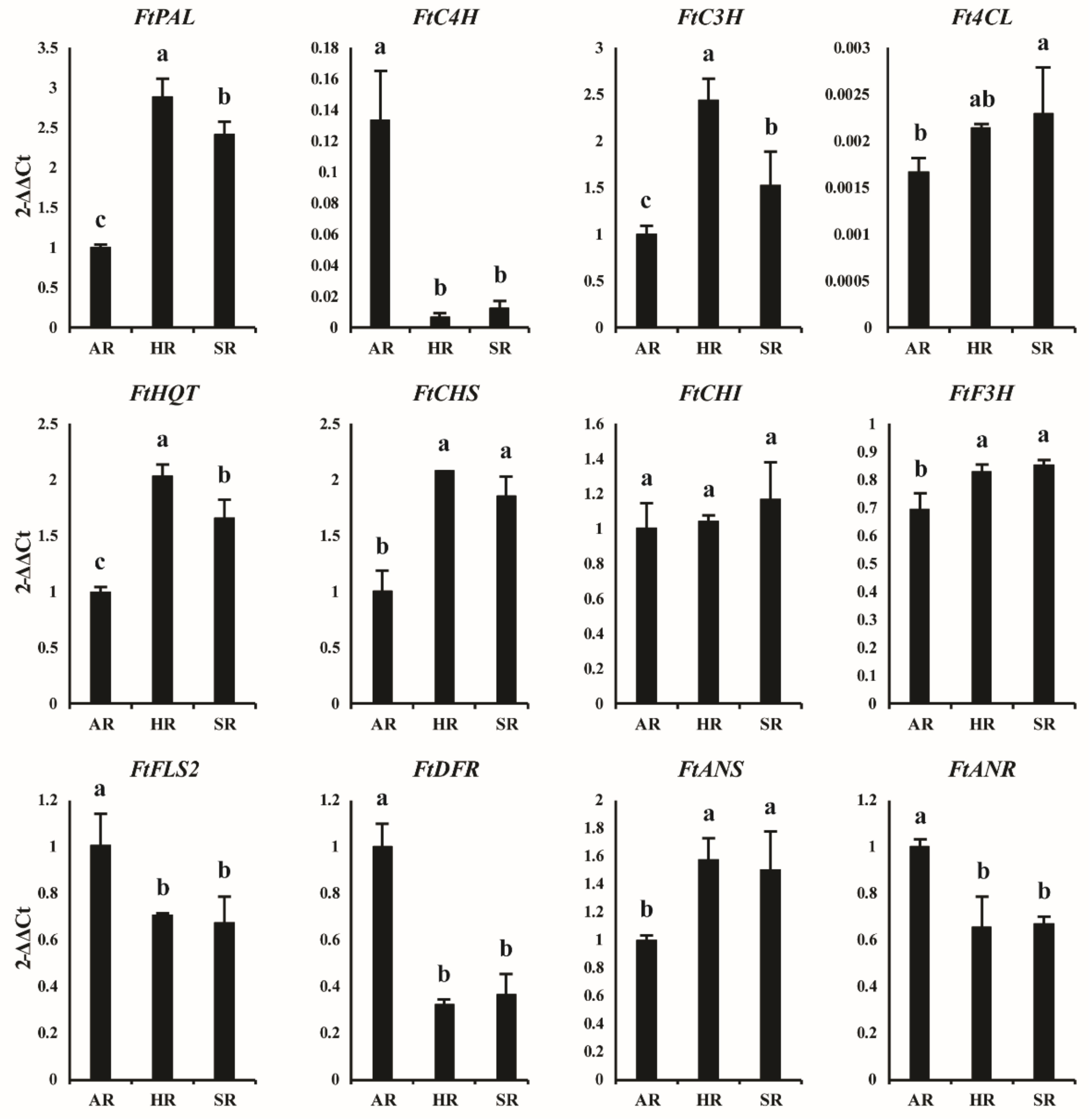
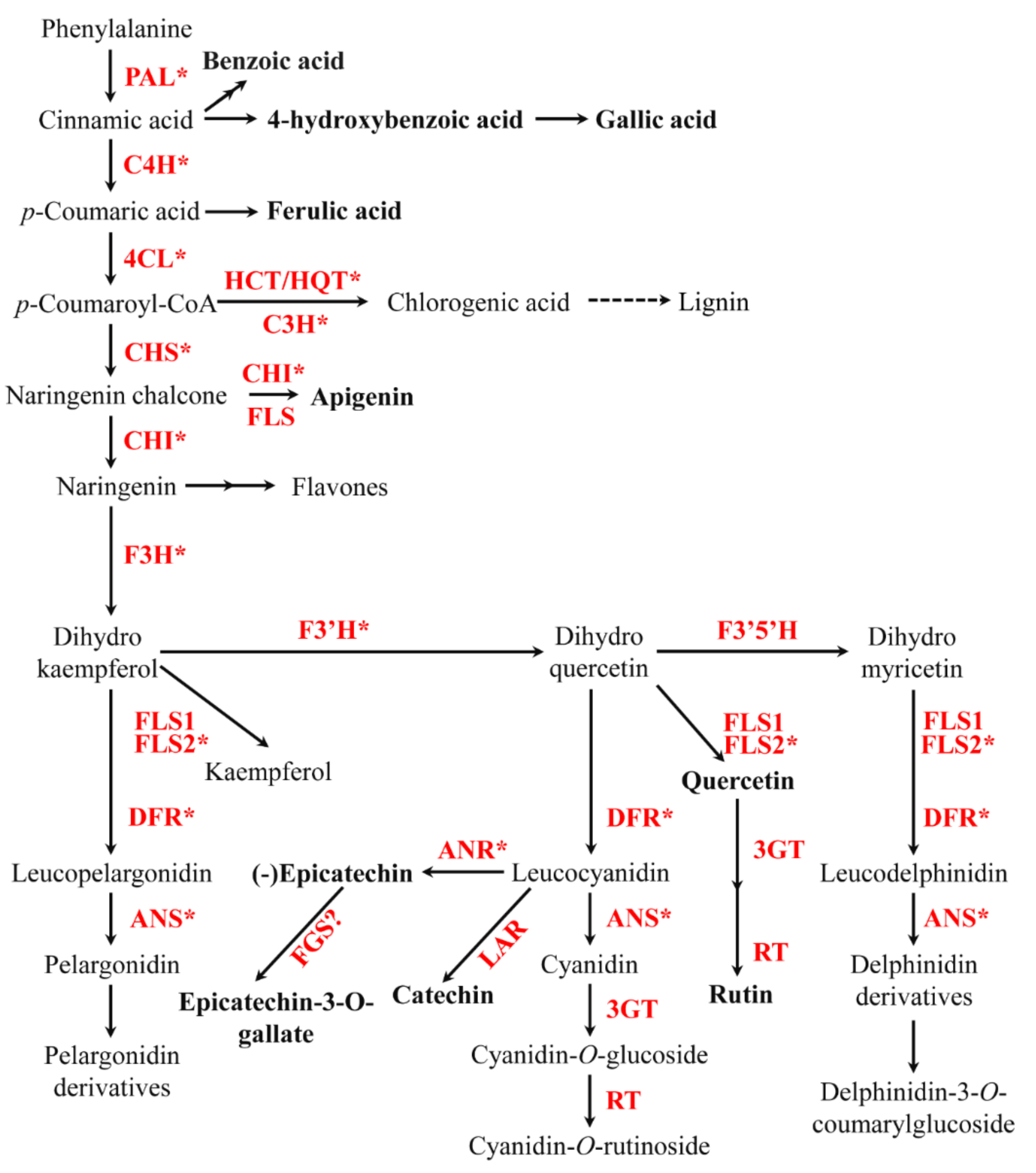
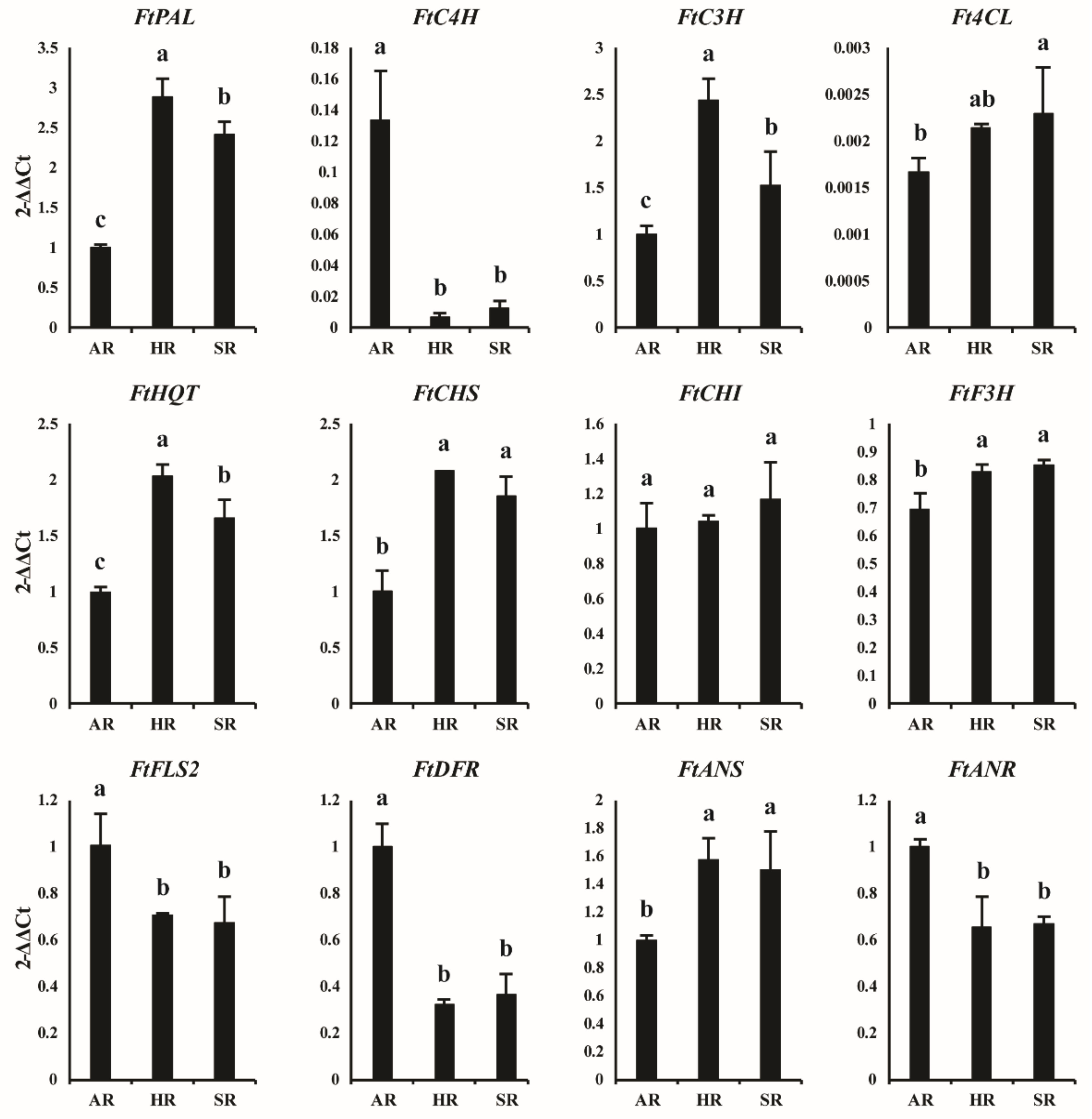
2.2. Phenolic Biosynthetic Pathway Gene and Its Expression Levels in Different Types of Roots
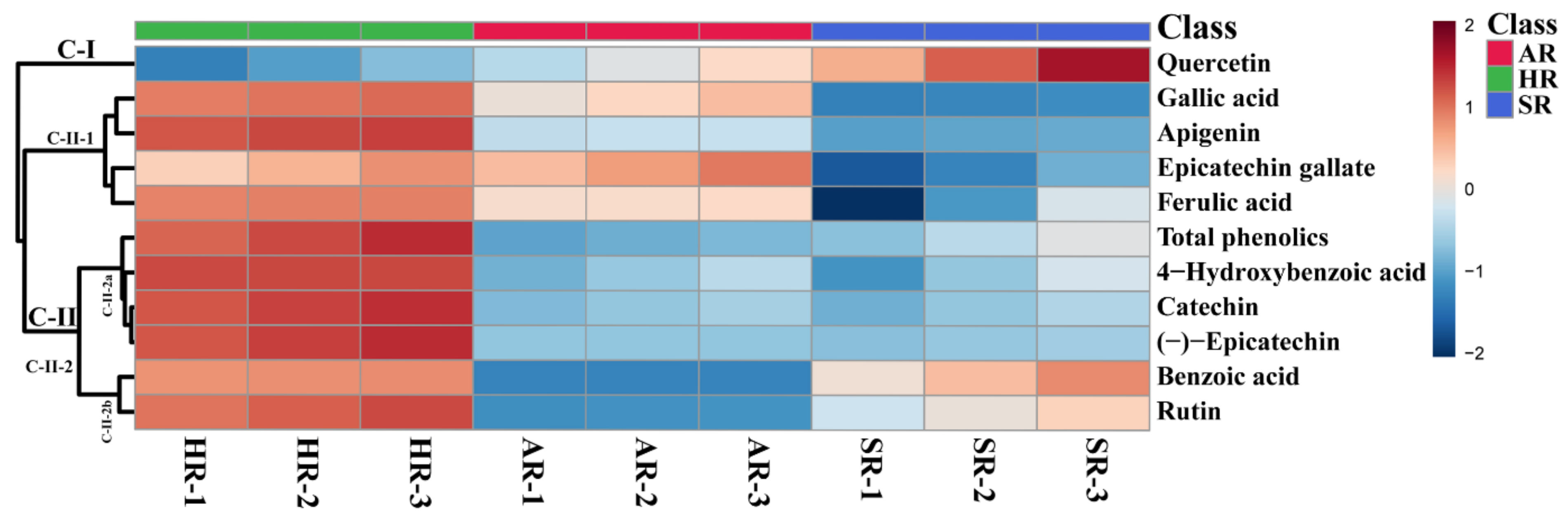
2.3. Phenolic Compound Content in Three Types of Root
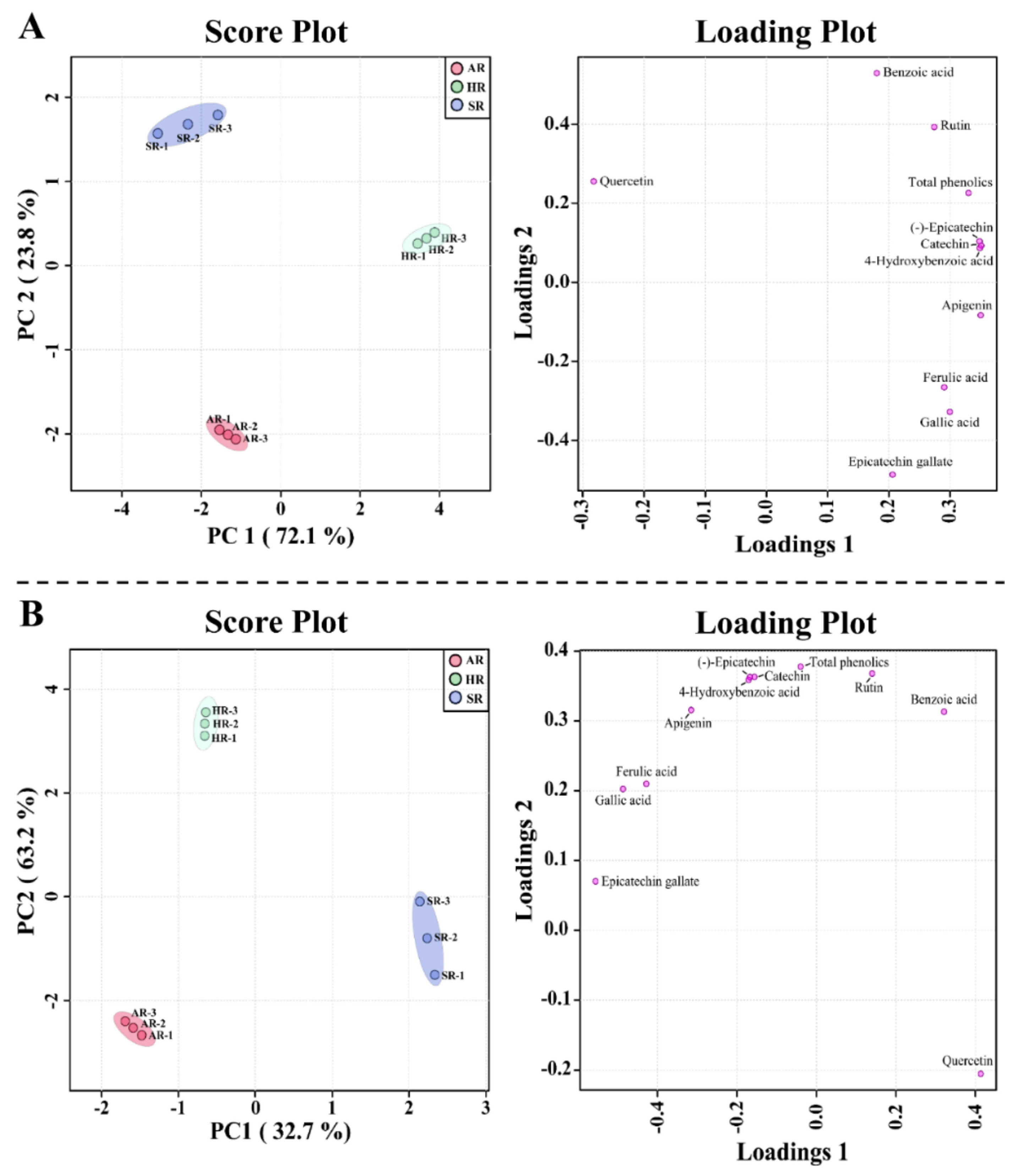
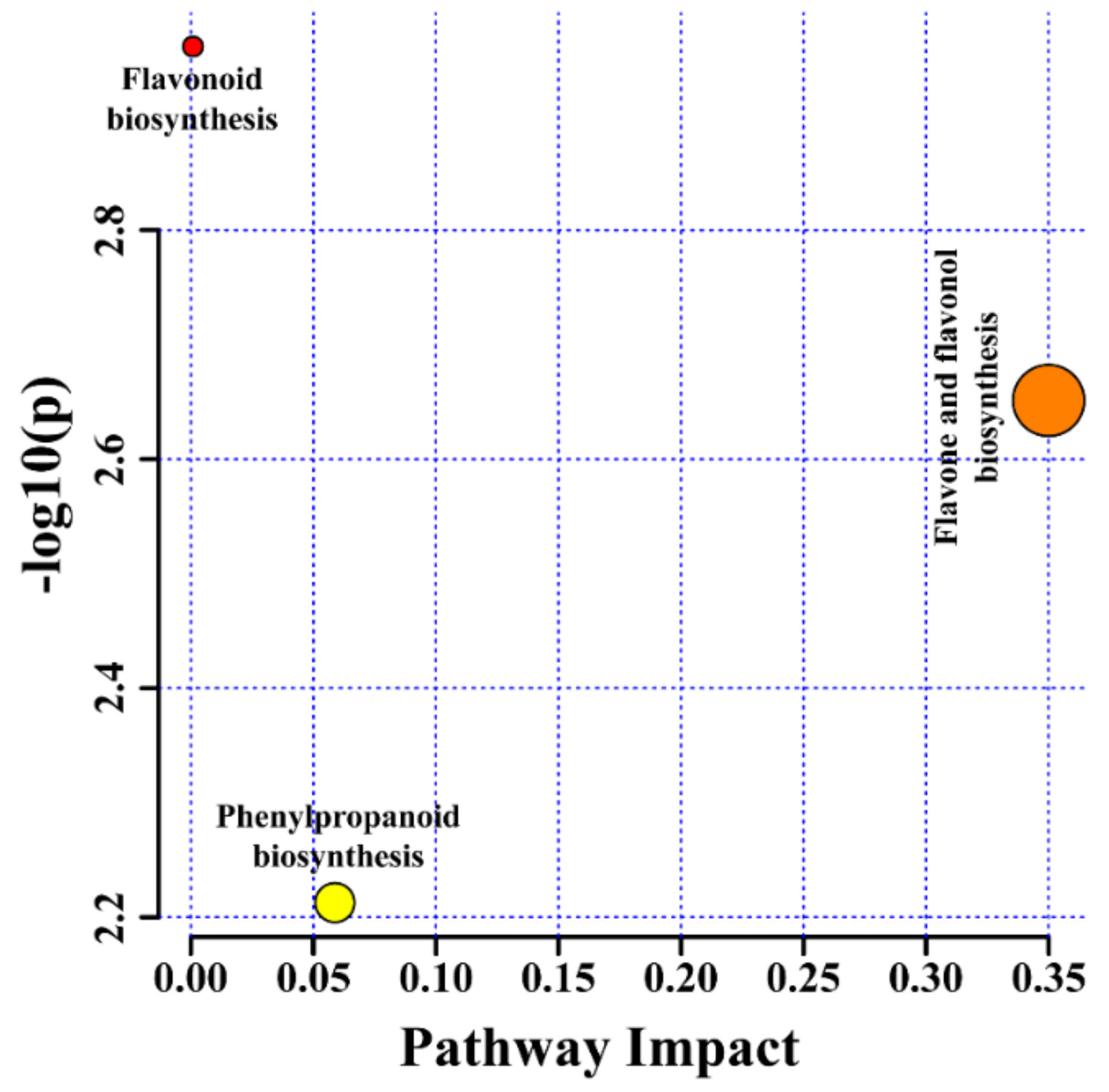
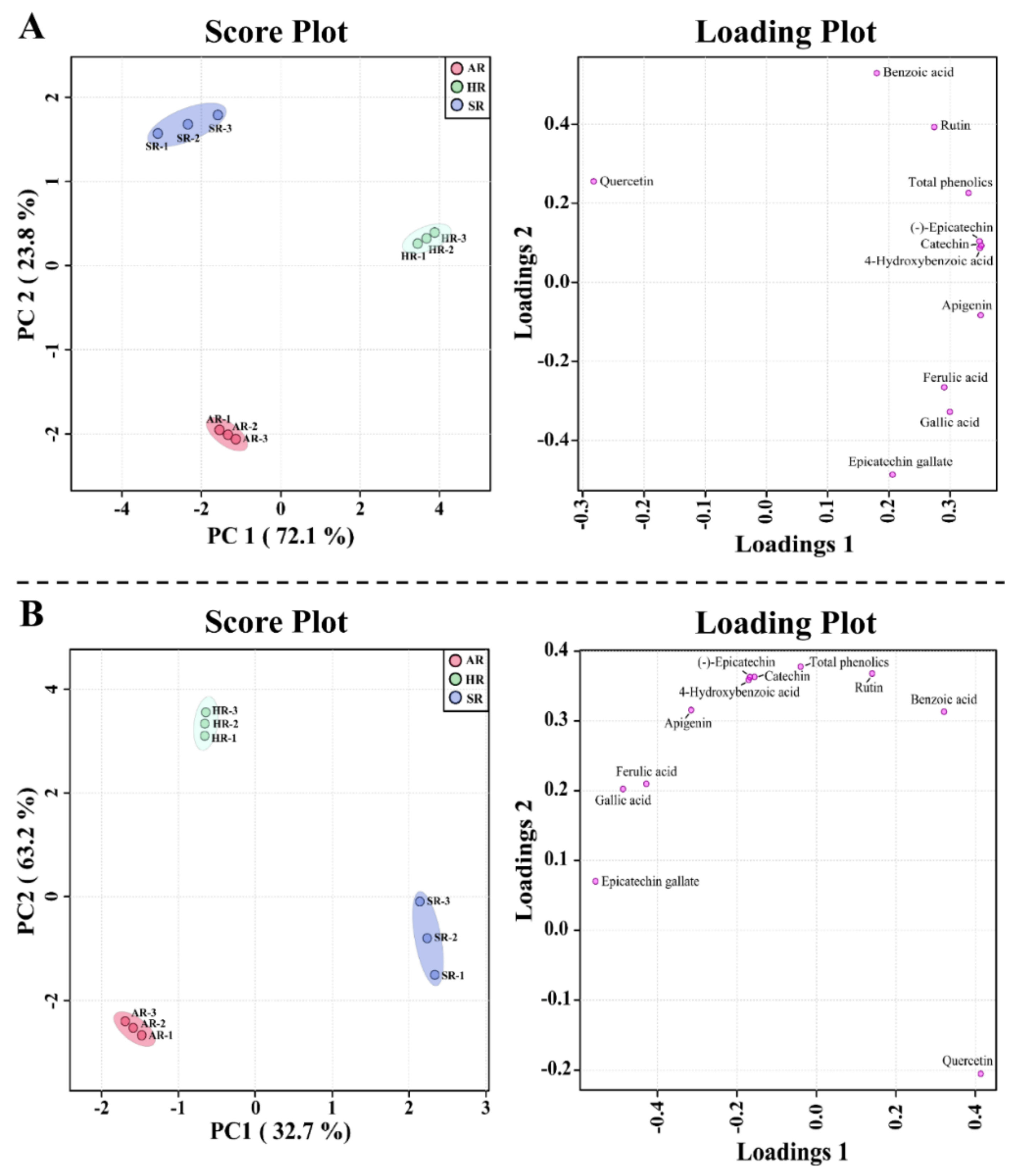
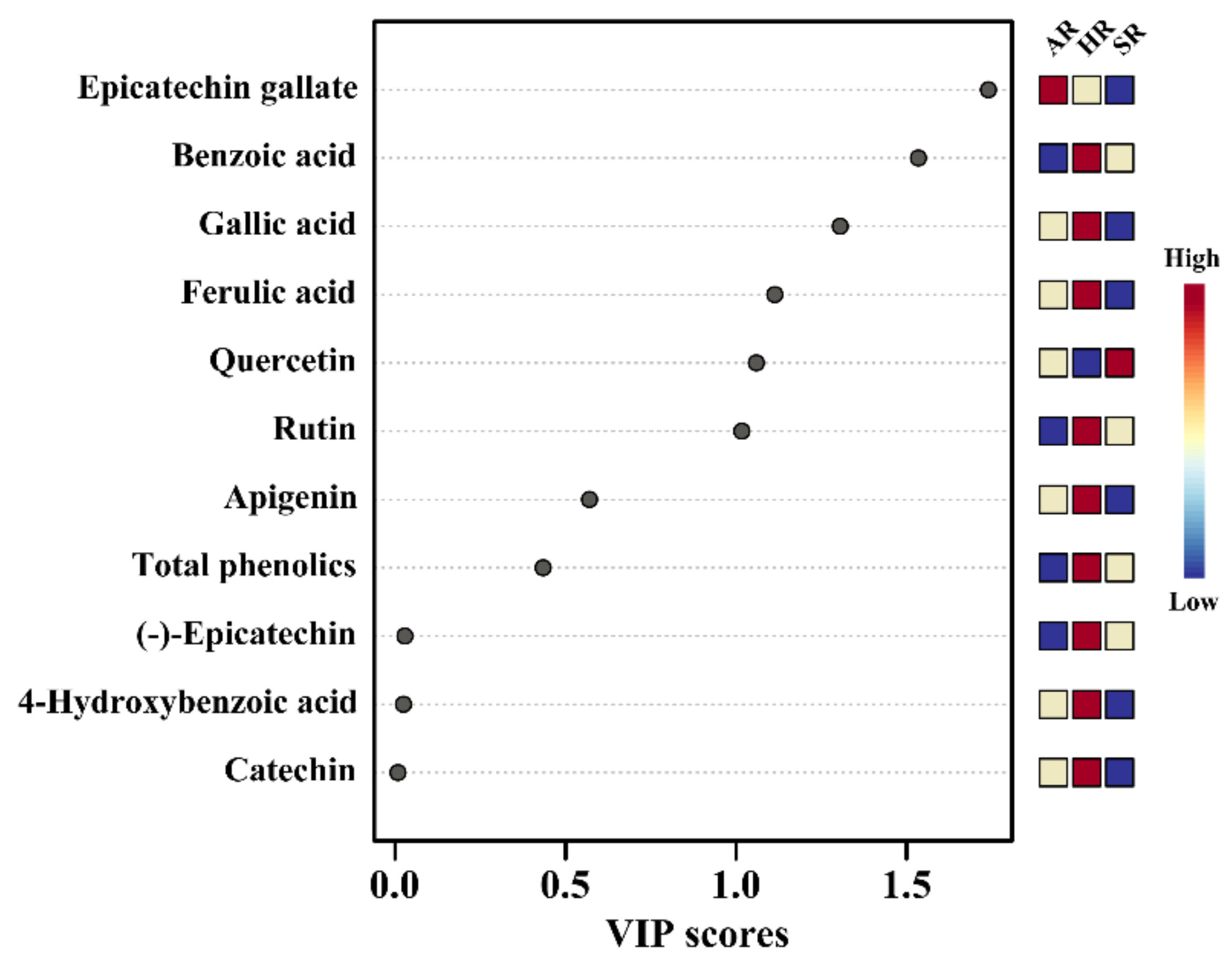
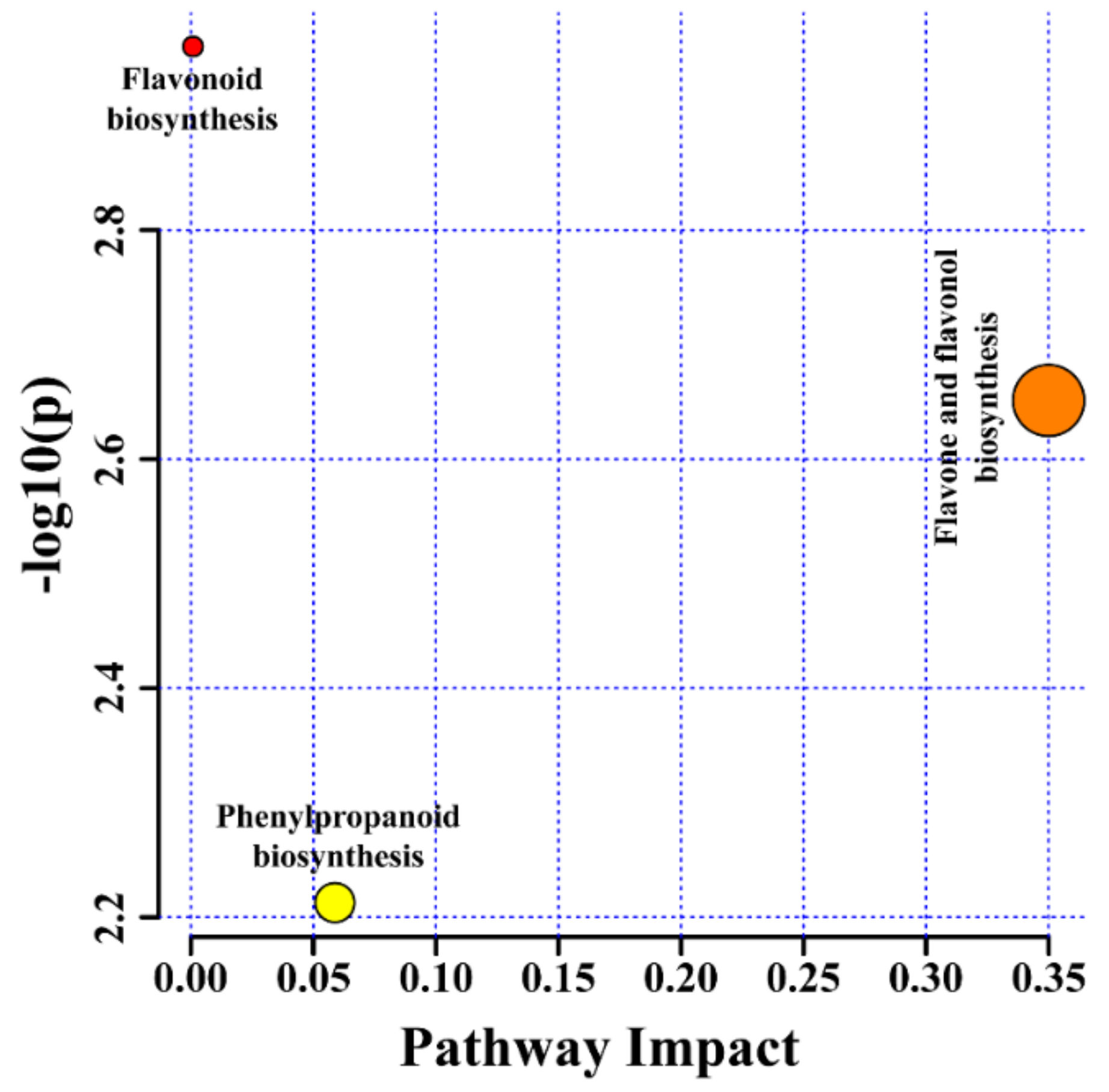
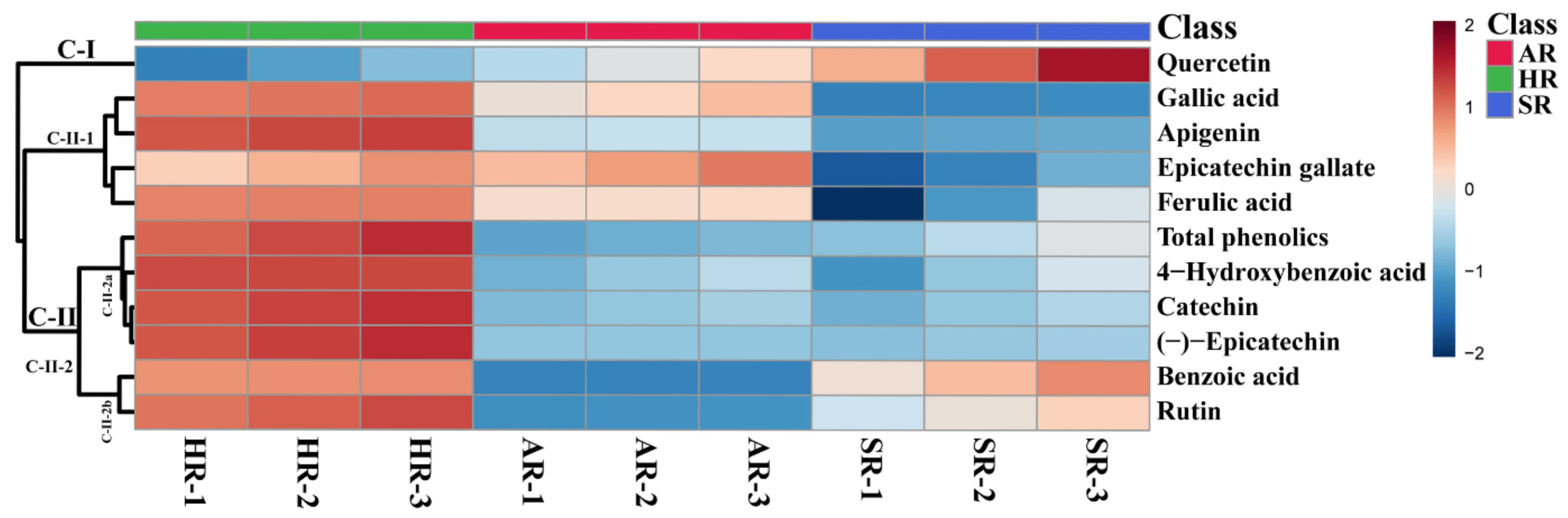
2.4. Metabolic Profiling of Different Types of Root Cultures
3. Materials and Methods
3.1. Three Types of Root Induction
3.2. qRT-PCR Analysis
3.3. HPLC Analysis
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fabjan, N.; Rode, J.; Košir, I.J.; Wang, Z.; Zhang, Z.; Kreft, I. Tartary buckwheat (Fagopyrum tataricum Gaertn.) as a source of dietary rutin and quercitrin. J. Agric. Food Chem. 2003, 51, 6452–6455. [Google Scholar] [CrossRef]
- Park, N.I.; Li, X.; Suzuki, T.; Kim, S.-J.; Woo, S.-H.; Park, C.H.; Park, S.U. Differential expression of anthocyanin biosynthetic genes and anthocyanin accumulation in tartary buckwheat cultivars ‘Hokkai T8’and ‘Hokkai T10’. J. Agric. Food Chem. 2011, 59, 2356–2361. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. Chemical composition and health effects of Tartary buckwheat. Food Chem. 2016, 203, 231–245. [Google Scholar] [CrossRef]
- Guillon, S.; Trémouillaux-Guiller, J.; Pati, P.K.; Gantet, P. Hairy roots: A powerful tool for plant biotechnological advances. In Bioactive Molecules and Medicinal Plants; Ramawat, K.G., Merillon, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 271–283. [Google Scholar]
- Bhatia, S.; Bera, T.; Dahiya, R.; Bera, T.; Bhatia, S.; Bera, T. Classical and nonclassical techniques for secondary metabolite production in plant cell culture. In Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Bhatia, S., Sharma, K., Dahiya, R., Bera, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 231–291. [Google Scholar]
- Su, W.W.; Lee, K.-T. Plant cell and hairy root cultures–Process characteristics, products, and applications. In Bioprocessing for Value-Added Products from Renewable Resources; Su, W.W., Lee, K.-T., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 263–292. [Google Scholar]
- Steffens, B.; Rasmussen, A. The physiology of adventitious roots. Plant Physiol. 2016, 170, 603–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonin, M.; Bergougnoux, V.; Nguyen, T.D.; Gantet, P.; Champion, A. What makes adventitious roots? Plants 2019, 8, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmat, E.; Kang, Y. Adventitious root culture for secondary metabolite production in medicinal plants: A review. J. Plant Biotechnol. 2019, 46, 143–157. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Chan, C.-L.; Yang, Q.-Q.; Li, H.-B.; Zhang, D.; Ge, Y.-Y.; Gunaratne, A.; Ge, J.; Corke, H. Bioactive compounds and beneficial functions of sprouted grains. In Sprouted Grains; Feng, H., Nemzer, B., DeVries, J.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 191–246. [Google Scholar]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.; Visser, R.G.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sathasivam, R.; Park, N.I.; Wu, Q.; Park, S.U. Enhancement of phenylpropanoid accumulation in tartary buckwheat hairy roots by overexpression of MYB transcription factors. Ind. Crop. Prod. 2020, 156, 112887. [Google Scholar] [CrossRef]
- Murthy, H.N.; Hahn, E.J.; Paek, K.Y. Adventitious roots and secondary metabolism. Chin. J. Biotechnol. 2008, 24, 711–716. [Google Scholar] [CrossRef]
- Sivanesan, I.; Jeong, B.R. Induction and establishment of adventitious and hairy root cultures of Plumbago zeylanica L. Afr. J. Biotechnol. 2009, 8, 5294–5300. [Google Scholar]
- Baskaran, P.; Jayabalan, N. Psoralen production in hairy roots and adventitious roots cultures of Psoralea coryfolia. Biotechnol. Lett. 2009, 31, 1073–1077. [Google Scholar] [CrossRef]
- Choffe, K.L.; Murch, S.J.; Saxena, P.K. Regeneration of Echinacea purpurea: Induction of root organogenesis from hypocotyl and cotyledon explants. Plant Cell Tissue Organ Cult. 2000, 62, 227–234. [Google Scholar] [CrossRef]
- Jenifer, U.; Francina Cecilia, K.; Ravindhran, R. In vitro adventitious root and hairy root cultures in Boerhaavia diffusa L. Int. J. Curr. Res. 2012, 4, 65–67. [Google Scholar]
- Swamy, S.; Puri, S.; Singh, A. Effect of auxins (IBA and NAA) and season on rooting of juvenile and mature hardwood cuttings of Robinia pseudoacacia and Grewia optiva. New For. 2002, 23, 143–157. [Google Scholar] [CrossRef]
- Sevón, N.; Oksman-Caldentey, K.-M. Agrobacterium rhizogenes-mediated transformation: Root cultures as a source of alkaloids. Planta Med. 2002, 68, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Shanks, J.V.; Morgan, J. Plant ‘hairy root’culture. Curr. Opin. Biotechnol. 1999, 10, 151–155. [Google Scholar] [CrossRef]
- Park, Y.J.; Thwe, A.A.; Li, X.; Kim, Y.J.; Kim, J.K.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Triterpene and flavonoid biosynthesis and metabolic profiling of hairy roots, adventitious roots, and seedling roots of Astragalus membranaceus. J. Agric. Food Chem. 2015, 63, 8862–8869. [Google Scholar] [CrossRef]
- Kim, Y.; Wyslouzil, B.E.; Weathers, P.J. Secondary metabolism of hairy root cultures in bioreactors. Vitr. Cell. Dev. Biol. Plant 2002, 38, 1–10. [Google Scholar] [CrossRef]
- Gómez-Martínez, H.; Gil-Muñoz, F.; Bermejo, A.; Zuriaga, E.; Badenes, M.L. Insights of phenolic pathway in fruits: Transcriptional and metabolic profiling in Apricot (Prunus armeniaca). Int. J. Mol. Sci. 2021, 22, 3411. [Google Scholar] [CrossRef]
- Wang, K.; Jin, P.; Han, L.; Shang, H.; Tang, S.; Rui, H.; Duan, Y.; Kong, F.; Kai, X.; Zheng, Y. Methyl jasmonate induces resistance against Penicillium citrinum in Chinese bayberry by priming of defense responses. Postharvest Biol. Technol. 2014, 98, 90–97. [Google Scholar] [CrossRef]
- Rashid, A.; Ali, V.; Khajuria, M.; Faiz, S.; Gairola, S.; Vyas, D. GC-MS based metabolomic approach to understand nutraceutical potential of Cannabis seeds from two different environments. Food Chem. 2021, 339, 128076. [Google Scholar] [CrossRef] [PubMed]
- De Falco, B.; Fiore, A.; Rossi, R.; Amato, M.; Lanzotti, V. Metabolomics driven analysis by UAEGC-MS and antioxidant activity of chia (Salvia hispanica L.) commercial and mutant seeds. Food Chem. 2018, 254, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Raval, S.S.; Mahatma, M.K.; Chakraborty, K.; Bishi, S.K.; Singh, A.L.; Rathod, K.J.; Jadav, J.K.; Sanghani, J.M.; Mandavia, M.K.; Gajera, H.P. Metabolomics of groundnut (Arachis hypogaea L.) genotypes under varying temperature regimes. Plant Growth Regul. 2018, 84, 493–505. [Google Scholar] [CrossRef]
- Yan, S.; Huang, W.; Gao, J.; Fu, H.; Liu, J. Comparative metabolomic analysis of seed metabolites associated with seed storability in rice (Oryza sativa L.) during natural aging. Plant Physiol. Biochem. 2018, 127, 590–598. [Google Scholar] [CrossRef]
- Jeon, J.; Kim, J.K.; Wu, Q.; Park, S.U. Comparative transcriptomic and metabolomic analyses in response to cold in Tartary buckwheat (Fagopyrum tataricum). bioRxiv 2018, 278432. [Google Scholar]
- Park, C.H.; Park, Y.E.; Yeo, H.J.; Park, N.I.; Park, S.U. Effect of light and dark on the phenolic compound accumulation in Tartary buckwheat hairy roots overexpressing ZmLC. Int. J. Mol. Sci. 2021, 22, 4702. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Auxin | Root Induction (%) | Number of Roots | Length of Roots (cm) |
|---|---|---|---|
| 1/2 SH | 66.67 | 5.25 ± 1.92 b | 2.1 ± 0.66 ab |
| NAA 0.1 | 61.11 | 4.25 ± 1.09 b | 1.9 ± 0.58 ab |
| NAA 0.5 | 75 | 2.75 ± 1.09 b | 1.9 ± 0.36 ab |
| NAA 1 | 58.33 | 3 ± 1.22 b | 2 ± 0.38 ab |
| IBA 0.1 | 66.67 | 4.5 ± 1.12 b | 2.7 ± 1.01 a |
| IBA 0.5 | 66.67 | 4.75 ± 3.34 b | 1.66 ± 0.58 ab |
| IBA 1 | 83.33 | 5.25 ± 1.48 b | 1.36 ± 0.39 b |
| IAA 0.1 | 58.33 | 10 ± 2.74 a | 2.14 ± 0.81 ab |
| IAA 0.5 | 91.67 | 10.25 ± 0.83 a | 1.96 ± 0.68 ab |
| IAA 1 | 83.33 | 12.25 ± 1.48 a | 1.76 ± 0.48 ab |
| Compounds | Adventitious Root (AR) | Hairy Root (HR) | Seedling Root (SR) |
|---|---|---|---|
| Gallic acid | 0.699 ± 0.068 b | 0.933 ± 0.019 a | 0.227 ± 0.021 c |
| Catechin | 1.616 ± 0.024 b | 2.029 ± 0.026 a | 1.614 ± 0.043 b |
| Benzoic acid | 0.169 ± 0.018 a | ND | 0.196 ± 0.001 a |
| 4-hydroxybenzoic acid | 0.193 ± 0.011 b | 0.286 ± 0.000 a | 0.192 ± 0.023 b |
| (-)-Epicatechin | 3.922 ± 0.003 b | 5.159 ± 0.087 a | 3.943 ± 0.046 b |
| Epicatechin gallate | 5.972 ± 0.109 a | 5.896 ± 0.111 a | 5.059 ± 0.187 b |
| Ferulic acid | 0.042 ± 0.000 a | 0.049 ± 0.000 a | 0.031 ± 0.009 b |
| Rutin | 16.133 ± 0.053 c | 21.288 ± 0.314 a | 18.780 ± 0.608 b |
| Quercetin | 2.283 ± 0.053 b | 2.123 ± 0.048 c | 2.491 ± 0.091 a |
| Apigenin | 0.003 ± 0.000 b | 0.007 ± 0.000 a | 0.002 ± 0.000 c |
| Total | 31.032 ± 0.339 c | 37.770 ± 0.605 a | 32.535 ± 1.029 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.; Sathasivam, R.; Nguyen, B.V.; Park, N.I.; Woo, S.-H.; Park, S.U. Expression Analysis of Phenylpropanoid Pathway Genes and Metabolomic Analysis of Phenylpropanoid Compounds in Adventitious, Hairy, and Seedling Roots of Tartary Buckwheat. Plants 2022, 11, 90. https://doi.org/10.3390/plants11010090
Choi M, Sathasivam R, Nguyen BV, Park NI, Woo S-H, Park SU. Expression Analysis of Phenylpropanoid Pathway Genes and Metabolomic Analysis of Phenylpropanoid Compounds in Adventitious, Hairy, and Seedling Roots of Tartary Buckwheat. Plants. 2022; 11(1):90. https://doi.org/10.3390/plants11010090
Chicago/Turabian StyleChoi, Minsol, Ramaraj Sathasivam, Bao Van Nguyen, Nam Il Park, Sun-Hee Woo, and Sang Un Park. 2022. "Expression Analysis of Phenylpropanoid Pathway Genes and Metabolomic Analysis of Phenylpropanoid Compounds in Adventitious, Hairy, and Seedling Roots of Tartary Buckwheat" Plants 11, no. 1: 90. https://doi.org/10.3390/plants11010090
APA StyleChoi, M., Sathasivam, R., Nguyen, B. V., Park, N. I., Woo, S.-H., & Park, S. U. (2022). Expression Analysis of Phenylpropanoid Pathway Genes and Metabolomic Analysis of Phenylpropanoid Compounds in Adventitious, Hairy, and Seedling Roots of Tartary Buckwheat. Plants, 11(1), 90. https://doi.org/10.3390/plants11010090