
Magnetic Immobilization and Growth of Nannochloropsis oceanica and Scenedasmus almeriensis

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
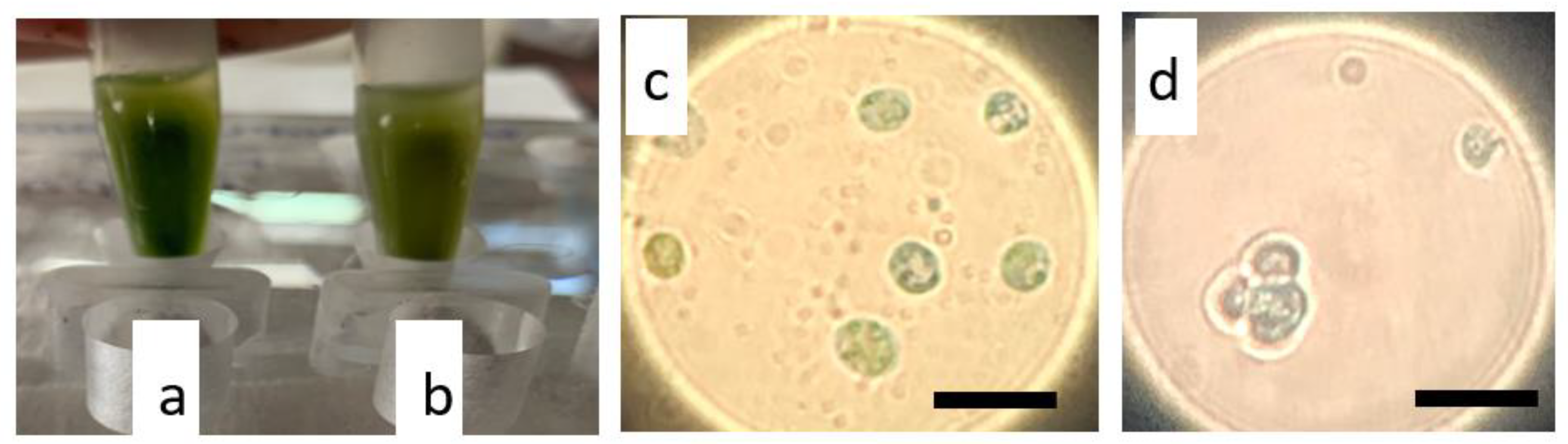
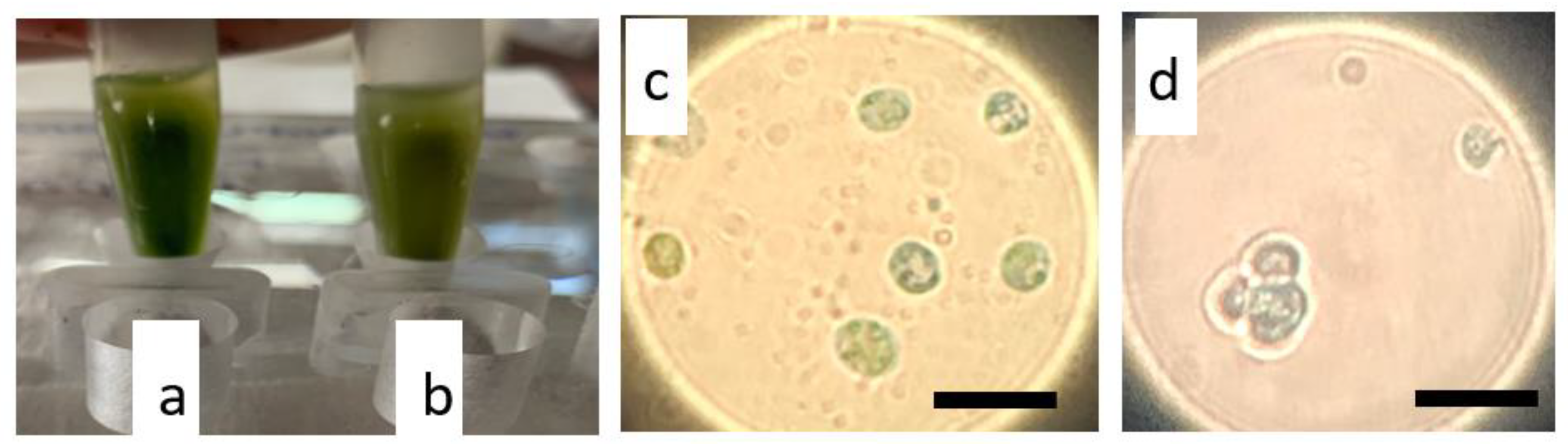
2.1. Enzymatic and Mechanical Treatment of N. oceanica and S. almeriensis Microalgae Cells Driving Protoplasts Formation
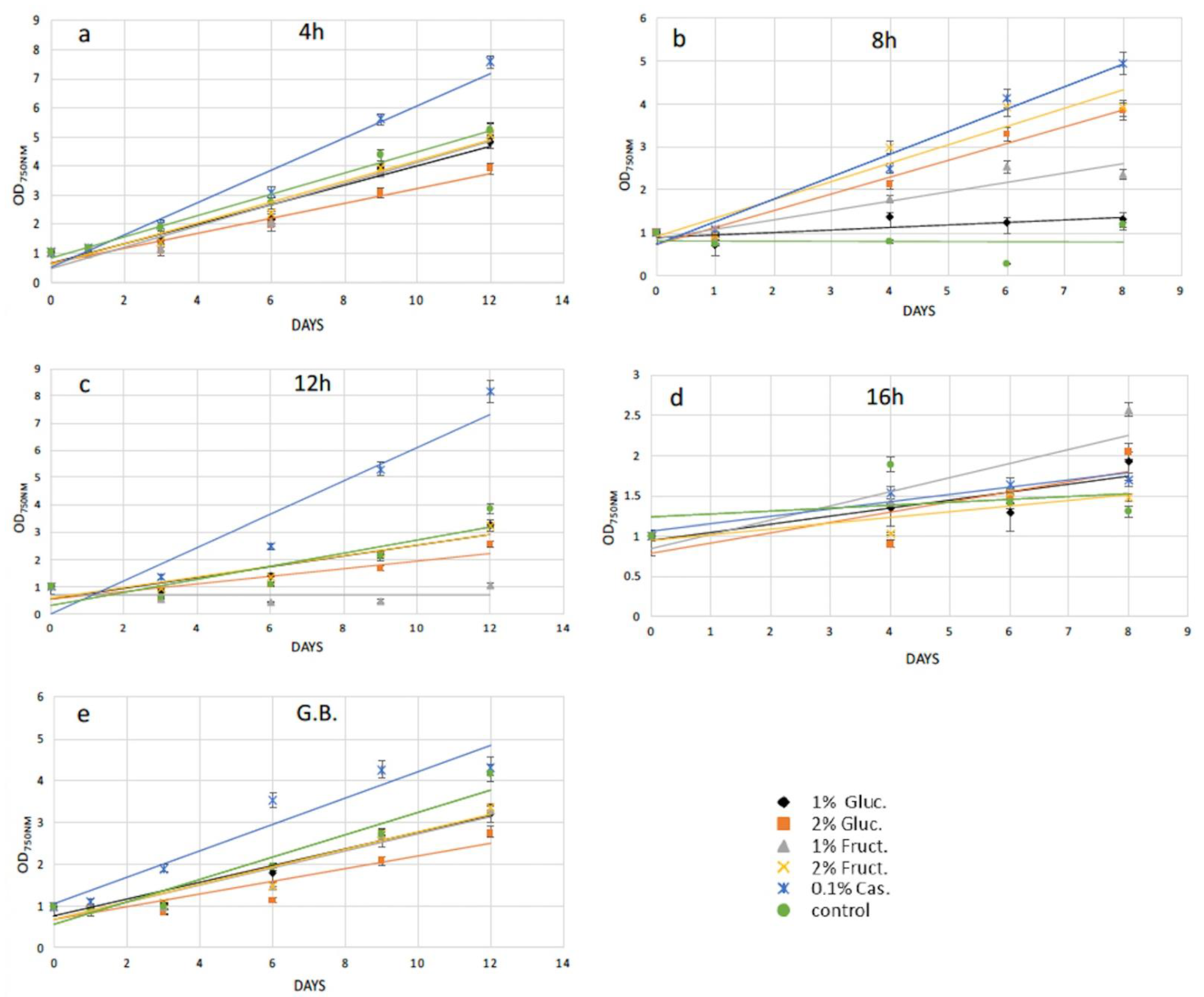
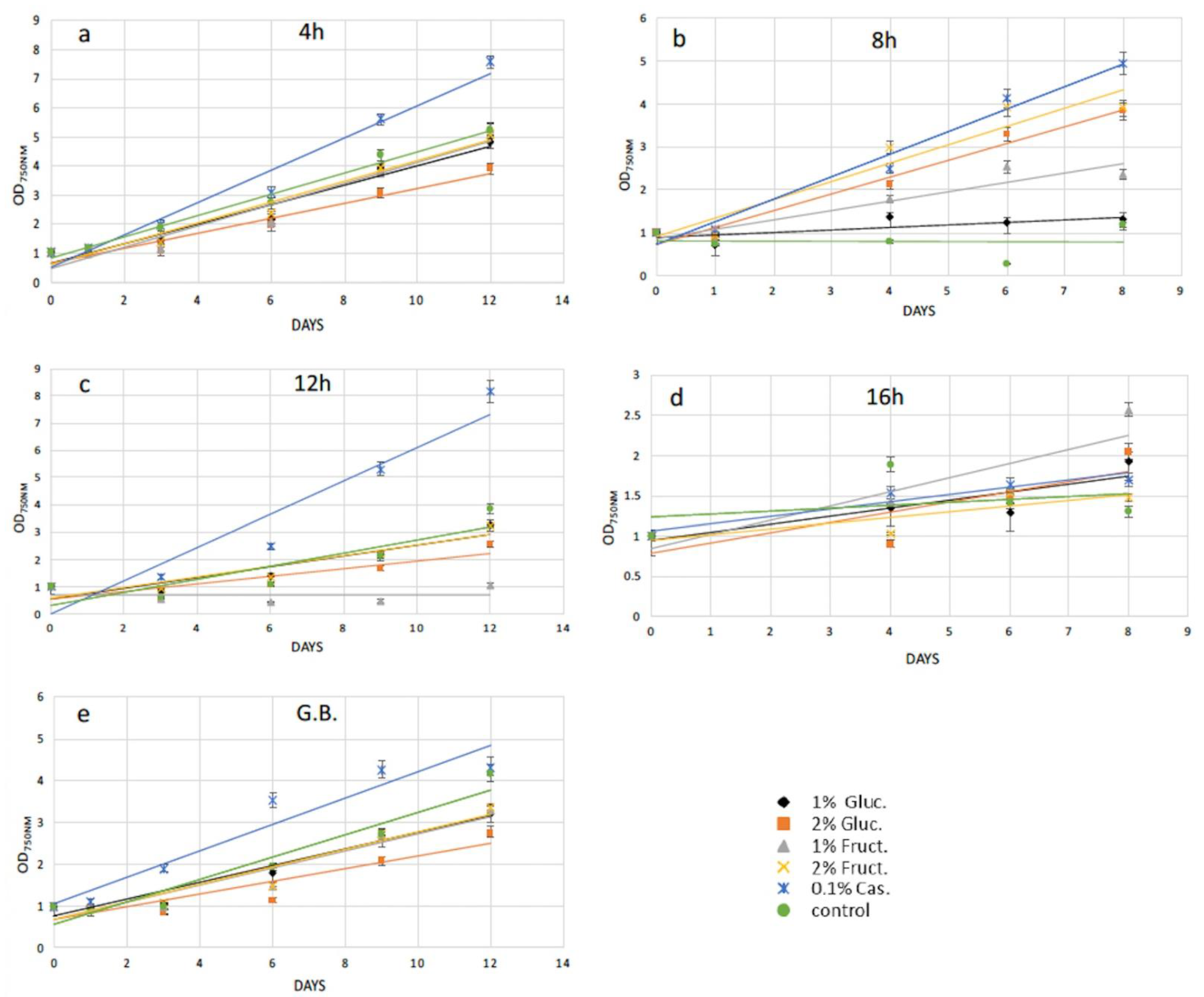
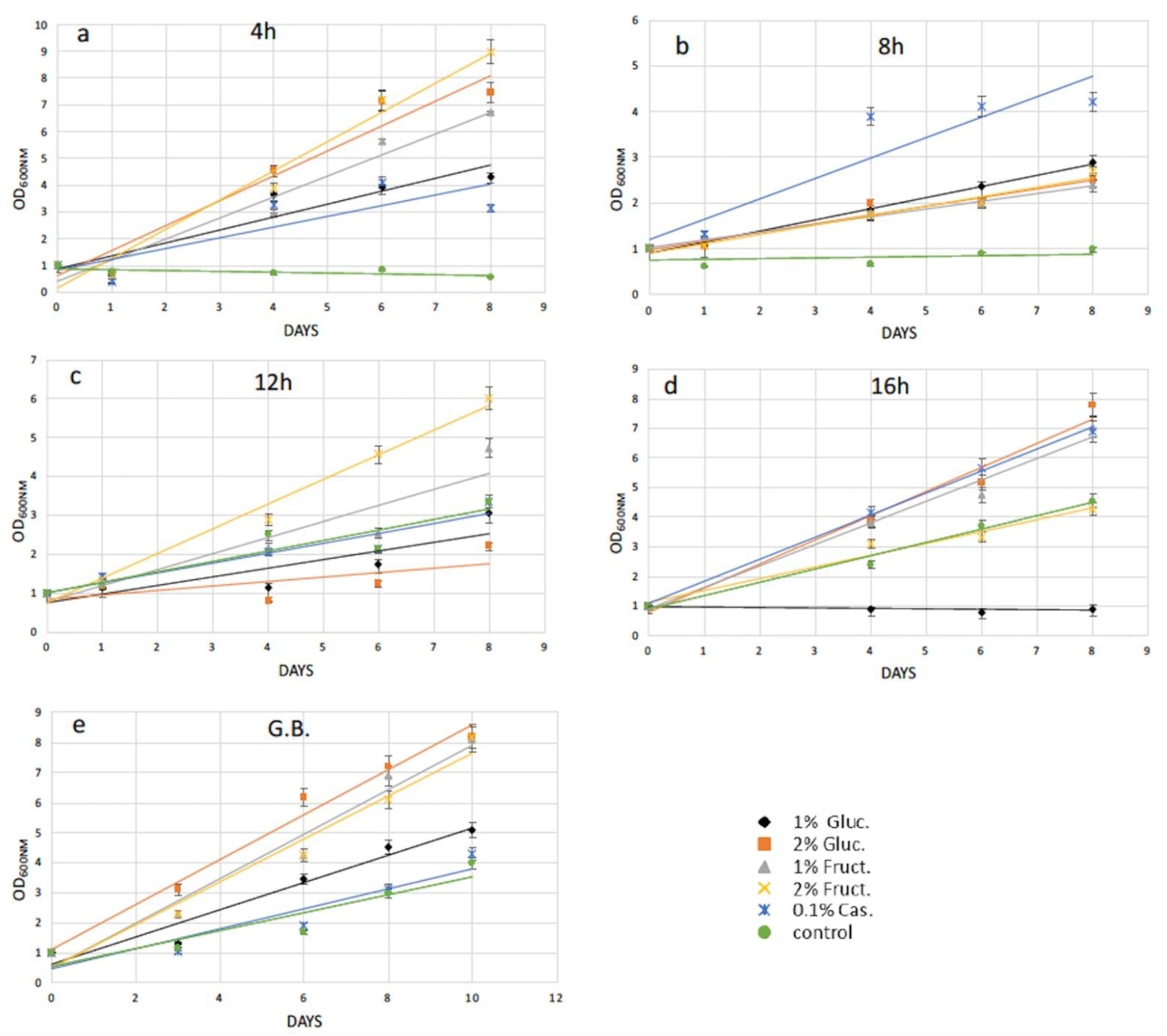
2.2. Protoplasts Regeneration Using Carbon and Amino Acids Supplements
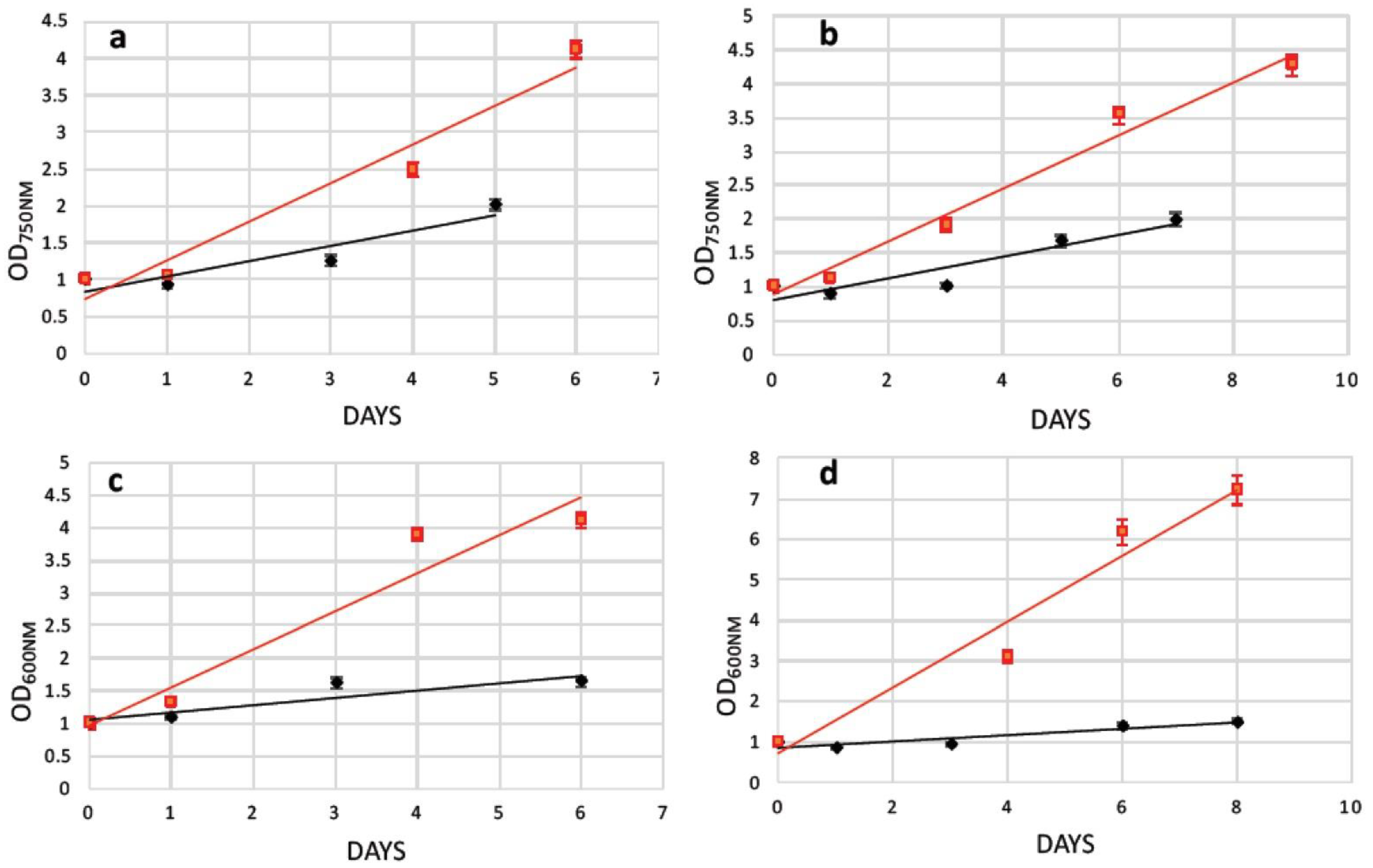
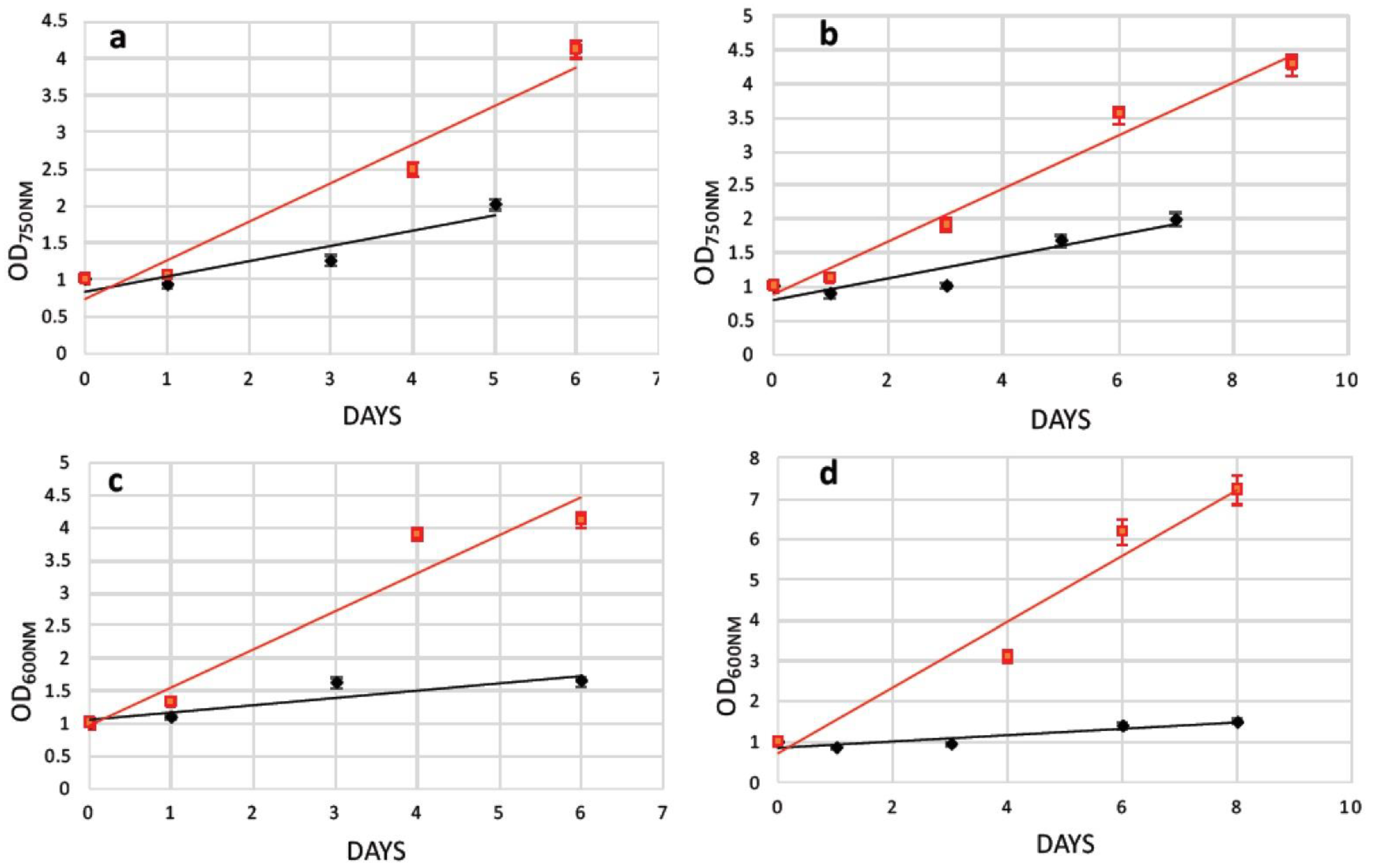
2.3. Transformation of N. oceanica and S. almeriensis Protoplasts with Magnetic Nanoparticles and Their Cultivation
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Protoplast Preparation
4.2.1. Enzymatic Cell Wall Digestion
4.2.2. Glass Beads Cell Wall Disruption
4.3. Scanning Electron Microscopy Imaging for Observation of Morphological Changes in Cells
4.4. Nanoparticles
4.5. Transformation
4.6. Protoplasts Regeneration before and after Electroporation
4.7. Transformation Efficiency
4.8. Prussian Blue Staining
4.9. Magnetic Cells Cultivation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harun, R.; Singh, M.; Forde, G.M.; Danquah, M.K. Bioprocess engineering of microalgae to produce a variety of consumer products. Renew. Sustain. Energy Rev. 2010, 14, 1037–1047. [Google Scholar] [CrossRef]
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Giraldo-Calderón, N.D.; Romo-Buchelly, R.J.; Arbeláez-Pérez, A.A.; Echeverri-Hincapié, D.; Atehortúa-Garcés, L. Microalgae biorefineries: Applications and emerging technologies. DYNA 2018, 85, 219–233. [Google Scholar] [CrossRef]
- Coll, J.M. Review. Methodologies for transferring DNA into eukaryotic microalgae. Span. J. Agric. Res. 2000, 4, 316–330. [Google Scholar] [CrossRef] [Green Version]
- Doron, L.; Segal, N.; Shapira, M. Transgene expression in microalgae—From tools to applications. Front. Plant Sci. 2016, 7, 505. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Lin, H.; Jiang, P. Advances in genetic engineering of marine algae. Biotechnol. Adv. 2012, 30, 1602–1613. [Google Scholar] [CrossRef] [PubMed]
- Faraco, M.; Di Sansebastiano, G.; Pietro, S.K.; Koes, R.E.; Quattrocchio, F.M. One Protoplast Is Not the Other! Plant Physiol. 2011, 156, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Verma, N.; Bansal, M.; Kumar, V. Protoplast fusion technology and its biotechnological applications. Chem. Eng. Trans 2008, 14, 113–120. [Google Scholar]
- Gerken, H.G.; Donohoe, B.; Knoshaug, E.P. Enzymatic cell wall degradation of Chlorella vulgaris and other microalgae for biofuels production. Planta 2013, 237, 239–253. [Google Scholar] [CrossRef]
- Gouveia, L.; Oliveira, A.C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2009, 36, 269–274. [Google Scholar] [CrossRef]
- Hibberd, D.J. Notes on the taxonomy and nomenclature of the algal classes Eustigmatophyceae and Tribophyceae (synonym Xanthophyceae). Bot. J. Linn. Soc. 1981, 82, 93–119. [Google Scholar] [CrossRef]
- Lubián, L.M.; Montero, O.; Moreno-Garrido, I.; Huertas, I.E.; Sobrino, C.; González-Del Valle, M.; Parés, G. Nannochloropsis (Eustigmatophyceae) as source of commercially valuable pigments. J. Appl. Phycol. 2000, 12, 249–255. [Google Scholar] [CrossRef]
- Savvidou, M.G.; Boli, E.; Logothetis, D.; Lymperopoulou, T.; Ferraro, A.; Louli, V.; Mamma, D.; Kekos, D.; Magoulas, K.; Kolisis, F.N. A study on the effect of macro-and micro-nutrients on nannochloropsis oceanica growth, fatty acid composition and magnetic harvesting efficiency. Plants 2020, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.C.; Ho, S.H.; Lee, D.J.; Chen, C.Y.; Huang, C.C.; Chang, J.S. Characterization, extraction and purification of lutein produced by an indigenous microalga Scenedesmus obliquus CNW-N. Biochem. Eng. J. 2013, 78, 24–31. [Google Scholar] [CrossRef]
- Mandal, S.; Mallick, N. Biodiesel production by the green Microalga scenedesmus obliquus in a recirculatory aquaculture system. Appl. Environ. Microbiol. 2012, 78, 5929–5934. [Google Scholar] [CrossRef] [Green Version]
- Mehariya, S.; Iovine, A.; Di Sanzo, G.; Larocca, V.; Martino, M.; Leone, G.P.; Casella, P.; Karatza, D.; Marino, T.; Musmarra, D.; et al. Supercritical fluid extraction of lutein from Scenedesmus almeriensis. Molecules 2019, 7, 1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffree, C.E. The Fine Structure of the Plant Cuticle. Annu. Plant Rev. 2007, 23, 11–125. [Google Scholar]
- Vieler, A.; Wu, G.; Tsai, C.H.; Bullard, B.; Cornish, A.J.; Harvey, C.; Reca, I.B.; Thornburg, C.; Achawanantakun, R.; Buehl, C.J.; et al. Genome, Functional Gene Annotation, and Nuclear Transformation of the Heterokont Oleaginous Alga Nannochloropsis oceanica CCMP1779. PLoS Genet. 2012, 8, e1003064. [Google Scholar] [CrossRef] [Green Version]
- Halim, R.; Hill, D.R.A.; Hanssen, E.; Webley, P.A.; Blackburn, S.; Grossman, A.R.; Posten, C.; Martin, G.J.O. Towards sustainable microalgal biomass processing: Anaerobic induction of autolytic cell-wall self-ingestion in lipid-rich: Nannochloropsis slurries. Green Chem. 2019, 11, 2967–2982. [Google Scholar] [CrossRef]
- Bisalputra, T. The origin of the pectic layer of the cell wall of Scenedesmus Quadricauda. Can. J. Bot. 1965, 43, 1549–1552. [Google Scholar] [CrossRef]
- Nemcova, Y. Detection of cell wall structural polysaccharides by cellulase-gold complexes and Detekce polysacharidu v bunecne stene. Czecg Phycol. 2003, 3, 31–36. [Google Scholar]
- Voigt, J.; Stolarczyk, A.; Zych, M.; Malec, P.; Burczyk, J. The cell-wall glycoproteins of the green alga Scenedesmus obliquus. The predominant cell-wall polypeptide of Scenedesmus obliquus is related to the cell-wall glycoprotein gp3 of Chlamydomonas reinhardtii. Plant Sci. 2014, 215–216, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kong, X.; Wang, Z.; Sun, Y.; Zhu, S.; Li, L.; Lv, P. Optimization of enzymatic hydrolysis for effective lipid extraction from microalgae Scenedesmus sp. Renew. Energy 2018, 125, 1049–1057. [Google Scholar] [CrossRef]
- Echeverri, D.; Romo, J.; Giraldo, N.; Atehortúa, L.; Echeverri, D.; Romo, J.; Giraldo, N.; Atehortúa, L. Microalgae protoplasts isolation and fusion for biotechnology research. Rev. Colomb. Biotecnol. 2019, 21, 101–112. [Google Scholar] [CrossRef]
- Ortiz-Matamoros, M.F.; Villanueva, M.A.; Islas-Flores, T. Genetic transformation of cell-walled plant and algae cells: Delivering DNA through the cell wall. Brief. Funct. Genom. 2018, 17, 26–33. [Google Scholar] [CrossRef]
- Spiden, E.M.; Scales, P.J.; Yap, B.H.J.; Kentish, S.E.; Hill, D.R.A.; Martin, G.J.O. The effects of acidic and thermal pretreatment on the mechanical rupture of two industrially relevant microalgae: Chlorella sp. and Navicula sp. Algal Res. 2015, 7, 5–10. [Google Scholar] [CrossRef]
- Davey, M.R.; Anthony, P.; Power, J.B.; Lowe, K.C. Plant protoplasts: Status and biotechnological perspectives. Biotechnol. Adv. 2005, 23, 131–171. [Google Scholar] [CrossRef] [PubMed]
- Papadakis, A.K.; Roubelakis-Angelakis, K.A. Oxidative stress could be responsible for the recalcitrance of plant protoplasts. Plant Physiol. Biochem. 2002, 40, 549–559. [Google Scholar] [CrossRef]
- Noda, J.; Mühlroth, A.; Bučinská, L.; Dean, J.; Bones, A.M.; Sobotka, R. Tools for biotechnological studies of the freshwater alga Nannochloropsis limnetica: Antibiotic resistance and protoplast production. J. Appl. Phycol. 2017, 29, 853–863. [Google Scholar] [CrossRef]
- Gan, Q.; Jiang, J.; Han, X.; Wang, S.; Lu, Y. Engineering the chloroplast genome of oleaginous marine microalga Nannochloropsis oceanica. Front. Plant Sci. 2018, 9, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abidin, A.A.Z.; Suntarajh, M.; Yusof, Z.N.B. Transformation of a Malaysian species of Nannochloropsis: Gateway to construction of transgenic microalgae as vaccine delivery system to aquatic organisms. Bioengineered 2020, 11, 1071–1079. [Google Scholar] [CrossRef]
- Savvidou, M.G.; Ferraro, A.; Hristoforou, E.; Mamma, D.; Kekos, D.; Kolisis, F.N. Incorporation of Magnetic Nanoparticles into Protoplasts of Microalgae Haematococcus pluvialis: A Tool for Biotechnological Applications. Molecules 2020, 25, 560. [Google Scholar] [CrossRef]
- Scholz, M.J.; Weiss, T.L.; Jinkerson, R.E.; Jing, J.; Roth, R.; Goodenough, U.; Posewitz, M.C.; Gerken, H.G. Ultrastructure and composition of the Nannochloropsis gaditana cell wall. Eukaryot. Cell 2014, 13, 1450–1464. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, T.; Kawai, T.; Kajiwara, T.; Ishida, Y. Protoplast isolation in the marine brown alga Dictyopteris prolifera (Dictyotales). Plant Cell Rep. 1995, 14, 30–33. [Google Scholar] [CrossRef]
- Gupta, V.; Kumar, M.; Kumari, P.; Reddy, C.R.K.; Jha, B. Optimization of protoplast yields from the red algae Gracilaria dura (C. Agardh) J. Agardh and G. verrucosa (Huds.) Papenfuss. J. Appl. Phycol. 2011, 23, 209–218. [Google Scholar] [CrossRef]
- Reddy, C.R.K.; Gupta, M.K.; Mantri, V.A.; Jha, B. Seaweed protoplasts: Status, biotechnological perspectives and needs. J. Appl. Phycol. 2008, 20, 619–632. [Google Scholar] [CrossRef]
- Chen, Y.C.; Shih, H.C. Development of protoplasts of Ulva fasciata (Chlorophyta) for algal seed stock. J. Phycol. 2000, 36, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Reshma, R.; Arumugam, M. Selective degradation of the recalcitrant cell wall of Scenedesmus quadricauda CASA CC202. Planta 2017, 26, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Pan, K.; Zhang, L.; Zhu, B.; Yang, G.; Zhang, X. Genetic transformation of Nannochloropsis oculata with a bacterial phleomycin resistance gene as dominant selective marker. J. Ocean Univ. China 2016, 25, 351–356. [Google Scholar] [CrossRef]
- Lu, Y.; Kong, R.; Hu, L. Preparation of protoplasts from Chlorella protothecoides. World J. Microbiol. Biotechnol. 2012, 28, 1827–1830. [Google Scholar] [CrossRef]
- Klochkova, T.A.; Kwak, M.S.; Kim, G.H. Proteomic profiles and ultrastructure of regenerating protoplast of bryopsis plumosa (Chlorophyta). Algae 2016, 31, 379–390. [Google Scholar] [CrossRef]
- Kim, G.H.; Klotchkova, T.A.; Kang, Y.M. Life without a cell membrane: Regeneration of protoplasts from disintegrated cells of the marine green alga Bryopsis plumosa. J. Cell Sci. 2001, 114, 2009–2014. [Google Scholar] [CrossRef] [PubMed]
- Polakovič, M.; Švitel, J.; Bučko, M.; Filip, J.; Neděla, V.; Ansorge-Schumacher, M.B.; Gemeiner, P. Progress in biocatalysis with immobilized viable whole cells: Systems development, reaction engineering and applications. Biotechnol. Lett. 2017, 39, 667–683. [Google Scholar] [CrossRef]
- Vasilieva, S.G.; Lobakova, E.S.; Lukyanov, A.A.; Solovchenko, A.E. Immobilized microalgae in biotechnology. Ecology 2016, 71, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Tagizadeh, S.M.; Ebrahiminezhad, A.; Ghoshoon, M.B.; Dehshahri, A.; Berenjian, A.; Ghasemi, Y. Impacts of magnetic immobilization on the growth and metabolic status of recombinant Pichia pastoris. Mol. Biotechnol. 2021, 63, 80–89. [Google Scholar] [CrossRef]
- Mann, J.E.; Myers, J. On pigments, growth, and photosynthesis of phaeodactylum tricornutum 2. J. Phycol. 1968, 4, 349–355. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | OD750nm (3rd Day after Recovery) | OD750nm (After Magnetic Separation) | Transformation Efficiency (%) |
|---|---|---|---|
| Enzymatic digestion (8 h) | 0.610 | 0.501 | 17.8 |
| Mechanical treatment | 0.912 | 0.825 | 10.7 |
| Conditions | OD600nm (3rd Day after Recovery) | OD600nm (After Magnetic Separation) | Transformation Efficiency (%) |
|---|---|---|---|
| Enzymatic digestion (8 h) | 0.834 | 0.679 | 18.6 |
| Mechanical treatment | 0.670 | 0.565 | 15.7 |
| Strains | Optical Density (1st Day) | Optical Density (3rd day) | Increment |
|---|---|---|---|
| N. oceanica (OD750nm) | 1.001 | 1.314 | +24% |
| S. almeriensis (OD600nm) | 1.097 | 1.305 | +16% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savvidou, M.G.; Ferraro, A.; Schinas, P.; Mamma, D.; Kekos, D.; Hristoforou, E.; Kolisis, F.N. Magnetic Immobilization and Growth of Nannochloropsis oceanica and Scenedasmus almeriensis. Plants 2022, 11, 72. https://doi.org/10.3390/plants11010072
Savvidou MG, Ferraro A, Schinas P, Mamma D, Kekos D, Hristoforou E, Kolisis FN. Magnetic Immobilization and Growth of Nannochloropsis oceanica and Scenedasmus almeriensis. Plants. 2022; 11(1):72. https://doi.org/10.3390/plants11010072
Chicago/Turabian StyleSavvidou, Maria G., Angelo Ferraro, Petros Schinas, Diomi Mamma, Dimitris Kekos, Evangelos Hristoforou, and Fragiskos N. Kolisis. 2022. "Magnetic Immobilization and Growth of Nannochloropsis oceanica and Scenedasmus almeriensis" Plants 11, no. 1: 72. https://doi.org/10.3390/plants11010072
APA StyleSavvidou, M. G., Ferraro, A., Schinas, P., Mamma, D., Kekos, D., Hristoforou, E., & Kolisis, F. N. (2022). Magnetic Immobilization and Growth of Nannochloropsis oceanica and Scenedasmus almeriensis. Plants, 11(1), 72. https://doi.org/10.3390/plants11010072