
Molecular Genetic Characteristics of Different Scenarios of Xylogenesis on the Example of Two Forms of Silver Birch Differing in the Ratio of Structural Elements in the Xylem

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Features of the Structure of Mature Xylem in Different Forms of Silver Birch
2.2. NAC Family Genes Identification in the Silver Birch Genome
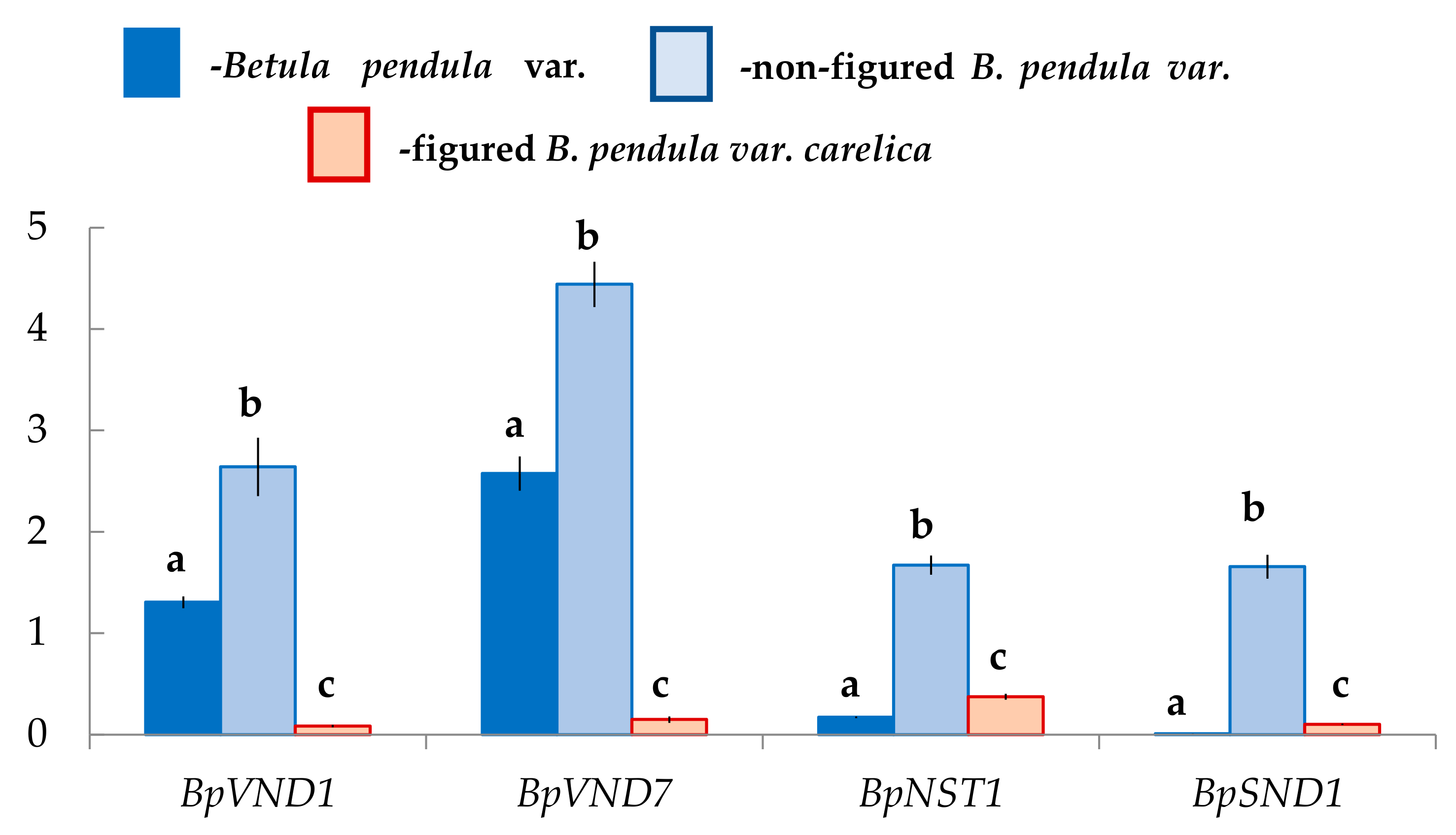
2.3. Expression of the Genes Encoding NAC-Domain Transcription Factors in the Differentiating Xylem of Different Forms of Silver Birch
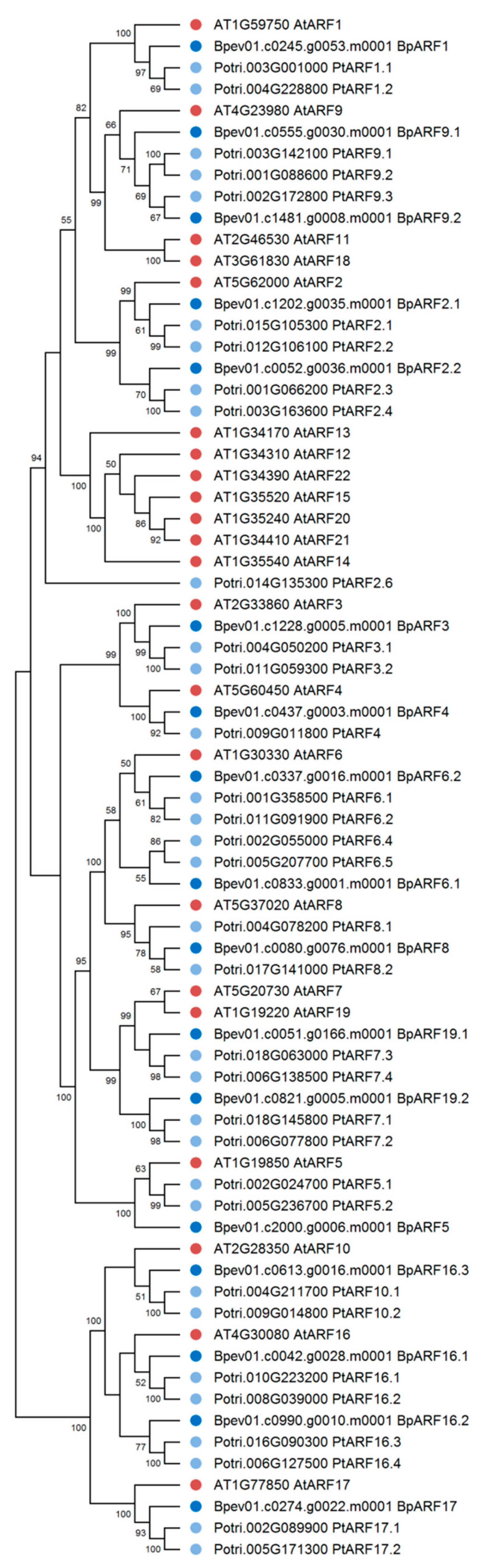
2.4. ARF Family Genes Identification in the Silver Birch Genome
2.5. HD-ZIP III Genes Identification in the Silver Birch Genome
2.6. Expression of the Genes Encoding Auxin-Dependent Transcription Factor BpARF5 and Transcription Factor BpHB8 in the Cambial Zone and Differentiating Xylem of Different Forms of Silver Birch
2.7. AuxRE cis-Elements in Promoters of Studied B. pendula Genes
3. Discussion
3.1. Distribution of BpARF5, BpHB8, BpVND, and BpNST Genes Expression during the Formation of Straight-Grained Wood of Silver Birch
3.2. The Formation of the Auxin-Deficient Phenotype of Karelian Birch Occurs against the Background of a Decrease in the Expression of BpARF5 and BpHB8, and It Is Accompanied by a Change in the Expression of the NAC Family Genes
3.3. Non-Figured Karelian Birch Plants Are of Particular Interest for the Study of the Development of Secondary Conducting Tissues
4. Materials and Methods
4.1. Study Objects
4.2. Plant Sampling
4.3. Microscopy
4.4. Gene Retrieval from the Silver Birch Genome by Bioinformatics Methods
4.5. qRT-PCR
4.6. Analysis of Promoters
4.7. Statistical Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Demura, T.; Ye, Z.-H. Regulation of plant biomass production. Curr. Opin. Plant Biol. 2010, 13, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Carpita, N.C. Progress in the biological synthesis of the plant cell wall: New ideas for improving biomass for bioenergy. Curr. Opin. Biotechnol. 2012, 23, 330–337. [Google Scholar] [CrossRef]
- Yang, J.H.; Wang, H. Molecular mechanisms for vascular development and secondary cell wall formation. Front. Plant Sci. 2016, 7, 356. [Google Scholar] [CrossRef] [PubMed]
- Růžička, K.; Ursache, R.; Hejátko, J.; Helariutta, Y. Xylem development—From the cradle to the grave. New Phytol. 2015, 207, 519–535. [Google Scholar] [CrossRef]
- Kondo, Y.; Tamaki, T.; Fukuda, H. Regulation of xylem cell fate. Front. Plant Sci. 2014, 5, 315. [Google Scholar] [CrossRef]
- Ye, Z.-H.; Zhong, R. Molecular control of wood formation in trees. J. Exp. Bot. 2015, 66, 4119–4131. [Google Scholar] [CrossRef]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.-C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Birol, I.; Raymond, A.; Jackman, S.D.; Pleasance, S.; Coope, R.; Taylor, G.A.; Yuen, M.M.S.; Keeling, C.I.; Brand, D.; Vandervalk, B.P.; et al. Assembling the 20 Gb white spruce (picea glauca) genome from whole-genome shotgun sequencing data. Bioinformatics 2013, 29, 1492–1497. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus Trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef]
- Myburg, A.A.; Grattapaglia, D.; Tuskan, G.A.; Hellsten, U.; Hayes, R.D.; Grimwood, J.; Jenkins, J.; Lindquist, E.; Tice, H.; Bauer, D.; et al. The genome of eucalyptus grandis. Nature 2014, 510, 356–362. [Google Scholar] [CrossRef]
- Salojärvi, J.; Smolander, O.-P.; Nieminen, K.; Rajaraman, S.; Safronov, O.; Safdari, P.; Lamminmäki, A.; Immanen, J.; Lan, T.; Tanskanen, J.; et al. Genome sequencing and population genomic analyses provide insights into the adaptive landscape of silver birch. Nat. Genet. 2017, 49, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, Y.; Yu, L.; Zheng, T.; Wang, S.; Yue, Z.; Jiang, J.; Kumari, S.; Zheng, C.; Tang, H.; et al. Genome sequence and evolution of Betula Platyphylla. Hortic. Res. 2021, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Jansson, S.; Douglas, C.J. Populus: A model system for plant biology. Ann. Rev. Plant Biol. 2007, 58, 435–458. [Google Scholar] [CrossRef]
- Douglas, C.J. Populus as a model tree. In Comparative and Evolutionary Genomics of Angiosperm Trees; Groover, A., Cronk, Q., Eds.; Plant Genetics and Genomics: Crops and Models; Springer International Publishing: Cham, Switzerland, 2017; Volume 21, pp. 61–84. ISBN 978-3-319-49327-5. [Google Scholar]
- Rodgers-Melnick, E.; Mane, S.P.; Dharmawardhana, P.; Slavov, G.T.; Crasta, O.R.; Strauss, S.H.; Brunner, A.M.; DiFazio, S.P. Contrasting patterns of evolution following whole genome versus tandem duplication events in Populus. Genome Res. 2012, 22, 95–105. [Google Scholar] [CrossRef]
- Wheeler, E.A. Inside wood—A web resource for hardwood anatomy. IAWA J. 2011, 32, 199–211. [Google Scholar] [CrossRef]
- Morris, H.; Brodersen, C.; Schwarze, F.W.M.R.; Jansen, S. The parenchyma of secondary xylem and its critical role in tree defense against fungal decay in relation to the CODIT model. Front. Plant Sci. 2016, 7, 1665. [Google Scholar] [CrossRef]
- Hynynen, J.; Niemisto, P.; Vihera-Aarnio, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of Birch (Betula Pendula Roth and Betula Pubescens Ehrh.) in Northern Europe. Forestry 2009, 83, 103–119. [Google Scholar] [CrossRef]
- Hagqvist, R.; Mikkola, A. Visakoivun Kasvatus Ja Käyttö; Metsäkustannus & Visaseurary: Hämeenlinna, Finland, 2008; ISBN 978-952-5694-03-1. [Google Scholar]
- Lyubavskaya, A.Y. Karelian Birch; Publishing House of Moscow State Forest University: Moscow, Russia, 2006. [Google Scholar]
- Novitskaya, L.; Nikolaeva, N.; Galibina, N.; Tarelkina, T.; Semenova, L. The greatest density of parenchyma inclusions in Karelian Birch wood occurs at confluences of phloem flows. Silva Fenn. 2016, 50, 1461–1478. [Google Scholar] [CrossRef][Green Version]
- Novitskaya, L.L.; Shulyakovskaya, T.A.; Galibina, N.A.; Ilyinova, M.K. Membrane lipid composition upon normal and patterned wood formation in Betula Pendula Roth. J. Plant Growth Regul. 2018, 37, 958–970. [Google Scholar] [CrossRef]
- Novitskaya, L.; Nikolaeva, N.; Tarelkina, T. Endogenous variability of the figured wood of Karelian Birch. Wulfenia 2016, 23, 175–188. [Google Scholar]
- Novitskaya, L.L. Karelian Birch: Mechanisms of Growth and Development of Structural Abnormalities; Verso: Petrozavodsk, Russia, 2008; ISBN 978-5-85039-229-1. [Google Scholar]
- Galibina, N.A.; Novitskaya, L.L.; Nikerova, K.M.; Moshkina, E.V.; Moshchenskaya, Y.L.; Borodina, M.N.; Sofronova, I.N.; Nikolaeva, N.N. Labile nitrogen availability in soil influences the expression of wood pattern in Karelian birch. Bot. Zhurnal 2019, 104, 1598–1609. [Google Scholar] [CrossRef]
- Paganova, V. Analysis of inheritance and growth of curly birch progenies from controlled hybridisation and possibilities of their utilisation for timber production in agricultural landscape. Czech J. Genet. Plant Breed. 2004, 40, 51–62. [Google Scholar] [CrossRef]
- Yermakov, V.I. Mechanisms of Adaptation of Birch to the Conditions of the North; Nauka: Leningrad, Russia, 1986. [Google Scholar]
- Novitskaya, L.L.; Tarelkina, T.V.; Galibina, N.A.; Moshchenskaya, Y.L.; Nikolaeva, N.N.; Nikerova, K.M.; Podgornaya, M.N.; Sofronova, I.N.; Semenova, L.I. The formation of structural abnormalities in Karelian Birch wood is associated with auxin inactivation and disrupted basipetal auxin transport. J. Plant Growth Regul. 2020, 39, 378–394. [Google Scholar] [CrossRef]
- Barilskaya, L.A. Comparative Analysis of Silver Birch and Karelian Birch Wood. Ph.D. Thesis, Forest Research Institute, Petrozavodsk, Russia, 1978. [Google Scholar]
- Galibina, N.A.; Novitskaya, L.L.; Krasavina, M.S.; Moshchenskaya, Y.L. Activity of sucrose synthase in trunk tissues of Karelian Birch during cambial growth. Russ. J. Plant Physiol. 2015, 62, 381–389. [Google Scholar] [CrossRef]
- Galibina, N.A.; Novitskaya, L.L.; Nikerova, K.M.; Moshchenskaya, Y.L.; Borodina, M.N.; Sofronova, I.N. Apoplastic invertase activity regulation in the cambial zone of Karelian Birch. Russ. J. Dev. Biol. 2019, 50, 20–29. [Google Scholar] [CrossRef]
- Galibina, N.A.; Novitskaya, L.L.; Krasavina, M.S.; Moshchenskaya, J.L. Invertase activity in trunk tissues of Karelian Birch. Russ. J. Plant Physiol. 2015, 62, 753–760. [Google Scholar] [CrossRef]
- Galibina, N.A.; Novitskaya, L.L.; Nikerova, K.M. Source–sink relations in the organs and tissues of silver birch during different scenarios of xylogenesis. Russ. J. Plant Physiol. 2019, 66, 308–315. [Google Scholar] [CrossRef]
- Kubo, M.; Udagawa, M.; Nishikubo, N.; Horiguchi, G.; Yamaguchi, M.; Ito, J.; Mimura, T.; Fukuda, H.; Demura, T. Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev. 2005, 19, 1855–1860. [Google Scholar] [CrossRef]
- Zhong, R.; Demura, T.; Ye, Z.-H. SND1, a NAC domain transcription factor, is a key regulator of secondary wall synthesis in fibers of Arabidopsis. Plant Cell 2006, 18, 3158–3170. [Google Scholar] [CrossRef]
- Nakano, Y.; Yamaguchi, M.; Endo, H.; Rejab, N.A.; Ohtani, M. NAC-MYB-based transcriptional regulation of secondary cell wall biosynthesis in land plants. Front. Plant Sci. 2015, 6, 288. [Google Scholar] [CrossRef]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef]
- Petricka, J.J.; Winter, C.M.; Benfey, P.N. Control of Arabidopsis root development. Ann. Rev. Plant Biol. 2012, 63, 563–590. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Demura, T. Transcriptional regulation of secondary wall formation controlled by NAC domain proteins. Plant Biotechnol. 2010, 27, 237–242. [Google Scholar] [CrossRef]
- Fang, Y.; You, J.; Xie, K.; Xie, W.; Xiong, L. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol. Genet. Genom. 2008, 280, 547–563. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC Transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2012, 1819, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lin, X.; Zhang, D.; Li, Q.; Zhao, X.; Chen, S. Genome-wide analysis of NAC gene family in Betula Pendula. Forests 2019, 10, 741. [Google Scholar] [CrossRef]
- Mitsuda, N.; Iwase, A.; Yamamoto, H.; Yoshida, M.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 2007, 19, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Richardson, E.A.; Ye, Z.-H. Two NAC domain transcription factors, SND1 and NST1, function redundantly in regulation of secondary wall synthesis in fibers of arabidopsis. Planta 2007, 225, 1603–1611. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Kubo, M.; Fukuda, H.; Demura, T. Vascular-related NAC-DOMAIN7 is involved in the differentiation of all types of xylem vessels in arabidopsis roots and shoots. Plant J. 2008, 55, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhong, R.; Ye, Z.-H. Arabidopsis NAC domain proteins, VND1 to VND5, are transcriptional regulators of secondary wall biosynthesis in vessels. PLoS ONE 2014, 9, e105726. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Yamaguchi, M.; Tamura, T.; Nakano, Y.; Nishikubo, N.; Yoneda, A.; Kato, K.; Kubo, M.; Kajita, S.; Katayama, Y.; et al. Multiple classes of transcription factors regulate the expression of Vascular-related NAC-DOMAIN7, a master switch of xylem vessel differentiation. Plant Cell Physiol. 2015, 56, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Sorce, C.; Giovannelli, A.; Sebastiani, L.; Anfodillo, T. Hormonal signals involved in the regulation of cambial activity, xylogenesis and vessel patterning in trees. Plant Cell Rep. 2013, 32, 885–898. [Google Scholar] [CrossRef]
- Aloni, R. Ecophysiological implications of vascular differentiation and plant evolution. Trees 2015, 29, 1–16. [Google Scholar] [CrossRef]
- Sundberg, B.; Uggla, C.; Tuominen, H. Cambial growth and auxin gradients. In Cell and Molecular Biology of Wood Formation; Savidge, R.A., Barnett, J.R., Napier, R., Eds.; BIOS Scientific Publishers Limited: Oxford, UK, 2000; pp. 169–188. [Google Scholar]
- Robert, H.S.; Grones, P.; Stepanova, A.N.; Robles, L.M.; Lokerse, A.S.; Alonso, J.M.; Weijers, D.; Friml, J. Local auxin sources orient the apical-basal axis in arabidopsis embryos. Curr. Biol. 2013, 23, 2506–2512. [Google Scholar] [CrossRef]
- Hardtke, C.S.; Berleth, T. The arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. EMBO J. 1998, 17, 1405–1411. [Google Scholar] [CrossRef]
- Hamann, T.; Benkova, E.; Bäurle, I.; Kientz, M.; Jürgens, G. The arabidopsis BODENLOS gene encodes an auxin response protein inhibiting MONOPTEROS-mediated embryo patterning. Genes Dev. 2002, 16, 1610–1615. [Google Scholar] [CrossRef]
- Xu, C.; Shen, Y.; He, F.; Fu, X.; Yu, H.; Lu, W.; Li, Y.; Li, C.; Fan, D.; Wang, H.C.; et al. Auxin-mediated Aux/IAA-ARF-HB signaling cascade regulates secondary xylem development in Populus. New Phytol. 2019, 222, 752–767. [Google Scholar] [CrossRef]
- Johnsson, C.; Jin, X.; Xue, W.; Dubreuil, C.; Lezhneva, L.; Fischer, U. The plant hormone auxin directs timing of xylem development by inhibition of secondary cell wall deposition through repression of secondary wall NAC-domain transcription factors. Physiol. Plant 2019, 165, 673–689. [Google Scholar] [CrossRef]
- Fischer, U.; Kucukoglu, M.; Helariutta, Y.; Bhalerao, R.P. The dynamics of cambial stem cell activity. Ann. Rev. Plant Biol. 2019, 70, 293–319. [Google Scholar] [CrossRef]
- Schrader, J.; Davis, J.; Mellerowicz, E.; Berglund, A.; Nilsson, P.; Hertzberg, M.; Sandberg, G. A High-resolution transcript profile across the wood-forming meristem of poplar identifies potential regulators of cambial stem cell identity. Plant Cell 2004, 16, 2278–2292. [Google Scholar] [CrossRef]
- Pascual, M.B.; de la Torre, F.; Cañas, R.A.; Cánovas, F.M.; Ávila, C. NAC transcription factors in woody plants. In Progress in Botany; Cánovas, F.M., Lüttge, U., Matyssek, R., Pretzsch, H., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 80, pp. 195–222. ISBN 978-3-030-10760-4. [Google Scholar]
- Cherenkov, P.; Novikova, D.; Omelyanchuk, N.; Levitsky, V.; Grosse, I.; Weijers, D.; Mironova, V. Diversity of cis-regulatory elements associated with auxin response in arabidopsis thaliana. J. Exp. Bot. 2018, 69, 329–339. [Google Scholar] [CrossRef]
- Galibina, N.A. Endogenous Mechanisms of Regulation of Xylogenesis in Woody Plants on the Example of Two Forms of Birch. Ph.D. Thesis, Komarov Botanical Institute of the Russian Academy of Sciences, St. Petersburg, Russia, 2018. [Google Scholar]
- Tarelkina, T.V.; Galibina, N.A.; Moshchenskaya, Y.L.; Nikerova, K.M.; Sofronova, I.N.; Nikolaeva, N.N.; Ivanova, D.S.; Semenova, L. Expression of genes involved in the transport and conjugation of auxin at different stages of cambial derivatives differentiation in two forms of Silver Birch. 2021. In Preparation. [Google Scholar]
- Ohashi-Ito, K.; Oda, Y.; Fukuda, H. Arabidopsis vascular-related NAC-DOMAIN6 directly regulates the genes that govern programmed cell death and secondary wall formation during xylem differentiation. Plant Cell 2010, 22, 3461–3473. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; Ye, Z.-H. Global analysis of direct targets of secondary wall NAC master switches in Arabidopsis. Mol. Plant 2010, 3, 1087–1103. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Mitsuda, N.; Ohtani, M.; Ohme-Takagi, M.; Kato, K.; Demura, T. Vascular-related NAC-DOMAIN 7 directly regulates the expression of a broad range of genes for xylem vessel formation: Direct target genes of VND7. Plant J. 2011, 66, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Mitsuda, N.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence. Plant Cell 2005, 17, 2993–3006. [Google Scholar] [CrossRef]
- Zhong, R.; Lee, C.; Ye, Z.-H. Functional characterization of poplar wood-associated NAC domain transcription factors. Plant Physiol. 2010, 152, 1044–1055. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; McCarthy, R.L.; Reeves, C.K.; Jones, E.G.; Ye, Z.-H. Transcriptional activation of secondary wall biosynthesis by rice and maize NAC and MYB transcription factors. Plant Cell Physiol. 2011, 52, 1856–1871. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, M.; Nishikubo, N.; Xu, B.; Yamaguchi, M.; Mitsuda, N.; Goué, N.; Shi, F.; Ohme-Takagi, M.; Demura, T. A NAC domain protein family contributing to the regulation of wood formation in poplar: NAC domain protein family regulates wood formation. Plant J. 2011, 67, 499–512. [Google Scholar] [CrossRef]
- Brackmann, K.; Qi, J.; Gebert, M.; Jouannet, V.; Schlamp, T.; Grünwald, K.; Wallner, E.-S.; Novikova, D.D.; Levitsky, V.G.; Agustí, J.; et al. Spatial specificity of auxin responses coordinates wood formation. Nat. Commun. 2018, 9, 875. [Google Scholar] [CrossRef]
- Doley, D.; Leyton, L. Effects of growth regulating substances and water potential on the development of secondary xylem in Fraxinus. New Phytol. 1968, 67, 579–594. [Google Scholar] [CrossRef]
- Aloni, R.; Zimmermann, M.H. The control of vessel size and density along the plant axis—A new hypothesis. Differentiation 1983, 24, 203–208. [Google Scholar] [CrossRef]
- Junghans, U.; Langenfeld-Heyser, R.; Polle, A.; Teichmann, T. Effect of auxin transport inhibitors and ethylene on the wood anatomy of poplar. Plant Biol. 2004, 6, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Aloni, R. The induction of vascular tissues by auxin. In Plant Hormones. Biosynthesis, Signal Transduction, Action! Davis, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 485–518. [Google Scholar]
- Sachs, T. Integrating cellular and organismic aspects of vascular differentiation. Plant Cell Physiol. 2000, 41, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Shchetinkin, S.V. Histogenesis of Figured Wood in Birch (Betula Pendula Roth Var. Carelica Merkl. and Betula Pendula Roth). Ph.D. Thesis, Voronezh State University, Voronezh, Russia, 1987. [Google Scholar]
- Tarelkina, T.V.; Novitskaya, L.L.; Galibina, N.A.; Moshchenskaya, Y.L.; Nikerova, K.M.; Nikolaeva, N.N.; Sofronova, I.N.; Ivanova, D.S.; Semenova, L.I. Expression analysis of key auxin biosynthesis, transport, and metabolism genes of Betula Pendula with special emphasis on figured wood formation in Karelian Birch. Plants 2020, 9, 1406. [Google Scholar] [CrossRef]
- Guilfoyle, T.J.; Hagen, G. Auxin response factors. Curr. Opin. Plant Biol. 2007, 10, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-B.; Xie, Z.-Z.; Hu, C.-G.; Zhang, J.-Z. A review of auxin response factors (ARFs) in plants. Front. Plant Sci. 2016, 7, 47. [Google Scholar] [CrossRef]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef]
- Normanly, J. Approaching cellular and molecular resolution of auxin biosynthesis and metabolism. Cold Spring Harb. Perspect. Biol. 2010, 2, a001594. [Google Scholar] [CrossRef]
- Ljung, K. Auxin metabolism and homeostasis during plant development. Development 2013, 140, 943–950. [Google Scholar] [CrossRef]
- Mollenhauer, H.H. Plastic embedding mixtures for use in electron microscopy. Stain Technol. 1964, 39, 111–114. [Google Scholar] [PubMed]
- IAWA. List of microscopic features for hardwood identification. IAWA Bull. 1989, 10, 219–332. [Google Scholar]
- Scholz, A.; Klepsch, M.; Karimi, Z.; Jansen, S. How to quantify conduits in wood? Front. Plant Sci. 2013, 4, 56. [Google Scholar] [CrossRef] [PubMed]
- Angyalossy, V.; Pace, M.R.; Evert, R.F.; Marcati, C.R.; Oskolski, A.A.; Terrazas, T.; Kotina, E.; Lens, F.; Mazzoni-Viveiros, S.C.; Angeles, G.; et al. IAWA list of microscopic bark features. IAWA J. 2016, 37, 517–615. [Google Scholar] [CrossRef]
- Zieminska, K.; Butler, D.W.; Gleason, S.M.; Wright, I.J.; Westoby, M. Fibre wall and lumen fractions drive wood density variation across 24 Australian angiosperms. AoB Plants 2013, 5, plt046. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bryant, S.H. CD-search: Protein domain annotations on the fly. Nucleic Acids Res. 2004, 32, W327–W331. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 978-0-19-513584-8. [Google Scholar]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zang, B.; Yao, H.S.; Huang, M.R. Isolation of high quality RNA and molecular manipulations with various tissues of Populus. Russ. J. Plant Physiol. 2009, 56, 716–719. [Google Scholar] [CrossRef]
- Moshchenskaya, Y.L.; Galibina, N.A.; Tarelkina, T.V.; Nikerova, K.M.; Chirva, O.V.; Novitskaya, L.L. Selection of reference genes for normalization of quantitative PCR data in real time in two forms of Silver Birch. Russ. J. Plant Physiol. 2021, 68, 214–221. [Google Scholar] [CrossRef]
- Rebrikov, D.V.; Korostin, D.O.; Ushakov, V.L.; Barsova, E.V.; Lukyanov, S.A. Application of Modern Methods of Molecular Biology for the Search and Cloning of Full-Length Nucleotide Sequences of CDNA; Publishing House of the NNIU MEPhI: Moscow, Russia, 2011. [Google Scholar]
- Chang, E.; Shi, S.; Liu, J.; Cheng, T.; Xue, L.; Yang, X.; Yang, W.; Lan, Q.; Jiang, Z. Selection of reference genes for quantitative gene expression studies in Platycladus Orientalis (Cupressaceae) using real-time PCR. PLoS ONE 2012, 7, e33278. [Google Scholar] [CrossRef] [PubMed]
- Chow, C.-N.; Lee, T.-Y.; Hung, Y.-C.; Li, G.-Z.; Tseng, K.-C.; Liu, Y.-H.; Kuo, P.-L.; Zheng, H.-Q.; Chang, W.-C. PlantPAN3.0: A New and updated resource for reconstructing transcriptional regulatory networks from ChIP-Seq experiments in plants. Nucleic Acids Res. 2019, 47, D1155–D1163. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | B. pendula Protein Name | Protein Length (aa) | Genome Location | Closest Homolog A. thaliana (% Identity) | Closest Homolog P. trichocarpa (% Identity) |
|---|---|---|---|---|---|
| Bpev01.c0022.g0005 | BpVND1 | 394 | Chr4: 1,788,544–1,796,957 | AtVND1 (57.3%) | PtVNS01 (73.5%) |
| Bpev01.c0411.g0006 | BpVND7 | 319 | Chr7: 25,758,566–25,760,964 | AtVND7 (55.1%) | PtVNS07 (69.2%) |
| Bpev01.c0001.g0043 | BpNST1 | 385 | Chr5: 20,598,169–20,599,517 | AtNST1 (54.0%) | PtVNS10 (65.9%) |
| Bpev01.c0522.g0030 | BpSND1 | 426 | Chr14: 273,787–275,254 | AtSND1 (48.9%) | PtVNS11 (69.2%) |
| Gene ID | B. pendula Protein Name | Protein Length (aa) | Genome Location | Closest Homolog A. thaliana (% Identity) | Closest Homolog P. trichocarpa (% Identity) |
|---|---|---|---|---|---|
| Bpev01.c0042.g0028 | BpARF16.1 | 688 | Chr5: 5,076,067–5,078,769 | AtARF16 (59.6%) | PtARF16.2 (67.6%) |
| Bpev01.c0051.g0166 | BpARF19.1 | 1135 | Chr10: 19,836,755–19,845,827 | AtARF19 (65.2%) | PtARF7.3 (74.8%) |
| Bpev01.c0052.g0036 | BpARF2.2 | 825 | Contig52: 293,588–301,375 | AtARF2 (47.2%) | PtARF2.3 (48.8%) |
| Bpev01.c0080.g0076 | BpARF8 | 849 | Chr2: 10,026,633–10,046,765 | AtARF8 (68.2%) | PtARF8.1 (79.6%) |
| Bpev01.c0245.g0053 | BpARF1 | 718 | Chr3: 494,058–504,888 | AtARF1 (67.3%) | PtARF1.2 (75.7%) |
| Bpev01.c0274.g0022 | BpARF17 | 592 | Chr1: 38,790,620–38,794,987 | AtARF17 (50.5%) | BpARF17.1 (65.3%) |
| Bpev01.c0337.g0016 | BpARF6.2 | 912 | Chr6: 7,510,485–7,517,394 | AtARF6 (73.0%) | PtARF6.2 (79.9%) |
| Bpev01.c0437.g0003 | BpARF4 | 799 | Chr13: 14,735,340–14,742,867 | AtARF4 (62.5%) | PtARF4 (71.2%) |
| Bpev01.c0555.g0030 | BpARF9.1 | 683 | Chr14: 1,409,167–1,413,267 | AtARF9 (56.9%) | PtARF9.1 (71.6%) |
| Bpev01.c0613.g0016 | BpARF16.3 | 713 | Chr13: 13,624,694–13,628,188 | AtARF16 (55.3%) | PtARF16.2 (61.1%) |
| Bpev01.c0821.g0005 | BpARF19.2 | 1109 | Chr7: 4,843,289–4,851,608 | AtARF19 (48.9%) | PtARF7.1 (79.8%) |
| Bpev01.c0833.g0001 | BpARF6.1 | 894 | Chr2: 16,657,839–16,679,834 | AtARF6 (67.6%) | PtARF6.5 (80.8%) |
| Bpev01.c0990.g0010 | BpARF16.2 | 702 | Chr8: 5,039,586–5,042,474 | AtARF16 (58.6%) | PtARF16.4 (76.2%) |
| Bpev01.c1202.g0035 | BpARF2.1 | 841 | Chr12: 25,724,880–25,729,200 | AtARF2 (66.6%) | PtARF2.2 (75.3%) |
| Bpev01.c1228.g0005 | BpARF3 | 753 | Chr6: 23,740,690–23,746,958 | AtARF3 (47.5%) | PtARF3.1 (64.1%) |
| Bpev01.c1481.g0008 | BpARF9.2 | 690 | Chr5: 21,299,075–21,303,311 | AtARF9 (55.1%) | PtARF9.1 (68.7%) |
| Bpev01.c2000.g0006 | BpAFR5 | 933 | Chr9: 2,615,628–2,622,870 | AtARF5 (58.9%) | PtARF5.2 (70.5%) |
| Gene ID | B. pendula Protein Name | Protein Length (aa) | Genome Location | Closest Homolog A. thaliana (% Identity) | Closest Homolog P. trichocarpa (% Identity) |
|---|---|---|---|---|---|
| Bpev01.c0135.g0014 | BpCORONA | 836 | Chr5: 19,578,340–19,586,093 | AtCORONA/ATHB15 (88.3%) | PtHB6 (92.7%) |
| Bpev01.c0371.g0002 | BpHB8 | 837 | Chr9: 13,192,071–13,200,917 | AtHB8 (80.8%) | PtHB8 (90.0%) |
| Bpev01.c0613.g0006 | BpREVOLUTA | 843 | Chr13: 13,883,876–13,890,572 | AtREVOLUTA/IFL1 (84.6%) | PtHB1 (86.3%) |
| Bpev01.c1631.g0001 | BpPHABULOSA | 833 | Chr6: 9,307,551–9,315,088 | AtPHABULOSA/ATHB14 (80.7%) | PtHB4 (91.7%) |
| Gene Name | Known AuxREs | Putative AuxREs | ||
|---|---|---|---|---|
| TGTCTC | TGTCCC | TGTGGG | GTCCCC | |
| BpHB8 | 1273 (−), 471 (+) | 470 (+) | ||
| BpVND1 | 458 (−), 46 (+) | 1666 (−), 447 (+) | ||
| BpVND7 | 805 (−), 703 (−), 28 (+) | 1956 (+) | 1427 (+), 372 (−), 308 (−) | |
| BpNST1 | 908 (−), 601 (−) | 1554 (−) | 491 (−) | |
| BpSND1 | 1630 (−) | 1746 (+), 288 (−) | 709 (+) | |
| Gene Name | Gene ID | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Tm, °C |
|---|---|---|---|---|
| Ef1a | Bpev01.c0437.g0013 | TCCTTGAGGCTCTTGACTTG | ATACCAGGCTTGATGACACC | 89.0 |
| GAPDH | Bpev01.c1040.g0016 | AGAATACAAGCCAGAACTCAAC | CTCTACCACCTCTCCAATCC | 85.9 |
| BpARF5 | Bpev01.c2000.g0006 | AGCGACACCTTCTCACAACTG | GCCTCACACCAACCAATAACTG | 84.9 |
| BpHB8 | Bpev01.c0371.g0002 | GTAGTGGAGTGGATGAGAATG | TCAAGAGCAGAGGCAAGG | 88.0 |
| BpVND1 | Bpev01.c0022.g0005 | AAGAGATAGAAGCGGATA | TTATTAACGGCAGAGATG | 76.8 |
| BpVND7 | Bpev01.c0411.g0006 | TTATGAACAGAATGAGTGGTA | GCAGTGGCTCTATTAGTC | 74.3 |
| BpNST1 | Bpev01.c0001.g0043 | AGTATATCAGTCAATGGA | AGTATATCAGTCAATGGA | 77.2 |
| BpSND1 | Bpev01.c0522.g0030 | AACTTCTACACTACTACC | CACATCTCTTGAATATCC | 76.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galibina, N.A.; Tarelkina, T.V.; Chirva, O.V.; Moshchenskaya, Y.L.; Nikerova, K.M.; Ivanova, D.S.; Semenova, L.I.; Serkova, A.A.; Novitskaya, L.L. Molecular Genetic Characteristics of Different Scenarios of Xylogenesis on the Example of Two Forms of Silver Birch Differing in the Ratio of Structural Elements in the Xylem. Plants 2021, 10, 1593. https://doi.org/10.3390/plants10081593
Galibina NA, Tarelkina TV, Chirva OV, Moshchenskaya YL, Nikerova KM, Ivanova DS, Semenova LI, Serkova AA, Novitskaya LL. Molecular Genetic Characteristics of Different Scenarios of Xylogenesis on the Example of Two Forms of Silver Birch Differing in the Ratio of Structural Elements in the Xylem. Plants. 2021; 10(8):1593. https://doi.org/10.3390/plants10081593
Chicago/Turabian StyleGalibina, Natalia A., Tatiana V. Tarelkina, Olga V. Chirva, Yulia L. Moshchenskaya, Kseniya M. Nikerova, Diana S. Ivanova, Ludmila I. Semenova, Aleksandra A. Serkova, and Ludmila L. Novitskaya. 2021. "Molecular Genetic Characteristics of Different Scenarios of Xylogenesis on the Example of Two Forms of Silver Birch Differing in the Ratio of Structural Elements in the Xylem" Plants 10, no. 8: 1593. https://doi.org/10.3390/plants10081593
APA StyleGalibina, N. A., Tarelkina, T. V., Chirva, O. V., Moshchenskaya, Y. L., Nikerova, K. M., Ivanova, D. S., Semenova, L. I., Serkova, A. A., & Novitskaya, L. L. (2021). Molecular Genetic Characteristics of Different Scenarios of Xylogenesis on the Example of Two Forms of Silver Birch Differing in the Ratio of Structural Elements in the Xylem. Plants, 10(8), 1593. https://doi.org/10.3390/plants10081593