Seed and seedling availability are the most limiting factors for restoration of high-value Tamaulipan thornforest habitat in the Lower Rio Grande Valley of south Texas. Seed supply depends on wild collection, which is labor-intensive and requires significant expertise. This challenge is compounded by low or highly variable germination rates of many thornforest species, and by knowledge gaps regarding the germination requirements and best horticultural practices for propagating thornforest species from seed. This study addressed key knowledge gaps and focused on three ecologically important thornforest species (Ebenopsis, Cordia, and Zanthoxylum) but encountered challenges common in thornforest seedling production. Germination rates were so low for two focal species that experiments yielded limited or no useful results. However, all Ebenopsis experiments were successful, and our investigation of gibberellic acid treatments and heat cracking of seed coats for Cordia yielded valuable data and compelling results.
3.1. Ebenopsis ebano (Texas ebony)
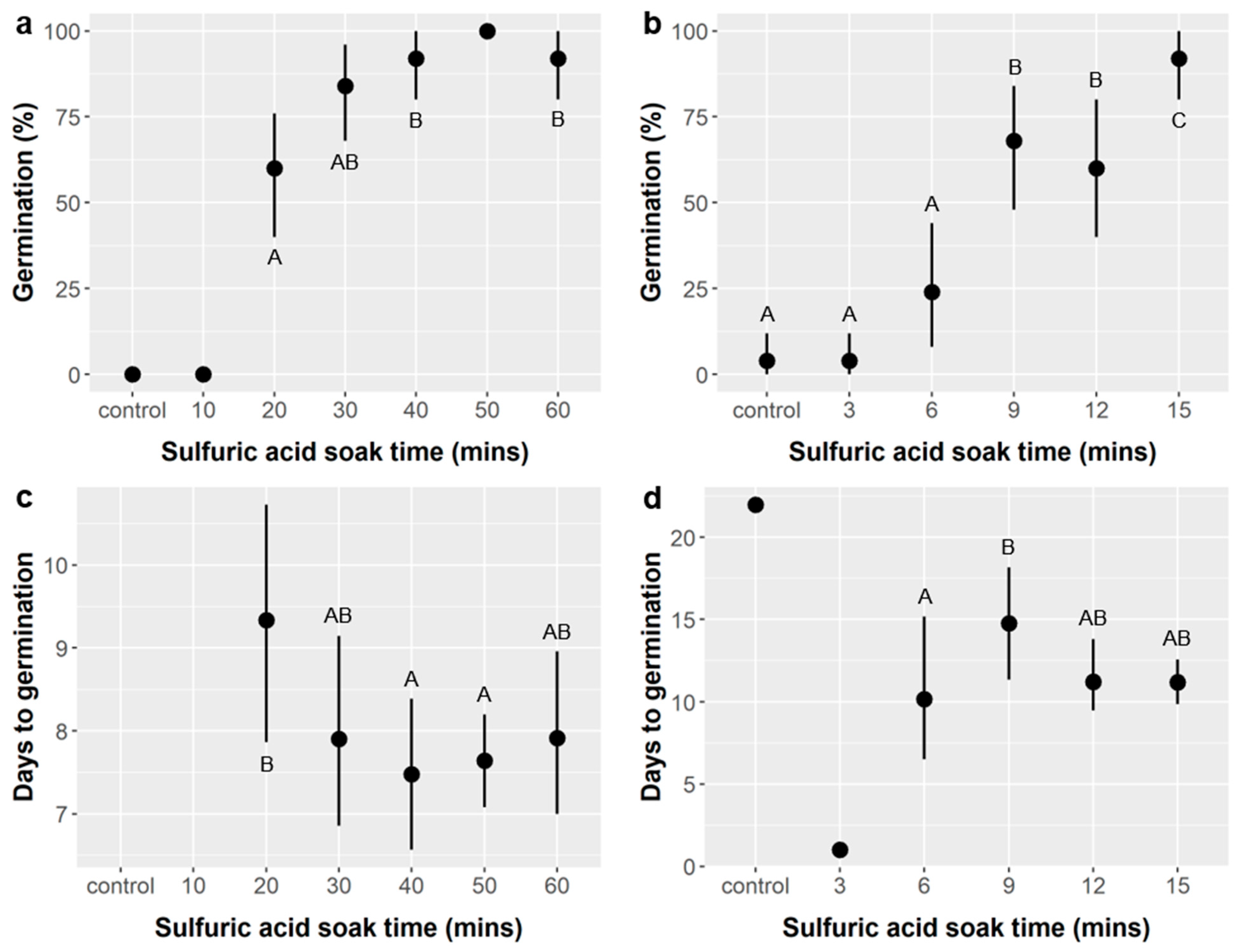
Ebenopsis seeds responded very strongly to sulfuric acid (SA) treatments. This was consistent with prior studies [
33] and current recommendations [
22,
31], and reflects germination requirements of other Fabaceae species with thick seed coats like
Ebenopsis [
32]. However, we identified an optimum SA soak time of 40–60 min, which is longer than the currently recommended SA soak time of 30–35 min [
31] and can be easily replicated by commercial growers (
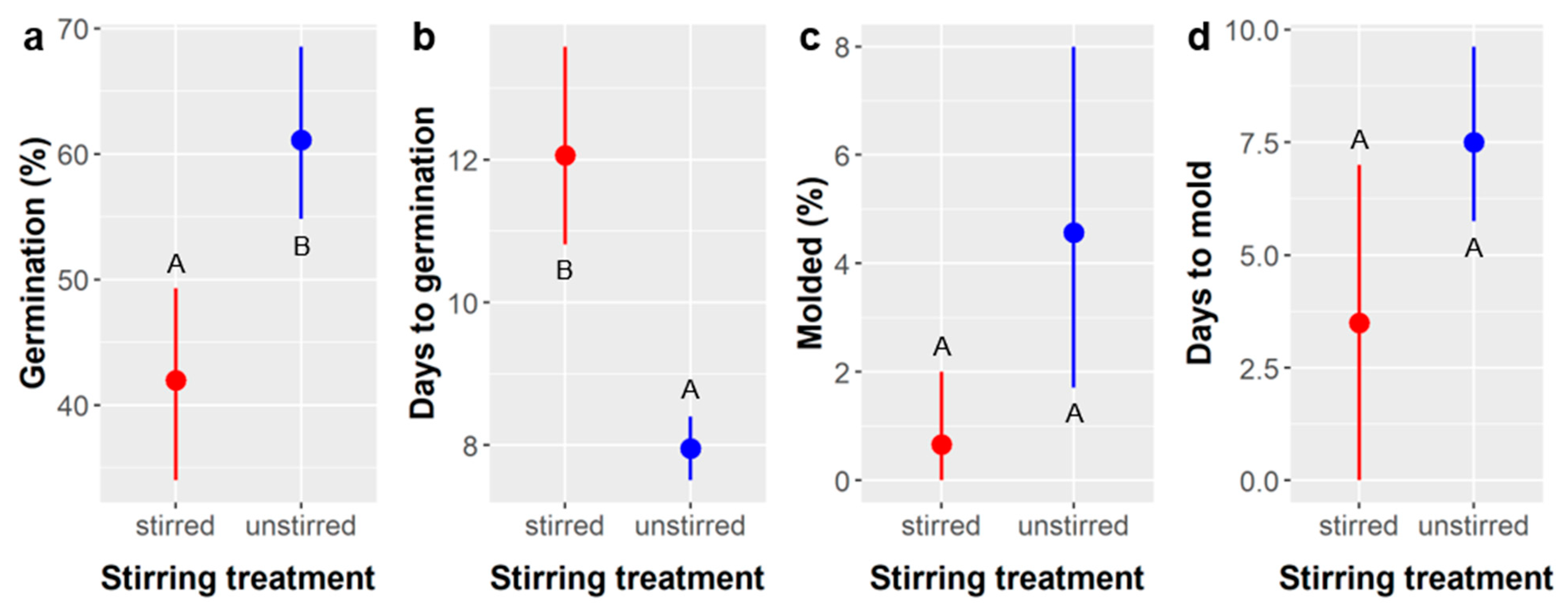
Figure 1). Stirring
Ebenopsis seeds during SA treatment can accelerate the scarification process and may reduce molding, but it is not necessary, and it increases the time required for germination (
Figure 2).
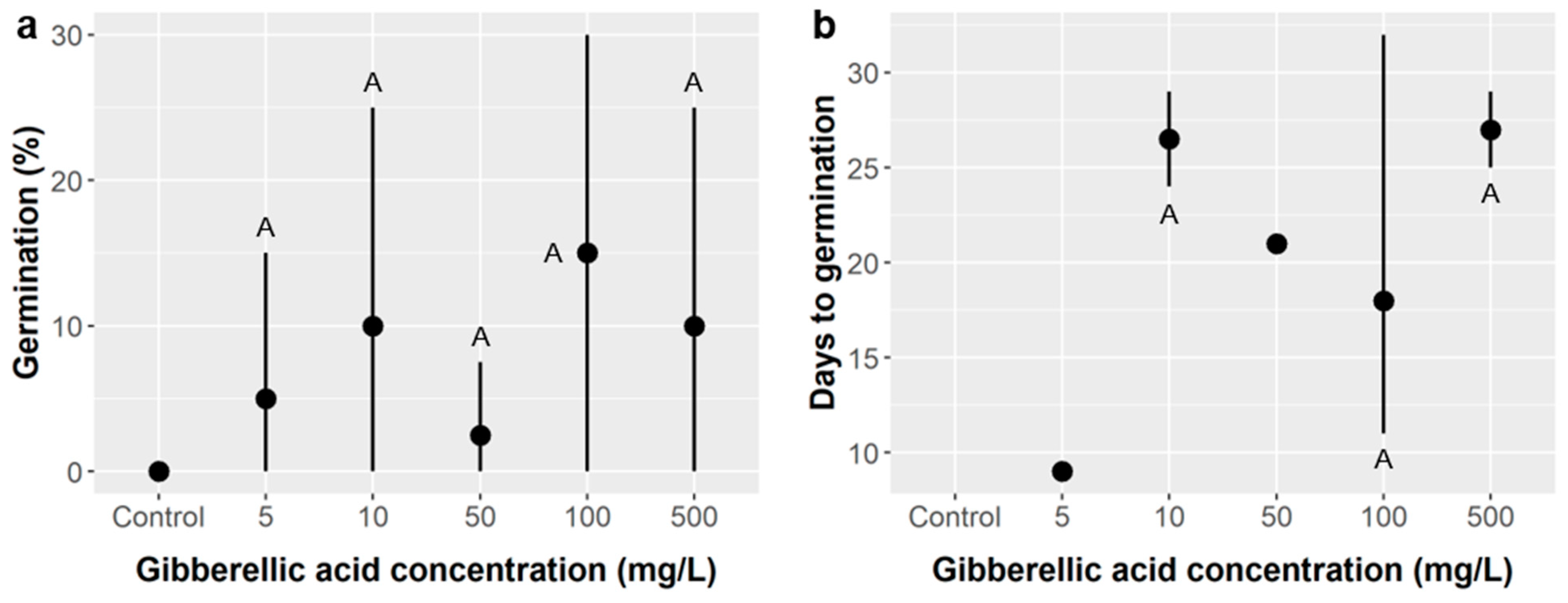
Gibberellic acid (GA) alone did not improve
Ebenopsis germination regardless of the dose tested (from 5–500 mg/L), but, mechanistically, this may be because the thick seed coat of
Ebenopsis prevented GA from reaching the embryo (
Figure 3). Thus, we cannot rule out the possibility that GA treatment could promote
Ebenopsis germination in combination with other seed treatments that increase permeability of the seed coat, such as SA treatment. The need to overcome consecutive dormancy mechanisms in this fashion is well established in the literature [
27,
32]. However, observed germination rates of over 90% in the most favorable SA treatments suggest the effects of GA are likely to be relatively weak compared to those of SA, but
Ebenopsis germination is often lower even with SA treatment [
22,
31], and GA treatment could have a larger effect in these cases. The effects of combining SA and GA treatments merit further investigation. Since the optimal SA soak time depends on seed coat thickness, which varies spatiotemporally, future study is also merited into whether 40–60 min is optimal across years and populations.
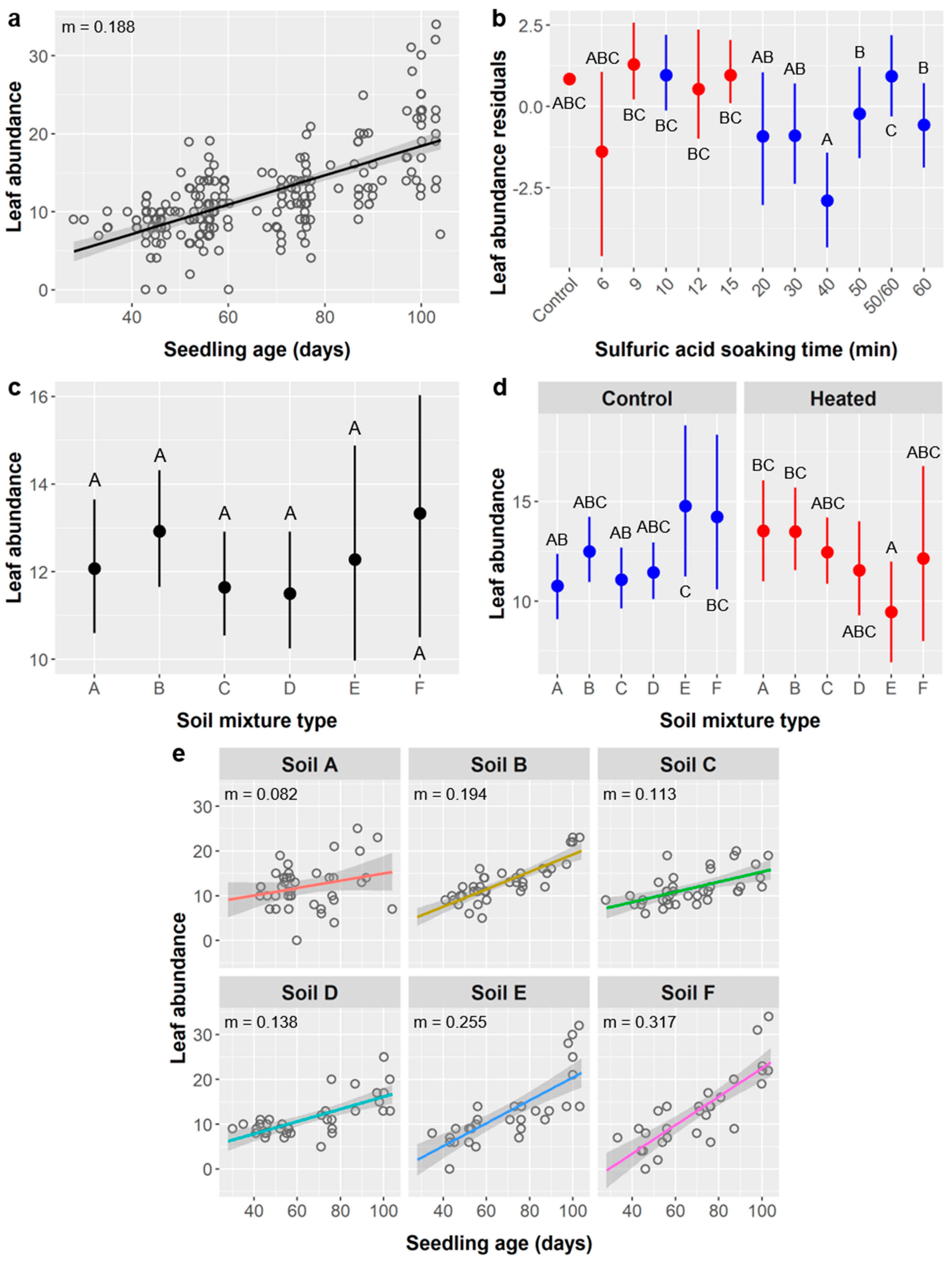
Potential impacts of seed treatments on seedling performance are important to consider, especially if negative effects could negate or exceed a treatment’s positive effects on germination. Importantly, SA treatments had minimal impacts on post-germination performance. SA treatment did not affect seedling survival or height (
Table 4 and
Table 5), and it had a weak effect on leaf abundance (
Table 6,
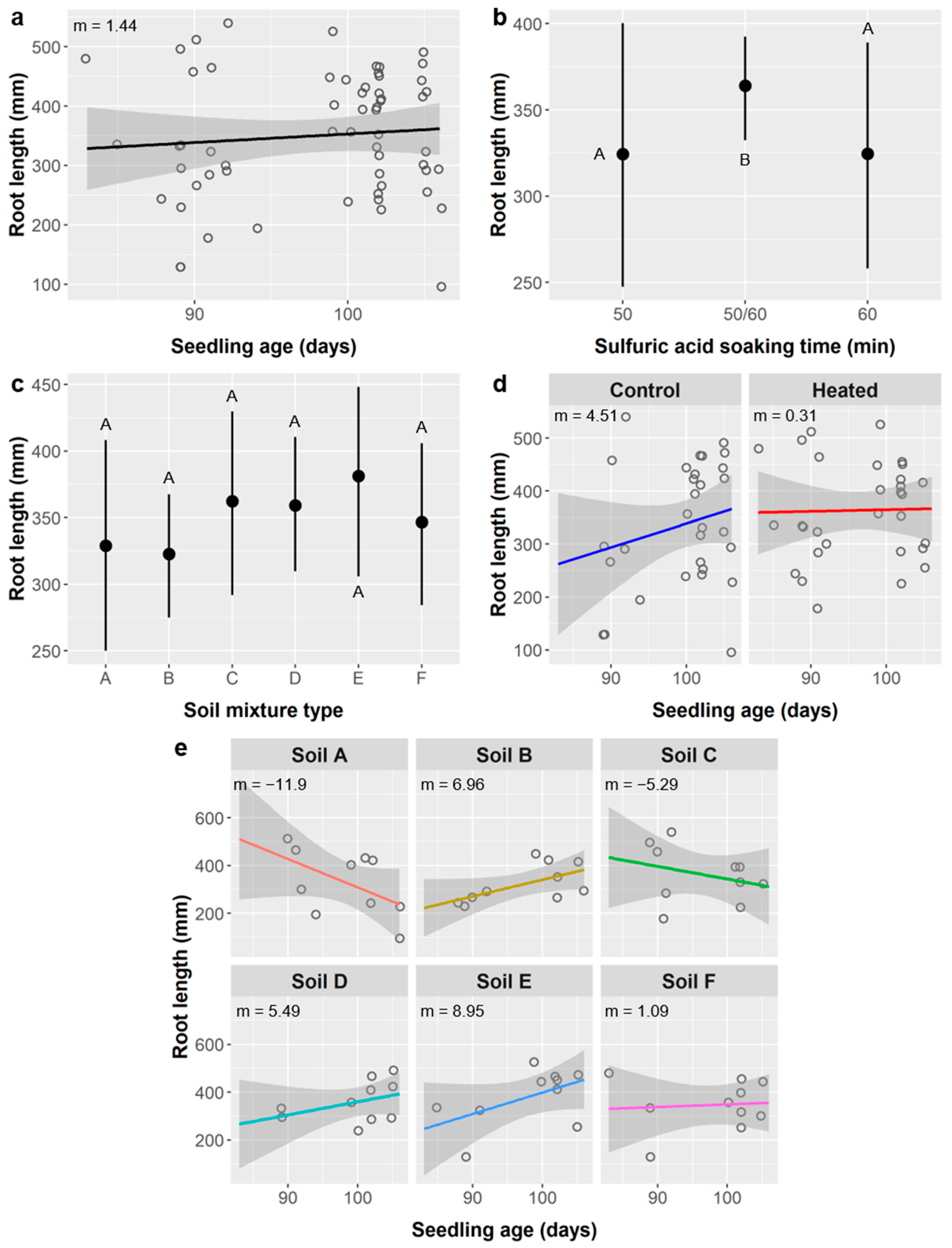
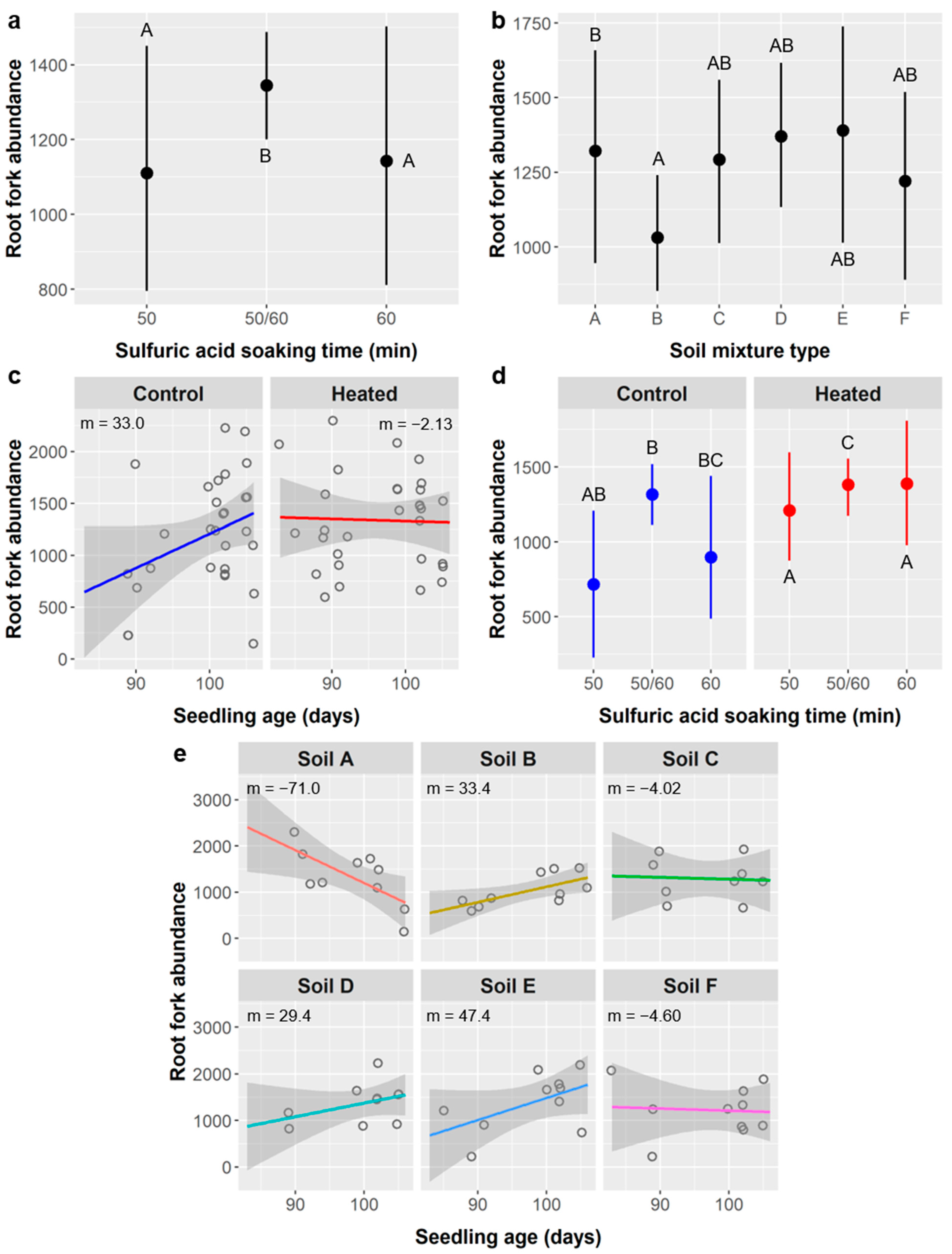
Figure 7b). Several root metrics did depend on SA treatment, including root length (
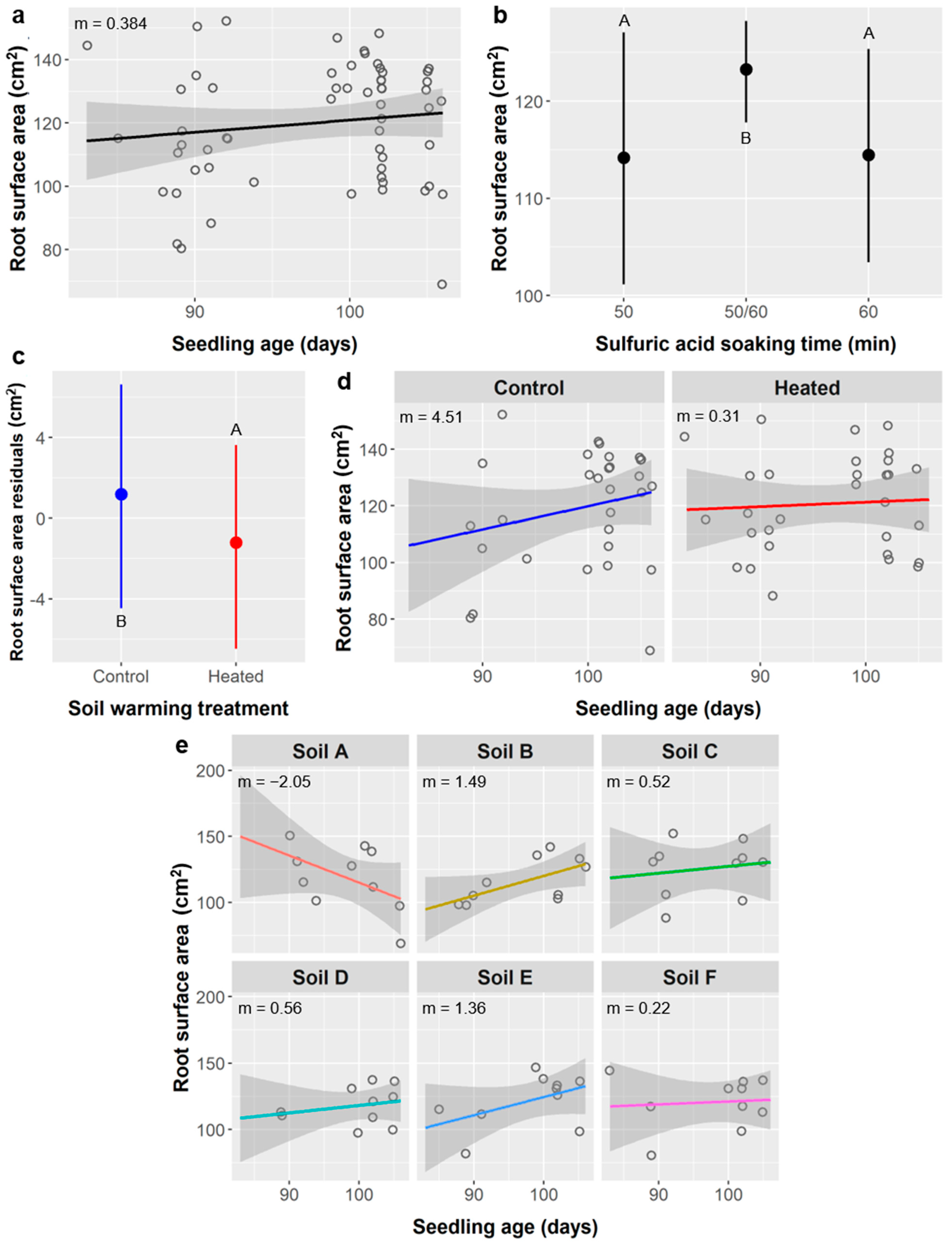
Figure 8b), surface area (
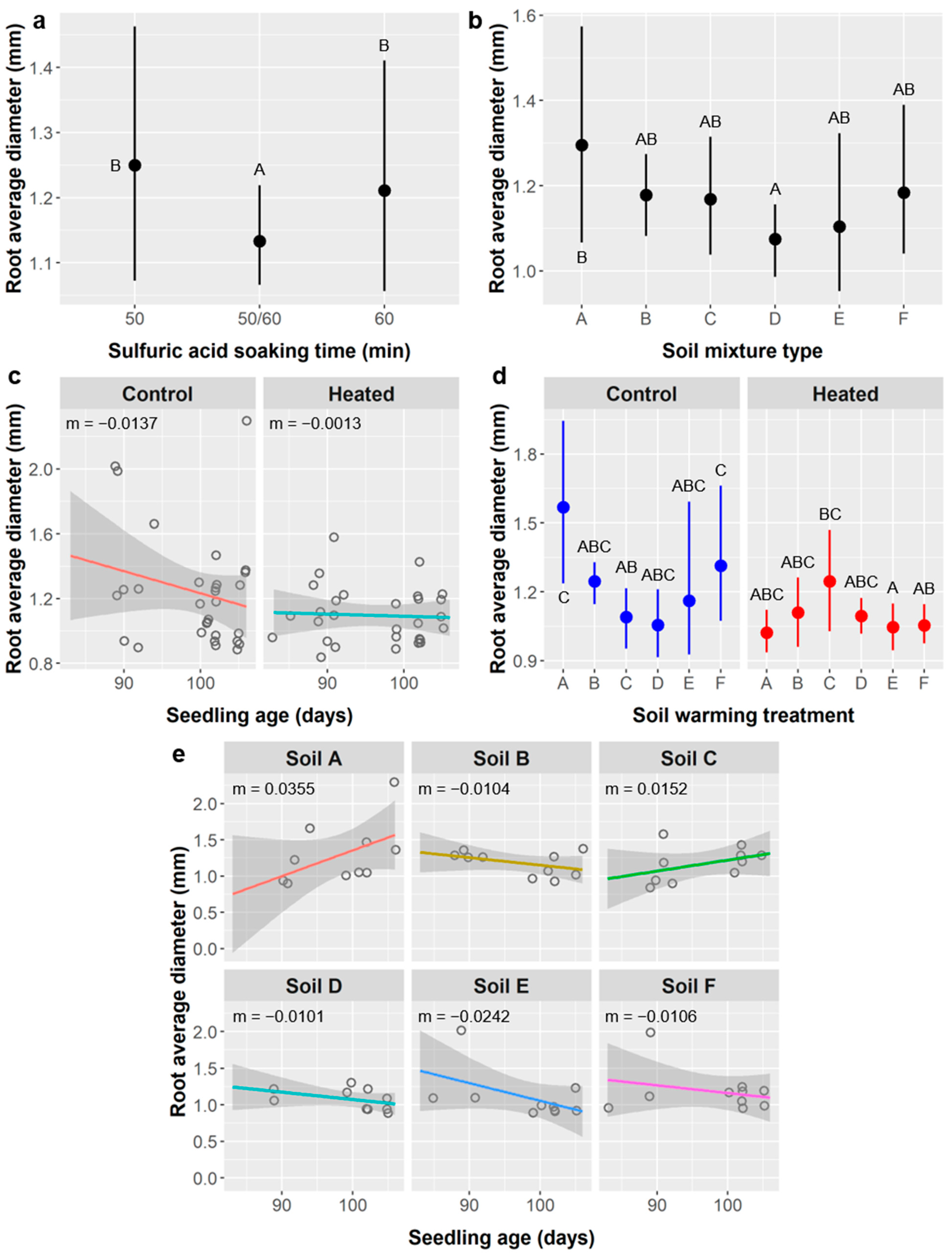
Figure 9b), diameter (
Figure 10a), fork abundance (
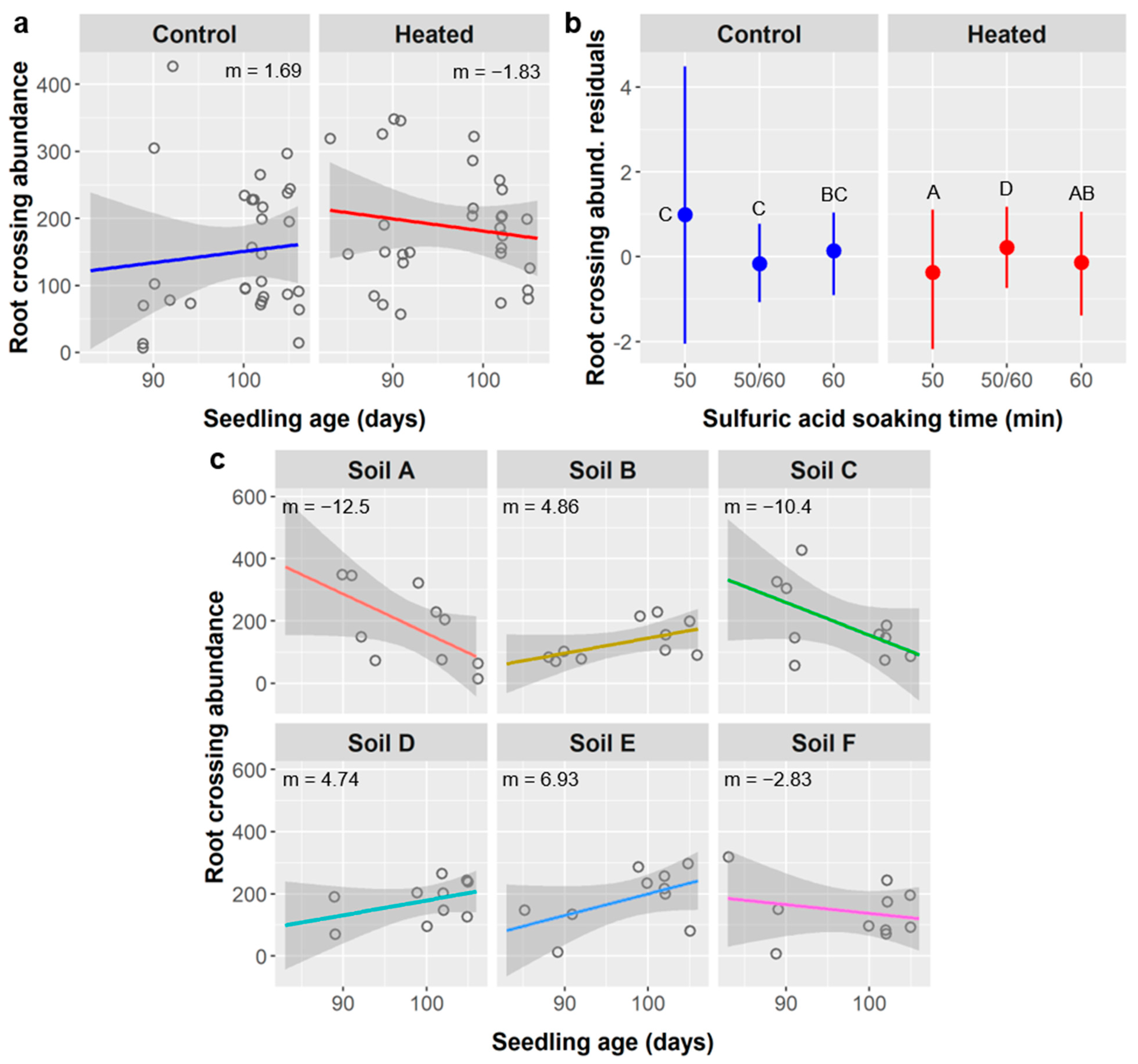
Figure 13d), and crossing abundance (
Figure 14b), all of these except root diameter were higher in the 50/60 SA treatment group. For leaves, after considering the effects of other factors, the shorter-duration stirred SA treatments had about two more leaves than the longer-duration unstirred SA treatments, which is a small but notable difference. Nevertheless, survival was not affected by SA treatment, and these differences are not significant enough to justify avoiding longer SA treatments if they confer ca. 20% higher germination of
Ebenopsis seeds.
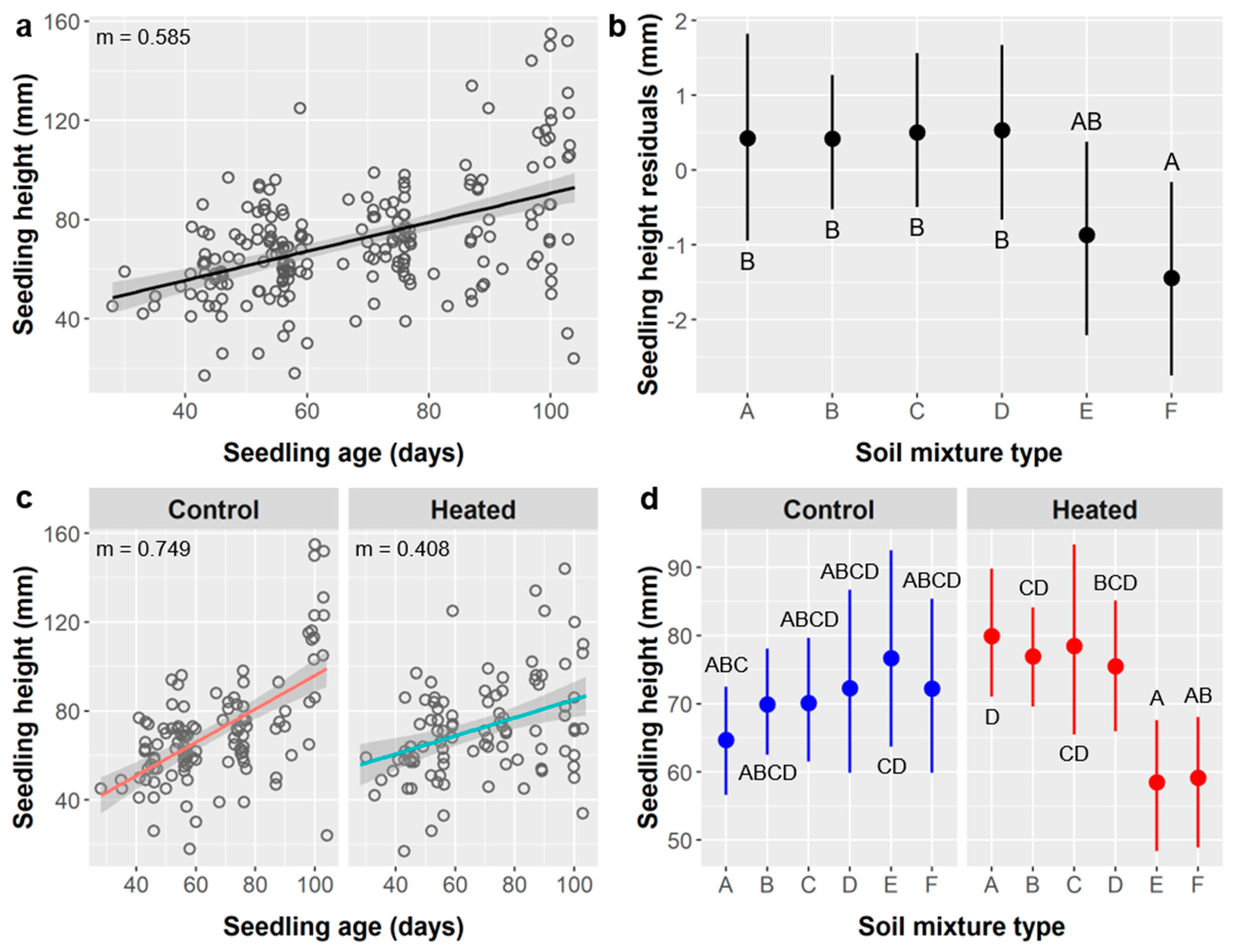
Furthermore, in these cases of leaf abundance and root metrics, the link between SA treatment and seedling performance may actually be driven by the relationship between seedling age and performance. For leaves, the stirred SA treatments and shorter unstirred SA treatments germinated significantly later and thus had younger seedlings at the time of data collection (mean age = 52 days for stirred SA treatments, 42 d for 10 min unstirred, and 74 d for ≥20 min unstirred). For all root metrics, the 50/60 SA group were seedlings from the 50 or 60 min unstirred SA treatments that were planted earlier and whose labels degraded, and were thus ca. 12.5 days older (mean age = 90 d for 50 min SA treatment, 89 d for 60 min SA, and 102 d for the 50/60 SA group).
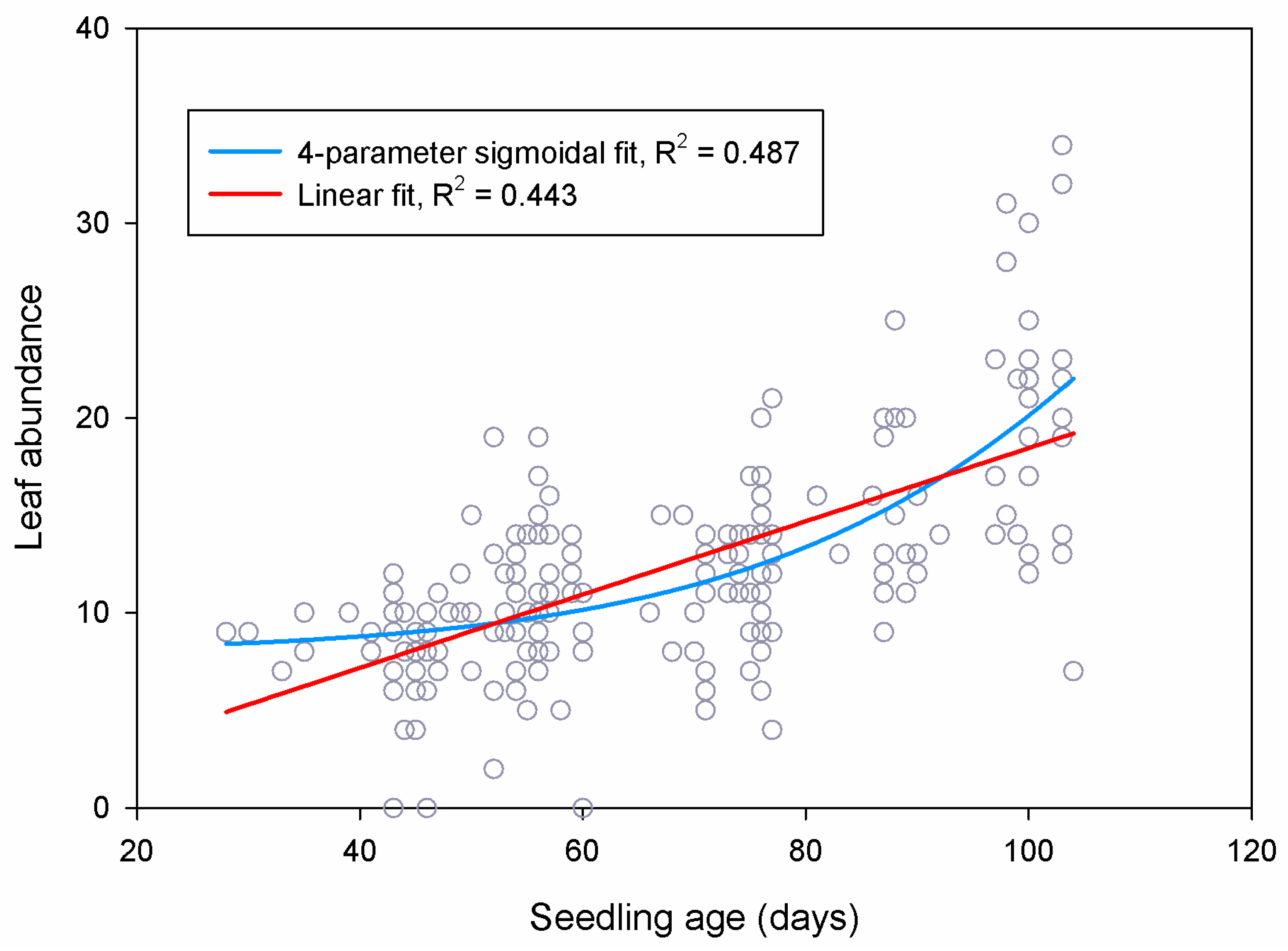
In theory, differences in age among treatment groups should be accounted for statistically by including seedling age in our models, which we did. However, this may fail to explain all the variance driven by seedling age if the relationship between age and performance is more complex than represented by our linear models, i.e., if the relationship is nonlinear. This appears to be the case in our data, and this is supported by prior studies documenting ontogenetic variation in plant growth strategies, with different patterns of carbon allocation to roots, stems, and leaves at different life stages [
57,
58,
59]. We fit a 4-parameter sigmoidal function to the relationship between leaf count and seedling age and performed a nonlinear least-squares regression using SigmaPlot 11 (Systat Software, Inc., Chicago, IL, USA). The sigmoidal relationship was significant (
p < 0.0001) and explained more variability (R
2 = 0.487) than the linear model (R
2 = 0.443) (
Figure 15).
The sigmoidal curve was relatively flat for seedlings aged ca. 20–60 days, which suggests a growth pattern emphasizing early foliation, which is consistent both with our observations and the typical growth strategies of plants like Ebenopsis that have large seeds with nutritional stores capable of fueling rapid growth immediately post-germination. Accelerated foliation after 60 days, as shown, is also consistent with ontogenetic growth strategies because plants become capable of faster absolute growth as they grow and their total photosynthetic capacity increases.
The relationships between SA treatment and root length, surface area, diameter, forks, and crossings are less easily explained as an effect of age because the linear relationships between these root metrics and seedling age are weak (
Figure 8,
Figure 9,
Figure 10,
Figure 13 and
Figure 14). There are three reasons why these relationships are weaker than those between root metrics and the differently-aged SA treatment groups: (1) a much shorter range of ages are represented (83–106 days); (2) there was high variability in root metrics that overshadowed age-related trends; (3) there were significant interactions between age and other factors like soil type and soil warming. Thus, the effects of SA on root metrics are probably more an artefact of our experimental design than they are evidence that SA seed treatments altered
Ebenopsis seedling performance.
Additionally, weakening the relationship between root morphology and age is the observation that, for root length, surface area, forks, and crossings, there was less variance at higher ages (and thus among the 50/60 SA group, specifically), and the peak values for older seedlings were comparable to the peak values among younger seedlings (e.g.,
Figure 8a,b and
Figure 9a,b). Taken together, this is evidence that the older seedlings became space-limited (root-bound) and their root growth and development were likely altered or restricted in some way, which is not surprising for 3-month-old seedlings being grown in ca. 200 mL of soil. Even if age is not the underlying factor driving differences between SA treatments, the effects of SA on root metrics were weak compared to the effects of soil type and soil warming. Thus, for both leaf abundance and root morphology, we found no compelling reasons to limit the use of SA to promote germination of
Ebenopsis seedlings.
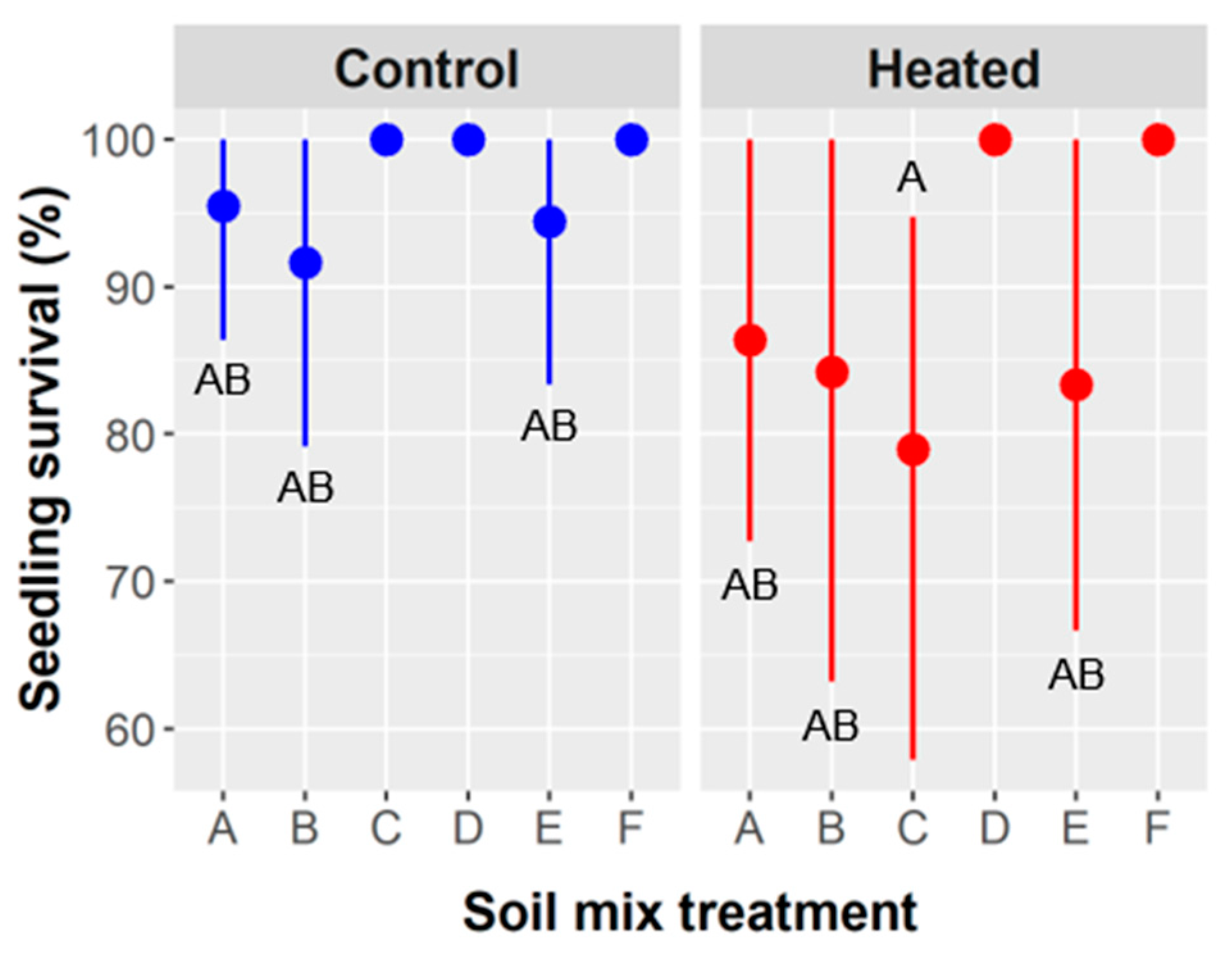
Soil type and soil warming were the only factors to influence survival, and they had the strongest effects on above- and belowground growth. Soil types D (50% peat, 25% sand, 25% vermiculite) and F (50% peat, 20% topsoil, 20% vermiculite, 10% perlite) were the only mixtures to have 100% survival in both soil warming treatments (
Figure 5). This may be because they had the greatest water-holding capacity, but soil F also had the lowest bulk density. Local topsoil is a sandy clay loam that drains quickly but holds more water than sand or perlite [
60], which are used horticulturally to improve drainage and soil aeration. Peat moss and vermiculite hold the most water and are often used for that reason. Soils D and F had combined totals of 75% and 70%, respectively, of peat plus vermiculite, with the next highest mixtures containing 50%, as found in soil types A (50% topsoil, 50% vermiculite), C (50% peat, 25% sand, 25% topsoil), and E (50% peat, 25% perlite, 25% sand). The mixtures with 50% peat + vermiculite had intermediate survival rates, whereas soil type B (50% topsoil, 25% perlite, 25% vermiculite) likely had the lowest water-holding capacity and exhibited the lowest seedling survival (
Figure 5).
Bulk density is another important soil property that influences root growth and varied considerably among our soil mixture treatments. Soil types A, B, D, and E all had bulk densities of approximately 0.690 g/cm
3 (range: 0.675–0.702 g/cm
3; see Methods), whereas soil F was least dense (0.480 g/cm
3), and soil C was most dense (0.892 g/cm
3). In the hot and semi-arid region of south Texas, water availability and water stress are of central importance to plant survival and performance, so it is reasonable that survival was governed by edaphic factors that influenced water availability (e.g., water-holding capacity and evaporation rate) and root development (e.g., bulk density, porosity). It follows mechanistically that survival was lower with soil warming (
Table 5,
Figure 5) because heated treatments would have had a higher evaporation rate and possibly higher transpiration.
The relationships between seedling performance and soil type and warming were more nuanced. Generally, aboveground growth decreased and belowground growth increased when soils were warmed via heating mats, but soil type had idiosyncratic effects on performance and often influenced seedling responses to soil warming. Greater belowground growth at the cost of lower aboveground growth, as observed, is a common response to water stress and is consistent with observed survival patterns. Mechanistically, as water stress increases, most plants will preferentially allocate more carbon to root growth (produce more or larger roots) to increase their capacity to uptake water, and a higher root:shoot ratio can increase tolerance of water stress by reducing transpirational losses in conjunction with greater water absorption capacity [
57]. Many of the specific responses to soil warming described above reflect this general pattern.
Other responses to warming reflect the same stress response. In unheated controls, we observed positive relationships between seedling age and root length, surface area, fork abundance, and crossing abundance, but in the soil warming treatments we saw neutral or weakly negative relationships between seedling age and the same variables (
Figure 8d,
Figure 9d,
Figure 13c and
Figure 14a). One would expect a positive relationship in both treatments and higher values in the warming treatment if warming increased water stress and plants responded by producing more roots, but not if belowground growth was limited by container size and seedlings had become space-limited, as we demonstrated above was likely the case. Importantly, the mean values for these root metrics in warming treatments are at the upper limit of the ranges of values observed in the unheated controls, which suggests seedlings in the control approached the container-imposed upper limit on root size as they reached the upper limits of age, but seedlings in the warming treatment had already reached that container-imposed limit by the lowest ages in the range analyzed. This is further evidence that container size affected seedling growth and root development.
We saw a similar but opposite pattern for root average diameter (
Figure 10c), but this is part of the same water stress response mechanism. Decreased root diameter can be consistent with both increased root growth and the water stress response because only secondary growth increases diameter, and it will not occur unless there is enough fine (low diameter) root mass to provide required water absorption. Most water is absorbed by fine roots with high surface area to volume ratios, so having relatively more fine roots (and thus a lower average root diameter) can improve water absorption capacity.
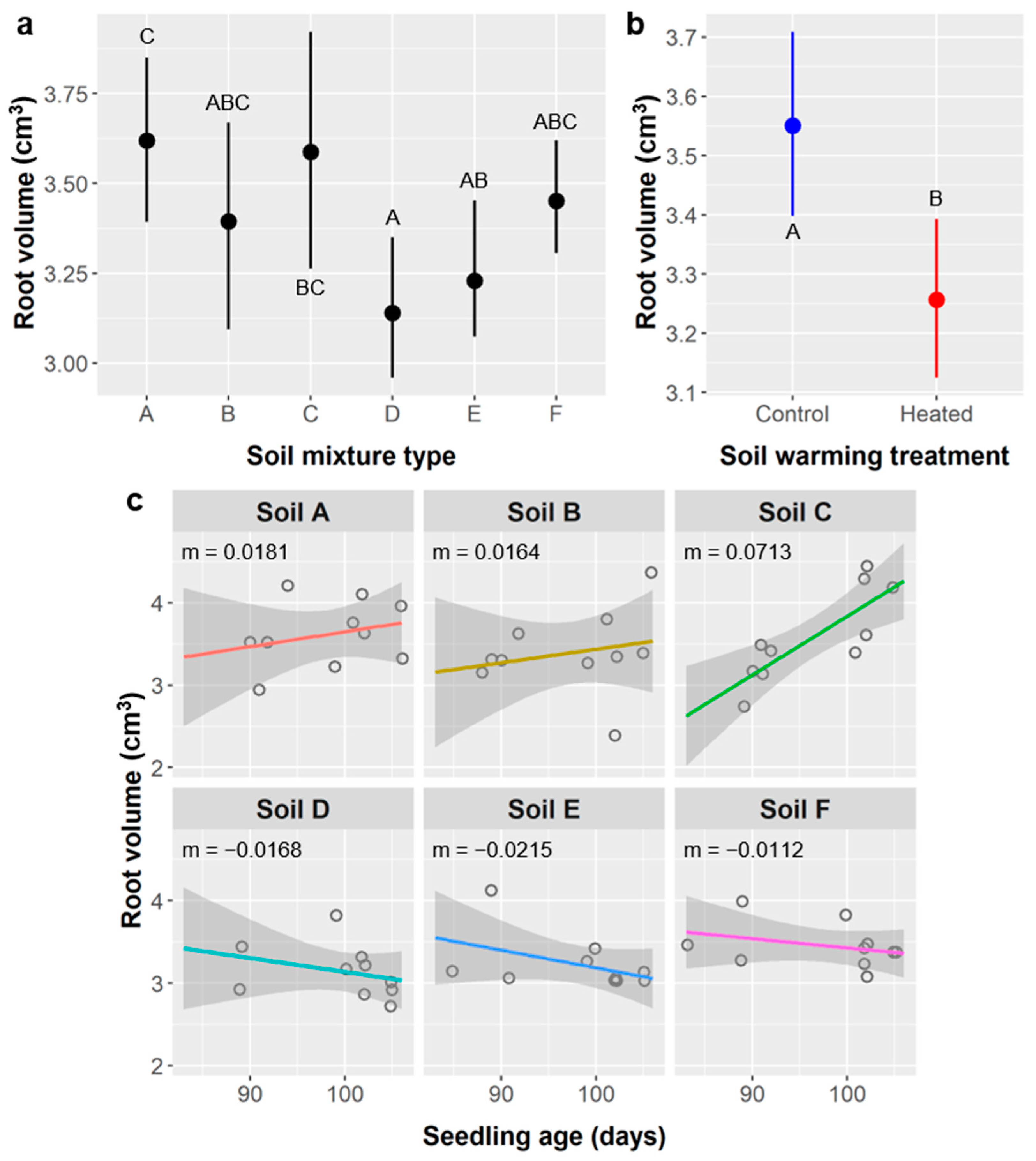
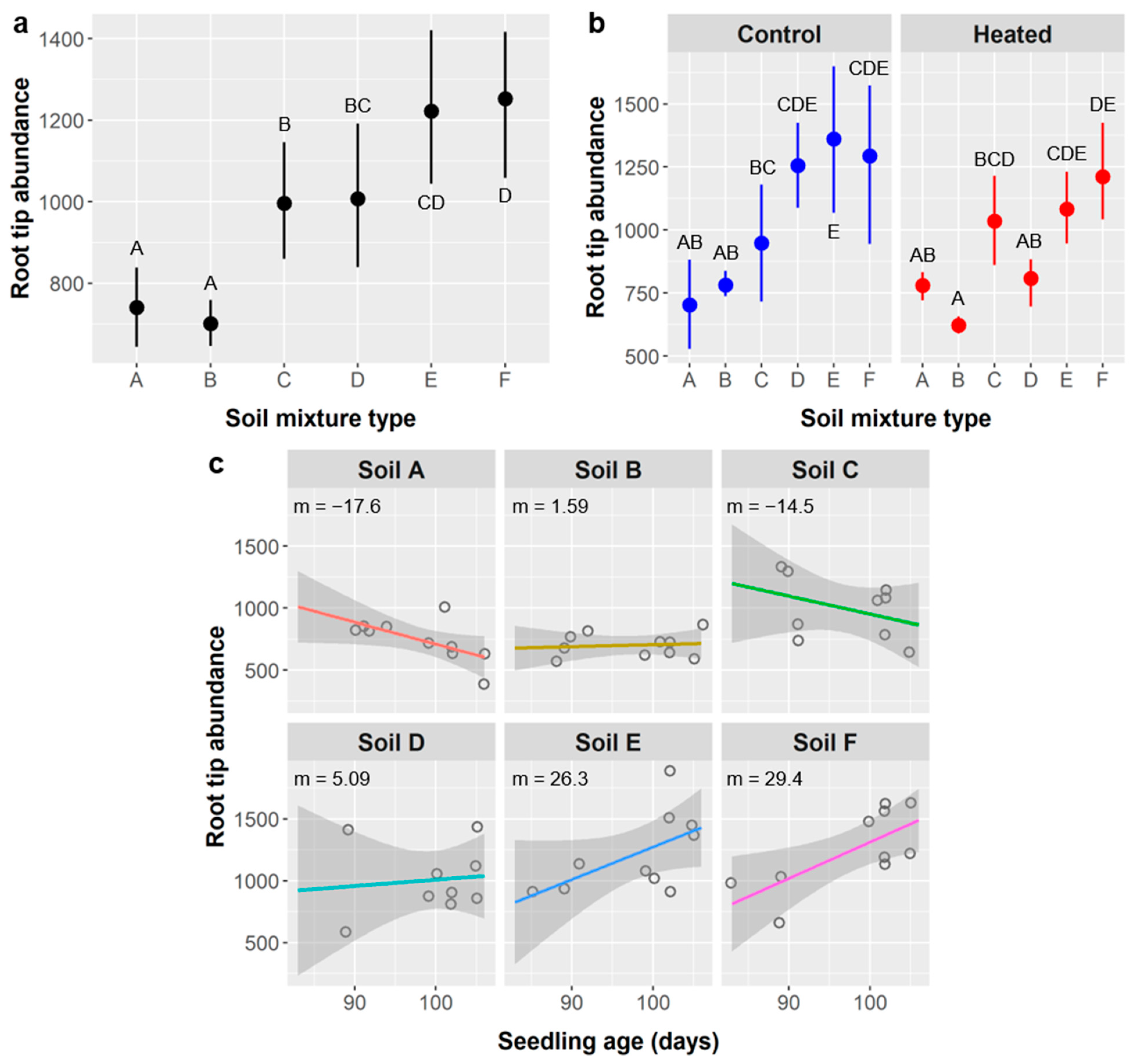
Soil type had more idiosyncratic effects on root metrics and often influenced relationships with age and seedling responses to soil warming. Relative differences in seedling performance attributable to the main effects of soil type are summarized in
Table 14. If we consider marginal effects (0.1 >
p ≥ 0.5), relative aboveground performance was highest in soil B, high in soil A, and mixed or intermediate in soils C, D, E, and F, whereas belowground performance was highest in soils A and C, intermediate in soils D, E, and F, and lowest in soil B. We expected the opposite pattern for seedling performance in soil B since it had the lowest water-holding capacity and seedlings typically allocate more carbon belowground in response to water scarcity. However, mechanistically, if soil B was least suitable for
Ebenopsis root growth, we would expect a “top-heavy” growth pattern and reduced survival, which we observed (survival was lowest in soil B;
Figure 5).
Soil mixtures with more peat moss (types C–F) had mixed results, which is somewhat surprising because high peat content usually promotes root growth horticulturally. Soil C had the highest bulk density, and high bulk density impedes root growth, but even the maximum bulk density of soil mixtures utilized in these experiments (0.892 g/cm
3) was well below the range in which bulk density begins to inhibit root growth [
61]. Soil F had the lowest bulk density, but its belowground performance was intermediate. These findings suggest that the general benefits of reduced bulk density (e.g., increased porosity, aeration, and water holding capacity) were present in all the soil mixture types utilized.
The overall leader in relative performance was soil A, which, notably, is the formulation recommended by USFWS. Soil A was half native topsoil and half vermiculite, which basically serves to improve the water-holding capacity of native soil without altering its chemistry (as peat does). This likely reflects adaptation by native plants to their native soils and may suggest the presence of important beneficial organisms in native soil. Whether the benefits of soil A translate to other Tamaulipan thornscrub species merits investigation.
Table 15 summarizes the nature of the linear relationships between seedling age and
Ebenopsis performance variables for each soil type. The prevalence of negative relationships between age and belowground performance metrics for soil A, and to a lesser degree soil C, suggests root growth became spatially limited most rapidly in these soil types, and may suggest that there was root growth outside of the containers that was not harvestable. Alternatively, but by the same logic, the prevalence of positive relationships between age and belowground metrics for soils B and E suggests roots were slowest to become spatially limited in these soil types, which is consistent with the hypothesis that soil mix B was least conductive for
Ebenopsis root growth.
Finally,
Table 16 summarizes the effects of soil warming on
Ebenopsis performance in each soil type. In soils E and F, warming had a negative effect on most performance metrics, and both contained 50% peat plus a relatively large proportion of drainage promoting materials (i.e., perlite, sand, or vermiculite). Heating likely had a relatively strong effect on increasing evaporation from these soil treatments. This would increase water stress but also promote root growth by triggering the water stress response, and these low density, high porosity soils should provide highly favorable conditions for root growth. The result may be belowground conditions that promote root growth while growth is limited by water availability, which could explain the mixed performance results (
Table 14) and variable relationships with age (
Table 15) observed for soils E and F. Soil warming had few effects in soils B, C, and D.
Interestingly, warming had positive aboveground effects and a strong negative effect on root diameter in soil A. Decreased average root diameter likely reflects a greater abundance of fine root mass in this context, which is supported by
Table 14 and the fact that aboveground performance increased. Mechanistically, higher evaporation with warming could have driven an increase in fine root mass by triggering the water stress response, and it could also reflect greater activity of beneficial soil organisms triggered by warming. Positive effects conferred by beneficial soil organisms might be observed most strongly in soil A because it was the most similar to native soil physically and chemically.
Further study is merited to differentiate between the roles of water stress and water availability in driving these patterns versus the effects of soil bulk density, soil porosity, and other physicochemical attributes like soil pH. The role of soil microorganisms in driving seedling survival and performance is also worth investigating, including whether natural or commercial soil inoculants can boost performance.
These germination and rearing practices are not confined to Ebenopsis. There are numerous thornscrub species in the Fabaceae with seeds and growth habits comparable to Ebenopsis, including four Acacia species. Some of these other legumes have exhibited low germination rates and substantially slower growth than Ebenopsis. Thus, these insights into Ebenopsis’s seed treatment and rearing methods can provide a useful foundation for future investigations into the propagation of other thornforest species.
3.2. Cordia boissieri (Mexican olive)
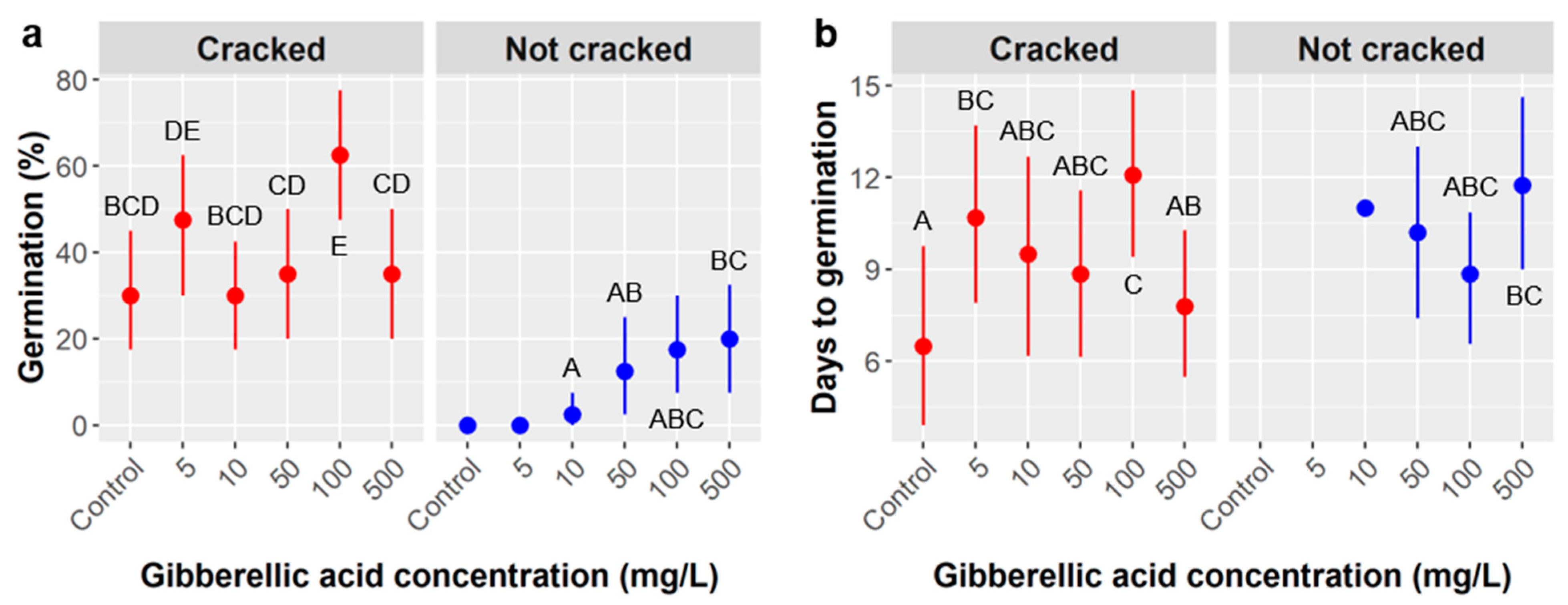
For
Cordia, we posit that treating seeds with 100 mg/L GA is optimal because increasing germination likelihood is most important, and this dose offers a large increase in germination from ca. 35% to over 60% (among cracked seeds) while imposing only a modest delay in germination timing. In practice, if germination takes 12 days instead of 9, as our data suggest for this dosage, this would impose only a minor inconvenience and is much less important than nearly doubling the likelihood of germination. Furthermore, the observed differences in germination timing were not statistically significant according to our ANOVA (
Table 3), which further supports the notion that differences in germination likelihood are paramount for
Cordia.
Like
Ebenopsis,
Cordia seeds have a hard exterior, but instead of being a thickened seed coat, it is the hardened endocarp or pit of the fruit, which is a drupe like a peach or cherry. Physical dormancy due to an impermeable seed coat has not been recognized in
Cordia’s family, the Boraginaceae, nor is it one of the forms of dormancy employed by the most familiar drupes of the Rosaceae [
25]. Nevertheless, our data suggests that physically breaking down the endocarp of
Cordia may enhance the likelihood of germination (
Figure 4). We cannot be sure, however, that the observed increase in germination was entirely due to the physical cracking of the endocarps achieved by desiccating the outer layer because this ‘heat cracking’ treatment was produced by accident on all the seeds from one population of
Cordia trees. It is possible the differences in germination between cracked and not cracked treatments are due to differences in viability between the populations tested. Unfortunately, seeds were highly limited, so we could not heat crack more seeds from additional populations to eliminate this confounding factor. The effects of heat cracking
Cordia seeds merits further study.
Furthermore, this uncertainty could have been partially, if not entirely, resolved by estimating the viability of all populations through tetrazolium testing or a similar approach. Tetrazolium testing in its own right is worthwhile in studies of thornscrub species because data on their seed viability is exceedingly limited, and this makes it currently impossible to link seed viability with other factors [
22]. For these reasons, among others, we argue that seed viability of thornscrub species should always be tested (e.g., via tetrazolium), especially alongside experimental factors in future studies of germination.
Among uncracked seeds, GA concentrations over 100 mg/L further increased Cordia germination, but the GA × cracking interaction raises mechanistic uncertainties. In the uncracked treatment, the increase in germination is neatly proportional to GA concentration, but, in the cracked treatment, there is no such proportional relationship. If the viability of seeds was similar among cracked and not cracked populations, the germination patterns observed would suggest that germination was largely driven by imbibition of water rather than GA dosage. In this scenario, if a limited amount of water penetrated Cordia seeds in the uncracked treatment, the GA dose may matter in those cases and could have produced the results observed. Alternatively, if cracked seeds had higher viability, that alone could explain observed differences in germination between the two groups, but it would not explain why germination was proportional to GA concentration in the uncracked seeds but not the cracked seeds. Instead, this might suggest that Cordia populations differ in both base viability and in their responses to GA.
In practice, if observed patterns hold true, GA treatment should be a useful means to enhance Cordia germination, and the heat cracking technique we discovered could prove exceptionally useful and cost-effective at both small and large scales of production.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}