
Climatic Diversity and Ecological Descriptors of Wild Tomato Species (Solanum sect. Lycopersicon) and Close Related Species (Solanum sect. Juglandifolia y sect. Lycopersicoides) in Latin America

 , , , and
, , , and 
Abstract
1. Introduction
2. Results
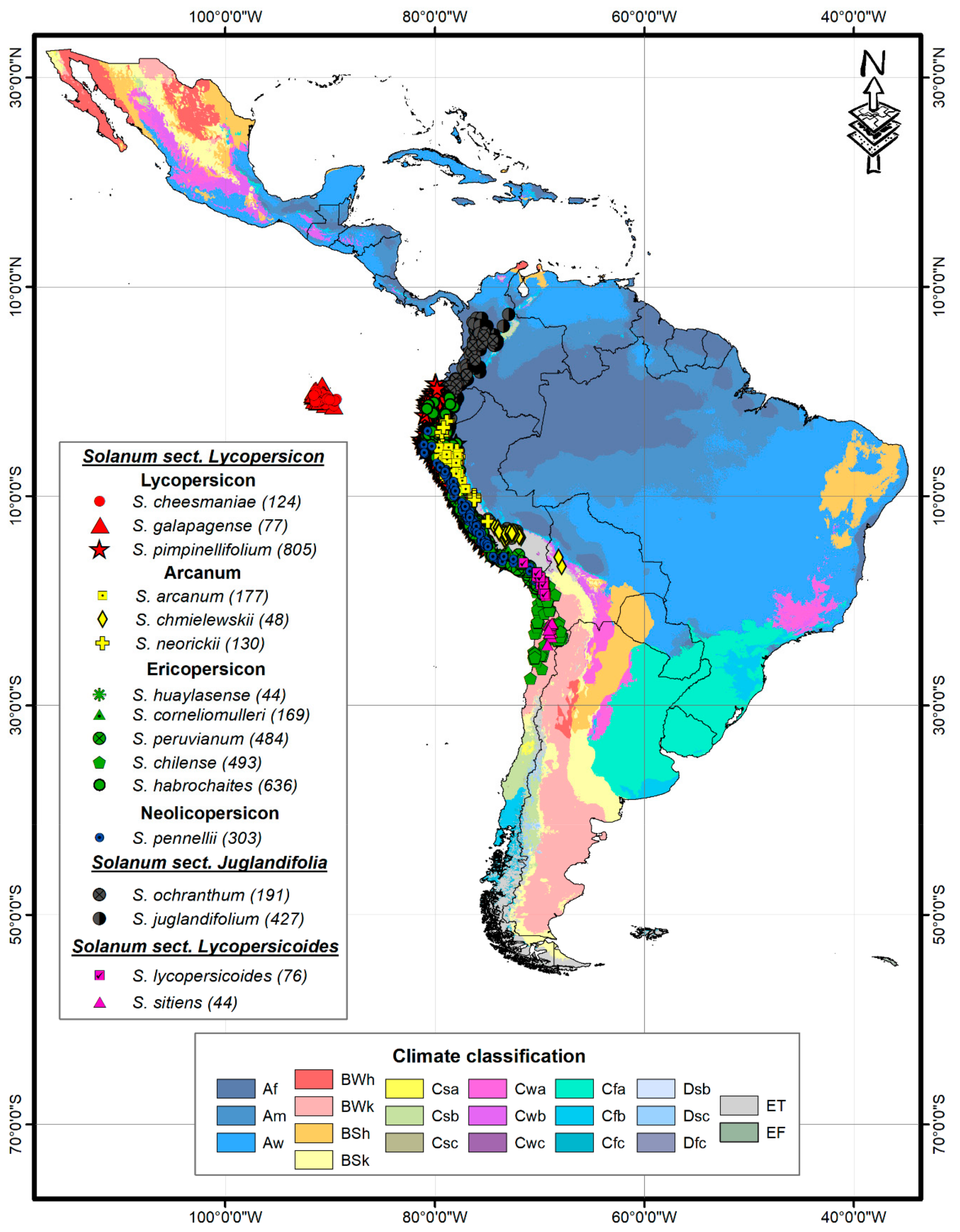
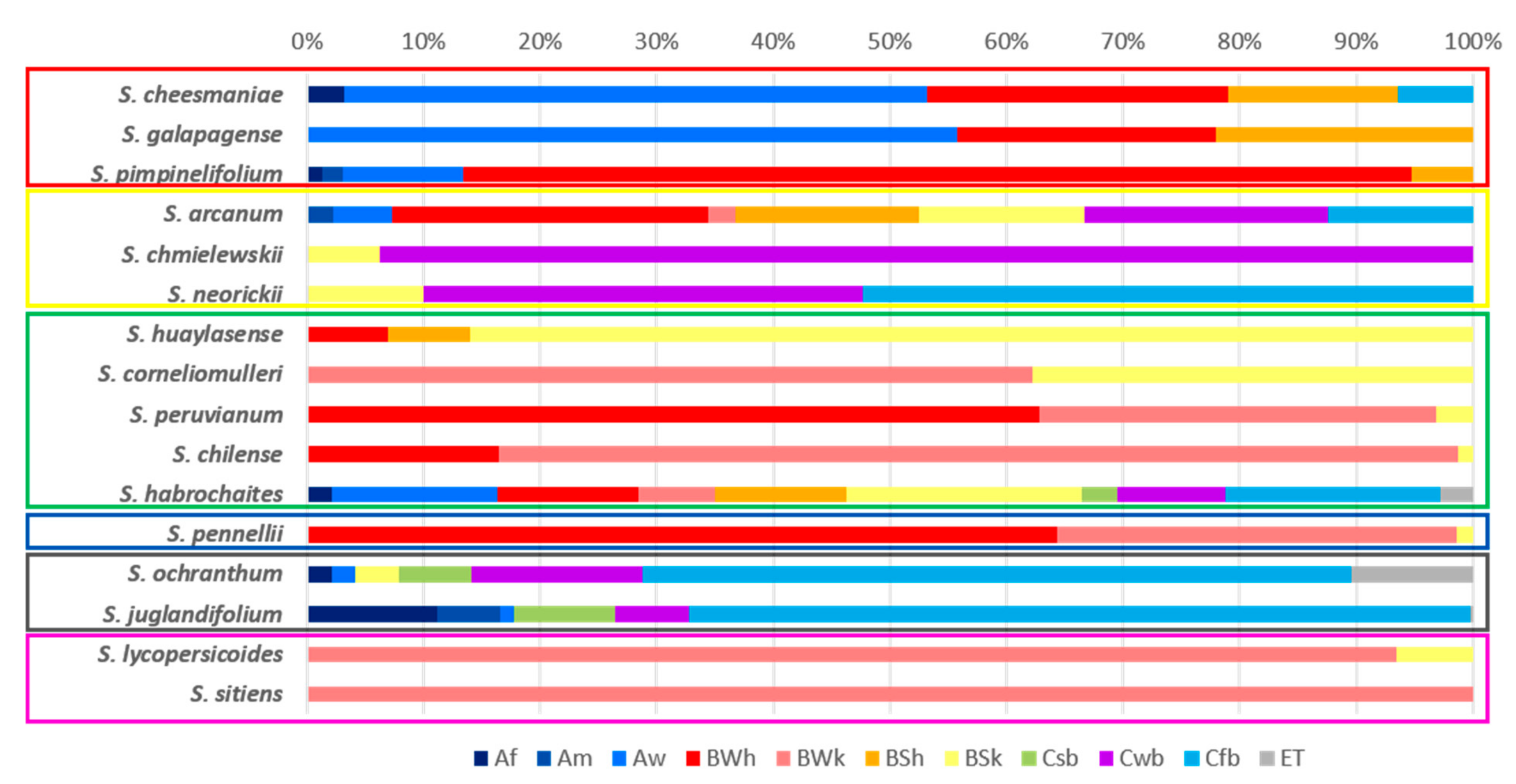
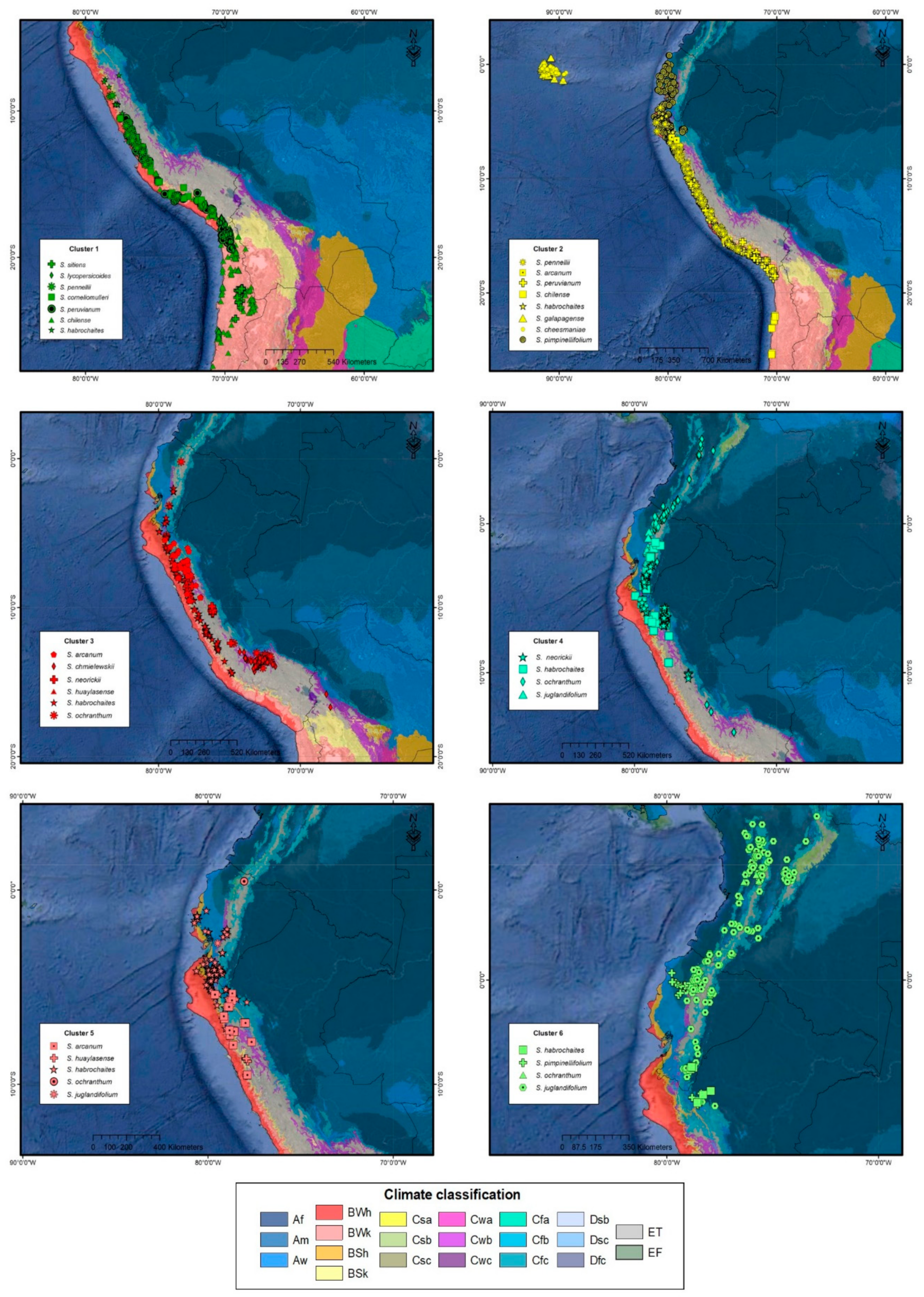
2.1. Climatic Diversity
2.2. Ecological Descriptors
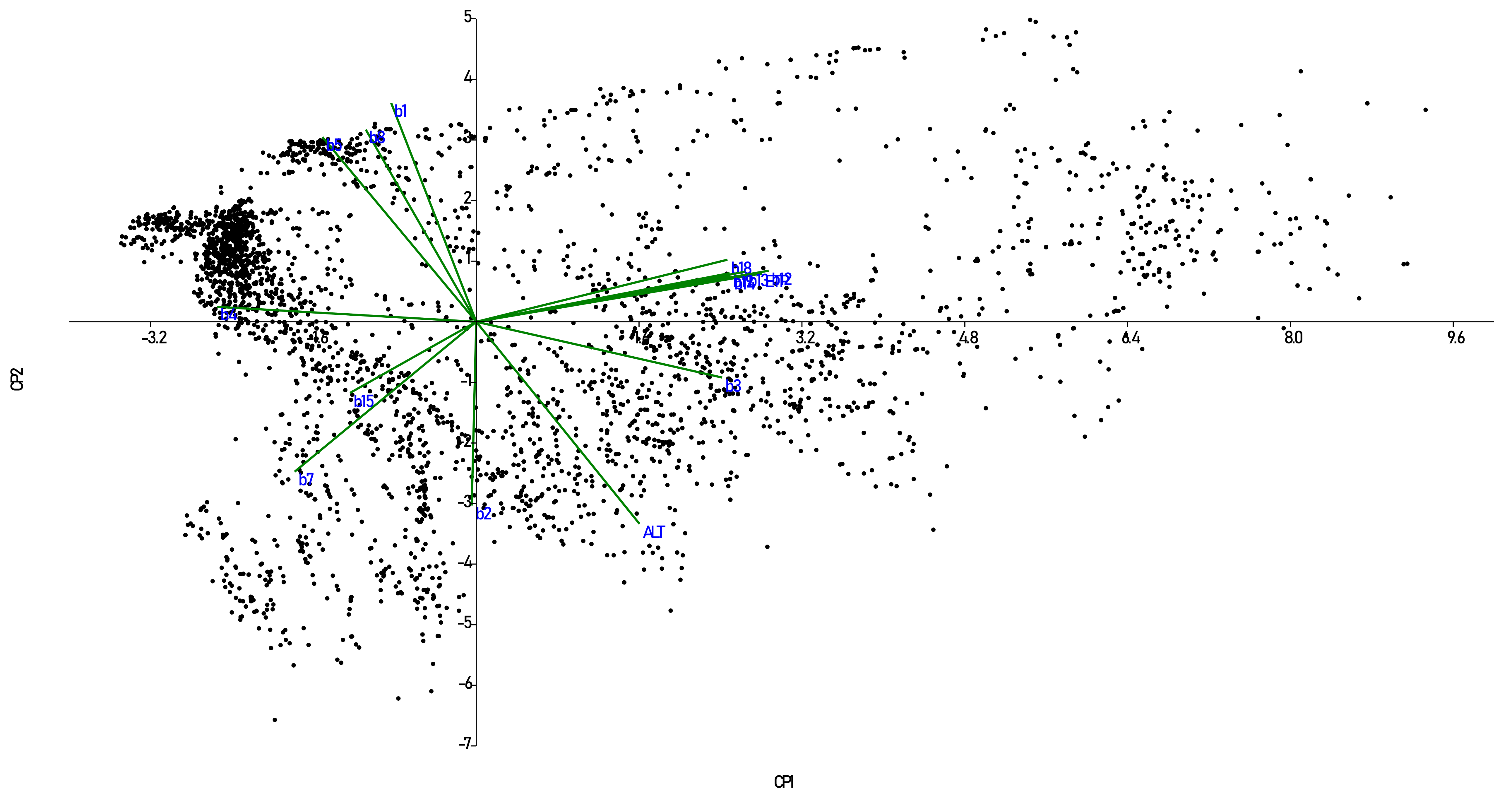
2.3. Statistical Analysis
3. Discussion
4. Materials and Methods
4.1. Database
4.2. Environmental Information
4.3. Climatic Diversity and Ecological Descriptors
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peralta, I.E.; Spooner, D.M.; Knapp, S. Taxonomy of wild tomatoes and their relatives (Solanum sect. Lycopersicoides, sect. Juglandifolia, sect. Lycopersicon; Solanaceae). Syst. Bot. Monogr. 2008, 84, 1–186. [Google Scholar]
- Razifard, H.; Ramos, A.; Valle, A.L.D.; Bodary, C.; Goetz, E.; Manser, E.J.; Li, X.; Zhang, L.; Visa, S.; Tieman, D.; et al. Evidence for Complex Domestication History of the Cultivated Tomato in Latin America. Mol. Biol. Evol. 2020, 37, 1118–1132. [Google Scholar] [CrossRef] [PubMed]
- Aflitos, S.; Schijlen, E.; de Jong, H.; de Ridder, D.; Smit, S.; Finkers, R.; Wang, J.; Zhang, G.; Li, N.; Mao, L.; et al. Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plants 2014, 80, 136–148. [Google Scholar]
- Causse, M.; Giovannoni, J.; Bouzayen, M.; Zouine, M. (Eds.) The Tomato Genome; Springer: Berlin, Germany, 2016; 259p. [Google Scholar] [CrossRef]
- Tropicos.org. Available online: https://tropicos.org/home (accessed on 20 December 2020).
- Rodríguez, F.; Feinan, W.; Cécile, A.; Tanksley, S.; Spooner, D.M. Do potatoes and tomatoes have a single evolutionary history, and what proportion of the genome supports this history? BMC Evol. Biol. 2009, 191. [Google Scholar] [CrossRef] [PubMed]
- Pease, J.B.; Haak, D.C.; Hahn, M.W.; Moyle, L.C. Phylogenomics Reveals Three Sources of Adaptive Variation during a Rapid Radiation. PLoS Biol. 2016, 14, e1002379. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Guzmán, E.; Vargas-Canela, D.; de Sánchez-González, J.J.; Lépiz-Idelfonso, R.; Rodríguez-Contreras, A.; Ruiz-Corral, J.A.; Puente-Ovalle, P.; Miranda-Medrano, R. Etnobotánica de Solanum var cerasiforme en el occidente de México. Nat. Desarro. 2009, 7, 45–57. [Google Scholar]
- Chávez-Servia, J.L.; Carrillo-Rodríguez, J.C.; Vera, A.; Rodríguez-Guzmán, E.; Lobato-Ortíz, R. Utilización Actual del Jitomate Silvestre Mexicano; Subsistema Nacional de Recursos Fitogenéticos para la Alimentación y la Agricultura, Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación, CIIDIR-Unidad Oaxaca del Instituto Politécnico Nacional, ITVO-Oaxaca: Oaxaca, México, 2009; 72p. [Google Scholar]
- Carrillo-Rodríguez, J.C.; Chávez-Servia, J.L. Caracterización agromorfológica de muestras de tomate de Oaxaca. Rev. Fitotec. Mex. 2010, 33, 1–6. [Google Scholar]
- Ramírez-Ojeda, G.; Ruiz-Corral, J.A.; Pérez-Mendoza, C.; Villavicencio-García, R.; Mena-Munguía, S.; Durán-Puga, N. Impactos del cambio climático en la distribución geográfica de Gossypium hirsutum L. en México. Rev. Mex. Cienc. Agríc. 2014, 5, 1885–1895. [Google Scholar] [CrossRef][Green Version]
- Ruiz-Corral, J.A.; Durán-Puga, N.; Sánchez-González, J.J.; Ron-Parra, J.; González-Eguiarte, D.R.; Holland, J.B.; Medina-García, G. Climatic adaptation and ecological descriptors of 42 Mexican Maize (Zea mays L.) races. Crop Sci. 2008, 48, 1502–1512. [Google Scholar] [CrossRef]
- Ruiz-Corral, J.A.; Sánchez-González, J.J.; Hernández-Casillas, J.M.; Willcox, M.C.; Ramírez-Ojeda, G.; Ramírez-Díaz, J.L.; González-Eguiarte, D.R. Identificación de razas de maíz adaptadas a condiciones deficientes de humedad mediante datos biogeográficos. Rev. Mex. Cienc. Agric. 2013, 4, 829–842. [Google Scholar] [CrossRef][Green Version]
- De la Paz-Gutierréz, S.; Sánchez-González, J.J.; Ruiz-Corral, J.A.; Ron-Parra, J.; Miranda-Medrano, R.; De la Cruz-Larios, L.; Lépiz-Idelfonso, R. Diversidad de especies insectiles en maíz y teocintle en México. Folia Entomo. Mex. 2010, 48, 103–118. [Google Scholar]
- Cervantes-Moreno, R.; Rodríguez-Pérez, J.E.; Carrillo-Fonseca, C.; Sahagún-Castellanos, J.; Rodríguez-Guzmán, E. Tolerancia de 26 colectas de tomates nativos de México al nematodo Meloidogyne incognita (Kofoid y White) Chitwood. Rev. Chapingo Ser. Hortic. 2014, 20, 5–18. [Google Scholar] [CrossRef]
- Marín-Montes, I.M.; Lobato-Ortiz, R.; Carrillo-Castañeda, G.; Rodríguez-Pérez, J.E.; García-Zavala, J.J.; Velasco-García, A.M. Riqueza alélica de poblaciones nativas de jitomate (Solanum lucopersicum L.) para el mejoramiento genético. Agrociencia 2019, 53, 355–370. [Google Scholar]
- Magallanes-López, A.M.; Martínez-Damián, M.T.; Sahagún-Castellanos, J.; Pérez-Flores, L.J.; Marín-Montes, I.M.; Rodríguez-Pérez, J.E. Post-harvest quality of 40 collections of tomato (Solanum lycopersicum L.) native of Mexico. Agrociencia 2020, 54, 779–795. [Google Scholar] [CrossRef]
- Córdova-Téllez, L.; Molina-Moreno, J.C. Conservación ex situ. In Recursos Fitogenéticos de México para la Alimentación y la Agricultura; Informe Nacional sobre el estado de los recursos fitogenéticos para la agricultura y la alimentación; Moreno, J.C.M., Téllez, L.C., Eds.; Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación y Sociedad Mexicana de Fitogenética, A.C: Chapingo, México, 2006; 172p. [Google Scholar]
- Florido, M.; Plana, D.; Álvarez, M.; Moya, C.; Regla, M.; Caballero, A. Evaluación del germoplasma de tomate (Solanum L. sección Lycopersicon) conservado Ex situ en Cuba utilizando prueba In vitro para predecir las respuestas a altas temperaturas. Cultiv. Trop. 2009, 30, 57–61. [Google Scholar]
- Juárez-López, P.; Castro-Brindis, R.; Colinas-León, T.; Ramírez-Vallejo, P.; Sandoval-Villa, M.; Reed, D.W.; Cisneros-Zeballos, L.; King, S. Evaluación de calidad de frutos de siete genotipos nativos de jitomate (Lycopersicum esculentum var. cerasiforme). Rev. Chapingo Ser. Hortic. 2009, 15, 5–9. [Google Scholar] [CrossRef]
- TGRC (Tomato Genetics Resource Center). Available online: https://tgrc.ucdavis.edu/ (accessed on 19 December 2020).
- Nakazato, T.; Warren, D.L.; Moyle, L.C. Ecological and geographic modes of species divergence in wild tomatoes. Am. J. Bot. 2010, 4, 680–693. [Google Scholar] [CrossRef]
- Grandillo, S.; Chetelat, R.; Knapp, S.; Spooner, D.; Peralta, I.E.; Cammareri, M.; Pérez, O.; Termolino, P.; Tripodi, P.; Chiusano, M.L.; et al. Solanum sect. Lycopersicon. In Wild Crop Relatives: Genomic and Breeding Resources Vegetables; Chittaranjan, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; Chapter 9; pp. 129–215. [Google Scholar] [CrossRef]
- Parra-Quinajo, M.; Iriondo, J.M.; Torres, E. Review. Applications of ecogeography and geographic information systems in conservation and utilization of plant genetic resources. Span. J. Agric. Res. 2012, 10, 419–429. [Google Scholar] [CrossRef]
- Hijmans, J.R.; Spooner, D.M. Geographic distribution of wild potato species. Am. J. Bot. 2001, 88, 2101–2112. [Google Scholar] [CrossRef]
- López-Soto, J.L.; Ruiz-Corral, J.A.; Sánchez-González, J.J.; Lépiz-Idelfonso, R. Adaptación climática de 25 especies de frijol silvestre (Phaseolus spp.) en la República Mexicana. Rev. Fitotec. Mex. 2005, 28, 221–230. [Google Scholar]
- Ruiz-Corral, J.A.; Sánchez-González, J.J.; Aguilar, M. Potential geographical distribution of teosinte in México: A GIS approach. Maydica 2001, 46, 105–110. [Google Scholar]
- Sánchez-González, J.J.; Ruiz-Corral, J.A.; Medina-García, G.; Ramírez-Ojeda, G.; de la Cruz-Larios, L.; Holland, J.B.; Miranda-Medrano, R.; García-Romero, G.E. Ecogeography of teosinte. PLoS ONE 2018, 13, e0192676. [Google Scholar] [CrossRef]
- Steiner, J.J.; Greene, S.L. Proposed ecological descriptors and their utility for plant germplasm collections. Crop Sci. 1996, 36, 439–451. [Google Scholar] [CrossRef]
- Lobo-Burle, M.; Torres-Cordeiro, C.M.; Fonseca, J.R.; de Melo, L.A.M.P.; de Belem-das, R.N.A.; Abadie, T. Characterization of germplasm according to environmental conditions at the collecting site using GIS: Two case studies from Brazil. Plant Genet. Resour. Newsl. 2013, 135, 1–11. [Google Scholar]
- Beck, H.E.; Zimmermann, N.E.; Mcvicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef]
- Nakazato, T.; Housworth, E.A. Spatial genetics of wild tomato species reveals roles of the Andean geography on demographic history. Am. J. Bot. 2011, 98, 88–98. [Google Scholar] [CrossRef]
- Bergougnoux, V. The history of tomato: From domestication to biopharming. Biotechnol. Adv. 2013, 32, 170–189. [Google Scholar] [CrossRef]
- Rick, C.M. Biosystematic studies in Lycopersicon and closely related species of Solanum. In The Biology and Taxonomy of Solanaceae; Hawkes, J.G., Lester, R.N., Skelding, A.D., Eds.; Academic Press: New York, NY, USA, 1979; pp. 667–677. [Google Scholar]
- Taylor, I.B. Biosystematic of the tomato. In The Tomato Crop: A Scientific Basis for Improvement; Atherton, I.G., Rudich, I., Eds.; Chapman and Hall: London, UK, 1986; pp. 1–34. [Google Scholar] [CrossRef]
- Flores-Hernández, L.A.; Lobato-Ortiz, R.; García-Zavala, J.J.; Molina-Galán, J.D.; Sangerman-Jarquín, D.M.; Velasco-Alvarado, M.J. Parientes silvestres del tomate como fuente de germoplasma para el mejoramiento genético de la especie. Rev. Fitotec. Mex. 2017, 40, 83–91. [Google Scholar] [CrossRef]
- Mittova, V.; Guy, M.; Tal, M.; Volokita, M. Salinity up-regulates the antioxidative system in root mitochondria and peroxisomes of the wild salt-tolerant tomato species Lycopersicon pennellii. J. Exp. Bot. 2004, 55, 1105–1113. [Google Scholar] [CrossRef]
- Xu, X.; Martin, B.; Comstock, J.P.; Vision, T.J.; Tauer, C.G.; Zhao, B.; Pausch, R.C.; Knapp, S. Fine mapping a QTL for carbon isotope composition in tomato. Theor. Appl. Genet. 2008, 117, 221–233. [Google Scholar] [CrossRef]
- Chetelat, R.T.; Pertuzé, R.A.; Faúndez, L.; Graham, E.B.; Jones, C.M. Distribution, ecology and reproductive biology of wild tomates and related nightshades from the Atacama Desert region of northern Chile. Euphytica 2009, 167, 77–93. [Google Scholar] [CrossRef]
- Foolad, M.R.; Lin, G.Y. Relationship between cold tolerance during seed germination and vegetative growth in tomato: Germplasm evaluation. J. Amer. Soc. Hort. Sci. 2000, 125, 679–683. [Google Scholar] [CrossRef]
- Venema, J.H.; Linger, P.; Van-Heusden, A.W.; Van-Hasselt, P.R.; Brüggemann, W. The inheritance of chilling tolerance in tomato (Lycopersicon spp.). Plant Biol. 2005, 7, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, X.; Chen, D.; Li, J.; Yi, Z.; Wang, A. A comparison of the low temperature transcriptomes of two tomato genotypes that differ in freezing tolerance: Solanum lycopersicum and Solanum habrochaites. BMC Plant Biol. 2015, 15, 132. [Google Scholar] [CrossRef]
- Zhao, L.; Qiu, C.; Li, J.; Chai, Y.; Kai, G.; Li, Z.; Sun, X.; Tang, K.X. Investigation of Disease Resistance and Cold Tolerance of Solanum lycopersicoides for Tomato Improvement. Hort. Sci. 2005, 40, 43–46. [Google Scholar] [CrossRef]
- Correll, D.S. The Potato and Its Wild Relatives; Texas Research Foundation, University of Michigan: Ann Arbor, MI, USA, 1962; Volume 4, pp. 1–606. [Google Scholar]
- Child, A. A synopsis of Solanum subgenus Potatoe (G. Don) (D’Arcy) (Tuberarium (Dun.) Bitter (s.l.)). Feddes Reper. 1990, 101, 209–235. [Google Scholar] [CrossRef]
- Peralta, I.E.; Spooner, D.M. Granule-Bound Starch Synthase (GBSSI) gene phylogeny of wild tomatoes (Solanum L. section Lycopersicon [Mill.] Wettst. subsection Lycopersicon). Am. J. Bot. 2001, 88, 1888–1902. [Google Scholar] [CrossRef]
- Marshall, J.A.; Knapp, S.; Davey, M.R.; Power, J.B.; Cocking, E.C.; Bennett, M.D.; Cox, A.V. Molecular systematics of Solanum section Lycopersicum (Lycopersicon) using the nuclear ITS rDNA region. Theor. Appl. Genet. 2001, 103, 1216–1222. [Google Scholar] [CrossRef]
- Álvarez, A.E.; van de Wiel, C.; Smulders, M.; Vosman, B. Use of microsatellites to evaluate genetic diversity and species relationships in the genus Lycopersicon. Theor. Appl. Genet. 2001, 103, 1283–1292. [Google Scholar] [CrossRef]
- Peralta, I.E.; Spooner, D.M. Morphological characterization and relationships of wild tomatoes (Solanum L. Section Lycopersicon [Mill.]). In A Festschrift for William G. D’Arcy: The Legacy of a Taxonomist; Keating, R.C., Hollowell, V.A., Croat, T.B., Eds.; Missouri Botanical Garden Press: St. Louis, MO, USA, 2005; pp. 227–257. [Google Scholar]
- Conesa, M.A.; Muir, C.D.; Roldán, E.J.; Molins, A.; Perdomo, J.A.; Galmés, J. Growth capacity in wild tomatoes and relatives correlates with original climate in arid and semi-arid species. Environ. Exp. Bot. 2017. [Google Scholar] [CrossRef]
- Arellano-Rodríguez, L.J.; Rodríguez-Guzmán, E.; Ron-Parra, J.; Martínez-Ramírez, J.L.; Lozoya-Saldaña, H.; Sánchez-Martínez, J.; Lépiz-Idelfonso, R. Evaluación de resistencia a Phytophthora en poblaciones silvestres de Solanum lycopersicum var. cerasiforme. Rev. Mex. Cien. Agríc. 2013, 4, 753–766. [Google Scholar] [CrossRef]
- Bonilla-Barrientos, O.; Lobato-Ortíz, R.; García-Zavala, J.J.; Cruz-Izquierdo, S.; Reyes-López, D.; Hernández-Leal, E.; Hernández-Bautista, A. Diversidad agronómica y morfológica de tomates arriñonados y tipo pimiento de uso local en Puebla y Oaxaca, México. Rev. Fitotec. Mex. 2014, 37, 129–139. [Google Scholar] [CrossRef]
- Maldonado-Peralta, R.; Ramírez-Vallejo, P.; González-Hernández, V.A.; Castillo-González, F.; Sandoval-Villa, M.; Livera-Muñoz, M.; Cruz-Huerta, N. Riqueza agronómica en colectas mexicanas de tomates (Solanum lycopersicum L.) nativos. Agroproductividad 2016, 9, 68–75. [Google Scholar]
- Velasco-Alvarado, M.J.; Lobato-Ortiz, R.; García-Zavala, J.J.; Castro-Brindis, R.; Cruz-Izquierdo, S.; Corona-Torres, T.; Moedano-Mariano, M.K. Mexican native tomatoes as rootstocks to increase fruit yield. Chilean J. Agric. Res. 2017, 77, 187–193. [Google Scholar] [CrossRef][Green Version]
- Flores-Hernández, L.A.; Lobato-Ortíz, R.; Sangerman-Jarquín, D.M.; García-Zavala, J.J.; Molina-Galán, J.D.; Velasco-Alvarado, M.J.; Marín-Montes, I.M. Genetic diversity within wild species of Solanum. Rev. Chapingo Ser. Hortic. 2018, 24, 85–96. [Google Scholar] [CrossRef]
- REMIB (Red Mundial de Información sobre Biodiversidad), Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. Available online: http://www.conabio.gob.mx/remib/ (accessed on 19 December 2020).
- GBIF (Global Biodiversity Information Facility). Available online: https://www.gbif.org/ (accessed on 19 December 2020).
- Solanaceae Source. A Global Taxonomic Source for the Nightshade Family. Available online: http://solanaceaesource.org/ (accessed on 15 December 2020).
- Worldclim. 2019. Available online: https://www.worldclim.org/data/index.html (accessed on 10 December 2020).
- Trabucco, A.; Zomer, R.J. Global High-Resolution Soil-Water Balance. Figshare. Fileset 2019. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 1. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteolologische Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Conover, W.J. Practical Nonparametric Statistics, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 1999; p. 584. [Google Scholar]
- SAS Institute. SAS/STAT User’s Guide: Software Version 9.3; Statistical Analysis System Institute: Cary, NC, USA, 2011. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section/Group | Solanum Species | Ecological Distribution | Altitude [1] | Altitude [23] |
|---|---|---|---|---|
| Section Lycopersicon Lycopersicon | S. cheesmaniae | Endemic to the Galapagos Islands. It inhabits dry, open, and rocky slopes, cold places. | 0–1300 | 0–1500 |
| S. galapagense | Endemic to the Galapagos Islands, on dry, open, and rocky slopes. | 0–600 (1500) | 0–650 | |
| S. pimpinellifolium | Southern region of Ecuador to the northern region of Chile. Dry slopes, plains, and associated with crop areas. | 0–500 | 0–500 | |
| Arcanum | S. arcanum | Northern Peru in inter-Andean dry valleys and coastal ecosystems with seasonal fog. Generally dry sites, rocky slopes. | 100–2500 | 500–3000 |
| S. chmielewskii | Southern zone of Peru and the northern zone of Bolivia. Wet and well-drained rocky slopes. | 2300–3000 | 1600–3100 | |
| S. neorickii | Southern Ecuador to southern Peru, in inter-Andean dry valleys. | 1950–3000 | 1500–2500 | |
| Ericopersicon | S. huaylasense | Northern and central Peru, on dry, open, and rocky slopes. | 1700–000 | 1000–900 |
| S. corneliomulleri | Southern Peru in regions with dry and rocky slopes. | 1000–3000 | 1000–3000 | |
| S. peruvianum | Central region of Peru to northern Chile in dry coastal deserts and seasonal mist ecosystems. | 0–600 | 0–2500 | |
| S. chilense | Coastal zone of Chile and northern Peru, on dry rocky slopes, and occasionally saline. | 0–3000 | 50–3500 | |
| S. habrochaites | Andean region of Ecuador and Peru in montane forests and dry slopes, occasionally found in seasonal fog ecosystems. | 400–3600 | 40–3300 | |
| Neolicopersicon | S. pennellii | North of Peru to the north of Chile, in areas of dry slopes, generally in flat areas. | 0–3000 | 0–1920 |
| Section Juglandifolia | S. ochranthum | Andean region of Colombia, Ecuador, and Peru, areas of mountain mesophilic forest. | 1900–4100 | 1200–3200 |
| S. juglandifollium | Andean region of Colombia, Ecuador, and Peru in areas of mountain mesophilic forest. | 1200–3100 | 1200–3100 | |
| Section Lycopersicoides | S. lycopersicoides | Southern area of Peru and northern Chile. In ravines and rocky slopes. | 1500–3700 | 1200–3700 |
| S. sitiens | Hyper-arid areas, northern region of Chile. | 2350–3500 | 2500–3500 |
| Sections/Group | Species | ALT | TEMP | DRAN | RAIN | EVAPO | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (m) | (°C) | (°C) | (mm) | (mm) | |||||||||||||||||
| Max | Min | Med | CV | Max | Min | Med | CV | Max | Min | Med | CV | Max | Min | Med | CV | Max | Min | Med | CV | ||
| Section Lycopersicon Lycopersicon | S. cheesmaniae | 1478 | 5 | 87 | 155.20 | 25 | 17.1 | 23.6 | 3.27 | 10.3 | 7.6 | 8.5 | 4.57 | 562 | 107 | 277 | 21.70 | 1125 | 187 | 454 | 42.24 |
| S. galapagense | 868 | 4 | 45 | 240.00 | 25 | 19.8 | 23.9 | 2.87 | 10.1 | 7.6 | 8.7 | 4.91 | 546 | 135 | 274 | 16.06 | 930 | 262 | 531 | 30.60 | |
| S. pimpinelifollium | 633 | 1 | 93 | 100.00 | 26.8 | 17.9 | 22.7 | 8.54 | 13.3 | 6.3 | 9.8 | 8.37 | 2989 | 1 | 70 | 157.86 | 1710 | 1 | 45 | 202.22 | |
| Arcanum | S. arcanum | 3292 | 132 | 1681 | 36.76 | 24.1 | 11.4 | 18.4 | 9.19 | 14.4 | 10.3 | 12.4 | 3.42 | 1193 | 22 | 527 | 43.45 | 1094 | 11 | 395 | 43.29 |
| S. chmielewskii | 3195 | 1953 | 2583 | 9.52 | 19.9 | 13 | 16.8 | 9.88 | 15.6 | 11.7 | 14.9 | 3.21 | 1318 | 504 | 944 | 18.51 | 874 | 429 | 647 | 16.74 | |
| S. neorickii | 3262 | 1705 | 2317 | 11.57 | 20.3 | 11.7 | 16.9 | 7.88 | 15.5 | 9.9 | 13.1 | 2.19 | 1366 | 426 | 816 | 23.53 | 1031 | 326 | 672 | 14.29 | |
| Ericopersicon | S. huaylasense | 3124 | 1141 | 2291 | 16.06 | 20.3 | 11.2 | 16.8 | 8.38 | 13.9 | 10.8 | 13.4 | 1.27 | 500 | 128 | 346 | 23.99 | 416 | 73 | 278 | 30.76 |
| S. corneliomulleri | 3097 | 1018 | 2344 | 18.75 | 18.3 | 9.6 | 14.1 | 12.95 | 16.6 | 9.4 | 12.3 | 4.07 | 434 | 19 | 201 | 60.70 | 354 | 12 | 141 | 48.58 | |
| S. peruvianum | 2617 | 2 | 528 | 124.03 | 20.9 | 11.7 | 18.6 | 6.98 | 15.5 | 4.7 | 9.4 | 15.61 | 434 | 0 | 25 | 92.00 | 324 | 0 | 13 | 130.77 | |
| S. chilense | 3995 | 0 | 1904 | 59.53 | 20.4 | 5.4 | 15.2 | 19.29 | 18.6 | 4.9 | 12.6 | 10.82 | 355 | 0 | 28 | 69.64 | 275 | 3 | 20 | 72.50 | |
| S. habrochaites | 3692 | 40 | 2033 | 33.78 | 25.8 | 7 | 16.6 | 17.30 | 14.5 | 6.5 | 11.8 | 5.26 | 2358 | 11 | 622 | 43.25 | 1682 | 8 | 555 | 43.69 | |
| Neolicopersicon | S. pennellii | 2921 | 5 | 822 | 53.16 | 25.1 | 10.5 | 18.4 | 7.38 | 13.7 | 6.2 | 10.2 | 8.35 | 404 | 1 | 49 | 95.92 | 289 | 0 | 33 | 96.97 |
| Section Juglandifolia | S. juglandifolium | 3353 | 1005 | 2195 | 14.49 | 22.9 | 8.9 | 15.8 | 10.68 | 12.5 | 7.2 | 9.1 | 1.94 | 3214 | 550 | 1895 | 28.79 | 1648 | 413 | 1177 | 10.24 |
| S. ochranthum | 4008 | 1195 | 2750 | 10.27 | 21.9 | 6.7 | 13.9 | 11.65 | 15.6 | 7.2 | 11.4 | 3.42 | 2358 | 507 | 1010 | 11.44 | 1474 | 286 | 818 | 13.57 | |
| Section Lycopersicoides | S. lycopersicoides | 3775 | 1290 | 2928 | 13.54 | 17.3 | 7.6 | 11.2 | 15.28 | 15.1 | 10.4 | 14.1 | 4.55 | 215 | 13 | 104 | 53.14 | 182 | 9 | 80 | 60.94 |
| S. sitiens | 3330 | 2276 | 2740 | 5.90 | 13.2 | 8.4 | 11.4 | 7.94 | 17.6 | 15.6 | 16.8 | 1.26 | 31 | 8 | 17 | 25.00 | 26 | 9 | 21 | 16.67 | |
| Group | CLUTER | Spe-Clim | ALT (msnm) | TEMP (°C) | DRAN (°C) | RAINF (mm) | EVAPO (mm) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Med | CV | Med | CV | Med | CV | Med | CV | Med | CV | |||
| E | 1 | CHI-5 | 2280 | 44.52 | 14.1 | 20.36 | 13.1 | 16.98 | 36 | 54.17 | 26 | 59.62 |
| E | 1 | CHI-7 | 3662 | 12.56 | 8.1 | 26.39 | 14.8 | 8.26 | 238 | 16.81 | 208 | 13.73 |
| E | 1 | COR-5 | 1989 | 22.08 | 15.6 | 6.65 | 12 | 13.81 | 130 | 46.15 | 94 | 36.9 |
| E | 1 | COR-7 | 2637 | 7.47 | 11.7 | 8.4 | 12.3 | 2.2 | 360 | 9.17 | 239 | 15.9 |
| E | 1 | HAB-29 | 3558 | 3.11 | 9.2 | 3.38 | 13.4 | 4.23 | 562 | 1.6 | 480 | 18.13 |
| E | 1 | HAB-5 | 1805 | 9.17 | 15.8 | 7.66 | 11.6 | 2.48 | 194 | 16.49 | 137 | 28.1 |
| Y | 1 | LYC-5 | 2881 | 13 | 11.6 | 13.83 | 14 | 4.78 | 93 | 59.68 | 76 | 61.18 |
| Y | 1 | LYC-7 | 3654 | 0 | 8.1 | 0 | 14.9 | 0 | 213 | 0 | 182 | 0.27 |
| N | 1 | PEN-5 | 1531 | 16.98 | 16.7 | 4.53 | 11 | 6.64 | 90 | 56.11 | 75 | 46.67 |
| N | 1 | PEN-7 | 2526 | 8.22 | 12.8 | 15.18 | 12.6 | 1.73 | 335 | 11.79 | 250 | 14.83 |
| E | 1 | PER-5 | 1713 | 29.19 | 16 | 7.72 | 11.6 | 12.98 | 55 | 104.55 | 51 | 77.45 |
| E | 1 | PER-7 | 2472 | 4.69 | 12.9 | 3.03 | 12.3 | 3.73 | 339 | 12.24 | 213 | 17.84 |
| Y | 1 | SIT-5 | 2740 | 5.9 | 11.4 | 7.94 | 16.8 | 0.95 | 17 | 25 | 21 | 16.67 |
| A | 2 | ARC-4 | 679 | 38.25 | 20 | 2.3 | 11.5 | 5.29 | 91 | 101.1 | 84 | 108.33 |
| L | 2 | CHE-1 | 620 | 13.43 | 21.3 | 3.6 | 8.7 | 8.23 | 305 | 7.46 | 988 | 6.4 |
| L | 2 | CHE-15 | 1013 | 10.27 | 19.7 | 2.61 | 10 | 2.72 | 328 | 4.81 | 992 | 4.39 |
| L | 2 | CHE-3 | 144 | 79.44 | 23.4 | 2.12 | 8.2 | 4.8 | 275 | 16 | 457 | 38.29 |
| L | 2 | CHE-4 | 47 | 63.98 | 24.3 | 0.69 | 8.9 | 1.66 | 202 | 22.4 | 290 | 7.67 |
| L | 2 | CHE-6 | 39 | 108.97 | 24.1 | 1.24 | 8.4 | 2.54 | 359 | 39.28 | 504 | 12.81 |
| E | 2 | CHI-4 | 385 | 90.78 | 18.7 | 2.31 | 9 | 20.18 | 7 | 71.43 | 7 | 21.43 |
| L | 2 | GAL-3 | 167 | 81.14 | 23.4 | 2.71 | 8.6 | 4.7 | 276 | 17.93 | 562 | 29.36 |
| L | 2 | GAL-4 | 20 | 77.5 | 24.4 | 1.27 | 9.1 | 4.32 | 249 | 18.07 | 322 | 39.91 |
| L | 2 | GAL-6 | 18 | 83.33 | 24.4 | 0.52 | 9.3 | 5.1 | 275 | 11.27 | 531 | 19.68 |
| E | 2 | HAB-4 | 326 | 110.43 | 20.6 | 9.33 | 11.3 | 6.28 | 114 | 57.46 | 77 | 92.86 |
| N | 2 | PEN-4 | 622 | 43.09 | 19.6 | 4.5 | 10 | 8.15 | 34 | 88.24 | 19 | 73.68 |
| L | 2 | PIM-3 | 27 | 414.81 | 25.2 | 1.42 | 9.1 | 5.01 | 1190 | 35.88 | 861 | 12.31 |
| L | 2 | PIM-4 | 95 | 87.37 | 22 | 8.03 | 10.1 | 12.63 | 50 | 119 | 25 | 224 |
| L | 2 | PIM-6 | 64 | 276.38 | 24.7 | 2.58 | 9.8 | 4.61 | 602 | 14.29 | 485 | 15.77 |
| A | 3 | ARC-12 | 2373 | 9.59 | 16.3 | 6.04 | 13.6 | 2.39 | 716 | 8.1 | 594 | 9.6 |
| A | 3 | ARC-15 | 2115 | 13.36 | 16.8 | 6.28 | 12.3 | 2.43 | 863 | 6.55 | 745 | 7.99 |
| A | 3 | ARC-7 | 2537 | 17.03 | 15.1 | 11.98 | 13.4 | 2.46 | 352 | 34.23 | 370 | 13.24 |
| A | 3 | CHM-12 | 2601 | 9.88 | 16.7 | 10.31 | 15 | 1.92 | 948 | 14.45 | 663 | 14.25 |
| A | 3 | CHM-7 | 2560 | 4.02 | 17.1 | 4.81 | 14.3 | 1.66 | 534 | 17.88 | 449 | 24.28 |
| E | 3 | HAB-12 | 2692 | 9.68 | 14.8 | 9.02 | 13.7 | 8.13 | 761 | 8.54 | 645 | 7.67 |
| E | 3 | HAB-7 | 2648 | 10.84 | 13.4 | 12.73 | 12.9 | 4.96 | 393 | 21.12 | 296 | 32.6 |
| E | 3 | HUA-7 | 2337 | 9.99 | 16.6 | 5.51 | 13.6 | 2.67 | 354 | 20.06 | 282 | 28.19 |
| A | 3 | NEO-12 | 2564 | 10.26 | 16.8 | 9.75 | 14.6 | 5.17 | 999 | 27.18 | 626 | 16.37 |
| A | 3 | NEO-7 | 2096 | 11.09 | 18.7 | 8.45 | 13.8 | 2.15 | 541 | 7.95 | 419 | 24.94 |
| J | 3 | OCH-12 | 3010 | 11.05 | 13.8 | 10.51 | 14.4 | 4.62 | 967 | 17.12 | 664 | 14.31 |
| J | 3 | OCH-7 | 2865 | 10.12 | 15 | 4.04 | 15.2 | 10.95 | 508 | 25 | 442 | 3.73 |
| E | 4 | HAB-15 | 2270 | 8.37 | 16.1 | 5.65 | 11.6 | 7.69 | 921 | 13.74 | 799 | 12.52 |
| E | 4 | HAB-9 | 2733 | 12.22 | 13.9 | 8.39 | 11 | 1.02 | 595 | 2.69 | 582 | 10.82 |
| J | 4 | JUG-12 | 2428 | 7.91 | 13.7 | 4.34 | 10 | 6.12 | 760 | 18.42 | 778 | 8.74 |
| J | 4 | JUG-29 | 3144 | 0 | 10.9 | 0 | 10.7 | 0 | 1222 | 0 | 1,015 | 0 |
| A | 4 | NEO-15 | 2153 | 11.18 | 16.9 | 6.68 | 11.8 | 4.17 | 817 | 15.09 | 719 | 7.82 |
| J | 4 | OCH-15 | 2626 | 9.12 | 14 | 9.08 | 11.2 | 8 | 1041 | 10.25 | 825 | 13.58 |
| J | 4 | OCH-29 | 3372 | 4.09 | 9.1 | 6.24 | 9.9 | 7.99 | 1010 | 6.76 | 812 | 11.02 |
| J | 4 | OCH-9 | 2725 | 5.42 | 13.8 | 5.12 | 10.9 | 6.91 | 1004 | 10.48 | 932 | 9.87 |
| A | 5 | ARC-2 | 1216 | 0 | 21.4 | 0 | 11.1 | 0 | 1193 | 0 | 1094 | 0 |
| A | 5 | ARC-3 | 1448 | 6.01 | 19.9 | 2.25 | 12.2 | 0.31 | 694 | 5.69 | 537 | 4.75 |
| A | 5 | ARC-5 | 1828 | 6.44 | 17.3 | 2.77 | 12.2 | 4.99 | 232 | 27.05 | 179 | 42.58 |
| A | 5 | ARC-6 | 1259 | 13.15 | 19.4 | 5.59 | 12.5 | 3.21 | 453 | 22.02 | 358 | 24.23 |
| E | 5 | HAB-3 | 1415 | 20.61 | 20.7 | 7.82 | 11.7 | 5.38 | 918 | 13.73 | 840 | 12.56 |
| E | 5 | HAB-6 | 1077 | 32.4 | 21.3 | 7.65 | 11.7 | 3.77 | 566 | 18.73 | 479 | 18.37 |
| E | 5 | HUA-4 | 1234 | 15.36 | 19.3 | 2.65 | 12.4 | 7.72 | 247 | 32.79 | 194 | 34.54 |
| E | 5 | HUA-6 | 1477 | 19.77 | 19.3 | 3.66 | 12.6 | 3.35 | 304 | 8.22 | 215 | 7.44 |
| J | 5 | JUG-3 | 1510 | 9.3 | 18.4 | 3.41 | 10.2 | 3.23 | 1,099 | 18.15 | 737 | 23.0 |
| J | 5 | OCH-3 | 1734 | 6.31 | 19.2 | 3.69 | 11.7 | 4.15 | 860 | 6.89 | 867 | 14.95 |
| E | 6 | HAB-1 | 629 | 51.55 | 24.2 | 3.79 | 10.9 | 2.06 | 1708 | 9.69 | 1549 | 8.78 |
| J | 6 | JUG-1 | 1515 | 10.73 | 19.7 | 5.06 | 9.6 | 6.74 | 2274 | 9.8 | 1363 | 6.64 |
| J | 6 | JUG-15 | 2275 | 9.78 | 15.4 | 8.23 | 9.1 | 7.35 | 1941 | 27.79 | 1183 | 8.54 |
| J | 6 | JUG-2 | 1325 | 8.42 | 19.6 | 3.03 | 8.6 | 2.76 | 2087 | 8.53 | 1189 | 3.36 |
| J | 6 | JUG-9 | 1933 | 12.78 | 16.6 | 6.31 | 9.0 | 5.15 | 1456 | 18.37 | 1148 | 3.53 |
| J | 6 | OCH-1 | 1472 | 10.09 | 19.9 | 4.57 | 9.9 | 5.47 | 2009 | 10.63 | 1333 | 12.87 |
| L | 6 | PIM-1 | 379 | 41.56 | 25.5 | 3.5 | 10.4 | 6.78 | 2080 | 20.82 | 1655 | 15.38 |
| L | 6 | PIM-2 | 289 | 60.14 | 24.3 | 3.52 | 8.2 | 1.62 | 2494 | 17.3 | 1221 | 5.2 |
| CP1 | CP2 | CP3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| EVAPO | BIO12 | BIO13 | ALT | BIO8 | BIO5 | BIO2 | BIO1 | BIO15 | BIO3 | |
| 1 | 137 d | 194 d | 69 c | 2526 b | 14.0 cd | 20.5 d | 12.6 b | 12.8 d | 127 a | 74.6 c |
| 2 | 389 c | 262 d | 54 c | 155 d | 25.1 a | 30.0 a | 9.0 e | 22.7 a | 89 b | 67.7 d |
| 3 | 521 c | 628 c | 117 b | 2562 ab | 16.5 c | 24.0 c | 13.7 a | 16.4 c | 80 c | 84.1 b |
| 4 | 805 b | 962 b | 141 b | 2675 a | 13.9 d | 20.7 d | 10.9 d | 13.8 cd | 43 d | 85.0 ab |
| 5 | 508 c | 630 c | 129 b | 1431 c | 20.3 b | 26.0 b | 11.9 c | 19.3 b | 92 bc | 86.6 a |
| 6 | 1277 a | 2044 a | 253 a | 1398 c | 19.8 b | 25.4 b | 9.3 e | 19.7 ab | 35 d | 88.9 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Ojeda, G.; Peralta, I.E.; Rodríguez-Guzmán, E.; Chávez-Servia, J.L.; Sahagún-Castellanos, J.; Rodríguez-Pérez, J.E. Climatic Diversity and Ecological Descriptors of Wild Tomato Species (Solanum sect. Lycopersicon) and Close Related Species (Solanum sect. Juglandifolia y sect. Lycopersicoides) in Latin America. Plants 2021, 10, 855. https://doi.org/10.3390/plants10050855
Ramírez-Ojeda G, Peralta IE, Rodríguez-Guzmán E, Chávez-Servia JL, Sahagún-Castellanos J, Rodríguez-Pérez JE. Climatic Diversity and Ecological Descriptors of Wild Tomato Species (Solanum sect. Lycopersicon) and Close Related Species (Solanum sect. Juglandifolia y sect. Lycopersicoides) in Latin America. Plants. 2021; 10(5):855. https://doi.org/10.3390/plants10050855
Chicago/Turabian StyleRamírez-Ojeda, Gabriela, Iris E. Peralta, Eduardo Rodríguez-Guzmán, José Luis Chávez-Servia, Jaime Sahagún-Castellanos, and Juan Enrique Rodríguez-Pérez. 2021. "Climatic Diversity and Ecological Descriptors of Wild Tomato Species (Solanum sect. Lycopersicon) and Close Related Species (Solanum sect. Juglandifolia y sect. Lycopersicoides) in Latin America" Plants 10, no. 5: 855. https://doi.org/10.3390/plants10050855
APA StyleRamírez-Ojeda, G., Peralta, I. E., Rodríguez-Guzmán, E., Chávez-Servia, J. L., Sahagún-Castellanos, J., & Rodríguez-Pérez, J. E. (2021). Climatic Diversity and Ecological Descriptors of Wild Tomato Species (Solanum sect. Lycopersicon) and Close Related Species (Solanum sect. Juglandifolia y sect. Lycopersicoides) in Latin America. Plants, 10(5), 855. https://doi.org/10.3390/plants10050855