
The Effect of Supplementary LED Lighting on the Morphological and Physiological Traits of Miniature Rosa × Hybrida ‘Aga’ and the Development of Powdery Mildew (Podosphaera pannosa) under Greenhouse Conditions


Abstract
1. Introduction
2. Results and Discussion
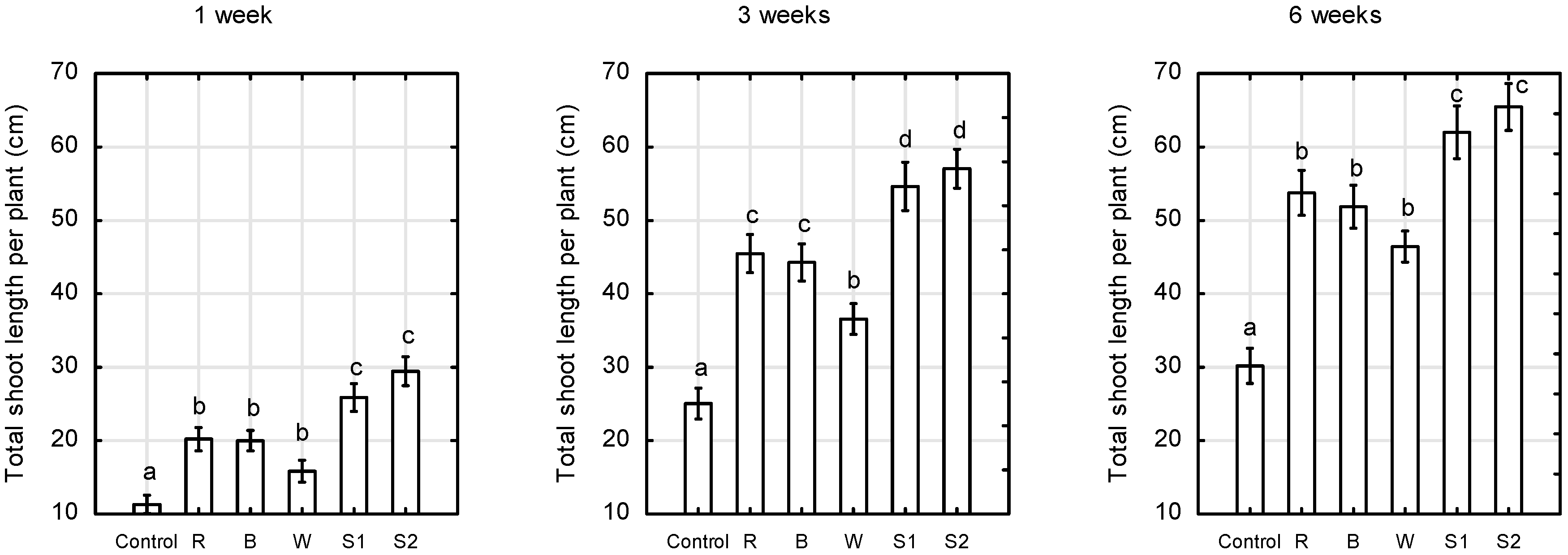
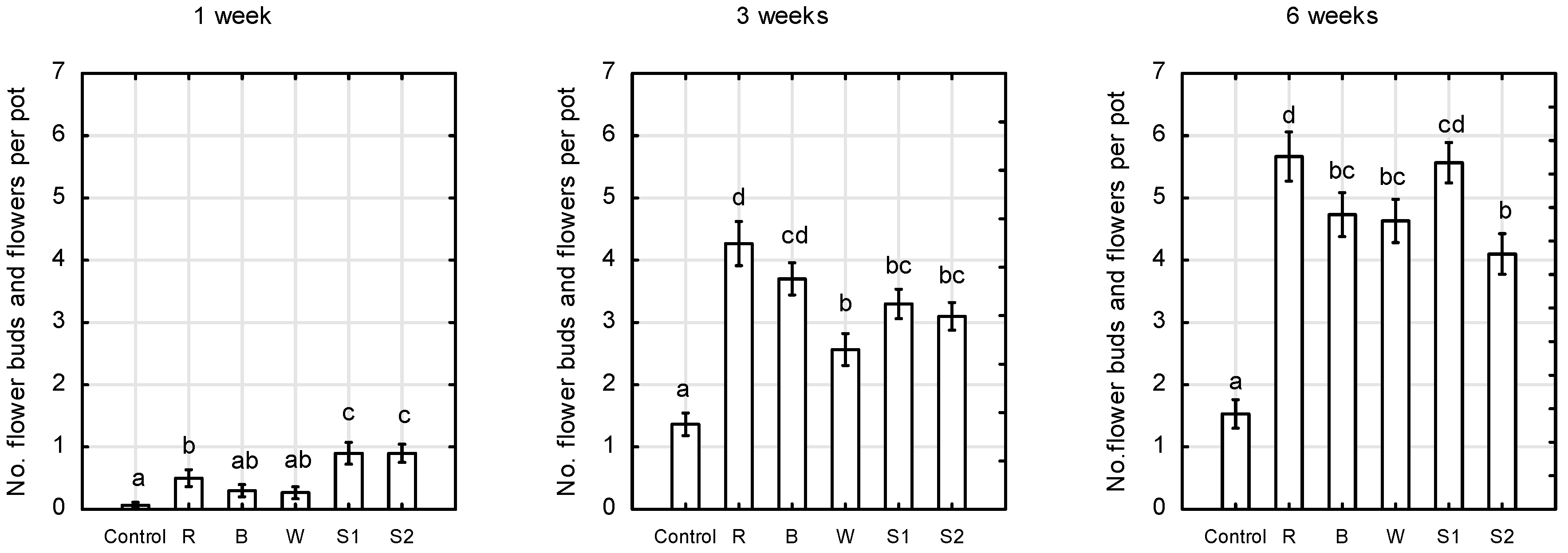
2.1. Plant Growth and Morphology
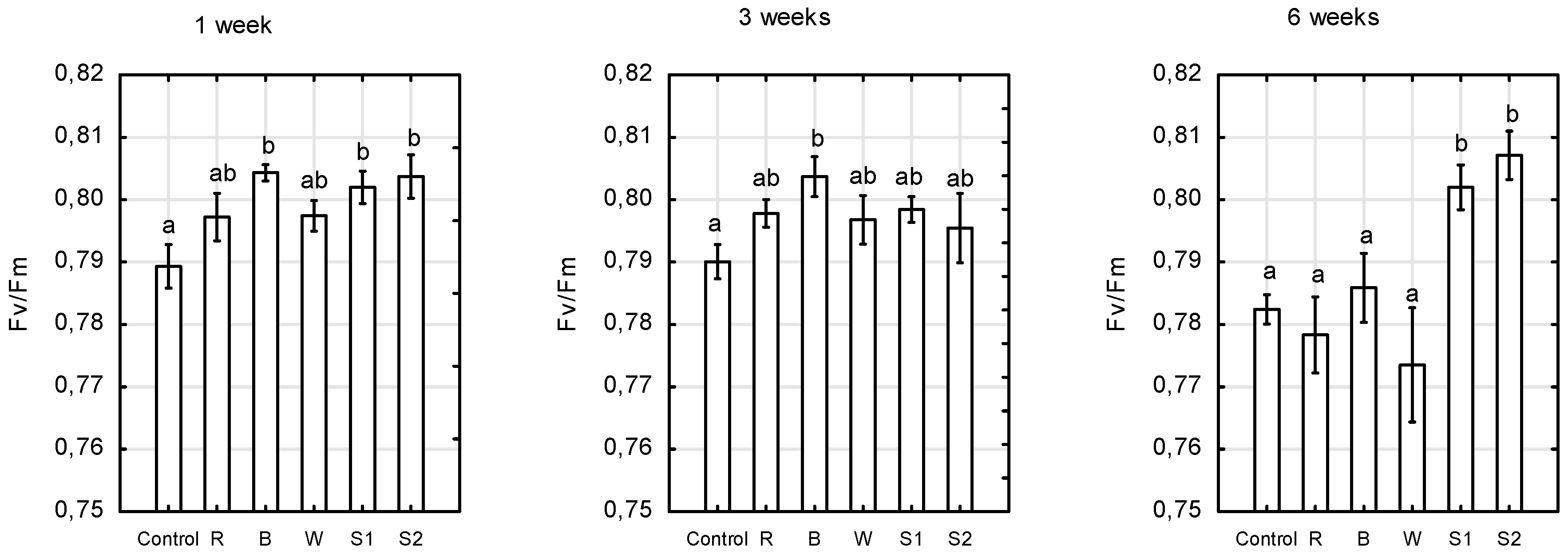
2.2. Chlorophyll Fluorescence
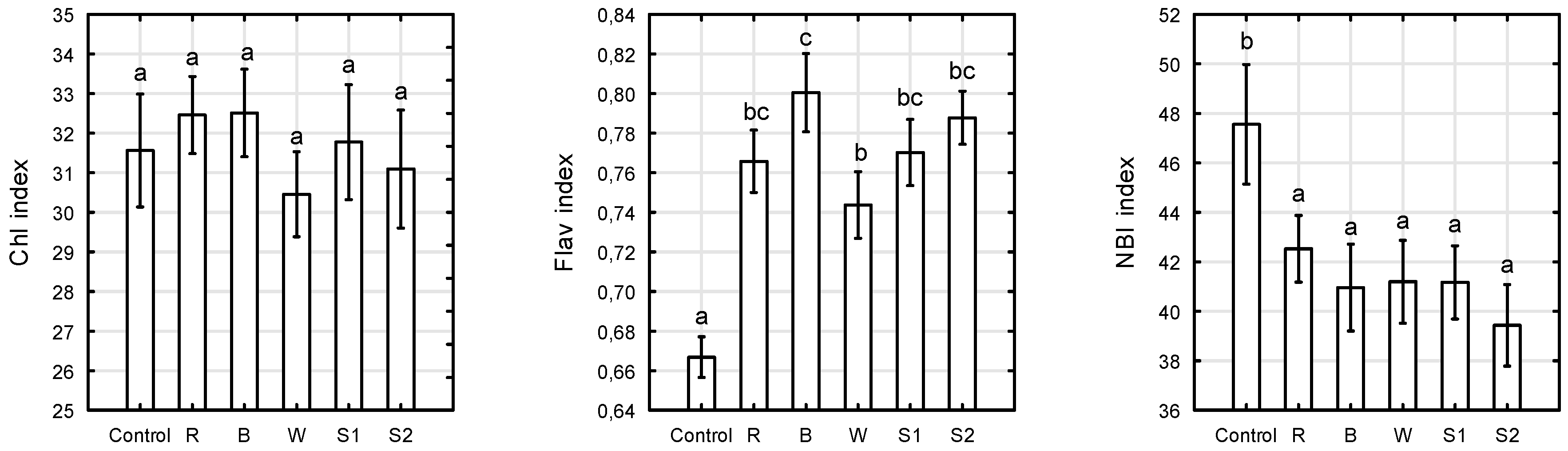
2.3. Chlorophyll, Flavonols, and Nitrogen Balance Indices
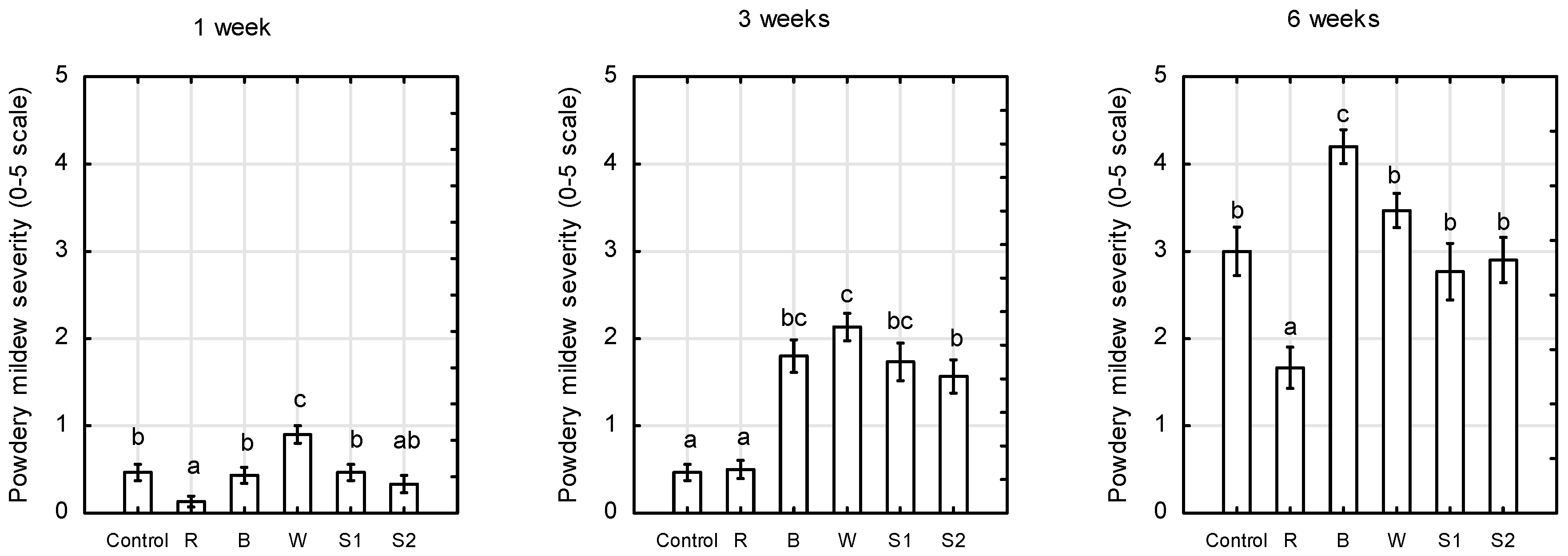
2.4. Disease Evaluation
3. Materials and Methods
3.1. Plant Material and Growth Conditions
3.2. Light Treatments
3.3. Growth Parameters
3.4. Chlorophyll, Flavonol, and Nitrogen Balance Indices
3.5. Chlorophyll Fluorescence Parameters
3.6. Disease Evaluation
3.7. Statistical Analysis
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eurostat. Horticultural Products. Flowers and Ornamental Plants—Production. In Statistics 2010–2019; Eurostat: Luxembourg, 2020. [Google Scholar]
- Moe, R.; Grimstad, S.O.; Gislerod, H.R. The use of artificial light in year round production of greenhouse crops in Norway. Acta Hortic. 2006, 711, 35–42. [Google Scholar] [CrossRef]
- Fanourakis, D.; Hyldgaard, B.; Giday, H.; Aulik, I.; Bouranis, D.; Körner, O.; Ottosen, C. Stomatal anatomy and closing ability is affected by supplementary light intensity in rose (Rosa hybrida L.). Hort. Sci. 2019, 46, 81–89. [Google Scholar] [CrossRef]
- Morrow, R.C. LED lighting in horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef]
- Olle, M.; Viršile, A. The effects of light-emitting diode lighting on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar] [CrossRef]
- Stutte, G.W. Commercial transition to LEDs: A pathway to high-value products. HortScience 2015, 50, 1297–1300. [Google Scholar] [CrossRef]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Ouzounis, T.; Giday, H.; Kjaer, K.H.; Ottosen, C.-O. LED or HPS in ornamentals? A case study in roses and campanulas. Eur. J. Hortic. Sci. 2018, 83, 166–172. [Google Scholar] [CrossRef]
- Ouzounis, T.; Fretté, X.; Rosenqvist, E.; Ottosen, C.-O. Spectral effects of supplementary lighting on the secondary metabolites in roses, chrysanthemums, and campanulas. J. Plant Physiol. 2014, 171, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.-O. Spectral effects of artificial light on plant physiology and secondary metabolism: A review. HortScience 2015, 50, 1128–1135. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M. An overview of LED lighting and spectral quality on plant photosynthesis. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Alsanius, B.W.; Karlsson, M.; Rosberg, A.K.; Dorais, M.; Naznin, M.T.; Khalil, S.; Bergstrand, K.-J. Light and microbial lifestyle: The impact of light quality on plant-microbe interactions in horticultural production systems—A review. Horticulturae 2019, 5, 41. [Google Scholar] [CrossRef]
- Davis, P.A.; Burns, C. Photobiology in protected horticulture. Food Energy Secur. 2016, 5, 223–238. [Google Scholar] [CrossRef]
- Paradiso, R.; Meinen, E.; Snel, J.F.H.; De Visser, P.; Van Ieperen, W.; Hogewoning, S.W.; Marcelis, L.F.M. Spectral dependence of photosynthesis and light absorptance in single leaves and canopy in rose. Sci. Hortic. 2011, 127, 548–554. [Google Scholar] [CrossRef]
- Abidi, F.; Girault, T.; Douillet, O.; Guillemain, G.; Sintes, G.; Laffaire, M.; Ben Ahmed, H.; Smiti, S.; Huché-Thélier, L.; Leduc, N. Blue light effects on rose photosynthesis and photomorphogenesis. Plant Biol. 2013, 15, 67–74. [Google Scholar] [CrossRef]
- Terfa, M.T.; Solhaug, K.A.; Gislerød, H.R.; Olsen, J.E.; Torre, S. A high proportion of blue light increases the photosynthesis capacity and leaf formation rate of Rosa × hybrida but does not affect time to flower opening. Physiol. Plant. 2013, 148, 146–159. [Google Scholar] [CrossRef]
- Terfa, M.T.; Olsen, J.E.; Torre, S. Blue light improves stomatal function and dark-induced closure of Rose leaves (Rosa x hybrida) developed at high air humidity. Front. Plant Sci. 2020, 11, 1036. [Google Scholar] [CrossRef]
- Kim, H.-H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green-light supplementation enhances lettuce growth under red- and blue-light-emitting diodes. HortScience 2004, 39, 1617–1622. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Xu, X.M.; Cui, J. Response of photosynthetic capacity of tomato leaves to different LED light wavelength. Environ. Exp. Bot. 2018, 150, 161–171. [Google Scholar] [CrossRef]
- Bayat, L.; Arab, M.; Aliniaeifard, S.; Seif, M.; Lastochkina, O.; Li, T. Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants 2018, 10. [Google Scholar] [CrossRef]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Tchélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environm. Experim. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Girault, T.; Bergougnoux, V.; Combes, D.; Viemont, J.D.; Leduc, N. Light controls shoot meristem organogenic activity and leaf primordia growth during bud burst in Rosa sp. Plant Cell Environm. 2008, 31, 1534–1544. [Google Scholar] [CrossRef]
- Debener, T.; Byrne, D.H. Disease resistance breeding in rose: Current status and potential of biotechnological tools. Plant Sci. 2014, 228, 107–117. [Google Scholar] [CrossRef]
- Ballaré, C.L.; Mazza, C.A.; Austin, A.T.; Pierik, R. Canopy light and plant health. Plant Physiol. 2012, 160, 145–155. [Google Scholar] [CrossRef]
- Carvalho, S.D.; Castillo, J.A. Influence of light on plant-phyllosphere interaction. Front. Plant Sci. 2018, 9, 1482. [Google Scholar] [CrossRef]
- Islam, S.Z.; Babadoos, M.; Honda, Y. Effect of red light treatment of seedlings of pepper, pumpkin, and tomato on the occurrence of Phytophthora damping-off. HortScience 2002, 37, 678–681. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Honda, Y.; Arase, S. Red-light-induced resistance in broad bean (Vicia faba L.) to leaf spot disease caused by Alternaria tenuissima. J. Phytopathol. 2003, 151, 86–91. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, Y.P.; Yu, H.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Light quality affects incidence of powdery mildew, expression of defense-related genes and associated metabolism in cucumber plants. Eur. J. Plant Pathol. 2010, 127, 125–135. [Google Scholar] [CrossRef]
- Kook, H.-S.; Park, S.-H.; Jang, Y.-J.; Lee, G.-W.; Kim, J.S.; Kim, H.M.; Oh, B.-T.; Chae, J.-C.; Lee, K.-J. Blue LED (light-emitting diodes)-mediated growth promotion and control of Botrytis disease in lettuce. Acta Agric. Scand. B Soil Plant Sci. 2013, 63, 271–277. [Google Scholar] [CrossRef]
- Suthaparan, A.; Stensvand, A.; Solhaug, K.A.; Torre, S.; Mortensen, L.M.; Gadoury, D.M.; Seem, R.C.; Gislerød, H.R. Suppression of powdery mildew (Podosphaera pannosa) in greenhouse roses by brief exposure to supplemental UV-B radiation. Plant Dis. 2012, 96, 1653–1660. [Google Scholar] [CrossRef]
- Suthaparan, A.; Torre, S.; Stensvand, A.; Herrero, M.L.; Pettersen, R.I.; Gadoury, D.M.; Gislerød, H.R. Specific light emitting diodes can suppress sporulation of Podosphaera pannosa on greenhouse roses. Plant Dis. 2010, 94, 1105–1110. [Google Scholar] [CrossRef]
- Aboody, M.; Mickymaray, S. Anti-fungal efficacy and mechanisms of flavonoids. Antibiotics 2020, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in biotic and abiotic stress responses, and microbial associations. Review. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Idris, A.; Linatoc, A.C.; Bakar, M.F.A.; Takai, Z.I.; Audu, Y. Effect of light quality and quantity on the accumulation of flavonoid in plant species. J. Sci. Technol. 2018, 10, 32–45. [Google Scholar] [CrossRef]
- Gam, D.T.; Khoi, P.H.; Ngoc, P.B.; Linh, L.K.; Hung, N.K.; Anh, P.; Thu, N.T.; Hien, N.; Khanh, T.D.; Ha, C.H. LED lights promote growth and flavonoid accumulation of Anoectochilus roxburghii and linked to the enhanced expression of several related genes. Plants 2020, 9, 1344. [Google Scholar] [CrossRef] [PubMed]
- Cerovic, Z.G.; Masdoumier, G.; Ghozlen, N.B.; Latouche, G. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant. 2012, 146, 251–260. [Google Scholar] [CrossRef]
- Wojdyła, A.T. Possibility of using natural and synthetic products in the protection of rose against Podosphaera pannosa (Wallr.) de Bary. Zesz. Probl. Postępów Nauk. Rol. 2016, 586, 89–98. [Google Scholar]
- Ahlman, L.; Bånkestad, D.; Wik, T. Using chlorophyll a fluorescence gains to optimize LED light spectrum for short term photosynthesis. Comput. Electron. Agric. 2017, 142, 224–234. [Google Scholar] [CrossRef]
- Crespel, L.; Le Bras, C.; Amoroso, T.; Unda Ulloa, M.G.; Morel, P.; Sakr, S. Genotype × light quality interaction on Rose architecture. Agronomy 2020, 10, 913. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Wang, Y.; Tong, Y.; Chu, H.; Chen, X.; Guo, H.; Yuan, H.; Yan, D.; Zheng, B. Effects of different light qualities on seedling growth and chlorophyll fluorescence parameters of Dendrobium officinale. Biologia 2017, 72, 735–744. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [PubMed]
- Centofante, A.R.; Neto, A.R.; Vasconcelos-Filho, S.C.; Oliveira, E.A.; Assis, E.S.; Rosa, M.; Silva, F.G. Light quality on the morphoanatomy and physiology of Campomanesia pubescens (DC.) O. Berg. seedlings. Sci. Hortic. 2020, 259, 108765. [Google Scholar] [CrossRef]
- Zheng, L.; Van Labeke, M.C. Long-term effects of red- and blue-light emitting diodes on leaf anatomy and photosynthetic efficiency of three ornamental pot plants. Front Plant Sci. 2017, 8, 917. [Google Scholar] [CrossRef]
- Wang, Z.; Tian, J.; Yu, B.; Yang, L.; Sun, Y. LED light spectrum affects the photosynthetic performance of Houttuynia cordata seedlings. Am. J. Opt. Photonics 2015, 3, 38–42. [Google Scholar] [CrossRef]
- Ouzounis, T.; Frette, X.; Ottosen, C.-O.; Rosenqvist, E. Spectral effects of LEDs on chlorophyll fluorescence and pigmentation in Phalaenopsis “Vivien” and “Purple Star”. Physiol. Plant. 2015, 154, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Stutte, G.W.; Edney, S.; Skerritt, T. Photoregulation of bioprotectant content of red leaf lettuce with light-emitting diodes. HortScience 2009, 44, 79–82. [Google Scholar] [CrossRef]
- Taulavuori, K.; Pyysalo, A.; Taulavuori, E.; Julkunen-Tiitto, R. Responses of phenolic acid and flavonoid synthesis to blue and blue-violet light depends on plant species. Environ. Exp. Bot. 2018, 150, 183–187. [Google Scholar] [CrossRef]
- Matysiak, B.; Kowalski, A. White, blue and red LED lighting on growth, morphology and accumulation of flavonoid compounds in leafy greens. Zemdirbyste-Agriculture 2019, 106, 281–286. [Google Scholar] [CrossRef]
- Nybakken, L.; Lie, N.H.; Julkunen-Titto, R.; Asplund, J.; Ohlson, M. Fertilization changes chemical defense in needles of mature Norway spruce (Picea abies). Front. Plant Sci. 2018, 9, 770. [Google Scholar] [CrossRef]
- Deng, B.; Li, Y.; Xu, D.; Qingqing, Y.; Guihua, L. Nitrogen availability alters flavonoid accumulation in Cyclocarya paliurus via the effects on the internal carbon/nitrogen balance. Sci. Rep. 2019, 9, 2370. [Google Scholar] [CrossRef]
- Padilla, F.M.; Pena-Fleitas, M.T.; Gallardo, M.; Thompson, R.B. Evaluation of optical sensor measurements of canopy reflectance and of leaf flavonols and chlorophyll contents to assess crop nitrogen status of muskmelon. Eur. J. Agron. 2014, 58, 39–52. [Google Scholar] [CrossRef]
- Lillo, C.; Meyer, C.; Lea, U.S.; Provan, F.; Oltedal, S. Mechanism and importance of post-translational regulation of nitrate reductasse. J. Exp. Bot. 2004, 55, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Téllez, L.I.; Estrada-Ortiz, E.; Gómez-Merino, F.C.; Becker, C.; Krumbein, A.; Schwarz, D. Flavonoid, nitrate and glucosinolate concentrations in Brassica species are differentially affected by photosynthetically active radiation, phosphate and phosphite. Front. Plant Sci. 2019, 10, 371. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, C.; Caceres, M.E.; Ceccarelli, M.; Pica, A.L.; Rugini, E.; Cristofori, V. Influence of Continuous Spectrum Light on Morphological Traits and Leaf Anatomy of Hazelnut Plantlets. Front. Plant Sci. 2019, 10, 1318. [Google Scholar] [CrossRef]
- Ballaré, C.L. Light regulation of plant defence. Annu. Rev. Plant Biol. 2014, 65, 335–363. [Google Scholar] [CrossRef] [PubMed]
- Neu, E.; Domes, H.S.; Menz, I.; Kaufmann, H.; Linde, M.; Debener, T. Interaction of roses with a biotrophic and a hemibiotrophic leaf pathogen leads to differences in defense transcriptome activation. Plant Mol. Biol. 2019, 99, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Wojdyła, A.T. Influence of some compounds on development of Sphaerotheca pannosa var. rosae. J. Plant Prot. Res. 2000, 40, 106–121. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LED Light Treatments | UV-A 340–400 nm | Blue 400–500 nm | Green-Yellow 500–600 nm | Red 600–700 nm | Far-Red 700–780 nm | R:FR Ratio |
|---|---|---|---|---|---|---|
| Control | 3.9 | 22.1 | 28.0 | 27.4 | 16.8 | 1.6 |
| R—Red | - | - | - | 100.0 | - | - |
| B—Blue | - | 100.0 | - | - | - | - |
| W—White | 0.1 | 24.4 | 44.4 | 28.9 | 2.2 | 13.1 |
| S1—R + B + W + FR high | 0.3 | 12.9 | 14.5 | 66.5 | 5.9 | 11.3 |
| S2—R + B + W + FR low | 0.2 | 16.3 | 10.3 | 49.4 | 23.8 | 2.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matysiak, B. The Effect of Supplementary LED Lighting on the Morphological and Physiological Traits of Miniature Rosa × Hybrida ‘Aga’ and the Development of Powdery Mildew (Podosphaera pannosa) under Greenhouse Conditions. Plants 2021, 10, 417. https://doi.org/10.3390/plants10020417
Matysiak B. The Effect of Supplementary LED Lighting on the Morphological and Physiological Traits of Miniature Rosa × Hybrida ‘Aga’ and the Development of Powdery Mildew (Podosphaera pannosa) under Greenhouse Conditions. Plants. 2021; 10(2):417. https://doi.org/10.3390/plants10020417
Chicago/Turabian StyleMatysiak, Bożena. 2021. "The Effect of Supplementary LED Lighting on the Morphological and Physiological Traits of Miniature Rosa × Hybrida ‘Aga’ and the Development of Powdery Mildew (Podosphaera pannosa) under Greenhouse Conditions" Plants 10, no. 2: 417. https://doi.org/10.3390/plants10020417
APA StyleMatysiak, B. (2021). The Effect of Supplementary LED Lighting on the Morphological and Physiological Traits of Miniature Rosa × Hybrida ‘Aga’ and the Development of Powdery Mildew (Podosphaera pannosa) under Greenhouse Conditions. Plants, 10(2), 417. https://doi.org/10.3390/plants10020417