
Establishment of a Novel and Efficient Agrobacterium-Mediated in Planta Transformation System for Passion Fruit (Passiflora edulis)

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Influences of Dark Incubation and Parafilm Wrapping on Shoot Regeneration Efficiency
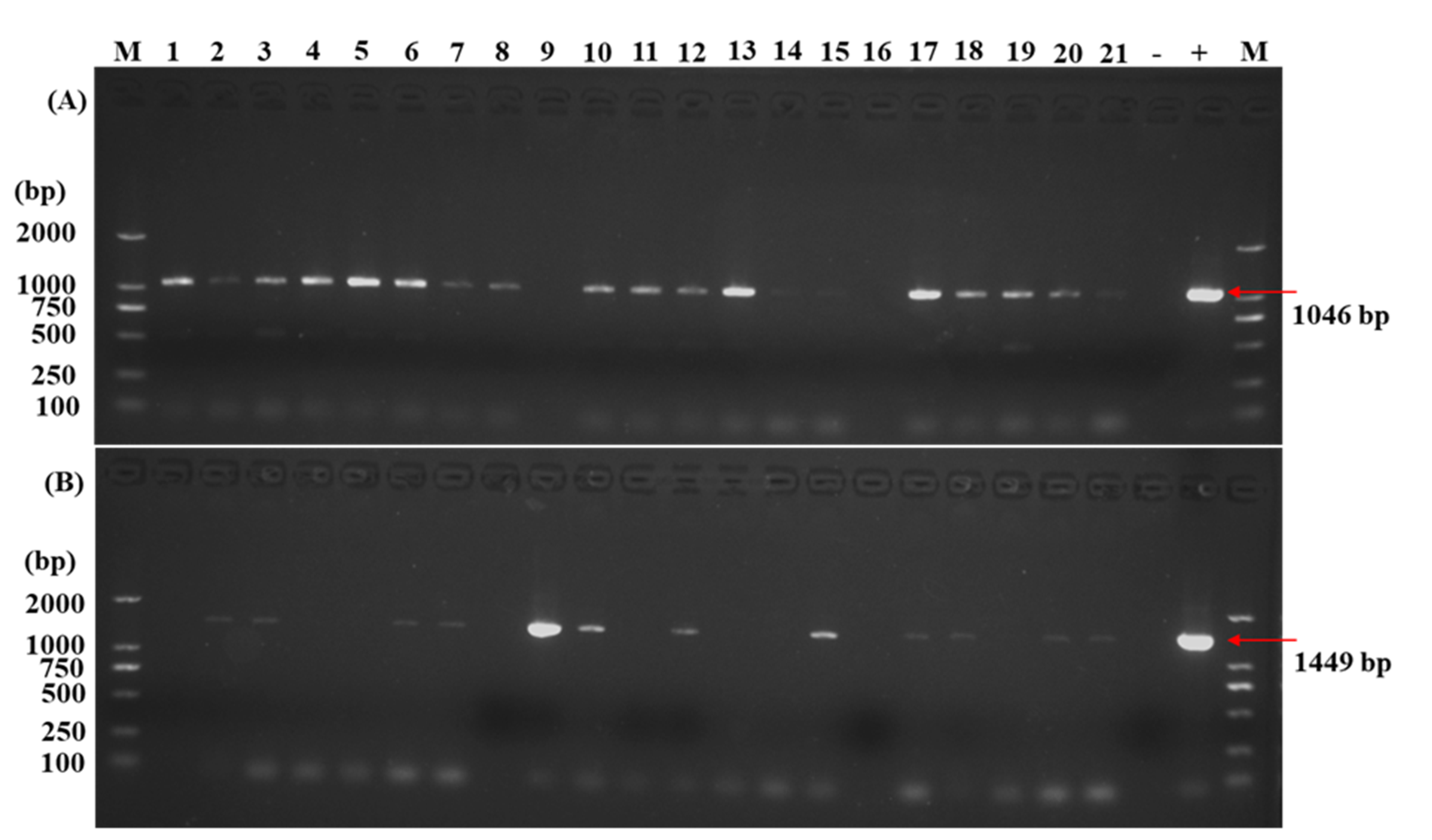
2.2. Molecular Identification of Transgenic Plants
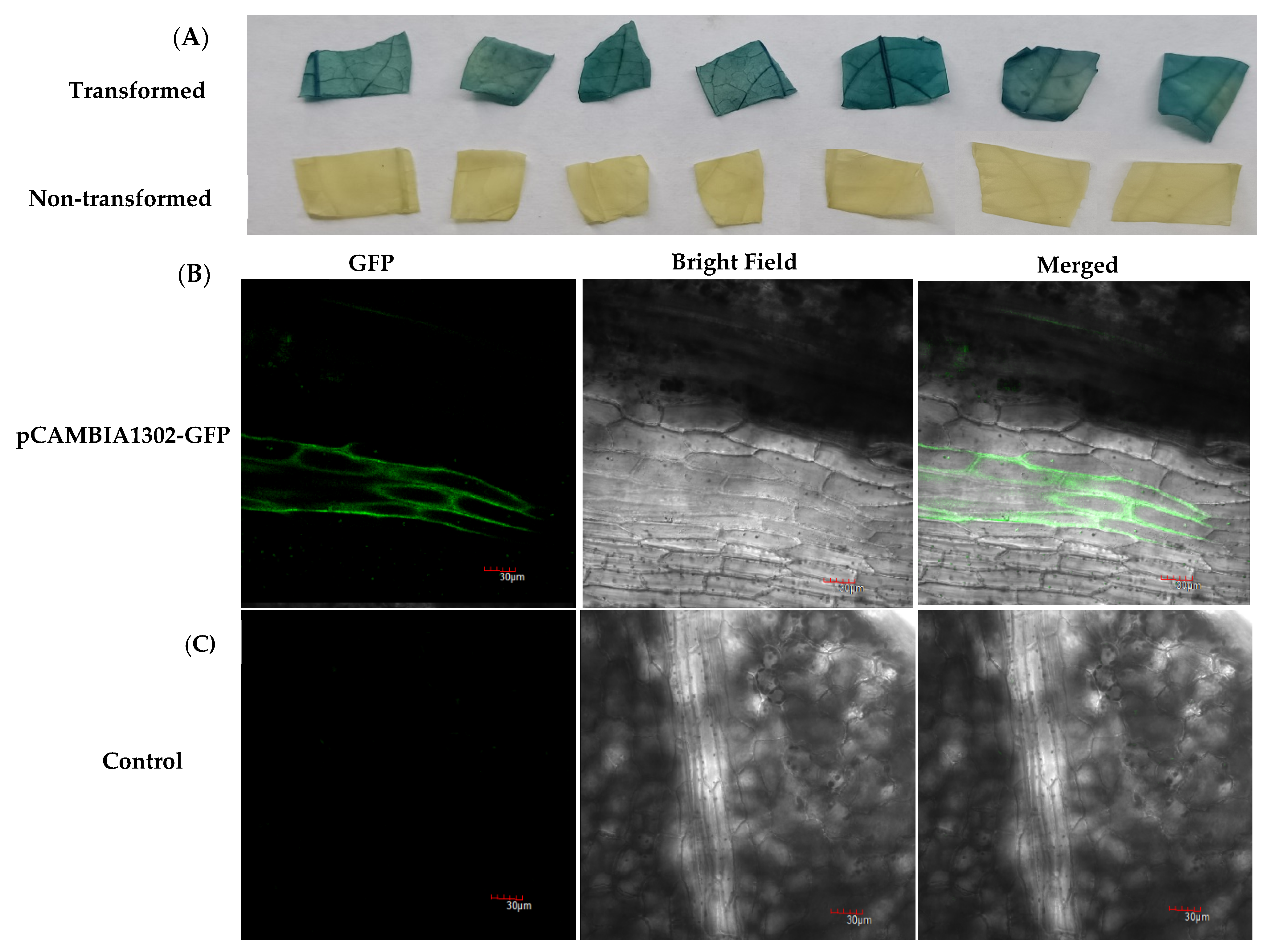
2.3. GUS Staining Assays and Visualization of GFP Fluorescence in Transgenic Plants
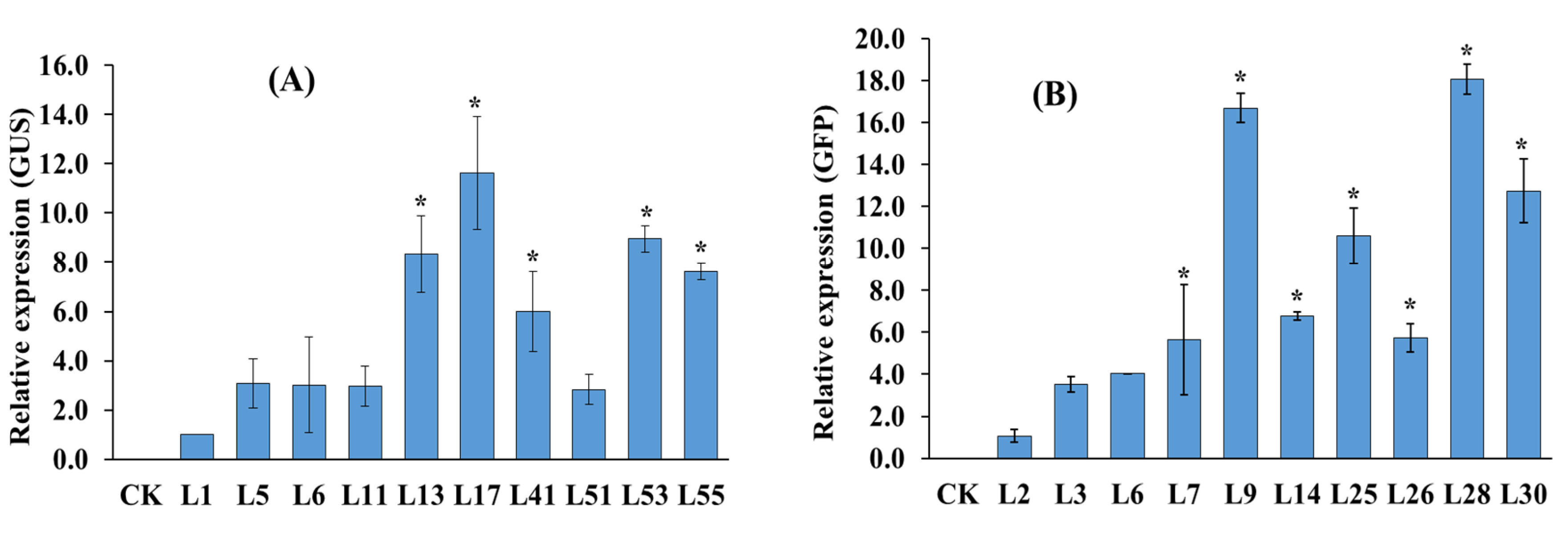
2.4. Expression Analysis of the GUS and GFP Genes by Quantitative Real-Time PCR
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Agrobacterium Strain and Binary Vector
4.3. Agrobacterium Culture Preparation
4.4. Experiment on Dark Incubation
4.5. In Planta Transformation of Passion Fruit Seedlings
4.6. Identification of Putative Transgenic Plant Lines
4.6.1. GUS Histochemical Staining Assay
4.6.2. GFP Fluorescence Visualization
4.6.3. Isolation of Genomic DNA and PCR Analysis for Transgenic Lines
4.7. RNA Extraction for qRT-PCR Analysis
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qiu, W.; Su, W.; Cai, Z.; Dong, L.; Li, C.; Xin, M.; Fang, W.; Liu, Y.; Wang, X.; Huang, Z. Combined analysis of transcriptome and metabolome reveals the potential mechanism of coloration and fruit quality in yellow and purple Passiflora edulis Sims. J. Agric. Food Chem. 2020, 68, 12096–12106. [Google Scholar] [CrossRef]
- Ramaiya, S.D.; Bujang, J.S.; Zakaria, M.H.; King, W.S.; Sahrir, M.A.S. Sugars, ascorbic acid, total phenolic content and total antioxidant activity in passion fruit (Passiflora) cultivars. J. Sci. Food Agric. 2013, 93, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Luan, F.; Yang, Y.; Wang, Z.; Zhao, Z.; Fang, J.; Wang, M.; Zuo, M.; Li, Y. Passiflora edulis: An insight into current researches on phytochemistry and pharmacology. Front. Pharmacol. 2020, 11, 617. [Google Scholar] [CrossRef]
- Zhang, R.; Lan, W.; Ding, J.; Ahmed, S.; Qin, W.; He, L.; Liu, Y. Effect of PLA/PBAT antibacterial film on storage quality of passion fruit during the shelf-life. Molecules 2019, 24, 3378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieczorek, A.; Wright, M.G. History of agricultural biotechnology: How crop development has evolved. Nat. Educ. Knowl. 2012, 3, 9. [Google Scholar]
- Gill, S.S.; Gill, R.; Tuteja, R.; Tuteja, N. Genetic engineering of crops: A ray of hope for enhanced food security. Plant Signal. Behav. 2014, 9, e28545. [Google Scholar] [CrossRef] [Green Version]
- Phillips, T. Genetically modified organisms (GMOs): Transgenic crops and recombinant DNA technology. Nat. Educ. 2008, 1, 213. [Google Scholar]
- Parmar, N.; Singh, K.H.; Sharma, D.; Singh, L.; Kumar, P.; Nanjundan, J.; Khan, Y.J.; Chauhan, D.K.; Thakur, A.K. Genetic engineering strategies for biotic and abiotic stress tolerance and quality enhancement in horticultural crops: A comprehensive review. 3 Biotech 2017, 7, 1–35. [Google Scholar] [CrossRef]
- Asande, L.K.; Omwoyo, R.O.; Oduor, R.O.; Nyaboga, E.N. A simple and fast Agrobacterium-mediated transformation system for passion fruit KPF4 (Passiflora edulis f. edulis× Passiflora edulis f. flavicarpa). Plant Methods 2020, 16, 1–12. [Google Scholar] [CrossRef]
- Manders, G.; Otoni, W.; Vaz, F.d.U.; Blackball, N.; Power, J.; Davey, M. Transformation of passionfruit (Passiflora edulis fv flavicarpa Degener.) using Agrobacterium tumefaciens. Plant Cell Rep. 1994, 13, 697–702. [Google Scholar] [CrossRef]
- Trevisan, F.; Mendes, B.; Maciel, S.; Vieira, M.; Meletti, L.; Rezende, J. Resistance to Passion fruit woodiness virus in transgenic passionflower expressing the virus coat protein gene. Plant Dis. 2006, 90, 1026–1030. [Google Scholar] [CrossRef]
- Monteiro-Hara, A.C.; Jadao, A.S.; Mendes, B.M.; Rezende, J.A.; Trevisan, F.; Mello, A.P.O.; Vieira, M.L.C.; Meletti, L.; Piedade, S.M.D.S. Genetic transformation of passionflower and evaluation of R1 and R2 generations for resistance to Cowpea aphid borne mosaic virus. Plant Dis. 2011, 95, 1021–1025. [Google Scholar] [CrossRef] [Green Version]
- Correa, M.F.; Pinto, A.P.C.; Rezende, J.A.M.; Harakava, R.; Mendes, B.M.J. Genetic transformation of sweet passion fruit (Passiflora alata) and reactions of the transgenic plants to Cowpea aphid borne mosaic virus. Eur. J. Plant Pathol. 2015, 143, 813–821. [Google Scholar] [CrossRef]
- Bunnag, S.; Chamnanpon, N. Regeneration and transformation of purple passion fruit (Passiflora edulis S.) via Agrobacterium tumefaciens. In Proceedings of the XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes (IHC2014): 1110, Brisbane, Australia, 17–22 August 2014; pp. 215–222. [Google Scholar]
- Tuhaise, S.; Nakavuma, J.L.; Adriko, J.; Ssekatawa, K.; Kiggundu, A. Establishment of a transformation protocol for Ugandas yellow passion fruit using the GUS gene. Afr. J. Biotechnol. 2019, 18, 416–425. [Google Scholar]
- da Silva, M.L.; Pinto, D.L.P.; Passos, A.B.; Marcelino-Guimarães, F.C.; Rossi, A.A.B.; Krause, W.; de Carvalho, I.F.; Batista, D.S.; Rocha, D.I.; Otoni, W.C. Novel and efficient transformation of wild passion fruit (Passiflora cincinnata Mast.) using sonication-assisted Agrobacterium-mediated transformation. Vitr. Cell. Dev. Biol.–Plant 2021, 57, 380–386. [Google Scholar] [CrossRef]
- Pathi, K.M.; Tula, S.; Tuteja, N. High frequency regeneration via direct somatic embryogenesis and efficient Agrobacterium-mediated genetic transformation of tobacco. Plant Signal. Behav. 2013, 8, e24354. [Google Scholar] [CrossRef] [Green Version]
- Ozdemir, B.S.; Budak, H. Application of tissue culture and transformation techniques in model species Brachypodium distachyon. In Brachypodium Genomics; Springer: Berlin, Germany, 2018; pp. 289–310. [Google Scholar]
- Tang, W. Additional virulence genes and sonication enhance Agrobacterium tumefaciens-mediated loblolly pine transformation. Plant Cell Rep. 2003, 21, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Nalapalli, S.; Tunc-Ozdemir, M.; Sun, Y.; Elumalai, S.; Que, Q. Morphogenic Regulators and Their Application in Improving Plant Transformation. In Rice Genome Engineering and Gene Editing; Springer: Berlin, Germany, 2021; pp. 37–61. [Google Scholar]
- Mishutkina, Y.V.; Gaponenko, A. Sugar beet (Beta vulgaris L.) morphogenesis in vitro: Effects of phytohormone type and concentration in the culture medium, type of explants, and plant genotype on shoot regeneration frequency. Russ. J. Genet. 2006, 42, 150–157. [Google Scholar] [CrossRef]
- Perrier, A.; Barberis, P.; Genin, S. Introduction of genetic material in Ralstonia solanacearum through natural transformation and conjugation. In Host-Pathogen Interactions; Springer: Berlin, Germany, 2018; pp. 201–207. [Google Scholar]
- Maggini, V.; Bettini, P.; Firenzuoli, F.; Bogani, P. An Efficient Method for the Genetic Transformation of Acmella oleracea L.(Spilanthes acmella Linn.) with Agrobacterium tumefaciens. Plants 2021, 10, 198. [Google Scholar] [CrossRef]
- Eibl, R.; Meier, P.; Stutz, I.; Schildberger, D.; Hühn, T.; Eibl, D. Plant cell culture technology in the cosmetics and food industries: Current state and future trends. Appl. Microbiol. Biotechnol. 2018, 102, 8661–8675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leva, A.; Petruccelli, R.; Rinaldi, L. Somaclonal variation in tissue culture: A case study with olive. Recent Adv. Plant Vitr. Cult. 2012, 7, 123–150. [Google Scholar]
- Islas-Osuna, M.; Tiznado-Hernández, M. Biotechnology and molecular biology of tropical and subtropical fruits. In Postharvest Biology and Technology of Tropical and Subtropical Fruits; Elsevier: Amsterdam, The Netherlands, 2011; pp. 315–380. [Google Scholar]
- Tuhaise, S.; Nakavuma, J.L.; Adriko, J.; Ssekatawa, K.; Kiggundu, A. In vitro regeneration of Ugandan passion fruit cultivars from leaf discs. BMC Res. Notes 2019, 12, 1–7. [Google Scholar] [CrossRef]
- Rizwan, H.M.; Irshad, M.; He, B.; Liu, S.; Lu, X.; Sun, Y.; Qiu, D. Role of reduced nitrogen for induction of embryogenic callus induction and regeneration of plantlets in Abelmoschus esculentus L. S. Afr. J. Bot. 2020, 130, 300–307. [Google Scholar] [CrossRef]
- Hwang, H.-H.; Yu, M.; Lai, E.-M. Agrobacterium-mediated plant transformation: Biology and applications. Arab. Book 2017, 15, e0186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.S.; Park, S.K.; Kim, B.C.; Kang, B.J.; Kim, D.U.; Nam, H.G. Stable genetic transformation of Arabidopsis thaliana by Agrobacterium inoculation in planta. Plant J. 1994, 5, 551–558. [Google Scholar] [CrossRef]
- Weigel, D.; Glazebrook, J. In planta transformation of Arabidopsis. CSH Protoc. 2006, 2006, 107–120. [Google Scholar] [CrossRef]
- Singh, P.; Kumar, K. Agrobacterium-mediated In-planta transformation of bread wheat (Triticum aestivum L.). J. Plant Biochem. Biotechnol. 2021, 3, 1–7. [Google Scholar]
- Basavaraju, S.N.; Lakshmikanth, R.Y.; Makarla, U. A modified in-planta transformation technique to generate stable gain-in function transformants in a recalcitrant indica rice genotype. Plant Physiol. Rep. 2020, 25, 231–244. [Google Scholar] [CrossRef]
- Zhang, Y.-y.; Zhang, D.-m.; Zhong, Y.; Chang, X.-j.; Hu, M.-l.; Cheng, C.-z. A simple and efficient in planta transformation method for pommelo (Citrus maxima) using Agrobacterium tumefaciens. Sci. Hortic. 2017, 214, 174–179. [Google Scholar] [CrossRef]
- Weeks, J.T.; Ye, J.; Rommens, C.M. Development of an in planta method for transformation of alfalfa (Medicago sativa). Transgenic Res. 2008, 17, 587–597. [Google Scholar] [CrossRef]
- Aslam, S.; Khan, S.H.; Ahmed, A.; Dandekar, A.M. The tale of cotton plant: From wild type to domestication, leading to its improvement by genetic transformation. Am. J. Mol. Biol. 2020, 10, 91–127. [Google Scholar] [CrossRef] [Green Version]
- Supartana, P.; Shimizu, T.; Nogawa, M.; Shioiri, H.; Nakajima, T.; Haramoto, N.; Nozue, M.; Kojima, M. Development of simple and efficient in planta transformation method for wheat (Triticum aestivum L.) using Agrobacterium tumefaciens. J. Biosci. Bioeng. 2006, 102, 162–170. [Google Scholar] [CrossRef]
- Yu, G.-R.; Yan, L.; Du, W.-P.; Jun, S.; Min, L.; Xu, L.-Y.; Xiao, F.-M.; Liu, Y.-S. Optimization of Agrobacterium tumefaciens-mediated immature embryo transformation system and transformation of glyphosate-resistant gene 2mG2-EPSPS in maize (Zea mays L.). J. Integr. Agric. 2013, 12, 2134–2142. [Google Scholar] [CrossRef]
- Marutani-Hert, M.; Bowman, K.D.; McCollum, G.T.; Mirkov, T.E.; Evens, T.J.; Niedz, R.P. A dark incubation period is important for Agrobacterium-mediated transformation of mature internode explants of sweet orange, grapefruit, citron, and a citrange rootstock. PLoS ONE 2012, 7, e47426. [Google Scholar]
- Yong, W.T.L.; Henry, E.S.; Abdullah, J.O. Enhancers of Agrobacterium-mediated Transformation of Tibouchina semidecandra Selected on the Basis of GFP Expression. Trop. Life Sci. Res. 2010, 21, 115. [Google Scholar]
- Que, Q.; Elumalai, S.; Li, X.; Zhong, H.; Nalapalli, S.; Schweiner, M.; Fei, X.; Nuccio, M.; Kelliher, T.; Gu, W. Maize transformation technology development for commercial event generation. Front. Plant Sci. 2014, 5, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuhaise, S. Establishment of a Transformation and Regeneration Protocol for Uganda’s Passion Fruit Using Gus Reporter Gene. Ph.D. Thesis, Makerere University, Kampala, Uganda, 2014. [Google Scholar]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In vitro plant tissue culture: Means for production of biological active compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyadarshi, S.; Sen, S. A revised scheme for mass propagation of Easter Lily. Plant Cell Tiss. Org. Cult. 1992, 30, 193–197. [Google Scholar] [CrossRef]
- Arzate-Fernandez, A.-M.; Nakazaki, T.; Okumoto, Y.; Tanisaka, T. Efficient callus induction and plant regeneration from filaments with anther in lily (Lilium longiflorum Thunb.). Plant Cell Rep. 1997, 16, 836–840. [Google Scholar] [CrossRef] [PubMed]
- Ramessar, K.; Peremarti, A.; Gómez-Galera, S.; Naqvi, S.; Moralejo, M.; Munoz, P.; Capell, T.; Christou, P. Biosafety and risk assessment framework for selectable marker genes in transgenic crop plants: A case of the science not supporting the politics. Transgenic Res. 2007, 16, 261–280. [Google Scholar] [CrossRef] [PubMed]
- Cubero, J.; Lastra, B.; Salcedo, C.; Piquer, J.; López, M.M. Systemic movement of Agrobacterium tumefaciens in several plant species. J. Appl. Microbiol. 2006, 101, 412–421. [Google Scholar] [CrossRef]
- Corte, E.-D.; Mahmoud, L.M.; Moraes, T.S.; Mou, Z.; Grosser, J.W.; Dutt, M. Development of improved fruit, vegetable, and ornamental crops using the CRISPR/Cas9 genome editing technique. Plants 2019, 8, 601. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, K.; Zeisler-Diehl, V.V.; Schreiber, L.; Bi, Y.-M.; Rothstein, S.J.; Ranathunge, K. Overexpression of ANAC046 promotes suberin biosynthesis in roots of arabidopsis Thaliana. Int. J. Mol. Sci. 2019, 20, 6117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodhi, M.A.; Ye, G.-N.; Weeden, N.F.; Reisch, B.I. A simple and efficient method for DNA extraction from grapevine cultivars andVitis species. Plant Mol. Biol. Report. 1994, 12, 6–13. [Google Scholar] [CrossRef]
- Munhoz, C.; Santos, A.; Arenhart, R.; Santini, L.; Monteiro-Vitorello, C.; Vieira, M. Analysis of plant gene expression during passion fruit–Xanthomonas axonopodis interaction implicates lipoxygenase 2 in host defence. Ann. Appl. Biol. 2015, 167, 135–155. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C T method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | With Parafilm | Without Parafilm | Mean Factor B (Dark Incubation) | |
|---|---|---|---|---|
| Dark incubation periods | 0 day | 37.33 ± 3.89 d | 9.33 ± 4.42 e | 23.33 ± 15.39 d |
| 5 days | 47.33 ± 9.04 c | 17.33 ± 4.90 e | 32.33 ± 17.57 c | |
| 10 days | 70.67 ± 6.46 b | 36.00 ± 4.42 d | 53.33 ± 19.18 b | |
| 15 days | 90.00 ± 5.96 a | 41.33 ± 5.42 c d | 65.67 ± 26.34 a | |
| 20 days | 87.33 ± 6.46 a | 48.67 ± 6.86 c | 68.00 ± 21.55 a | |
| Mean Factor A (Parafilm) | 66.53 ± 22.53 a | 30.53 ± 16.06 b | ||
| LSD Factor A (p ≤ 0.05) | 7.025 | |||
| LSD Factor B (p ≤ 0.05) | 5.832 | |||
| LSD Interaction A × B (p ≤ 0.05) | 8.248 | |||
| No. of Plants | % | |
|---|---|---|
| Regeneration efficiency a | 215/250 | 86.0 ± 4.0 |
| Overall transformation efficiency b | 62/215 | 28.8 ± 4.6 |
| GUS assays efficiency c | 36/44 | 81.7 ± 5.4 |
| GFP fluorescence efficiency d | 10/18 | 56.7 ± 8.2 |
| Single bud rate e | 47/62 | 74.4 ± 8.8 |
| Multiple bud rate f | 16/62 | 25.7 ± 4.5 |
| PCR GUS-positive efficiency g | 44/110 | 40.0 ± 5.0 |
| PCR GFP-positive efficiency h | 18/105 | 17.1 ± 5.9 |
| Cultivars | Vector and Agrobacterium Strain | EXPLANTS | Methods | Efficiency (%) | References |
|---|---|---|---|---|---|
| Passiflora edulis Sims. f. flavicarpa. IAC-275 and IAC-277. | pCAMBIA2300; EHA105 | Leaf disks | In vitro regeneration and transformation | 0.11–0.21 | [11] |
| Passiflora edulis Sims. f. flavicarpa, IAC-275 and IAC-277. | pCAMBIA2300; EHA105 | Leaf disks | In vitro regeneration and transformation | 0.19–0.67 | [12] |
| Passiflora alata | pCABMVdsCP; EHA105 | Hypocotyls | In vitro regeneration and transformation | 0.89 | [13] |
| Passiflora edulis f. flavicarpa. | pCAMBIA2301; JM109 | Leaf disks | In vitro regeneration and transformation | 0.456 | [15] |
| Passiflora edulis f. edulis × Passiflora edulis f. flavicarpa = KPF4. | pCAMBIA1301; LBA4404 | Leaf disks | In vitro regeneration and transformation | 0.67 | [9] |
| Passiflora cincinnata Mast. | pCAMBIA1304; LB4404 | Anthers | In vitro somatic embryogenesis and sonication-assisted transformation | 28.26 | [16] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizwan, H.M.; Yang, Q.; Yousef, A.F.; Zhang, X.; Sharif, Y.; Kaijie, J.; Shi, M.; Li, H.; Munir, N.; Yang, X.; et al. Establishment of a Novel and Efficient Agrobacterium-Mediated in Planta Transformation System for Passion Fruit (Passiflora edulis). Plants 2021, 10, 2459. https://doi.org/10.3390/plants10112459
Rizwan HM, Yang Q, Yousef AF, Zhang X, Sharif Y, Kaijie J, Shi M, Li H, Munir N, Yang X, et al. Establishment of a Novel and Efficient Agrobacterium-Mediated in Planta Transformation System for Passion Fruit (Passiflora edulis). Plants. 2021; 10(11):2459. https://doi.org/10.3390/plants10112459
Chicago/Turabian StyleRizwan, Hafiz Muhammad, Qiang Yang, Ahmed Fathy Yousef, Xiaoxue Zhang, Yasir Sharif, Jia Kaijie, Meng Shi, Han Li, Nigarish Munir, Xuelian Yang, and et al. 2021. "Establishment of a Novel and Efficient Agrobacterium-Mediated in Planta Transformation System for Passion Fruit (Passiflora edulis)" Plants 10, no. 11: 2459. https://doi.org/10.3390/plants10112459
APA StyleRizwan, H. M., Yang, Q., Yousef, A. F., Zhang, X., Sharif, Y., Kaijie, J., Shi, M., Li, H., Munir, N., Yang, X., Wei, X., Oelmüller, R., Cheng, C., & Chen, F. (2021). Establishment of a Novel and Efficient Agrobacterium-Mediated in Planta Transformation System for Passion Fruit (Passiflora edulis). Plants, 10(11), 2459. https://doi.org/10.3390/plants10112459