
Monoseeding Increases Peanut (Arachis hypogaea L.) Yield by Regulating Shade-Avoidance Responses and Population Density



Abstract
:1. Introduction
2. Results
2.1. Plant Growth and Development
2.2. Chlorophyll Content and Net Photosynthetic Rate
2.3. Dry Matter Accumulation
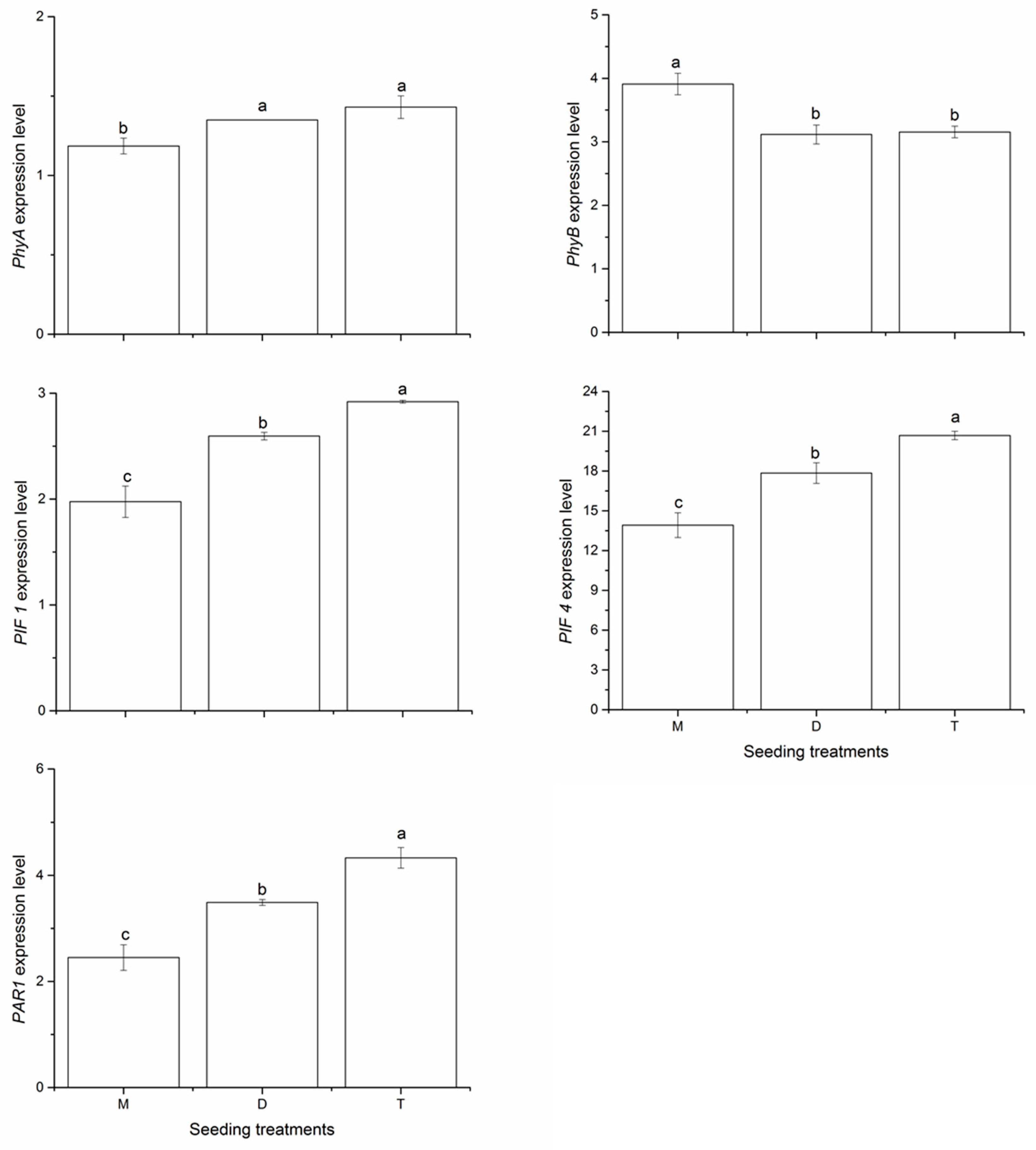
2.4. Expression of Shade-Avoidance Response Genes
2.5. Yield and Yield Components
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Data Collection
4.2.1. Plant Traits
4.2.2. Dry-Matter Accumulation
4.2.3. Chlorophyll Content and Photosynthetic Parameters
4.2.4. RNA Extraction and qRT-PCR Analysis
4.2.5. Peanut Yield and Yield Components
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boccalandro, H.E.; Ploschuk, E.L.; Yanovsky, M.J.; Sánchez, R.A.; Gatz, C.; Casal, J.J. Increased phytochrome B alleviates density effects on tuber yield of field potato crops. Plant. Physiol. 2003, 133, 1539–1546. [Google Scholar] [CrossRef] [Green Version]
- USDA-ERS. Recent Trends in GE Adoption. Available online: https://www.ers.usda.gov/data-products/adoption-of-genetically-engineered-crops-in-the-us/recent-trends-in-ge-adoption/ (accessed on 8 February 2020).
- Adams, C.; Thapa, S.; Kimura, E. Determination of a plant population density threshold for optimizing cotton lint yield: A synthesis. Field Crop. Res. 2019, 230, 11–16. [Google Scholar] [CrossRef]
- Schmitt, H.R.; Donley, J.L.; Antonucci, R.R.; Hutchings, J.B.; Kinney, A.L.; Pringle, J.E. A hubble space telescope survey of extended [O lll] 5007 A emission in a far-infreaed-selected samples of seyfert galaxies: Results. Astrophys. J. 2003, 597, 768–779. [Google Scholar] [CrossRef] [Green Version]
- Pantazopoulou, C.K.; Bongers, F.J.; Pierik, R. Reducing shade avoidance can improve Arabidopsis canopy performance against competitors. Plant. Cell Environ. 2021, 44, 1130–1141. [Google Scholar] [CrossRef] [PubMed]
- Ballaré, C.L.; Casal, J.J. Light signals perceived by crop and weed plants. Field Crop. Res. 2000, 67, 149–160. [Google Scholar] [CrossRef]
- Huché-Thélier, L.; Lanrent, C.; Jose, L.G.; Philippe, M.; Soulaiman, S.; Nathalie, L. Light signaling and plant responses to blue and UV radiations-Perspectives for applications in horticulture. Environ. Exp. Bot. 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Smith, H.; Whitelam, G.C. The shade avoidance syndrome: Multiple responses mediated by multiple phytochromes. Plant. Cell Environ. 1997, 20, 840–844. [Google Scholar] [CrossRef] [Green Version]
- Fraser, D.P.; Hayes, S.; Franklin, K.A. Photoreceptor crosstalk in shade avoidance. Curr. Opin. Plant. Biol. 2016, 33, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gommers, C.M.M.; Visser, R.J.W.; Onge, K.R.S.; Voesenek, L.A.C.; Pierik, R. Shade tolerance: When growing tall is not an option. Trend Plant. Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H.C.; Ravi, K.; Aashish, R.; Julie, M.P.; Brad, T.T.; Yasunori, I.; Ciera, C.M.; Kristina, Z.; John, J.H.; Maloof, J.N.; et al. Light-Induced Indeterminacy Alters Shade-Avoiding Tomato Leaf Morphology. Plant. Physiol. 2015, 169, 2030–2047. [Google Scholar] [CrossRef] [Green Version]
- Page, W.M.; Scott, A.F.; Kevin, L.C.; John, E.M.; Willam, L.R. Opportunities to improve adaptability and yield in grasses: Lessons from sorghum. Crop. Sci. 2002, 42, 1791–1799. [Google Scholar]
- Kroon, H.D.; Huber, H.; Stuefer, J.F.; Groenendael, J.M.V. A modular concept of phenotypic plasticity in plants. New Phytol. 2005, 166, 73–82. [Google Scholar] [CrossRef]
- Iwabe, R.; Koyama, K.; Komamura, R. Shade Avoidance and Light Foraging of a Clonal Woody Species, Pachysandra terminalis. Plants 2021, 10, 809. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, F.; Pierik, R.; Millenaar, F.F.; Voesenek, L.A.; Straeten, D.V.D. Reaching out of the shade. Curr. Opin. Plant. Biol. 2005, 8, 462–468. [Google Scholar] [CrossRef]
- Zhang, Y.; Mayba, O.; Pfeiffer, A.; Shi, H.; Tepperman, J.M.; Speed, T.P.; Quail, P.H. A quartet of PIF bHLH factors provides a transcriptionally centered signaling hub that regulates seedling morphogenesis through differential expression patterning of shared target genes in Arabidopsis. PLoS Genet. 2013, 9, e1003244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carriedo, L.G.; Maloof, J.N.; Brady, S.M. Molecular control of crop shade avoidance. Curr. Opin. Plant. Biol. 2016, 30, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Dai, J.; Zhang, Y.; Kong, X.; Li, C.; Dong, H. Topical shading substantially inhibits vegetative branching by altering leaf photosynthesis and hormone contents of cotton plants. Field Crop. Res. 2019, 238, 18–26. [Google Scholar] [CrossRef]
- Ding, Z.; Zhang, Y.; Xiao, Y.; Liu, F.; Wang, M.; Zhu, X.; Liu, P.; Sun, Q.; Wang, W.; Peng, M.; et al. Transcriptome response of cassava leaves under natural shade. Sci. Rep. 2016, 6, 31673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, D.E. Horicultural requirements for mechanical pepper harvesting. Proc. 1st Int. Conf. on Fruit, Nut and Vegetable Harvesting Mechanization, Bet Dagan, Israel. Am. Soc. Agric. Eng. Publ. 1984, 389–396. Available online: https://aces.nmsu.edu/pubs/research/horticulture/CTF13/welcome.html (accessed on 8 February 2020).
- Zhang, J.L.; Geng, Y.; Guo, F.; Li, X.; Wan, S.B. Research progress on the mechanism of improving peanut yield by single-seed precision sowing. J. Integr. Agric. 2019, 18, 2–10. [Google Scholar]
- Zhang, J.L.; Guo, F.; Yang, D.Q.; Meng, J.J.; Yang, S.; Wang, X.Y.; Tao, S.X.; Li, X.G.; Wan, S.B. Effects of single-seed precision sowing on population structure and yield of peanuts with super-high yield cultivation. Sci. Agric. Sin. 2015, 48, 3757–3766. [Google Scholar]
- Liang, X.Y.; Guo, F.; Zhang, J.L.; Meng, J.J.; Li, L.; Wan, S.B.; Li, X.G. Effects of single-seed sowing on canopy microenvironment, photosynthetic characteristics and pod yield of peanut (Arachis hypogaca). Chin. J. Appl. Ecol. 2015, 26, 3700–3706. [Google Scholar]
- Deguchi, R.; Koyama, K. Photosynthetic and morphological acclimation to high and low light environments in Petasites japonicus subsp. giganteus. Forests 2020, 11, 1365. [Google Scholar] [CrossRef]
- Wan, Y.; Zhang, Y.; Zhang, M.; Hong, A.; Liu, Y. Shade effects on growth, photosynthesis and chlorophyll fluorescence parameters of three Paeonia species. Peer J. 2020, 8, e9316. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Y.; Hao, G.Y.; Guo, J.J.; Liu, Z.H.; Cao, K.F. Differentiation in leaf physiological traits related to shade and drought tolerance underlies contrasting adaptations of two Cyclobalanopsis (Fagaceae) species at the seedling stage. Forests 2020, 11, 844. [Google Scholar] [CrossRef]
- Nilsson, M.C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Liu, J.L.; Mahoney, K.J.; Sikkema, P.H.; Swanton, C.J. The importance of light quality in crop-weed competition. Weed Res. 2009, 49, 217–224. [Google Scholar] [CrossRef]
- Li, J.; Li, G.; Wang, H.; Deng, X.W. Phytochrome signaling mechanisms. Arab. Book/Am. Soc. Plant. Biol. 2011, 9, e0148. [Google Scholar] [CrossRef] [Green Version]
- Lorrain, S.; Allen, T.; Duek, P.D.; Whitelam, G.C.; Fankhause, C. Phytochrome-mediated inhibition of shade avoidance involves degradation of growth-promoting bHLH transcription factors. Plant. J. 2008, 53, 312–323. [Google Scholar] [CrossRef] [Green Version]
- Leivar, P.; Monte, E.; Oka, Y.; Liu, T.; Carle, C.; Castillon, A.; Huq, E.; Quail, P.H. Multiple phytochrome-interacting bHLH transcription factors repress premature seedling photomorphogenesis in darkness. Curr. Biol. 2008, 18, 1815–1823. [Google Scholar] [CrossRef] [Green Version]
- Ciolfi, A.; Sessa, G.; Possenti, M.; Salvucci, S.; Carabelli, M.; Morelli, G.; Ruberti, L. Dynamics of the shade-avoidance response in Arabidopsis. Plant. Physiol. 2013, 163, 331–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef]
- Franklin, K.A.; Quail, P.H. Phytochrome functions in Arabidopsis development. J. Exp. Bot. 2010, 61, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.; Bradley, M.; Harberd, N.P.; Whitelam, G.C. Photoresponses of light-grown Arabidopsis (phytochrome A is required for the day length extensions). Plant. Physiol. 1994, 105, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Song, M.; Yang, Q.; Su, L.; Hou, P.; Guo, L.; Zheng, X.; Xi, Y.; Meng, F.; Xiao, Y.; et al. Both PHYTOCHROME RAPIDLY REGULATED1 (PAR1) and PAR2 promote seedling photomorphogenesis in multiple light signaling pathways. Plant. Physiol. 2014, 164, 841–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig-Villanova, I.; Bou-Torrent, J.; Galstyan, A.; Carretero-Paulet, L.; Portolés, S.; Rodríguez-Concepción, M.; Martínez-García, J.F. Interaction of shade avoidance and auxin responses: A role for two novel atypical bHLH proteins. EMBO J. 2007, 26, 4756–4767. [Google Scholar] [CrossRef] [Green Version]
- Suprapto, A.; Sugito, Y.; Sitompul, S.M. Sudaryono Study of growth, yield and radiation energy conversion efficiency on arieties and different plant population of peanut. Procedia Environ. Sci. 2013, 17, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.L.; Li, W.J.; Tang, W.; Zhang, D.M.; Li, Z.H.; Lu, H.Q.; Eneji, A.E.; Dong, H.Z. Manipulation of dry matter accumulation and partitioning with plant density in relation to yield stability of cotton under intensive management. Field Crop. Res. 2015, 180, 207–215. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.C.M.; Ren, L.; Farmer, A.D.; Pandey, M.K.; et al. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Dang, P.; Bausher, M.G.; Holbrook, C.C.; Lee, R.D.; Lynch, R.E.; Guo, B.Z. Identification of transcripts involved in resistance responses to leaf spot disease caused by Cercosporidium personatum in peanut (Arachis hypogaea L.). Phytopathology 2005, 95, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.; Moorman, A.F. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Zhang, G.C.; Dai, L.X.; Ding, H.; Ci, D.W.; Ning, T.Y.; Yang, J.S.; Zhao, X.H.; Yu, H.Q.; Zhang, Z.M. Response and adaptation to the accumulation distribution of phtosynthetic product in peanut under salt stress. J. Integr. Agric. 2020, 19, 690–699. [Google Scholar] [CrossRef]

{kind=link}
| Years | Parameters | |||
|---|---|---|---|---|
| Main Stem Height (cm) | Main Stem Diameter (mm) | Number of Branches | Number of Nodes | |
| 2018 | 26.8 ± 10.9 a | 3.1 ± 1.2 a | 5.7 ± 2.8 a | 6.7 ± 2.6 a |
| 2019 | 25.3 ± 11.8 a | 3.1 ± 1.3 a | 5.6 ± 2.7 a | 7.9 ± 2.4 b |
| Years | Parameters | |||
|---|---|---|---|---|
| Main Stem Height (cm) | Main Stem Diameter (mm) | Number of Branches | Number of Nodes | |
| Seedling stage | 14.5 ± 2.4 d | 1.9 ± 0.2 c | 3.1 ± 1.5 c | 4.8 ± 1.4 b |
| Flowering and pegging stage | 19.8 ± 3.9 c | 2.4 ± 0.5 c | 5.0 ± 1.9 bc | 6.1 ± 1.2 b |
| Pod filling stage | 28.0 ± 3.5 b | 3.4 ± 0.7 b | 6.1 ± 2.0 ab | 8.3 ± 1.8 a |
| Maturity stage | 41.6 ± 6.5 a | 4.6 ± 0.9 a | 8.3 ± 2.5 a | 9.9 ± 1.8 a |
| Treatment | Main Stem Height (cm) | Main Stem Diameter (mm) | Number of Branches | Number of Nodes |
|---|---|---|---|---|
| M | 22.2 ± 9.3 a | 3.8 ± 1.4 a | 7.9 ± 2.5 a | 8.9 ± 2.5 a |
| D | 25.7 ± 10.9 a | 3.1 ± 1.1 ab | 5.5 ± 2.1 b | 7.1 ± 2.3 ab |
| T | 30.1 ± 13.1 a | 2.4 ± 0.8 b | 3.6 ± 1.6 b | 5.8 ± 1.8 b |
| Parameters | ||||
|---|---|---|---|---|
| Main Stem Height (cm) | Main Stem Diameter (mm) | Number of Branches | Number of Nodes | |
| Year (Y) | 49.9 ** | 0.0 ** | 0.2 NS | 10.9 ** |
| Growth stage (G) | 2464.1 ** | 17.8 ** | 114.5 ** | 83.2 ** |
| Treatment (T) | 379.1 ** | 6.9 ** | 148.8 ** | 83.0 ** |
| Y×G | 53.2 ** | 0.1 ** | 1.2 ** | 3.7 ** |
| Y×T | 0.1 NS | 0.0 ** | 4.1 ** | 2.7 NS |
| G×T | 29.1 ** | 0.5 ** | 2.0 ** | 1.0 ** |
| Y×G×T | 0.7 NS | 0.0 NS | 0.2 NS | 2.5 ** |
| Years | SPAD |
|---|---|
| 2018 | 41.2 ± 2.5 a |
| 2019 | 37.2 ± 4.0 b |
| Growth Stages | SPAD |
|---|---|
| Flowering and pegging stage | 37.4 ± 4.5 b |
| Maturity stage | 40.8 ± 2.2 a |
| Treatment | SPAD | Net Photosynthetic Rate(Pn) (μmol CO2 m−2 s−1) |
|---|---|---|
| M | 41.9 ± 2.9 a | 21.5 ± 1.2 a |
| D | 38.6 ± 3.5 b | 19.4 ± 0.7 b |
| T | 36.3 ± 3.4 b | 16.8 ± 0.7 c |
| Year | Growth Stage | Treatment | Root Dry Weight (g/Plant) | Stem and Petiole Dry Weight (g/Plant) | Leaf Dry Weight (g/Plant) | Pod Dry Weight (g/Plant) |
|---|---|---|---|---|---|---|
| 2018 | Seedling stage | M | 0.9 ± 0.0 a | 4.6 ± 0.2 a | 6.8 ± 0.4 a | - |
| D | 0.6 ± 0.0 b | 3.9 ± 0.0 b | 5.6 ± 0.4 b | - | ||
| T | 0.4 ± 0.0 c | 3.1 ± 0.3 c | 4.7 ± 0.1 c | - | ||
| Flowering and pegging stage | M | 1.0 ± 0.1 a | 7.4 ± 0.1 a | 11.6 ± 0.3 a | - | |
| D | 0.8 ± 0.0 b | 5.7 ± 0.0 b | 9.8 ± 0.2 b | - | ||
| T | 0.6 ± 0.0 c | 4.6 ± 0.4 c | 7.7 ± 0.1 c | - | ||
| Maturity stage | M | 3.3 ± 0.2 a | 16.3 ± 0.7 a | 14.1 ± 0.5 a | 44.1 ± 0.6 a | |
| D | 2.4 ± 0.2 b | 12.2 ± 0.3 b | 11.9 ± 0.2 b | 32.9 ± 3.6 b | ||
| T | 1.8 ± 0.1 c | 10.1 ± 0.7 c | 9.1 ± 0.3 c | 19.8 ± 2.5 c | ||
| 2019 | Seedling stage | M | 0.5 ± 0.0 a | 2.9 ± 0.1 a | 4.1 ± 0.2 a | - |
| D | 0.3 ± 0.0 b | 2.2 ± 0.2 b | 3.1 ± 0.1 b | - | ||
| T | 0.3 ± 0.0 c | 1.7 ± 0.0 c | 2.0 ± 0.3 c | - | ||
| Flowering and pegging stage | M | 1.3 ± 0.1 a | 7.8 ± 0.0 a | 9.3 ± 1.1 a | - | |
| D | 1.0 ± 0.0 b | 5.2 ± 0.1 b | 6.4 ± 0.2 b | - | ||
| T | 0.7 ± 0.1 c | 3.9 ± 0.0 c | 4.9 ± 0.1 c | - | ||
| Maturity stage | M | 2.9 ± 0.0 a | 18.3 ± 1.3 a | 13.6 ± 0.2 a | 43.6 ± 3.8 a | |
| D | 2.1 ± 0.1 b | 14.8 ± 0.0 b | 12.0 ± 0.3 b | 33.8 ± 0.8 b | ||
| T | 1.6 ± 0.0 c | 11.9 ± 0.1 c | 8.8 ± 0.8 c | 20.9 ± 1.3 c | ||
| Mean Square | ||||||
| Year (Y) | 0.1 ** | 0.1 NS | 32.2 ** | 0.1 NS | ||
| Growth stage (G) | 11.4 ** | 384.2 ** | 154.7 ** | - | ||
| Treatments (T) | 1.8 ** | 41.4 ** | 41.6 ** | 562.7 ** | ||
| Y × G | 0.2 ** | 10.4 ** | 6.2 ** | - | ||
| Y × T | 0.0 NS | 0.1 NS | 0.0 NS | 0.7 NS | ||
| G × T | 0.3 ** | 6.4 ** | 2.2 ** | - | ||
| Y × G × T | 0.0 NS | 0.2 NS | 0.2 NS | - | ||
| Treatment | Pods Number per Plant | 100 Pod Weight (kg) | Shelling Percentage (%) | Pod Yield (kg/hm2) |
|---|---|---|---|---|
| M | 23.61 ± 0.49 a | 0.21 ± 0.01 a | 76 ± 0.76 a | 11,683.75 ± 145.58 a |
| D | 19.15 ± 0.70 b | 0.19 ± 0.01 b | 73 ± 0.59 b | 10,277.00 ± 290.87 b |
| T | 14.52 ± 3.48 c | 0.16 ± 0.01 c | 70 ± 0.74 c | 8848.75 ± 238.05 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Zhang, J.; Wang, X.; Zeng, R.; Chen, Y.; Zhang, H.; Wan, S.; Zhang, L. Monoseeding Increases Peanut (Arachis hypogaea L.) Yield by Regulating Shade-Avoidance Responses and Population Density. Plants 2021, 10, 2405. https://doi.org/10.3390/plants10112405
Chen T, Zhang J, Wang X, Zeng R, Chen Y, Zhang H, Wan S, Zhang L. Monoseeding Increases Peanut (Arachis hypogaea L.) Yield by Regulating Shade-Avoidance Responses and Population Density. Plants. 2021; 10(11):2405. https://doi.org/10.3390/plants10112405
Chicago/Turabian StyleChen, Tingting, Jialei Zhang, Xinyue Wang, Ruier Zeng, Yong Chen, Hui Zhang, Shubo Wan, and Lei Zhang. 2021. "Monoseeding Increases Peanut (Arachis hypogaea L.) Yield by Regulating Shade-Avoidance Responses and Population Density" Plants 10, no. 11: 2405. https://doi.org/10.3390/plants10112405
APA StyleChen, T., Zhang, J., Wang, X., Zeng, R., Chen, Y., Zhang, H., Wan, S., & Zhang, L. (2021). Monoseeding Increases Peanut (Arachis hypogaea L.) Yield by Regulating Shade-Avoidance Responses and Population Density. Plants, 10(11), 2405. https://doi.org/10.3390/plants10112405