
Seasonal Variability of Photosynthetic Microbial Eukaryotes (<3 µm) in the Kara Sea Revealed by 18S rDNA Metabarcoding of Sediment Trap Fluxes



Abstract
:
1. Introduction
2. Materials and Methods
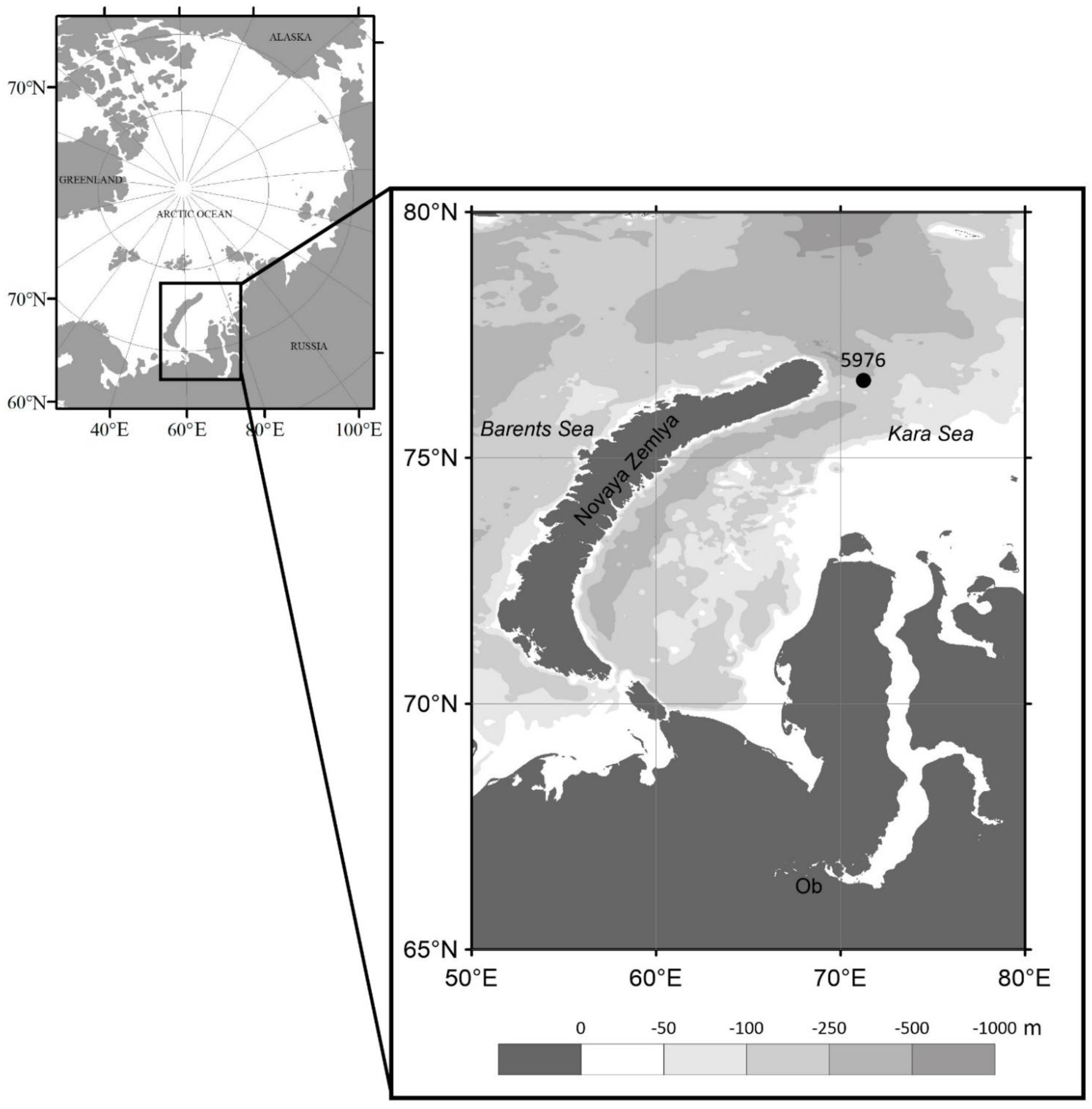
2.1. Study Area and Sampling
2.2. Picophytoplankton Enumeration
2.3. Chlorophyll a Measurement
2.4. DNA Isolation
2.5. NGS Read Processing and Phylogenetic Analysis
2.6. Statistical Analyses
3. Results
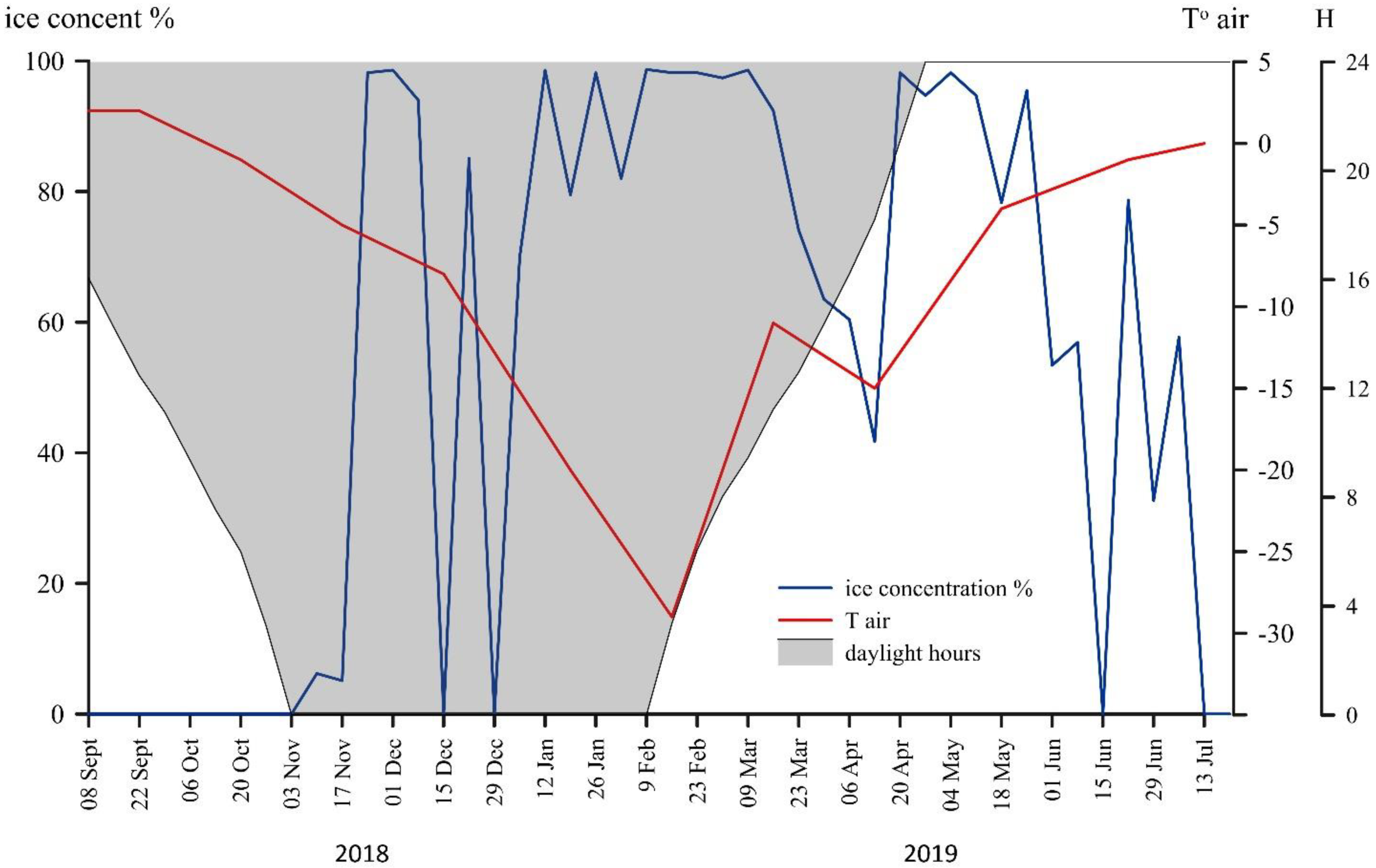
3.1. Environmental Conditions
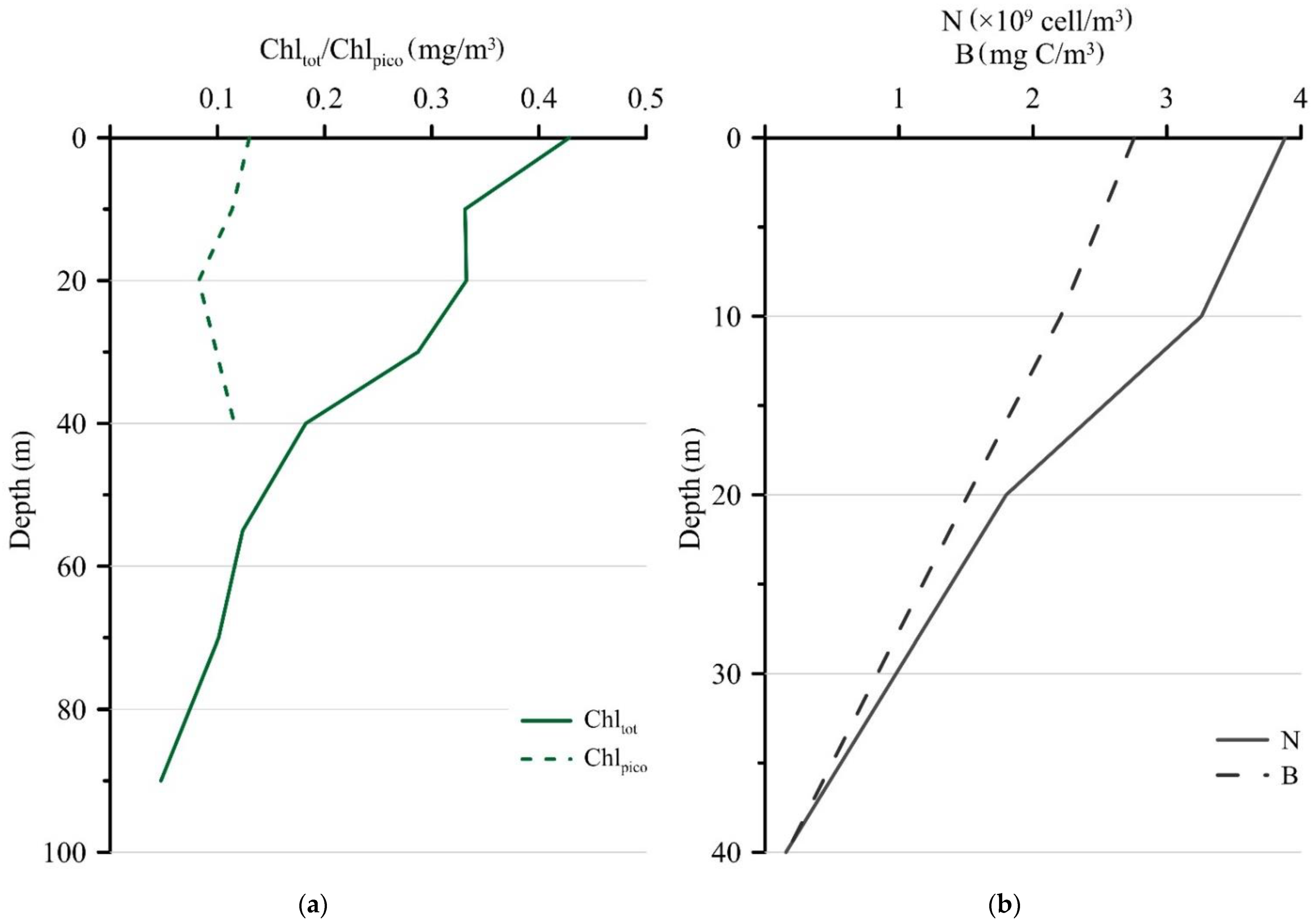
3.2. Chltot, Chlpico, Abundance, and Biomass of Photosynthetic Picoeukaryotes at Station 5976 While Setting the Sediment Trap
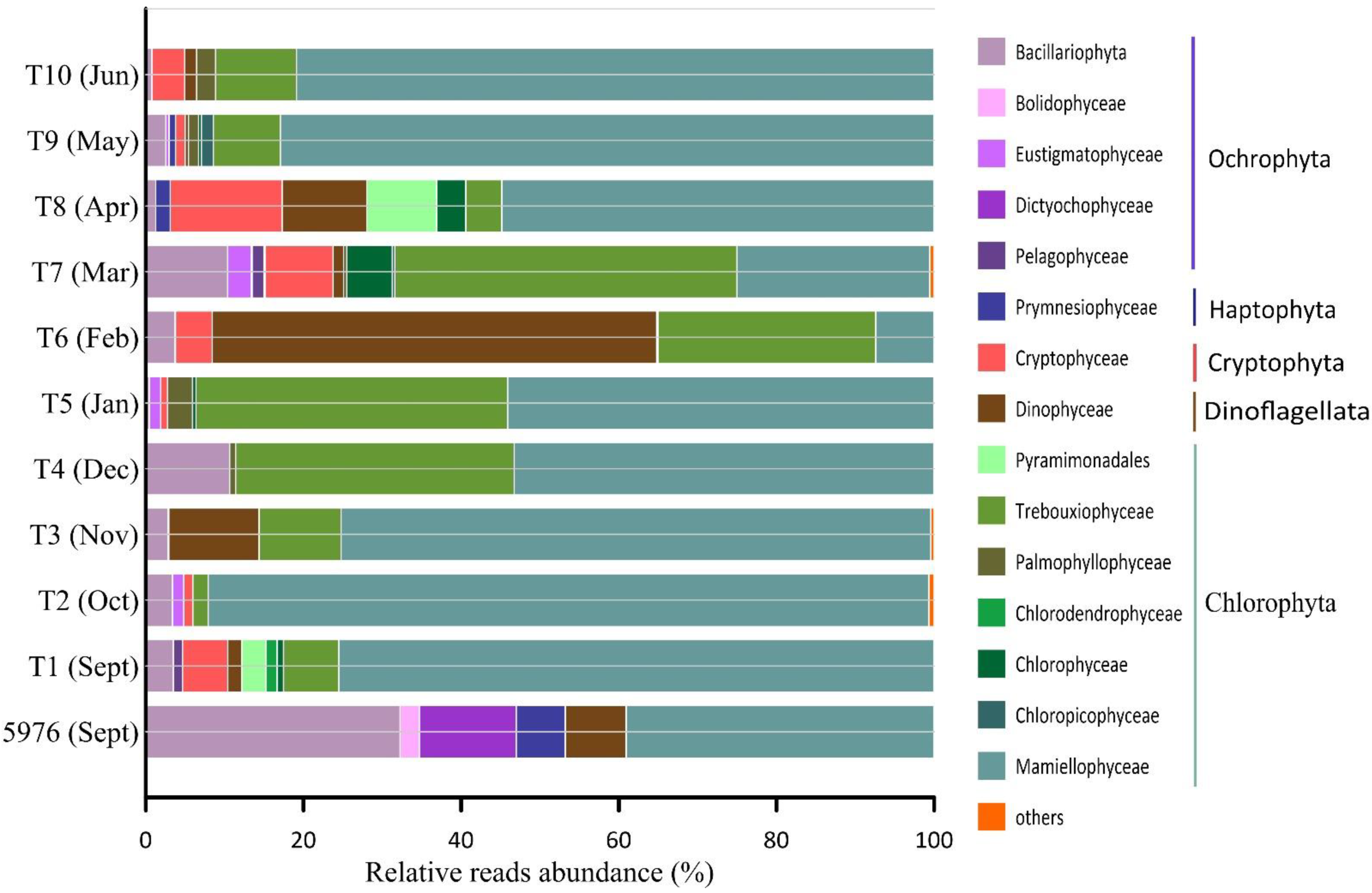
3.3. General Characteristics of Microbial Eukaryote Taxonomic Composition in Samples Filtered through a 3 µm Pore Size Filter
3.4. Variability in Pelagic PE Diversity in the Samples Filtered through a 3 µm Pore Size Filter
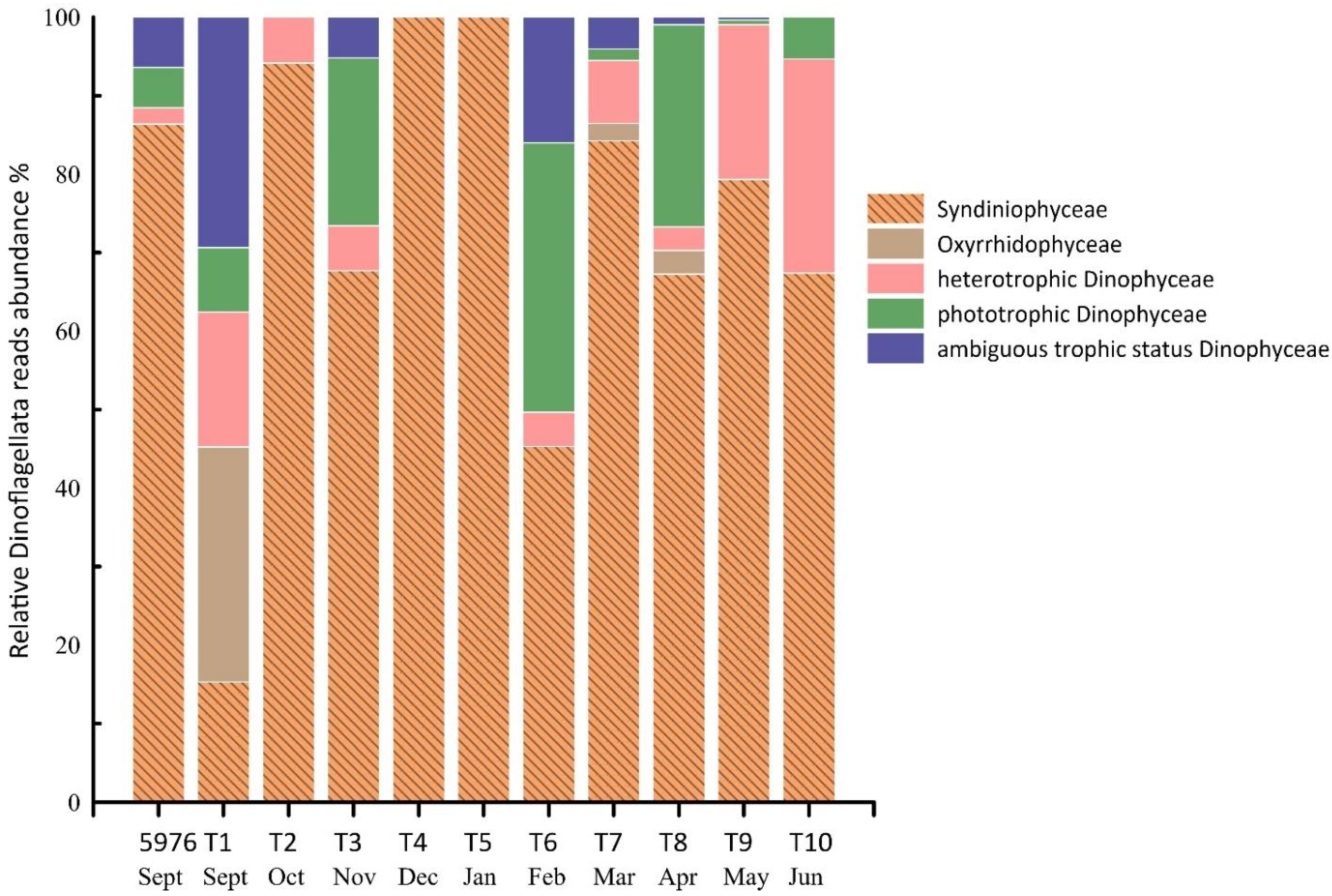
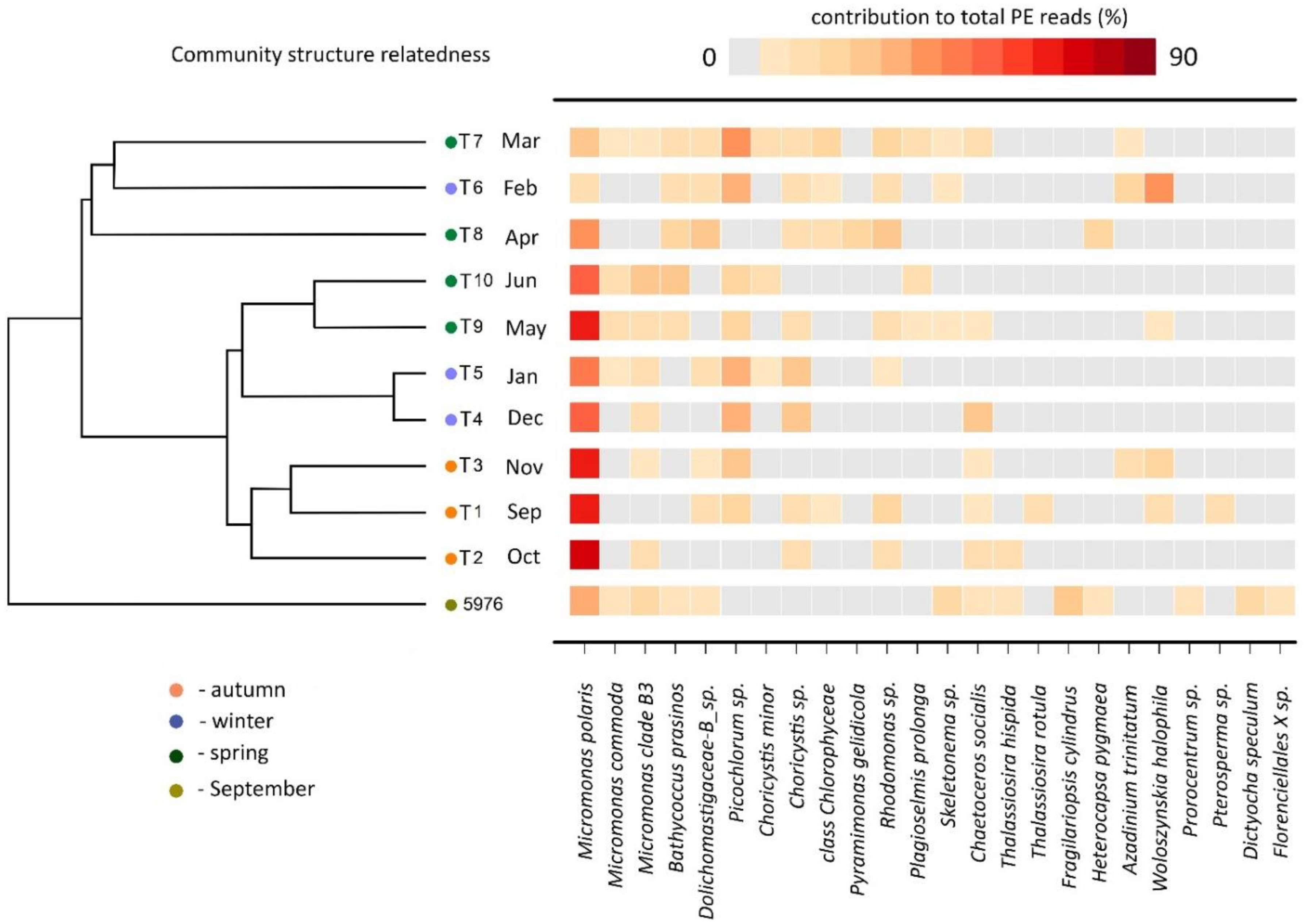
3.5. PE Community Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gosselin, M.; Levasseur, M.; Wheeler, P.A.; Horner, R.A.; Booth, B.C. New measurements of phytoplankton and ice algal production in the Arctic Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 1997, 44, 1623–1644. [Google Scholar] [CrossRef]
- Sherr, E.B.; Sherr, B.F.; Wheeler, P.A.; Thompson, K. Temporal and spatial variation in stocks of autotrophic and hetero-trophic microbes in the upper water column of the central Arctic Ocean. Deep-Sea Res. Part I Oceanogr. Res. Pap. 2003, 50, 557–571. [Google Scholar] [CrossRef]
- Bhavya, P.S.; Lee, J.H.; Lee, H.W.; Kang, J.J.; Lee, J.H.; Lee, D.; An, S.H.; Stockwell, D.A.; Whitledge, T.E.; Lee, S.H. First in situ estima-tions of small phytoplankton carbon and nitrogen uptake rates in the Kara, Laptev, and East Siberian seas. Biogeosciences 2018, 15, 5503–5517. [Google Scholar] [CrossRef] [Green Version]
- Booth, B.C.; Horner, R.A. Microalgae on the arctic ocean section, 1994: Species abundance and biomass. Deep Sea Res. Part II Top. Stud. Oceanogr. 1997, 44, 1607–1622. [Google Scholar] [CrossRef]
- Tremblay, G.; Belzile, C.; Gosselin, M.; Poulin, M.; Roy, S.; Tremblay, J. Late summer phytoplankton distribution along a 3500 km transect in Canadian Arctic waters: Strong numerical dominance by picoeukaryotes. Aquat. Microb. Ecol. 2009, 54, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Demidov, A.; Sukhanova, I.; Belevich, T.; Flint, M.; Gagarin, V.; Sergeeva, V.; Eremeeva, E.; Fedorov, A. Size-fractionated surface Phytoplankton in the Kara and Laptev Seas: Environmental control and spatial variability. Mar. Ecol. Prog. Ser. 2021, 664, 59–77. [Google Scholar] [CrossRef]
- Kilias, E.S.; Nöthig, E.-M.; Wolf, C.; Metfies, K. Picoeukaryote Plankton Composition off West Spitsbergen at the Entrance to the Arctic Ocean. J. Eukaryot. Microbiol. 2014, 61, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Metfies, K.; von Appen, W.-J.; Kilias, E.; Nicolaus, A.; Nöthig, E.-M. Biogeography and Photosynthetic Biomass of Arctic Marine Pico-Eukaroytes during Summer of the Record Sea Ice Minimum 2012. PLoS ONE 2016, 11, e0148512. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; Steele, J.; Caporaso, J.G.; Steinbrück, L.; Reeder, J.; Temperton, B.; Huse, S.M.; McHardy, A.C.; Knight, R.; Joint, I.; et al. Defining seasonal marine microbial community dynamics. ISME J. 2011, 6, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, C.; McClelland, J.W.; Dunton, K.H.; Crump, B.C. Strong Seasonality in Arctic Estuarine Microbial Food Webs. Front. Microbiol. 2019, 10, 2628. [Google Scholar] [CrossRef]
- Serreze, M.C.; Barry, R. Processes and impacts of Arctic amplification: A research synthesis. Glob. Planet. Chang. 2011, 77, 85–96. [Google Scholar] [CrossRef]
- Li, W.K.W.; Carmack, E.C.; McLaughlin, F.; Nelson, R.J.; Williams, W.J. Spacefor-time substitution in predicting the state of picoplankton and nanoplankton in a changing Arctic Ocean. J. Geophys. Res. Ocean. 2013, 118, 5750–5759. [Google Scholar] [CrossRef]
- Lovejoy, C.; Vincent, W.F.; Bonilla, S.; Roy, S.; Martineau, M.-J.; Terrado, R. Distribution, phylogeny and growth of cold-adapted picoprasinophytes in arctic seas. J. Phycol. 2007, 43, 78–89. [Google Scholar] [CrossRef]
- Morán, X.A.G.; López-Urrutia, Á.; Calvo-Díaz, A.; Li, W.K.W. Increasing importance of small phytoplankton in a warmer ocean. Glob. Chang. Biol. 2010, 16, 1137–1144. [Google Scholar] [CrossRef]
- Acevedo-Trejos, E.; Brandt, G.; Steinacher, M.; Merico, A. A glimpse into the future composition of marine phytoplankton communities. Front. Mar. Sci. 2014, 1, 15. [Google Scholar] [CrossRef] [Green Version]
- Fischer, G.; Karakas, G. Sinking rates and ballast composition of particles in the Atlantic Ocean: Implications for the or-ganic carbon fluxes to the deep ocean. Biogeosciences 2009, 6, 85–102. [Google Scholar] [CrossRef] [Green Version]
- Reigstad, M.; Riser, C.W.; Wassmann, P.; Ratkova, T. Vertical export of particulate organic carbon: Attenuation, composition and loss rates in the northern Barents Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 2308–2319. [Google Scholar] [CrossRef]
- Buesseler, K.O.; Lamborg, C.H.; Boyd, P.W.; Lam, P.J.; Trull, T.W.; Bidigare, R.R.; Bishop, J.K.B.; Casciotti, K.L.; Dehairs, F.; Elskens, M.; et al. Revisiting Carbon Flux Through the Ocean’s Twilight Zone. Science 2007, 316, 567–570. [Google Scholar] [CrossRef] [Green Version]
- Metfies, K.; Bauerfeind, E.; Wolf, C.; Sprong, P.; Frickenhaus, S.; Kaleschke, L.; Nicolaus, A.; Nöthig, E.-M. Protist Communities in Moored Long-Term Sediment Traps (Fram Strait, Arctic)–Preservation with Mercury Chloride Allows for PCR-Based Molecular Genetic Analyses. Front. Mar. Sci. 2017, 4, 301. [Google Scholar] [CrossRef] [Green Version]
- Meshram, A.R.; Vader, A.; Kristiansen, S.; Gabrielsen, T.M. Microbial Eukaryotes in an Arctic Under-Ice Spring Bloom North of Svalbard. Front. Microbiol. 2017, 8, 1099. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, R.; Lepere, C.; Jardillier, L.E.; Not, F.; Bouman, H.; Mead, A.; Scanlan, D.J. A global perspective on marine photosyn-thetic picoeukaryote community structure. ISME J. 2013, 7, 922–936. [Google Scholar] [CrossRef] [Green Version]
- Marquardt, M.; Vader, A.; Stübner, E.I.; Reigstad, M.; Gabrielsen, T.M. Strong Seasonality of Marine Microbial Eukaryotes in a High-Arctic Fjord (Isfjorden, in West Spitsbergen, Norway). Appl. Environ. Microbiol. 2016, 82, 1868–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordeev, V.V.; Martin, J.M.; Sidorov, I.S.; Sidorova, M.V. A reassessment of the Eurasian river input of water, sediment, ma-jor elements, and nutrients to the Arctic Ocean. Am. J. Sci. 1996, 296, 664–691. [Google Scholar] [CrossRef]
- Stain, R. Circum Arctic river discharge and its geological record. Int. J. Earth Sci. 2000, 89, 447–449. [Google Scholar] [CrossRef]
- Sukhanova, I.N.; Flint, M.V.; Druzhkova, E.I.; Sazhin, A.F.; Sergeeva, V. Phytoplankton in the northwestern Kara Sea. Oceanology 2015, 55, 547–560. [Google Scholar] [CrossRef]
- Belevich, T.A.; Ilyash, L.V.; Demidov, A.B.; Flint, M.V. Picophytoplankton Distribution at the Ob River Section and in the Western Part of the Kara Sea. Oceanology 2019, 59, 871–880. [Google Scholar] [CrossRef]
- Belevich, T.A.; Milyutina, I.A.; Troitsky, A.V.; Flint, M.V. Picophytoplankton in Blagopoluchia Bay (Novaya Zemlya Archipelago) and Adjacent Part of the Kara Sea. Oceanology 2020, 60, 473–482. [Google Scholar] [CrossRef]
- Belevich, T.A.; Milyutina, I.A.; Abyzova, A.G.; Troitsky, A.V. The pico-sized Mamiellophyceae and a novel Bathycoccus clade from the summer plankton of Russian Arctic Seas and adjacent waters. FEMS Microbiol. Ecol. 2021, 97, 2. [Google Scholar] [CrossRef] [PubMed]
- Verity, P.G.; Robertson, C.Y.; Tronzo, C.R.; Andrews, M.G.; Nelson, J.R.; Sieracki, M. Relationships between cell volume and the carbon and nitrogen content of marine photosynthetic nanoplankton. Limnol. Oceanogr. 1992, 37, 1434–1446. [Google Scholar] [CrossRef]
- DuRand, M.D.; Olson, R.J.; Chisholm, S. Phytoplankton population dynamics at the Bermuda Atlantic Time-series station in the Sargasso Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 1983–2003. [Google Scholar] [CrossRef]
- Holm-Hansen, O.; Riemann, B. Chlorophyll a Determination: Improvements in Methodology. Oikos 1978, 30, 438–447. [Google Scholar] [CrossRef]
- Belevich, T.A.; Ilyash, L.V.; Milyutina, I.A.; Logacheva, M.D.; Goryunov, D.V.; Troitsky, A. Photosynthetic Picoeukaryotes in the Land-Fast Ice of the White Sea, Russia. Microb. Ecol. 2017, 75, 582–597. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 25 February 2021).
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2006; 192p, Available online: https://www.primer-e.com/ (accessed on 31 October 2021).
- Tragin, M.; Vaulot, D. Novel diversity within marine Mamiellophyceae (Chlorophyta) unveiled by metabarcoding. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Belevich, T.A.; Ilyash, L.V.; Milyutina, I.A.; Logacheva, M.D.; Troitsky, A.V. Metagenomics of bolidophyceae in plankton and ice of the White Sea. Biochemistry 2017, 82, 1538–1548. [Google Scholar] [CrossRef] [PubMed]
- Mosharov, S.A.; Demidov, A.B.; Simakova, U. Peculiarities of the primary production process in the Kara Sea at the end of the vegetation season. Oceanology 2016, 56, 84–94. [Google Scholar] [CrossRef]
- Demidov, A.B.; Mosharov, S.A.; Makkaveev, P.N. Evaluation of the influence of abiotic and biotic factors on primary production in the Kara Sea in autumn. Oceanology 2015, 55, 535–546. [Google Scholar] [CrossRef]
- Drits, A.; Pasternak, A.; Arashkevich, E.; Kravchishina, M.; Sukhanova, I.; Sergeeva, V.; Flint, M. Influence of riverine discharge and timing of ice retreat on particle sedimentation patterns on the Laptev Sea shelf. J. Geophys. Res. Oceans 2021, 126, e2021JC017462. [Google Scholar] [CrossRef]
- Sergeeva, V.M.; Sukhanova, I.N.; Druzhkova, E.I.; Mosharov, S.A.; Kremenetskiy, V.V.; Poluhin, A.A. The structure and distribution of the phytoplankton community in the deep region of the Northern Kara Sea. Oceanology 2016, 56, 107–113. [Google Scholar] [CrossRef]
- Lovejoy, C.; Potvin, M. Microbial eukaryotic distribution in a dynamic Beaufort Sea and the Arctic Ocean. J. Plankton Res. 2010, 33, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Iversen, K.R.; Seuthe, L. Seasonal microbial processes in a high-latitude fjord (Kongsfjorden, Svalbard): I. Heterotrophic bacteria, picoplankton and nanoflagellates. Polar Biol. 2011, 34, 731–749. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; He, J.; Lin, L.; Jin, H. Dominance of picophytoplankton in the newly open surface water of the central Arctic Ocean. Polar Biol. 2015, 38, 1081–1089. [Google Scholar] [CrossRef]
- Bezzubova, E.M.; Drits, A.V.; Mosharov, S.A. Effect of Mercury Chloride on the Chlorophyl a and Pheophytin Content in Marine Microalgae: Measuring the Flow of Autotrophic Phytoplankton Using Sediment Traps Data. Oceanology 2018, 58, 479–486. [Google Scholar] [CrossRef]
- Gran, H.H. Diatomaceae from the ice-floes and plankton of the Arctic Ocean; McLaren Books Ltd., ABA(associate), PBFA: Largs, UK, 1904; Volume 4, pp. 3–74. [Google Scholar]
- Poulin, M.; Daugbjerg, N.; Gradinger, R.; Ilyash, L.; Ratkova, T.; von Quillfeldt, C. The pan-Arctic biodiversity of marine pe-lagic and sea-ice unicellular eukaryotes: A first-attempt assessment. Mar. Biodivers. 2011, 41, 13–28. [Google Scholar] [CrossRef]
- Degerlund, M.; Eilertsen, H.C. Main Species Characteristics of Phytoplankton Spring Blooms in NE Atlantic and Arctic Waters (68–80° N). Chesap. Sci. 2010, 33, 242–269. [Google Scholar] [CrossRef]
- Sørensen, N.; Daugbjerg, N.; Gabrielsen, T.M. Molecular diversity and temporal variation of picoeukaryotes in two Arctic fjords, Svalbard. Polar Biol. 2012, 35, 519–533. [Google Scholar] [CrossRef]
- Romari, K.; Vaulot, D. Composition and temporal variability of picoeukaryote communities at a coastal site of the Eng-lish Channel from 18S rDNA sequences. Limnol. Oceanogr. 2004, 49, 784–798. [Google Scholar] [CrossRef] [Green Version]
- Comeau, A.M.; Philippe, B.; Thaler, M.; Gosselin, M.; Poulin, M.; Lovejoy, C. Protists in Arctic drift and land-fast sea ice. J. Phycol. 2013, 49, 229–240. [Google Scholar] [CrossRef]
- Kataoka, T.; Yamaguchy, H.; Sato, M.; Watanabe, T.; Taniuchi, Y.; Kuwata, A.; Kawachi, M. Seasonal and geographical dis-tribution of near-surface small photosynthetic eukaryotes in the western North Pacific determined by pyrosequencing of 18S rDNA. FEMS Microbiol. Ecol. 2017, 93, fiw229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Not, F.; Del Campo, J.; Balagué, V.; De Vargas, C.; Massana, R. New Insights into the Diversity of Marine Picoeukaryotes. PLoS ONE 2009, 4, e7143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, D.H. The Ciliated Protozoa; Springer Science + Business Media B.V.: New York, NY, USA, 2008; Correct Printing 2010; p. 605. [Google Scholar]
- De Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahé, F.; Logares, R.; Lara, E.; Berney, C.; Le Bescot, N.; Probert, I.; et al. Ocean plankton. Eukaryotic plankton diversity in the sunlit ocean. Science 2015, 348, 1261605. [Google Scholar] [CrossRef] [Green Version]
- Giner, C.R.; Balagué, V.; Krabberød, A.K.; Ferrera, I.; Reñé, A.; Garcés, E.; Gasol, J.M.; Logares, R.; Massana, R. Quantifying long-term recurrence in planktonic microbial eukaryotes. Mol. Ecol. 2019, 28, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Gimmler, A.; Korn, R.; De Vargas, C.; Audic, S.; Stoeck, T. The Tara Oceans voyage reveals global diversity and distribution patterns of marine planktonic ciliates. Sci. Rep. 2016, 6, 33555. [Google Scholar] [CrossRef] [Green Version]
- Guidi, L.; Tara Oceans Consortium Coordinators; Chaffron, S.; Bittner, L.; Eveillard, D.; Larhlimi, A.; Roux, S.; Darzi, Y.; Audic, S.; Berline, L.; et al. Plankton networks driving carbon export in the oligotrophic ocean. Nat. Cell Biol. 2016, 532, 465–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Wang, C.; Liu, D.; He, N.; Deng, X. Hg tolerance and biouptake of an isolated pigmentation yeast Rhodotorula mucilaginosa. PLoS ONE 2017, 12, e0172984. [Google Scholar] [CrossRef]
- Bachy, C.; López-García, P.; Vereshchaka, A.; Moreira, D. Diversity and Vertical Distribution of Microbial Eukaryotes in the Snow, Sea Ice and Seawater Near the North Pole at the End of the Polar Night. Front. Microbiol. 2011, 2, 106. [Google Scholar] [CrossRef] [Green Version]
- Majaneva, M.; Rintala, J.-M.; Piisilä, M.; Fewer, D.P.; Blomster, J. Comparison of wintertime eukaryotic community from sea ice and open water in the Baltic Sea, based on sequencing of the 18S rRNA gene. Polar Biol. 2012, 35, 875–889. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Kohlmeyer, E. Marine Mycology: The Higher Fungi; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Newell, S. Fungal biomass and productivity. Methods Microbiol. 2001, 30, 357–372. [Google Scholar] [CrossRef]
- Stock, A.; Jürgens, K.; Bunge, J.; Stoeck, T. Protistan diversity in suboxic and anoxic waters of the Gotland Deep (Baltic Sea) as revealed by 18S rRNA clone libraries. Aquat. Microb. Ecol. 2009, 55, 267–284. [Google Scholar] [CrossRef]
- Jobard, M.; Rasconi, S.; Sime-Ngando, T. Diversity and functions of microscopic fungi: A missing component in pelagic food webs. Aquat. Sci. 2010, 72, 255–268. [Google Scholar] [CrossRef]
- López-García, P.; Rodríguez-Valera, F.; Pedrós-Alió, C.; Moreira, D. Unexpected diversity of small eukary-otes in deep-sea Antarctic plankton. Nature 2001, 409, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Moon-van der Staay, S.Y.; De Wachter, R.; Vaulot, D. Oceanic 18S rDNA sequences from picoplankton reveal unsuspect-ed eukaryotic diversity. Nature 2001, 409, 607–610. [Google Scholar] [CrossRef]
- Moreira, D.; López-García, P. Are hydrothermal vents oases for parasitic protists? Trends Parasitol. 2003, 119, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Guillou, L.; Viprey, M.; Chambouvet, A.; Welsh, R.M.; Kirkham, A.R.; Massana, R.; Scanlan, D.J.; Worden, A.Z. Widespread occurrence and genetic diversity of marine parasitoids belonging toSyndiniales(Alveolata). Environ. Microbiol. 2008, 10, 3349–3365. [Google Scholar] [CrossRef] [Green Version]
- Lovejoy, C.; Massana, R.; Pedrós-Alió, C. Diversity and Distribution of Marine Microbial Eukaryotes in the Arctic Ocean and Adjacent Seas. Appl. Environ. Microbiol. 2006, 72, 3085–3095. [Google Scholar] [CrossRef] [Green Version]
- Stoeck, T.; Kasper, J.; Bunge, J.; Leslin, C.; Ilyin, V.; Epstein, S. Protistan Diversity in the Arctic: A Case of Paleoclimate Shaping Modern Biodiversity? PLoS ONE 2007, 2, e728. [Google Scholar] [CrossRef] [Green Version]
- Worden, A.Z.; Not, F. Ecology and Diversity of Picoeukaryotes. In Microbial Ecology of the Oceans; Wiley: Hoboken, NJ, USA, 2008; pp. 159–205. [Google Scholar]
- Monier, A.; Comte, J.; Babin, M.; Forest, A.; Matsuoka, A.; Lovejoy, C. Oceanographic structure drives the assembly processes of microbial eukaryotic communities. ISME J. 2015, 9, 990–1002. [Google Scholar] [CrossRef] [Green Version]
- Simmons, M.P.; Bachy, C.; Sudek, S.; van Baren, M.J.; Sudek, L.; Ares, M., Jr.; Worden, A.Z. Intron invasions trace algal specia-tion and reveal nearly identical arctic and Antarctic Micromonas populations. Mol. Biol. Evol. 2015, 32, 2219–2235. [Google Scholar] [CrossRef] [Green Version]
- Simon, N.; Barlow, R.G.; Marie, M.; Partensky, F.; Vaulot, D. Characterization of oceanic photosynthetic picoeukaryotes by flow cytometry1. J. Phycol. 1994, 30, 922–935. [Google Scholar] [CrossRef]
- Vaulot, D.; Eikrem, W.; Viprey, M.; Moreau, H. The diversity of small eukaryotic phytoplankton (<3 µm) in marine eco-systems. FEMS Microbiol. Rev. 2008, 32, 795–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatsepin, G.; Zavialov, P.O.; Kremenetskiy, V.V.; Poyarkov, S.G.; Soloviev, D.M. The upper desalinated layer in the Kara Sea. Oceanology 2010, 50, 657–667. [Google Scholar] [CrossRef]
- Foflonker, F.; Mollegard, D.; Ong, M.; Yoon, H.S.; Bhattacharya, D. Genomic Analysis of Picochlorum Species Reveals How Microalgae May Adapt to Variable Environments. Mol. Biol. Evol. 2018, 35, 2702–2711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthiessen, J.; Kunz-Pirrung, M.; Mudie, P.J. Freshwater chlorophycean algae in recent marine sediments of the Beaufort, Laptev and Kara Seas (Arctic Ocean) as indicators of river runoff. Acta Diabetol. 2000, 89, 470–485. [Google Scholar] [CrossRef]
- Metfies, K.; Gescher, C.; Frickenhaus, S.; Niestroy, R.; Wichels, A.; Gerdts, G.; Knefelkamp, B.; Wiltshire, K.; Medlin, L.K. Contribution of the class cryptophyceae to phytoplankton structure in the german bight1. J. Phycol. 2010, 46, 1152–1160. [Google Scholar] [CrossRef]
- Mikkelsen, D.M.; Rysgaard, S.; Glud, R.N. Microalgal composition and primary production in Arctic sea ice: A seasonal study from Kobbefjord (Kangerluarsunnguaq), West Greenland. Mar. Ecol. Prog. Ser. 2008, 368, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Vallières, C.; Retamal, L.; Ramlal, P.; Osburn, C.; Vincent, W.F. Bacterial production and microbial food web structure in a large arctic river and the coastal Arctic Ocean. J. Mar. Syst. 2008, 74, 756–773. [Google Scholar] [CrossRef]
- Coupel, P.; Jin, H.Y.; Joo, M.; Horner, R.; Bouvet, H.A.; Garcon, V.; Sicre, M.-A.; Gascard, J.-C.; Chen, J.F.; Ruiz-Pino, D. Arctic phy-toplankton in unusually low sea ice cover. Biogeosci. Discuss 2012, 9, 2055–2093. [Google Scholar] [CrossRef] [Green Version]
- Piwosz, K.; Kownacka, J.; Ameryk, A.; Zalewski, M.; Pernthaler, J. Phenology of cryptomonads and the CRY1 lineage in a coastal brackish lagoon (Vistula Lagoon, Baltic Sea). J. Phycol. 2016, 52, 626–637. [Google Scholar] [CrossRef]
- Medlin, L.K.; Piwosz, K.; Metfies, K. Uncovering hidden biodiversity in the Cryptophyta: Clone library studies at the Hel-goland Time Series Site in the Southern German Bight indentifies the cryptophycean clade potentially responsible for the ma-jority of its genetic diversity during the spring bloom. Vie et Millieu Life Environ. 2017, 67, 27–32. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Date/ Period | Total No. of V4 Tag Quality Sequences | Quality Sequences | No. of Protist Reads | No. of Protist ASVs | No. of Photosynthetic Eukaryotes Reads | No. of Photosynthetic Eukaryotes ASVs | Chao1 |
|---|---|---|---|---|---|---|---|---|
| 5976 | 8 September | 304,129 | 129,900 | 127,795 | 89 | 27,164 | 28 | 22 |
| T1 | September | 157,252 | 118,966 | 94,543 | 62 | 36,344 | 16 | 13 |
| T2 | October | 110,720 | 86,035 | 74,141 | 47 | 38,955 | 13 | 11 |
| T3 | November | 202,474 | 160,279 | 139,922 | 66 | 74,172 | 19 | 14 |
| T4 | December | 127,790 | 93,930 | 71,426 | 45 | 21,589 | 13 | 7 |
| T5 | January | 260,009 | 217,018 | 177,565 | 71 | 47,926 | 22 | 15 |
| T6 | February | 190,536 | 146,055 | 125,063 | 106 | 33,073 | 31 | 19 |
| T7 | March | 236,408 | 153,141 | 133,153 | 257 | 40,360 | 81 | 46 |
| T8 | April | 221,949 | 145,227 | 94,280 | 85 | 15,347 | 21 | 13 |
| T9 | May | 232,284 | 159,887 | 137,194 | 169 | 45,242 | 56 | 26 |
| T10 | June | 385,572 | 105,569 | 76,089 | 55 | 30,434 | 17 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belevich, T.A.; Milyutina, I.A.; Troitsky, A.V. Seasonal Variability of Photosynthetic Microbial Eukaryotes (<3 µm) in the Kara Sea Revealed by 18S rDNA Metabarcoding of Sediment Trap Fluxes. Plants 2021, 10, 2394. https://doi.org/10.3390/plants10112394
Belevich TA, Milyutina IA, Troitsky AV. Seasonal Variability of Photosynthetic Microbial Eukaryotes (<3 µm) in the Kara Sea Revealed by 18S rDNA Metabarcoding of Sediment Trap Fluxes. Plants. 2021; 10(11):2394. https://doi.org/10.3390/plants10112394
Chicago/Turabian StyleBelevich, Tatiana A., Irina A. Milyutina, and Aleksey V. Troitsky. 2021. "Seasonal Variability of Photosynthetic Microbial Eukaryotes (<3 µm) in the Kara Sea Revealed by 18S rDNA Metabarcoding of Sediment Trap Fluxes" Plants 10, no. 11: 2394. https://doi.org/10.3390/plants10112394
APA StyleBelevich, T. A., Milyutina, I. A., & Troitsky, A. V. (2021). Seasonal Variability of Photosynthetic Microbial Eukaryotes (<3 µm) in the Kara Sea Revealed by 18S rDNA Metabarcoding of Sediment Trap Fluxes. Plants, 10(11), 2394. https://doi.org/10.3390/plants10112394