
Morphology and Anatomy of Branch–Branch Junctions in Opuntia ficus-indica and Cylindropuntia bigelovii: A Comparative Study Supported by Mechanical Tissue Quantification

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation
2.2. Molecular Genetic Determination
2.3. Morphometric Analyses
2.4. Magnetic Resonance Imaging
2.5. Anatomical Analyses
2.6. Tissue Mechanics
2.7. Statistics
3. Results
3.1. Molecular Genetic Determination
3.2. Morphometric Analyses
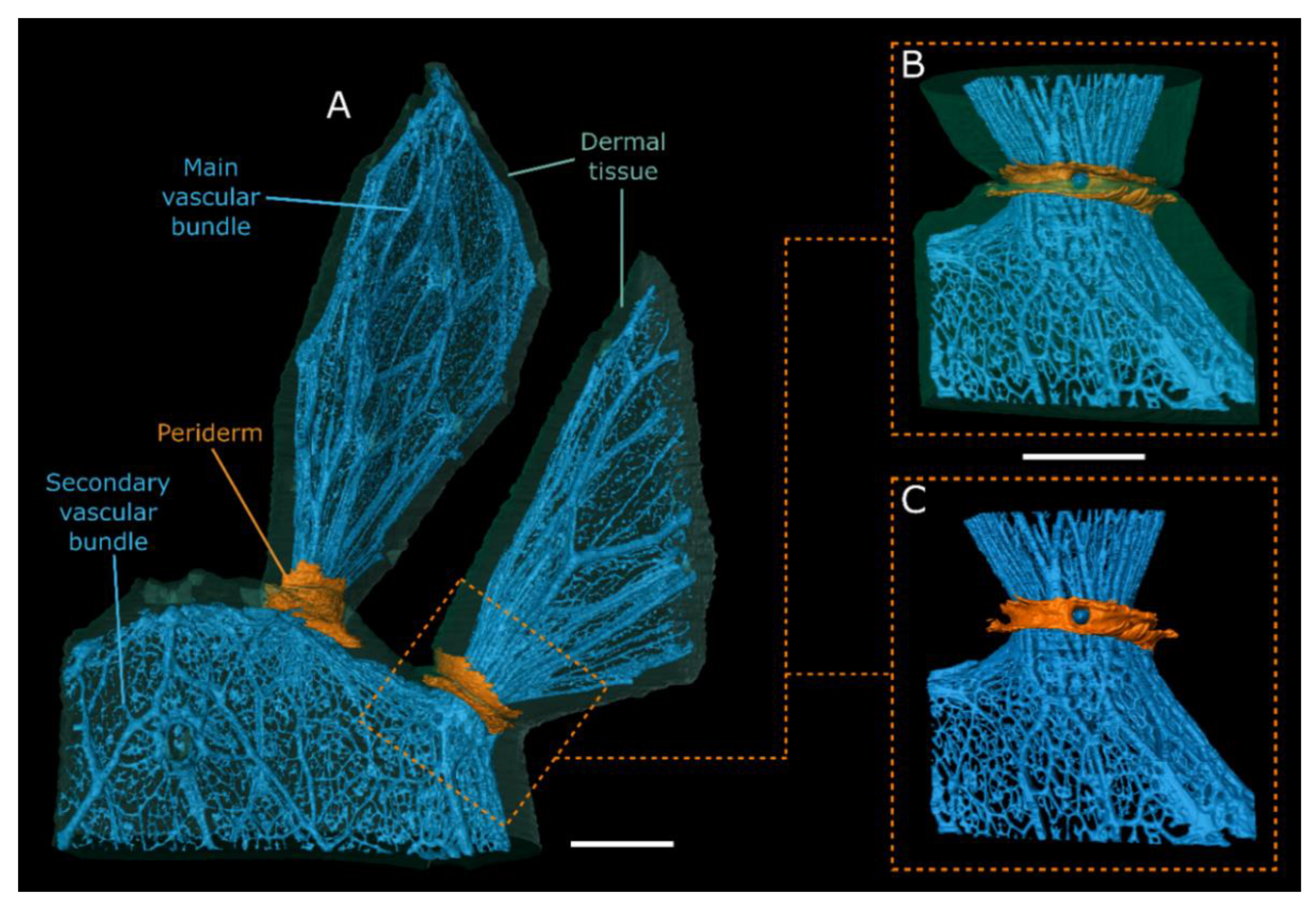
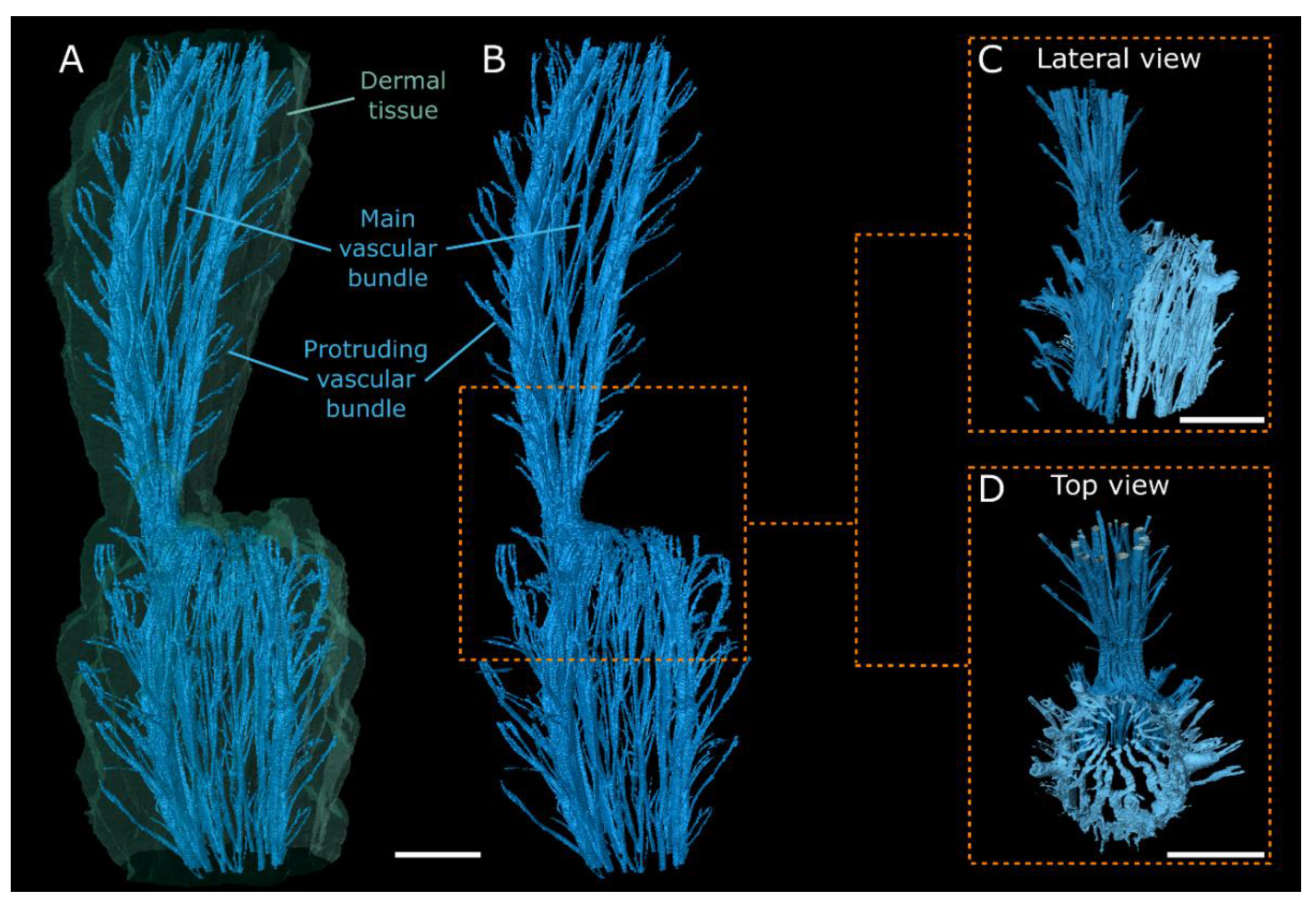
3.3. MRI Analyses
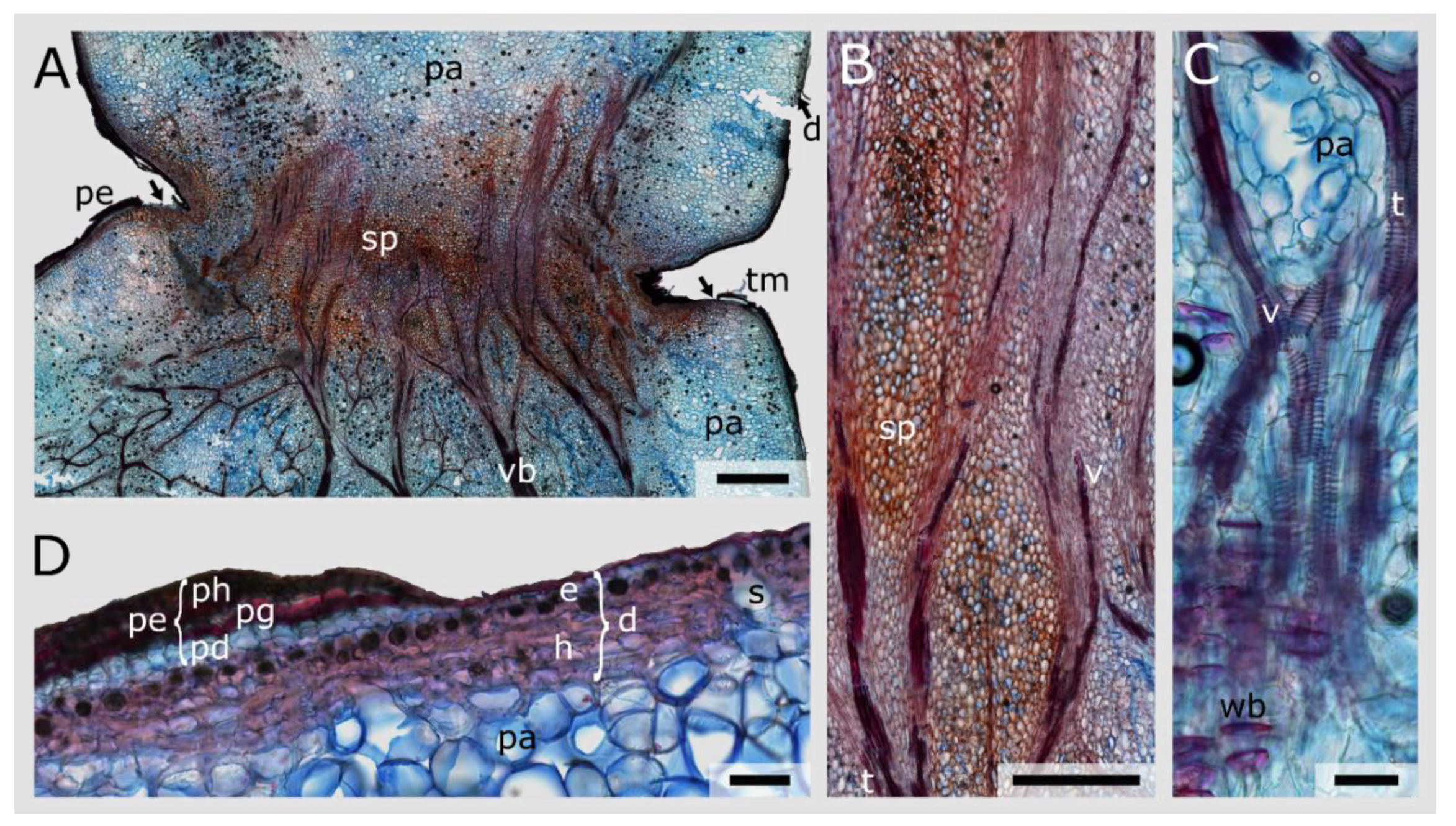
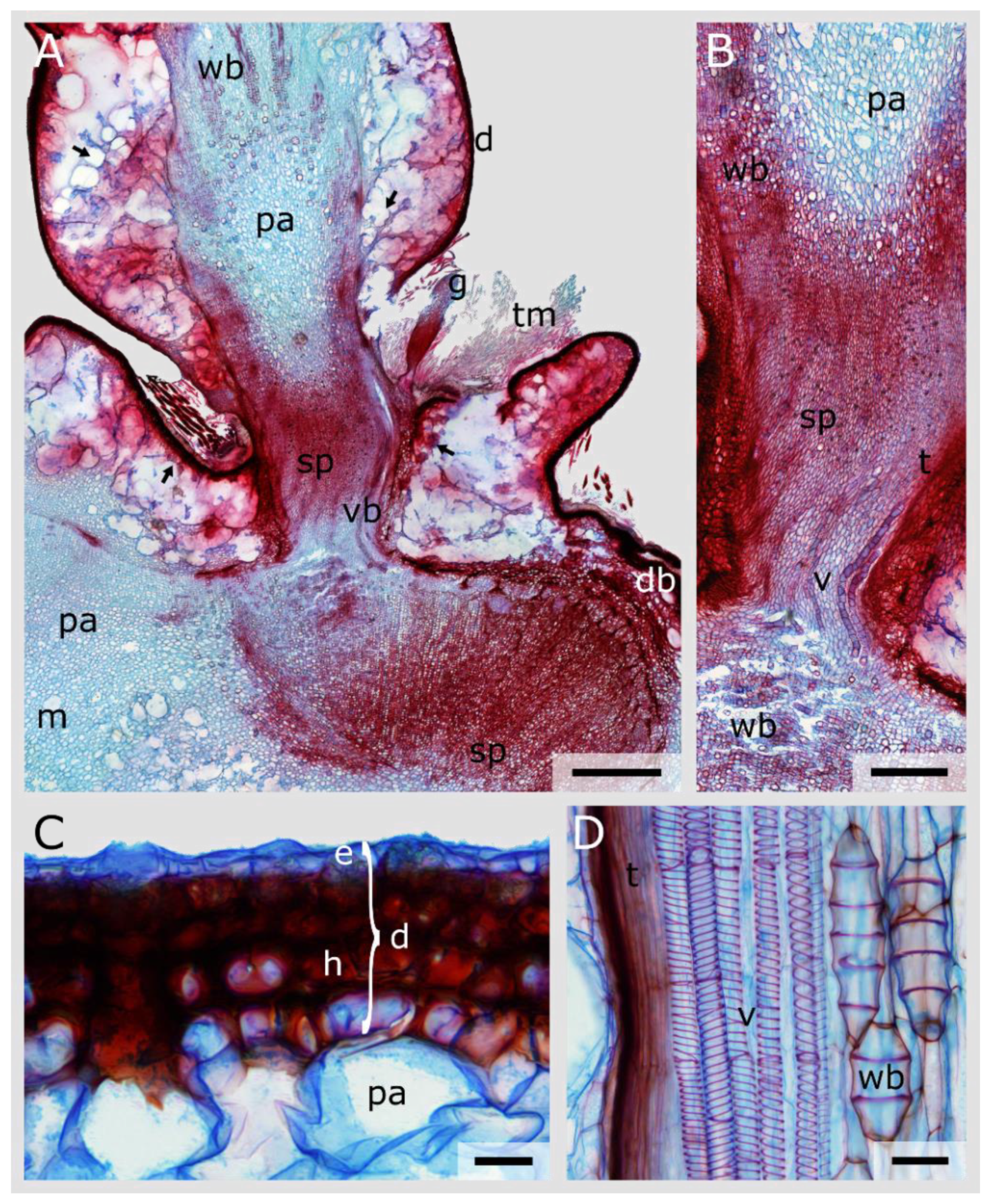
3.4. Junction Histology
3.5. Periderm Formation in Opuntia Ficus-Indica
3.6. Tissue Mechanics
4. Discussion
4.1. Junction Morphology
4.2. Junction Histology
4.3. Tissue Mechanics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nobel, P.S. Cacti: Biology and Uses; University of California Press: Berkeley, CA, USA, 2002. [Google Scholar]
- Benson, L. The Cacti of the United States and Canada; Stanford University Press: Stanford, CA, USA, 1982. [Google Scholar]
- Griffith, M.P.; Porter, J.M. Phylogeny of Opuntioideae (Cactaceae). Int. J. Plant Sci. 2009, 170, 107–116. [Google Scholar] [CrossRef]
- Mauseth, J.D. Structure-function relationships in highly modified shoots of Cactaceae. Ann. Bot. 2006, 98, 901–926. [Google Scholar] [CrossRef]
- Conde, L.F. Anatomical Comparisons of Five Species of Opuntia (Cactaceae). Ann. Mo. Bot. Gard. 1975, 62, 425. [Google Scholar] [CrossRef]
- Mauseth, J.D. Anatomical features, other than wood, in subfamily Opuntioideae (Cactaceae). Haseltonia 2005, 11, 113–125. [Google Scholar] [CrossRef]
- Nobel, P.S.; Cavelier, J.; Andrade, J.L. Mucilage in cacti: Its apoplastic capacitance, associated solutes, and influence on tissue 5. J. Exp. Bot. 1992, 43, 641–648. [Google Scholar] [CrossRef]
- Mauseth, J.D. Release of whole cells of Nopalea (Cactaceae) into secretory canals. Bot. Gaz. 1980, 141, 15–18. [Google Scholar] [CrossRef]
- Mauseth, J.D. Wood in the cactus subfamily Opuntioideae has extremely diverse structure. Bradleya 2006, 24, 93–106. [Google Scholar] [CrossRef]
- Reyes-Rivera, J.; Soto-Hernández, M.; Canché-Escamilla, G.; Terrazas, T. Structural characterization of lignin in four cacti wood: Implications of lignification in the growth form and succulence. Front. Plant Sci. 2018, 9, 1518. [Google Scholar] [CrossRef]
- Reyes-Agüero, J.A.; Valiente-Banuet, A. Reproductive biology of Opuntia: A review. J. Arid Environ. 2006, 64, 549–585. [Google Scholar] [CrossRef]
- Nobel, P.S.; Gibson, A.C. The Cactus Primer; Harvard University Press: Cambridge, MA, USA, 1986. [Google Scholar]
- Bobich, E.G. Vegetative reproduction, population structure, and morphology of Cylindropuntia fulgida var. mamillata in a desert grassland. Int. J. Plant Sci. 2005, 166, 97–104. [Google Scholar] [CrossRef]
- Allen, L.J.S.; Allen, E.J.; Kunst, C.R.G.; Sosebee, R.E. A diffusion model for dispersal of Opuntia imbricata (Cholla) on rangeland. J. Ecol. 1991, 79, 1123. [Google Scholar] [CrossRef]
- Del Carmen Mandujano, M.; Montana, C.; Mendez, I.; Golubov, J. The relative contributions of sexual reproduction and clonal propagation in Opuntia rastrera from two habitats in the Chihuahuan Desert. J. Ecol. 1998, 86, 911–921. [Google Scholar] [CrossRef]
- Grant, V.; Grant, K.A. Clonal microspecies of hybrid origin in the Opuntia lindheimeri group. Bot. Gaz. 1980, 141, 101–106. [Google Scholar] [CrossRef]
- Addicott, F.T. Abscission; University of California Press: Berkeley, CA, USA, 1982. [Google Scholar]
- Nobel, P.S.; Meyer, R.W. Biomechanics of cladodes and cladode-cladode junctions for Opuntia ficus-indica (Cactaceae). Am. J. Bot. 1991, 78, 1252–1259. [Google Scholar] [CrossRef]
- Bobich, E.G.; Nobel, P.S. Vegetative reproduction as related to biomechanics, morphology and anatomy of four cholla cactus species in the Sonoran Desert. Ann. Bot. 2001, 87, 485–493. [Google Scholar] [CrossRef][Green Version]
- Bobich, E.G.; Nobel, P.S. Biomechanics and anatomy of cladode junctions for two Opuntia (Cactaceae) species and their hybrid. Am. J. Bot. 2001, 88, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Bobich, E.G.; Nobel, P.S. Cladode junction regions and their biomechanics for arborescent Platyopuntias. Int. J. Plant Sci. 2002, 163, 507–517. [Google Scholar] [CrossRef]
- Masselter, T.; Haushahn, T.; Schwager, H.; Milwich, M.; Nathanson, R.; Gude, M.; Cichy, F.; Hufenbach, W.; Neinhuis, C.; Speck, T. Biomimetic fibre-reinforced composites inspired by branched plant stems. In Design and Nature V: Comparing Design in Nature with Science and Engineering; WIT Press: Southampton, UK, 2010; pp. 411–420. [Google Scholar]
- Schwager, H.; Masselter, T.; Speck, T.; Neinhuis, C. Functional morphology and biomechanics of branch-stem junctions in columnar cacti. Proc. Biol. Sci. B 2013, 280, 20132244. [Google Scholar] [CrossRef]
- Bouakba, M.; Bezazi, A.; Boba, K.; Scarpa, F.; Bellamy, S. Cactus fibre/polyester biocomposites: Manufacturing, quasi-static mechanical and fatigue characterisation. Compos. Sci. Technol. 2013, 74, 150–159. [Google Scholar] [CrossRef]
- Greco, A.; Maffezzoli, A. Rotational molding of biodegradable composites obtained with PLA reinforced by the wooden backbone of opuntia ficus indica cladodes. J. Appl. Polym. Sci. 2015, 132, 42447. [Google Scholar] [CrossRef]
- Mannai, F.; Elhleli, H.; Ammar, M.; Passas, R.; Elaloui, E.; Moussaoui, Y. Green process for fibrous networks extraction from Opuntia (Cactaceae): Morphological design, thermal and mechanical studies. Ind. Crop. Prod. 2018, 126, 347–356. [Google Scholar] [CrossRef]
- De Vivo, L.; Matsushita, A.K.; Kupor, D.; Luna, J.; Tierra, B.A.; Sah, R.L.; Lubarda, V.A.; Meyers, M.A.; McKittrick, J.M.; Krysl, P.; et al. Cholla cactus frames as lightweight and torsionally tough biological materials. Acta Biomater. 2020, 112, 213–224. [Google Scholar] [CrossRef]
- Mylo, M.D.; Krüger, F.; Speck, T.; Speck, O. Self-Repair in Cacti Branches: Comparative Analyses of Their Morphology, Anatomy, and Biomechanics. Int. J. Mol. Sci. 2020, 21, 4630. [Google Scholar] [CrossRef] [PubMed]
- Majure, L.C.; Puente, R.; Griffith, M.P.; Judd, W.S.; Soltis, P.S.; Soltis, D.E. Phylogeny of Opuntia s.s. (Cactaceae): Clade delineation, geographic origins, and reticulate evolution. Am. J. Bot. 2012, 99, 847–864. [Google Scholar] [CrossRef]
- Sutton, M.A.; Mingqi, C.; Peters, W.H.; Chao, Y.J.; McNeill, S.R. Application of an optimized digital correlation method to planar deformation analysis. Image Vis. Comput. 1986, 4, 143–150. [Google Scholar] [CrossRef]
- Doube, M.; Kłosowski, M.M.; Arganda-Carreras, I.; Cordelières, F.P.; Dougherty, R.P.; Jackson, J.S.; Schmid, B.; Hutchinson, J.R.; Shefelbine, S.J. BoneJ: Free and extensible bone image analysis in ImageJ. Bone 2010, 47, 1076–1079. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arppe, A. DescTools: Tools for Descriptive Statistics; R Package Version 0.99; R Foundation for Statistical Computing: Vienna, Austria, 2016; p. 18. [Google Scholar]
- Janzen, D. Chihuahuan Desert nopaleras: Defaunated big mammal vegetation. Annu. Rev. Ecol. Evol. Syst. 1986, 17, 595–636. [Google Scholar] [CrossRef]
- Niklas, K.J. Plant Biomechanics: An Engineering Approach to Plant Form and Function; University of Chicago Press: Chicago, IL, USA, 1992. [Google Scholar]
- Niklas, K.J. Plant Allometry: The Scaling of Form and Process; University of Chicago Press: Chicago, IL, USA, 1994. [Google Scholar]
- Langer, M.; Speck, T.; Speck, O. Petiole-lamina transition zone: A functionally crucial but often overlooked leaf trait. Plants 2021, 10, 774. [Google Scholar] [CrossRef]
- Köckenberger, W. Functional imaging of plants by magnetic resonance experiments. Trends Plant Sci. 2001, 6, 286–292. [Google Scholar] [CrossRef]
- Blümler, P.; Windt, C.W.; van Dusschoten, D. Magnetic resonance of plants. Nova Acta Leopold. 2009, 357, 17–30. [Google Scholar]
- Hesse, L.; Masselter, T.; Leupold, J.; Spengler, N.; Speck, T.; Korvink, J.G. Magnetic resonance imaging reveals functional anatomy and biomechanics of a living dragon tree. Sci. Rep. 2016, 6, 32685. [Google Scholar] [CrossRef] [PubMed]
- Hesse, L.; Leupold, J.; Speck, T.; Masselter, T. A qualitative analysis of the bud ontogeny of Dracaena marginata using high-resolution magnetic resonance imaging. Sci. Rep. 2018, 8, 9881. [Google Scholar] [CrossRef] [PubMed]
- Hesse, L.; Bunk, K.; Leupold, J.; Speck, T.; Masselter, T. Structural and functional imaging of large and opaque plant specimens. J. Exp. Bot. 2019, 70, 3659–3678. [Google Scholar] [CrossRef] [PubMed]
- Malik, F.T.; Clement, R.M.; Gethin, D.T.; Kiernan, M.; Goral, T.; Griffiths, P.; Beynon, D.; Parker, A.R. Hierarchical structures of cactus spines that aid in the directional movement of dew droplets. Philos. Trans. A Math. Phys. Eng. Sci. 2016, 374, 2073. [Google Scholar] [CrossRef]
- Niklas, K.J. Dependency of the tensile modulus on transverse dimensions, water potential, and cell number of pith parenchyma. Am. J. Bot. 1988, 75, 1286–1292. [Google Scholar] [CrossRef]
- Taylor, J.E.; Whitelaw, C.A. Signals in abscission. New Phytol. 2001, 151, 323–340. [Google Scholar] [CrossRef]
- Yu, Y.; Leyva, P.; Tavares, R.L.; Kellogg, E.A. The anatomy of abscission zones is diverse among grass species. Am. J. Bot. 2020, 107, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Green, D.W.; Winandy, J.E.; Kretschmann, D.E. Mechanical properties of wood. In Wood Handbook: Wood as an Engineering Material; General Technical Report FPL.; GTR-113; USDA Forest Service, Forest Products Laboratory: Madison, WI, USA, 1999; pp. 4.1–4.45. [Google Scholar]
- Speck, T.; Burgert, I. Plant stems: Functional design and mechanics. Annu. Rev. Mater. Res. 2011, 41, 169–193. [Google Scholar] [CrossRef]
- Bastola, A.K.; Soffiatti, P.; Behl, M.; Lendlein, A.; Rowe, N.P. Structural performance of a climbing cactus: Making the most of softness. J. R. Soc. Interface 2021, 18, 20210040. [Google Scholar] [CrossRef]
- Hesse, L.; Kampowski, T.; Leupold, J.; Caliaro, S.; Speck, T.; Speck, O. Comparative analyses of the self-sealing mechanisms in leaves of Delosperma cooperi and Delosperma ecklonis (Aizoaceae). Int. J. Mol. Sci. 2020, 21, 5768. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| O. ficus-indica | C. bigelovii | Statistical Difference p-Value & Significance Level | |
|---|---|---|---|
| Sample Size N | 12 | 12 | |
| Morphometric Variable | Median (IQR) | Median (IQR) | |
| Junction area | 25.02 | 15.42 | 0.0068 |
| [mm2] | (15.10) | (7.22) | ** b |
| Branch area | 382.09 | 395.58 | 0.4713 |
| [mm2] | (138.04) | (103.41) | n.s. a |
| Area ratio | 0.068 | 0.038 | 0.0011 |
| [ ] | (0.040) | (0.012) | ** b |
| Junction Imajor-axis | 43.86 | 19.56 | 0.0056 |
| [mm4] | (37.42) | (18.62) | ** b |
| Junction Iminor-axis | 56.96 | 26.55 | 0.0173 |
| [mm4] | (45.66) | (29.02) | * b |
| Junction Imajor-axis/Iminor-axis | 0.851 | 0.717 | 0.0678 |
| [ ] | (0.203) | (0.087) | n.s. a |
| Branch Imajor-axis | 2045.41 | 12,600.93 | >0.0001 |
| [mm4] | (1207.33) | (6356.29) | *** b |
| Branch Iminor-axis | 98,875.21 | 13,321.64 | >0.0001 |
| [mm4] | (35,309.61) | (7028.65) | *** b |
| Branch Imajor-axis/Iminor-axis | 0.021 | 0.935 | >0.0001 |
| [ ] | (0.017) | (0.029) | *** a |
| Ratio Imajor-axis: junction/branch | 0.02606 | 0.00133 | >0.0001 |
| [ ] | (0.04370) | (0.00068) | *** b |
| Ratio Iminor-axis: junction/branch | 0.00081 | 0.00192 | 0.0014 |
| [ ] | (0.00033) | (0.00063) | ** a |
| Junction torsion constant K | 110.78 | 43.21 | 0.01 |
| [mm4] | (81.76) | (30.94) | * b |
| Branch torsion constant K | 4776.45 | 24,909.54 | >0.0001 |
| [mm4] | (2854.11) | (13,002.07) | *** b |
| Ratio torsion constant: | 0.0222 | 0.00017 | >0.0001 |
| junction/branch [ ] | (0.05398) | (0.00010) | *** b |
| Tissue | O. ficus-indica | C. bigelovii | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Dermal Tissue | Dermal Tissue Covered with Periderm | Dermal Tissue | |||||||
| Sample Orientation | Longitudinal | Transverse | Longitudinal | Transverse | “Longitudinal” | ||||
| Sample Size N | 12 | 12 | 12 | 12 | 12 | ||||
| 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | ||
| Mechanical Variable | Median (IQR) | Median (IQR) | Median (IQR) | Median (IQR) | Median (IQR) | ||||
| Thickness [mm] | 0.22 A | 0.20 A | 0.26 B | 0.27 B | 0.16 C | ||||
| (0.03) | (0.06) | (0.07) | (0.04) | (0.02) | |||||
| Strength [MPa] | 4.08 A | 3.96 A | 12.21 B | 8.83 BC | 13.36 B | 6.23 C | 11.21 B | ||
| (0.86) | (0.72) | (1.54) | (2.96) | (5.72) | (1.29) | (4.39) | |||
| Elastic modulus [MPa] | 44.09 AB | 40.76 A | 463.95 C | 414.45 C | 39.51 AB | 68.68 B | |||
| (13.73) | (15.59) | (179.48) | (256.97) | (17.95) | (57.09) | ||||
| Fracture energy [mJ/mm2] | 11.94 A | 11.31 A | 4.82 B | 3.20 B | 3.30 B | ||||
| (2.69) | (3.68) | (4.08) | (2.06) | (1.84) | |||||
| Deformation at break [%] | 17.24 A | 20.90 A | 15.00 A | 4.54 B | 10.16 AC | 4.49 BC | 19.51 A | ||
| (3.41) | (2.72) | (2.23) | (2.32) | (1.37) | (0.81) | (8.78) | |||
| Poisson’s ratio [ ] | 0.65 A | 0.52 AB | 0.34 AC | 0.21 BCDE | 0.64 AD | 0.08 E | |||
| (0.21) | (0.33) | (0.22) | (0.23) | (0.18) | (0.09) | ||||
| Tissue | O. ficus-indica | C. bigelovii | |||
|---|---|---|---|---|---|
| Vascular Bundle | Vascular Bundle | Vascular Bundle | |||
| (Young Branch) | (Older Branch) | ||||
| Sample Size N | 12 | 12 | 12 | ||
| 6 | 6 | 6 | 6 | ||
| Mechanical Variable | Median (IQR) | Median (IQR) | Median (IQR) | ||
| Diameter [mm] | 0.153 A | 0.321 B | 0.856 C | 1.068 D | |
| (0.105) | (0.101) | (0.167) | (0.399) | ||
| Strength [MPa] | 56.24 A | 51.11 A | 2.09 B | ||
| (56.52) | (35.75) | (1.25) | |||
| Elastic modulus [MPa] | 1233.83 A | 1673.97 A | 7.44 B | ||
| (1061.79) | (384.52) | (2.89) | |||
| Fracture energy [mJ/mm2] | 36.20 A | 41.90 A | 2.25 B | ||
| (48.82) | (63.03) | (1.56) | |||
| Deformation at break [%] | 10.86 A | 12.21 A | 5.33 A | 65.39 B | 44.11 B |
| (7.36) | (3.36) | (5.70) | (24.54) | (10.99) | |
| Feature | O. ficus-indica | C. bigelovii |
|---|---|---|
| Life-Form | ||
| Growth-form | stem succulent | |
| tree-like | shrubby | |
| Main propagation | sexual via fruits/seeds | vegetative via shed offshoots |
| Branch–branch junction stability | stable | fragile (abscission) |
| Similarities | ||
| Morphometry | distinct cross-sectional taper towards the junctions | |
| Morphology | net-like arrangement of vascular bundles that close together circularly within the junctions | |
| Anatomy | changes in tissue characteristics from the branches to the junctions: lumen of parenchyma changes from large to small and the amount of wide-band tracheids changes to a smaller number | |
| Mechanical properties | comparable stiffness of the dermal tissues (without periderm coverage) | |
| Dissimilarities | ||
| Morphometry | C. bigelovii: significantly lower absolute and relative junction cross-sectional areas | |
| Morphology | O. ficus-indica: periderm formation around lateral junctions | |
| Anatomy | O. ficus-indica: periderm formed as wound tissue by outgrowing areoles | |
| Mechanical properties | O. ficus-indica: periderm stiffens dermal tissue by a factor of about 10 | |
| O. ficus-indica: strength and stiffness of vascular fibres are higher by a factor of about 25 and 200, respectively | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mylo, M.D.; Hesse, L.; Masselter, T.; Leupold, J.; Drozella, K.; Speck, T.; Speck, O. Morphology and Anatomy of Branch–Branch Junctions in Opuntia ficus-indica and Cylindropuntia bigelovii: A Comparative Study Supported by Mechanical Tissue Quantification. Plants 2021, 10, 2313. https://doi.org/10.3390/plants10112313
Mylo MD, Hesse L, Masselter T, Leupold J, Drozella K, Speck T, Speck O. Morphology and Anatomy of Branch–Branch Junctions in Opuntia ficus-indica and Cylindropuntia bigelovii: A Comparative Study Supported by Mechanical Tissue Quantification. Plants. 2021; 10(11):2313. https://doi.org/10.3390/plants10112313
Chicago/Turabian StyleMylo, Max D., Linnea Hesse, Tom Masselter, Jochen Leupold, Kathrin Drozella, Thomas Speck, and Olga Speck. 2021. "Morphology and Anatomy of Branch–Branch Junctions in Opuntia ficus-indica and Cylindropuntia bigelovii: A Comparative Study Supported by Mechanical Tissue Quantification" Plants 10, no. 11: 2313. https://doi.org/10.3390/plants10112313
APA StyleMylo, M. D., Hesse, L., Masselter, T., Leupold, J., Drozella, K., Speck, T., & Speck, O. (2021). Morphology and Anatomy of Branch–Branch Junctions in Opuntia ficus-indica and Cylindropuntia bigelovii: A Comparative Study Supported by Mechanical Tissue Quantification. Plants, 10(11), 2313. https://doi.org/10.3390/plants10112313