
Comparative Analysis of the Effect of Carbon- and Titanium-Ions Irradiation on Morpho-Anatomical and Biochemical Traits of Dolichos melanophthalmus DC. Seedlings Aimed to Space Exploration





Abstract
:1. Introduction
2. Results
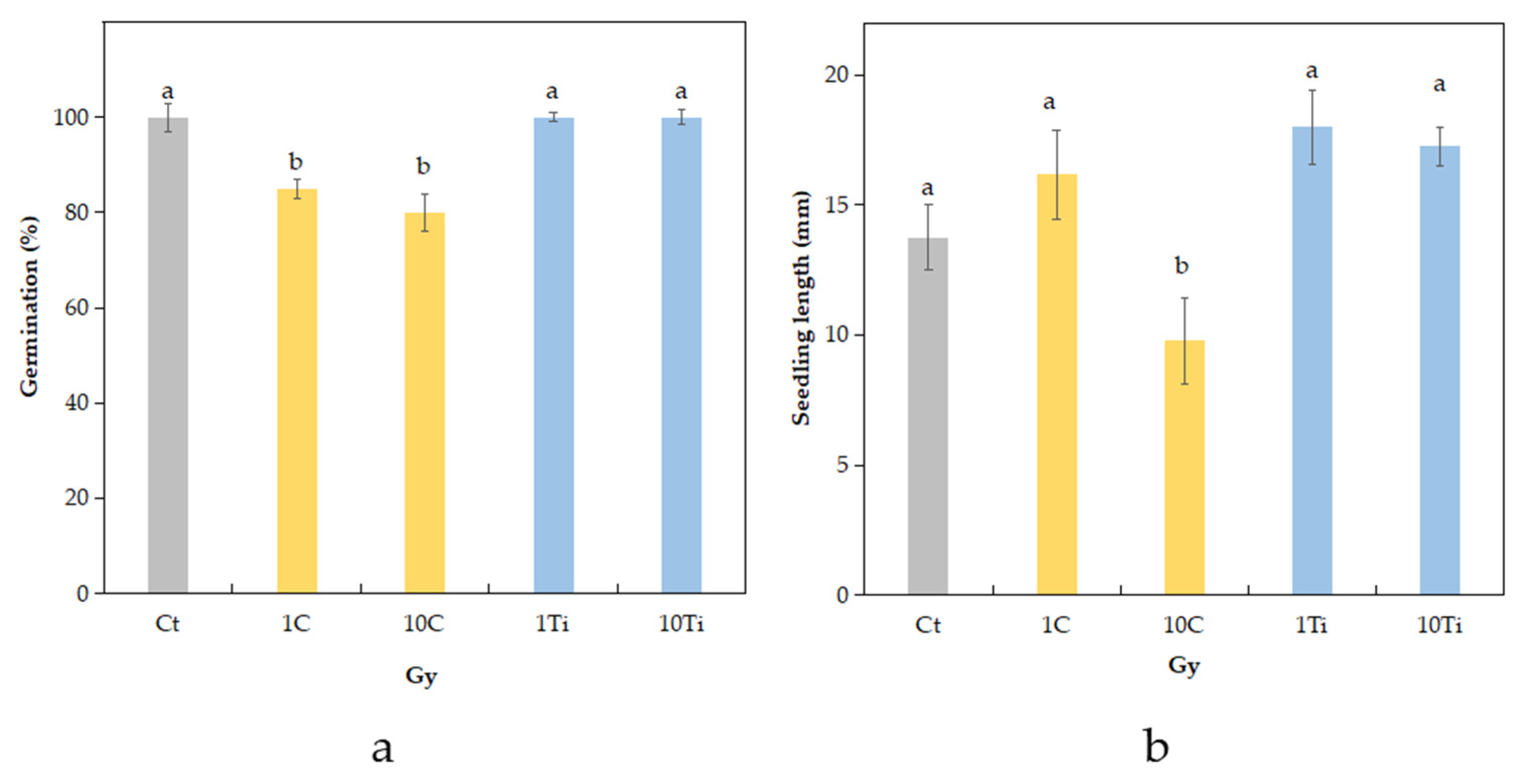
2.1. Germination Rate and Seedling Length
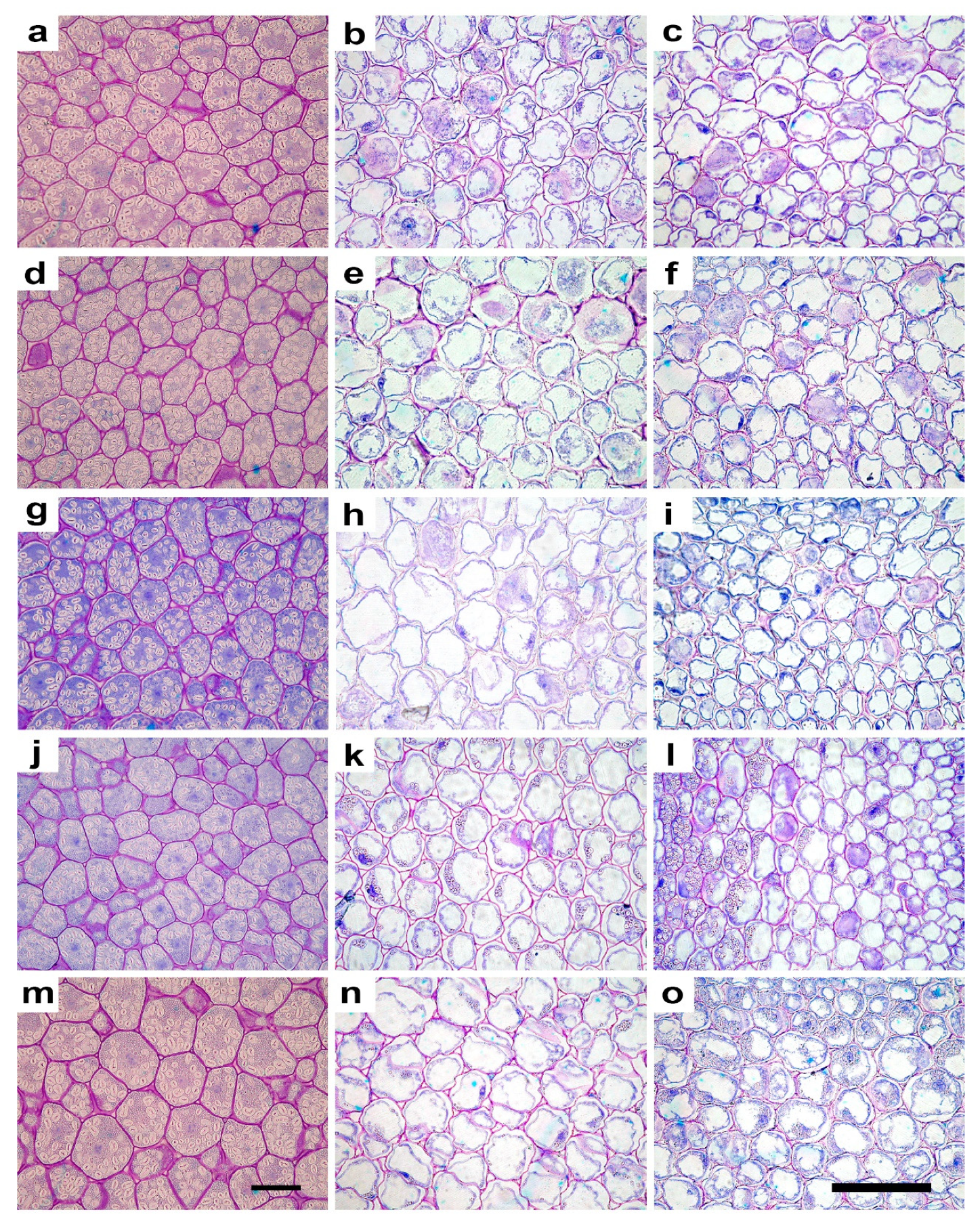
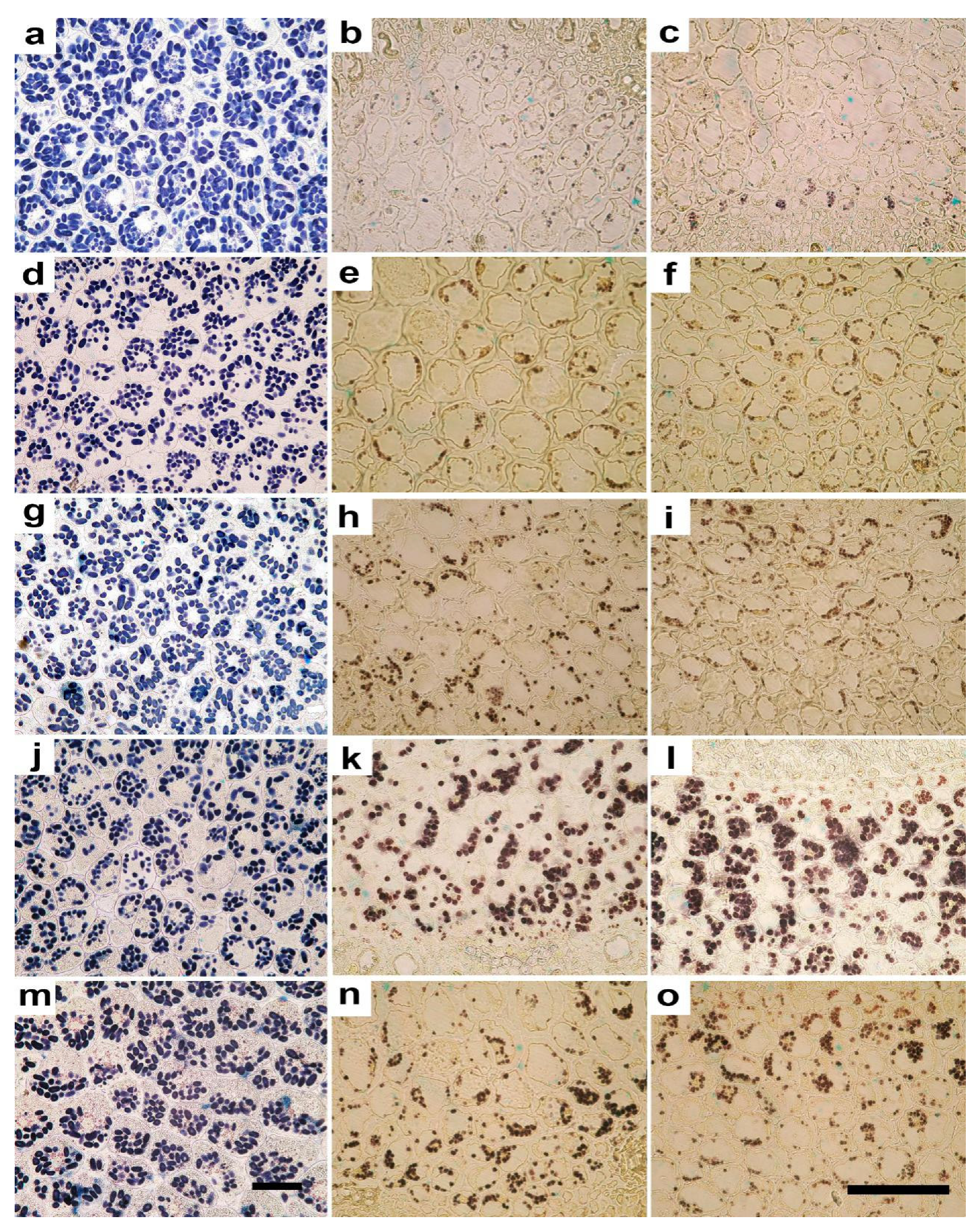
2.2. Anatomy
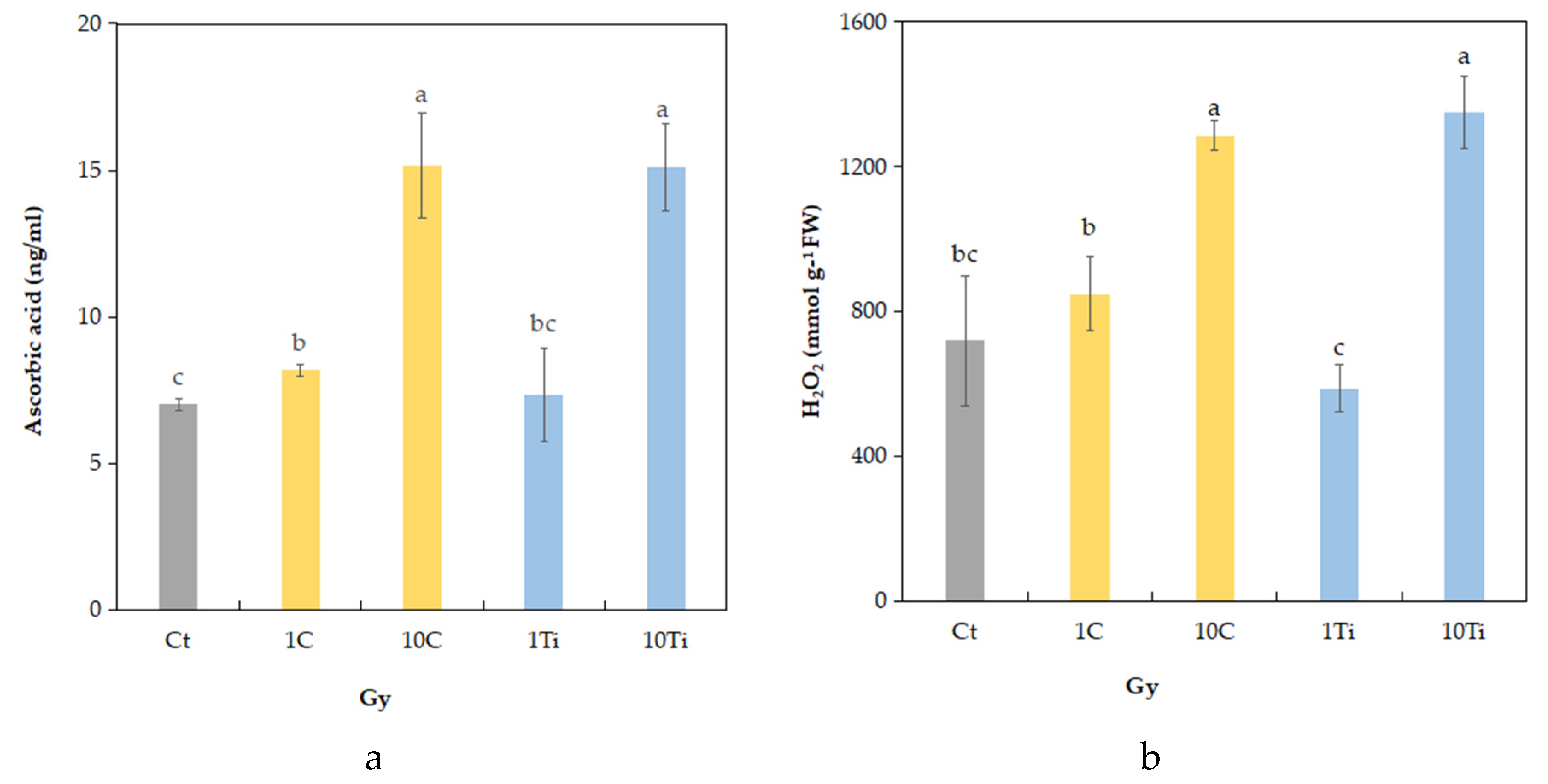
2.3. Biochemical Traits
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Seed Germination and Morphology
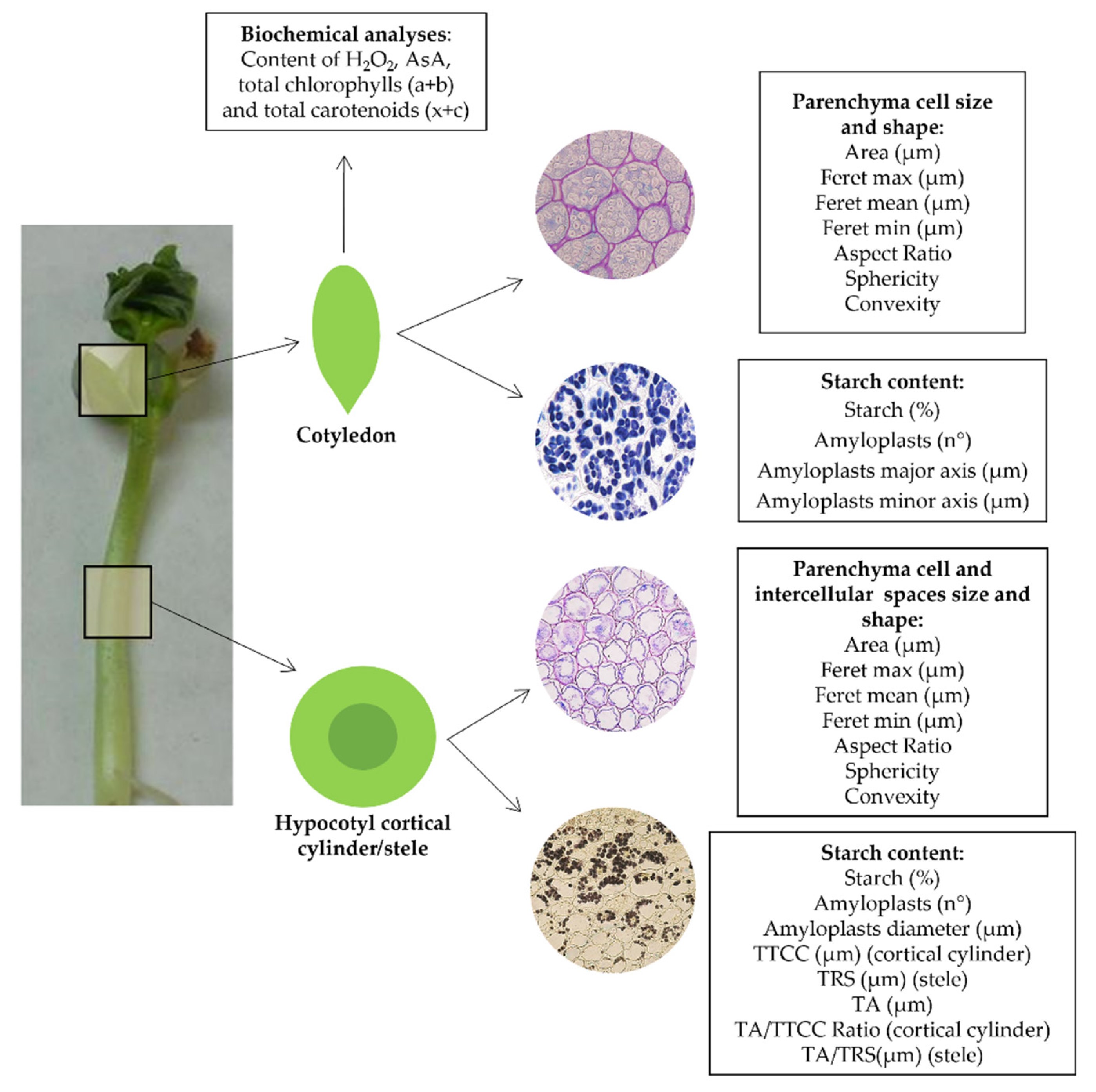
4.3. Light Microscopy and Digital Image Analysis
4.4. Biochemical Analyses
4.5. Data Elaboration
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Portree, D.S. Humans to Mars: Fifty Years of Mission Planning, 1950–2000; National Aeronautics and Space Administration: Washington, DC, USA, 2001. [Google Scholar]
- Wheeler, R.M. Crop Production for Advanced Life Support Systems-Observations from the Kennedy Space Center Breadboard Project; NASA: Washington, DC, USA, 2003. [Google Scholar]
- De Pascale, S.; Arena, C.; Aronne, G.; De Micco, V.; Pannico, A.; Paradiso, R.; Rouphael, Y. Biology and crop production in Space environments: Challenges and opportunities. Life Sci. Space Res. 2021, 29, 30–37. [Google Scholar] [CrossRef]
- Mitchell, C.A. Bioregenerative life-support systems. Am. J. Clin. Nutr. 1994, 60, 820S–824S. [Google Scholar] [CrossRef]
- Paradiso, R.; De Micco, V.; Buonomo, R.; Aronne, G.; Barbieri, G.; De Pascale, S. Soilless cultivation of soybean for Bioregenerative Life-Support Systems: A literature review and the experience of the MELiSSA Project—Food characterisation Phase I. Plant Biol. 2014, 16, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Nelson, P.E. Conditions and constraints of food processing in space. Food Technol. 1994, 48, 113–204. [Google Scholar]
- De Micco, V.; Arena, C.; Pignalosa, D.; Durante, M. Effects of sparsely and densely ionizing radiation on plants. Rad. Environ. Biophys. 2011, 50, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; De Micco, V.; Macaeva, E.; Quintens, R. Space radiation effects on plant and mammalian cells. Acta Astronaut. 2014, 104, 419–431. [Google Scholar] [CrossRef]
- Durante, M. Space radiation protection: Destination Mars. Life Sci. Space Res. 2014, 1, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Nagamatsu, A.; Nenoi, M.; Fujimori, A.; Kakinuma, S.; Katsube, T.; Wang, B.; Tsuruoka, C.; Shirai, T.; Nakamura, A.J.; et al. Space Radiation Biology for “Living in Space”. Biomed. Res. Int. 2020, 2020, 4703286. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.A. Elemental and isotopic composition of the galactic cosmic rays. Annu. Rev. Nucl. Part. Sci. 1983, 33, 323–382. [Google Scholar] [CrossRef]
- Zeitlin, C.; Hassler, D.M.; Cucinotta, F.A.; Ehresmann, B.; Wimmer-Schweingruber, R.F.; Brinza, D.E.; Kang, S.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Measurements of energetic particle radiation in transit to Mars on the Mars Science Laboratory. Science 2013, 340, 1080–1084. [Google Scholar] [CrossRef] [Green Version]
- Norbury, J.W.; Schimmerling, W.; Slaba, T.C.; Azzam, E.I.; Badavi, F.F.; Baiocco, G.; Benton, E.; Bindi, V.; Blakely, E.A.; Blattnig, S.R.; et al. Galactic cosmic ray simulation at the NASA Space Radiation Laboratory. Life Sci. Space Res. 2016, 8, 38–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, M.J.; Kim, D.Y.; Ahn, J.W.; Kang, S.Y.; Seo, Y.W.; Kim, J.B. Comparison of radiosensitivity response to acute and chronic gamma irradiation in colored wheat. Genet. Mol. Biol. 2018, 41, 611–623. [Google Scholar] [CrossRef] [Green Version]
- Kovalchuk, I.; Molinier, J.; Yao, Y.L.; Arkhipov, A.; Kovalchuk, O. Transcriptome analysis reveals fundamental differences in plant response to acute and chronic exposure to ionizing radiation. Mutat. Res. 2007, 624, 101–113. [Google Scholar] [CrossRef]
- Simonsen, L.C.; Slaba, T.C.; Guida, P.; Rusek, A. NASA’s first ground-based Galactic Cosmic Ray Simulator: Enabling a new era in space radiobiology research. PLoS Biol. 2020, 18, e3000669. [Google Scholar] [CrossRef]
- Schuy, C.; Weber, U.; Durante, M. Hybrid active-passive space radiation simulation concept for GSI and the future FAIR facility. Front. Phys. 2020, 8, 337. [Google Scholar] [CrossRef]
- Le Caër, S. Water radiolysis: Influence of oxide surfaces on H2 production under ionizing radiation. Water 2011, 3, 235–253. [Google Scholar] [CrossRef] [Green Version]
- Caplin, N.; Willey, N. Ionizing radiation, higher plants, and radioprotection: From acute high doses to chronic low doses. Front. Plant Sci. 2018, 9, 847. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; De Micco, V.; Aronne, G.; Pugliese, M.G.; Virzo, A.; DeMaio, A. Response of Phaseolus vulgaris L. plants to low-LET ionizing radiation: Growth and oxidative stress. Acta Astronaut. 2014, 91, 107–114. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; da Silva, J.A.T.; Fujita, M. Plant Response and Tolerance to Abiotic Oxidative Stress: Antioxidant Defense is a Key Factor. In Crop Stress and Its Management: Perspectives and Strategies; Venkateswarlu, B., Shanker, A.K., Shanker, C., Maheswari, M., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 261–315. ISBN 978-94-007-2220-0. [Google Scholar]
- Gudkov, S.V.; Grinberg, M.A.; Sukhov, V.; Vodeneev, V. Effect of ionizing radiation on physiological and molecular processes in plants. J. Environ. Radioact. 2019, 202, 8–24. [Google Scholar] [CrossRef]
- Medina, F.J.; Herranz, R.; Arena, C.; Aronne, G.; De Micco, V. Growing Plants under Generated Extra-Terrestrial Environments: Effects of Altered Gravity and Radiation. Gener. Appl. Extra-Terr. Environ. Earth 2015, 235–250. [Google Scholar]
- Endo, T.R.; Gill, B.S. The Deletion Stocks of Common Wheat. J. Hered. 1996, 87, 295–307. [Google Scholar] [CrossRef]
- Kim, J.-H.; Moon, Y.R.; Lee, M.H.; Kim, J.H.; Wi, S.G.; Park, B.J.; Kim, C.S.; Chung, B.Y. Photosynthetic capacity of Arabidopsis plants at the reproductive stage tolerates gamma irradiation. J. Radiat. Res. 2011, 52, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Wi, S.G.; Chung, B.Y.; Kim, J.H.; Baek, M.H.; Yang, D.H.; Lee, J.W.; Kim, J.S. Ultrastructural changes of cell organelles in Arabidopsis stems after gamma irradiation. J. Plant Biol. 2005, 48, 195–200. [Google Scholar] [CrossRef]
- Arena, C.; De Micco, V.; De Maio, A. Growth alteration and leaf biochemical responses in Phaseolus Vulgaris exposed to different doses of ionizing radiation. Plant Biol. 2014, 16, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Shikazono, N.; Tanaka, A.; Kitayama, S.; Watanabe, H.; Tano, S. LET dependence of lethality in Arabidopsis thaliana irradiated by heavy ions. Rad. Environ. Biophys. 2002, 41, 159–162. [Google Scholar] [CrossRef]
- Wei, L.J.; Yang, Q.; Xia, H.M.; Furusawa, Y.; Guan, S.H.; Xin, P.; Sun, Y.Q. Analysis of cytogenetic damage in rice seeds induced by energetic heavy ions on-ground and after spaceflight. J. Radiat. Res. 2006, 47, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; Turano, M.; Mele, B.H.; Cataletto, P.; Furia, M.; Pugliese, M.; De Micco, V. Anatomy, photochemical activity, and DNA polymorphism in leaves of dwarf tomato irradiated with X-rays. Biol. Plant. 2017, 61, 305–314. [Google Scholar] [CrossRef]
- Ling, A.P.K.; Ung, Y.C.; Hussein, S.; Harun, A.R.; Tanaka, A.; Yoshihiro, H. Morphological and biochemical responses of Oryza sativa L. (cultivar MR219) to ion beam irradiation. J. Zhejiang Univ. Sci. B 2013, 14, 1132–1143. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, Z.; Zhou, L.; Qu, Y.; Lu, D.; Yu, L.; Du, Y.; Jin, W.; Li, W. Relationship between plant growth and cytological effect in root apical meristem after exposure of wheat dry seeds to carbon ion beams. Nucl. Instrum. Methods Phys. Res. Sect. B 2013, 305, 9–15. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, E.; Hay Mele, B.; Cataletto, P.R.; Turano, M.; Simoniello, P.; De Micco, V. Suitability of Solanum lycopesicum L. ‘Microtom’ for growth in Bioregenerative Life Support Systems: Exploring the effect of high-LET ionising radiation on photosynthesis, leaf structure and fruit trait. Plant Biol. 2019, 21, 615–626. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; de Pascale, S.; Paradiso, R.; Aronne, G. Microgravity effects on different stages of higher plant life cycle and completion of the seed-to-seed cycle. Plant Biol. 2014, 16, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Amitrano, C.; Arena, C.; De Pascale, S.; De Micco, V. Light and Low Relative Humidity Increase Antioxidants Content in Mung Bean (Vigna radiata L.) Sprouts. Plants 2020, 9, 1093. [Google Scholar] [CrossRef]
- De Micco, V.; Amitrano, C.; Vitaglione, P.; Ferracane, R.; Pugliese, M.; Arena, C. Effect of light quality and ionising radiation on morphological and nutraceutical traits of sprouts for astronauts’ diet. Acta Astronaut. 2021, 185, 188–197. [Google Scholar] [CrossRef]
- Gommers, C.M.M.; Monte, E. Seedling Establishment: A Dimmer Switch-Regulated Process between Dark and Light Signaling. Plant Physiol. 2018, 176, 1061–1074. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, S.; Kumar, M.; Kumar, P.; Gupta, V.K.; Singhal, R.K.; Yadav, A.; Singh, B. Metabolic and biochemical changes caused by gamma irradiation in plants. J. Radioanal. Nucl. Chem. 2014, 300, 199–212. [Google Scholar] [CrossRef]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, Oxidative Damage and Oxygen Deprivation Stress: A Review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, E.; Vitale, L.; Costanzo, G.; Velikova, V.; Tsonev, T.; Simoniello, P.; De Micco, V.; Arena, C. Light Spectral Composition Influences Structural and Eco-Physiological Traits of Solanum lycopersicum L. cv.‘Microtom’ in Response to High-LET Ionizing Radiation. Plants 2021, 10, 1752. [Google Scholar] [CrossRef]
- Humphries, T.; Chauhan, B.S.; Florentine, S.K. Environmental factors effecting the germination and seedling emergence of two populations of an aggressive agricultural weed; Nassella trichotoma. PLoS ONE 2018, 13, e0199491. [Google Scholar] [CrossRef]
- De Micco, V.; Paradiso, R.; Aronne, G.; De Pascale, S.; Quarto, M.; Arena, C. Leaf anatomy and photochemical behaviour of Solanum lycopersicum L. plants from seeds irradiated with low-LET ionising radiation. Sci. World J. 2014, 2014, 428141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hase, Y.; Yamaguchi, M.; Inoue, M.; Tanaka, A. Reduction of survival and induction of chromosome aberrations in tobacco irradiated by carbon ions with different linear energy transfers. Int. J. Radiat. Biol. 2002, 78, 799–806. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Arena, C.; Aronne, G. Anatomical alterations of Phaseolus vulgaris L. mature leaves irradiated with X-rays. Plant Biol. 2014, 16, 187–193. [Google Scholar] [CrossRef]
- Skagen, E.B.; Iversen, T.-H. Simulated weightlessness and hyper-g results in opposite effects on the regeneration of the cortical microtubule array in protoplasts from Brassica napus hypocotyls. Physiol. Plant. 1999, 106, 318–325. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Aronne, G.; Joseleau, J.P.; Ruel, K. Xylem development and cell wall changes of soybean seedlings grown in space. Ann. Bot. 2008, 101, 661–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, D.J. Plant cell growth and elongation. eLS 2014. [Google Scholar] [CrossRef]
- Shigo, A. Compartmentalization: A conceptual framework for understanding how trees grow and defend themselves. Annu. Rev. Phytopathol 1984, 22, 189–214. [Google Scholar] [CrossRef]
- Streb, S.; Zeeman, S.C. Starch metabolism in Arabidopsis. Am. Soc. Plant Biol. 2012, 10, e0160. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Beckles, D.M. Dynamic changes in the starch-sugar interconversion within plant source and sink tissues promote a better abiotic stress response. J. Plant Physiol. 2019, 234, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Krasavina, M.S.; Burmistrova, N.A.; Raldugina, G.N. The role of carbohydrates in plant resistance to abiotic stresses. In Emerging Technologies and Management of Crop Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2014; pp. 229–270. [Google Scholar]
- Todaka, D.; Matsushima, H.; Morohashi, Y. Water stress enhances β-amylase activity in cucumber cotyledons. J. Exp. Bot. 2000, 51, 739–745. [Google Scholar] [CrossRef]
- Rosa, M.; Hilal, M.; González, J.A.; Prado, F.E. Low-temperature effect on enzyme activities involved in sucrose-starch partitioning in salt-stressed and salt-acclimated cotyledons of quinoa (Chenopodium quinoa Willd.) seedlings. Plant Physiol. Biochem. 2009, 47, 300–307. [Google Scholar] [CrossRef]
- Chen, H.-J.; Chen, J.-Y.; Wang, S.-J. Molecular regulation of starch accumulation in rice seedling leaves in response to salt stress. Acta Physiol. Plant. 2008, 30, 135–142. [Google Scholar] [CrossRef]
- Dkhil, B.B.; Denden, M. Salt stress induced changes in germination, sugars, starch and enzyme of carbohydrate metabolism in Abelmoschus esculentus L. (Moench.) seeds. Afr. J. Agric. Res. 2010, 5, 408–415. [Google Scholar]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, H.; Wu, J.; Xie, F. Effect of Drought Stress during Soybean R2–R6 Growth Stages on Sucrose Metabolism in Leaf and Seed. Int. J. Mol. Sci. 2020, 21, 618. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Lihavainen, J.; Keinänen, M.; Keski-Saari, S.; Kontunen-Soppela, S.; Sõber, A.; Oksanen, E. Artificially decreased vapour pressure deficit in field conditions modifies foliar metabolite profiles in birch and aspen. J. Exp. Bot. 2016, 67, 4367–4378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.W.; Montagu, M.V.; Inze, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N.; Benzie, I.J.J.; Strain, J.J.; Favell, D.; Fletcher, J. Plant L-ascorbic acid: Chemistry, function, metabolism, bioavailability and effects of processing. J. Sci. Food Agric. 2000, 80, 825–860. [Google Scholar] [CrossRef]
- Amitrano, C.; Rouphael, Y.; De Pascale, S.; De Micco, V. Modulating vapor pressure deficit in the plant micro-environment may enhance the bioactive value of lettuce. Horticulturae 2021, 7, 32. [Google Scholar] [CrossRef]
- Rosales, M.A.; Ruiz, J.M.; Hernández, J.; Soriano, T.; Castilla, N.; Romero, L. Antioxidant content and ascorbate metabolism incherry tomato exocarp in relation to temperature and solar radiation. J. Sci. Food Agric. 2006, 86, 1545–1551. [Google Scholar] [CrossRef]
- Islamian, J.P.; Mehrali, H. Lycopene as a carotenoid provides radioprotectant and antioxidant effects by quenching radiation-induced free radical singlet oxygen: An overview. Cell J. 2015, 16, 386–391. [Google Scholar]
- Viuda-Martos, M.; Sanchez-Zapata, E.; Sayas-Barbera, E.; Sendra, E.; Perez-Alvarez, J.A.; Fernandez-Lopez, J. Tomato and tomato byproducts. Human benefits of lycopene and its application to meat products: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1032–1049. [Google Scholar] [CrossRef]
- Mak, C.; Yap, T.C. Inheritance of seed protein content and other agronomic characters in long bean (Vigna sesquipedalis Fruw.). Theor. Appl. Genet. 1980, 56, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Kazama, Y.; Hirano, T.; Saito, H.; Liu, Y.; Ohbu, S.; Hayashi, Y.; Abe, T. Characterization of highly efficient heavy-ion mutagenesis in Arabidopsis thaliana. BMC Plant Biol. 2011, 11, 161. [Google Scholar] [CrossRef] [Green Version]
- Reale, L.; Gigante, D.; Landucci, F.; Ferranti, F.; Venanzoni, R. Morphological and histo-anatomical traits reflect die-back in Phragmites australis (Cav.) Steud. Aquat. Bot. 2012, 103, 122–128. [Google Scholar] [CrossRef]
- Ruzin, S.E. Sectioning and mounting. In Plant Microtechnique and Microscopy; Oxford University Press: Oxford, UK, 1999; pp. 73–85. [Google Scholar]
- Van Buggenhout, S.; Grauwet, T.; Van Loey, A. Structure/processing relation of vacuum infused strawberry tissue frozen under different conditions. Eur. Food Res. Technol. 2008, 226, 437–448. [Google Scholar] [CrossRef]
- Sergiev, L.; Alexieva, V.; Karanova, E. Effect of spermine, atrazine and combination between them on some endogenous protective systems and stress markers in plants. Comp. Ren. Acad. Bulg. Sci. 1997, 51, 121–124. [Google Scholar]
- Vitale, E.; Velikova, V.; Tsonev, T.; Ferrandino, I.; Capriello, T.; Arena, C. The Interplay between Light Quality and Biostimulant Application Affects the Antioxidant Capacity and Photosynthetic Traits of Soybean (Glycine max L. Merrill). Plants 2021, 10, 861. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cotyledons | ||||||
|---|---|---|---|---|---|---|
| Ct | 1C | 10C | 1Ti | 10Ti | S | |
| Parenchyma Cells | ||||||
| Area (µm) | 5243 ± 180.9 b | 4631 ± 267.9 b | 6740 ± 278.9 a | 4947 ± 201.2 b | 6928 ± 374.5 a | *** |
| Feret max (µm) | 99.68 ± 2.075 b | 98.81 ± 3.413 b | 113.0 ± 3.079 a | 95.13 ± 2.205 b | 112.5 ± 2.625 a | *** |
| Feret mean (µm) | 87.36 ± 1.491 b | 84.56 ± 2.561 b | 98.73 ± 2.295 a | 84.94 ± 1.726 b | 99.26 ± 2.425 a | *** |
| Feret min (µm) | 72.94 ± 1628 b | 67.24 ± 1962 b | 81.28 ± 2067 a | 72.10 ± 1835 b | 83.15 ± 2615 a | *** |
| Aspect Ratio | 1.341 ± 0.037 a | 1.457 ± 0.041 a | 1.385 ± 0.038 a | 1.311 ± 0.033 a | 1.363 ± 0.034 a | NS |
| Sphericity | 0.595 ± 0.035 a | 0.526 ± 0.034 a | 0.550 ± 0.028 a | 0.589 ± 0.032 a | 0.571 ± 0.029 a | NS |
| Convexity | 0.957 ± 0.007 a | 0.904 ± 0.021 c | 0.947 ± 0.007 ab | 0.936 ± 0.020 b | 0.952 ± 0.008 ab | *** |
| Starch (%) | 49.03 ± 0.009 a | 46.66 ± 0.023 a | 34.67 ± 0.010 b | 35.46 ± 0.024 b | 35.71 ± 0.011 b | *** |
| Amyloplasts (n°) | 20.78 ± 0.697 b | 23.35 ± 0.962 a | 20.28 ± 0.801 b | 18.67 ± 0.914 b | 20.58 ± 1024 b | * |
| Amyloplasts major axis (µm) | 24.73 ± 0.556 a | 20.57 ± 0.484 c | 23.91 ± 0.450 ab | 22.58 ± 0.594 b | 23.05 ± 0.459 b | *** |
| Amyloplasts minor axis (µm) | 13.01 ± 0.316 a | 11.05 ± 0.238 b | 12.91 ± 0.244 a | 11.70 ± 0.364 b | 12.76 ± 0.330 a | *** |
| Hypocotyls—Cortical Cylinder | ||||||
|---|---|---|---|---|---|---|
| Parameters | Ct | 1C | 10C | 1Ti | 10Ti | S |
| Parenchyma Cells | ||||||
| Area (µm) | 1086 ± 56.57 ab | 1217 ± 87.22 a | 665.2 ± 54.66 c | 933.9 ± 80.77 b | 1145 ± 110.6 ab | *** |
| Feret max (µm) | 43.96 ± 1.298 a | 44.32 ± 1.427 a | 35.79 ± 1.451 b | 42.03 ± 2.124 a | 43.27 ± 2.071 a | ** |
| Feret mean (µm) | 39.41 ± 1.027 a | 40.81 ± 1.387 a | 31.43 ± 1.213 b | 36.57 ± 1.701 a | 39.43 ± 1.880 a | *** |
| Feret min (µm) | 33.79 ± 0.863 ab | 36.44 ± 1.394 a | 26.13 ± 1.260 c | 30.51 ± 1.375 b | 34.82 ±1.742 a | *** |
| Aspect Ratio | 1.271 ± 0.027 ab | 1.196 ± 0.021 b | 1.366 ± 0.065 a | 1.334 ± 0.025 a | 1.222 ± 0.022 b | ** |
| Sphericity | 0.668 ± 0.032 ab | 0.731 ± 0.030 a | 0.619 ± 0.051 b | 0.586 ± 0.027 b | 0.680 ± 0.029 ab | * |
| Convexity | 0.942 ± 0.009 ab | 0.949 ± 0.009 a | 0.924 ± 0.013 c | 0.930 ± 0.012 bc | 0.946 ± 0.014 a | ** |
| Starch (%) | 0.782 ± 0.016 d | 1.558 ± 0.021 cd | 2.879 ± 0.012 c | 12.93 ± 0.028 a | 6.416 ± 0.013 b | *** |
| Amyloplasts (n°) | 7.730 ± 0.727 bc | 5.780 ± 1.145 c | 7.700 ± 1.356 bc | 12.33 ± 0.702 a | 10.73 ± 0.794 ab | *** |
| Amyloplasts diameter (µm) | 2.082 ± 0.116 d | 2.872 ± 0.114 c | 2.339 ± 0.197 d | 5.236 ± 0.216 a | 3.424 ± 0.180 b | *** |
| TTCC (µm) | 364.1 ± 8.804 b | 407.7 ± 11.30 a | 328.5 ± 11.94 c | 357.7 ± 9.420 bc | 421.2 ± 15.37 a | *** |
| TA (µm) | 79.83 ± 20.72 c | 141.3 ± 42.86 bc | 96.09 ± 25.47 c | 263.0 ± 12.96 a | 222.9 ± 42.28 ab | ** |
| TA/TTCC Ratio | 0.304 ± 0.064 c | 0.702 ± 0.018 a | 0.291 ± 0.079 c | 0.734 ± 0.026 a | 0.510 ± 0.084 b | *** |
| Intercellular Spaces | ||||||
| Area (µm) | 13.73 ± 1.529 a | 21.38 ± 4.501 a | 17.28 ± 3.808 a | 12.44 ± 1.683 a | 12.54 ± 1.858 a | NS |
| Feret max (µm) | 6.697 ± 0.546 a | 8.082 ± 0.933 a | 7.572 ± 1.268 a | 6.409 ± 0.644 a | 5.930 ± 0.391 a | NS |
| Feret mean (µm) | 5.606 ± 0.424 a | 6.646 ± 0.731 a | 6.021 ± 0.921 a | 5.290 ± 0.456 a | 4.919 ± 0.355 a | NS |
| Feret min (µm) | 4.199 ± 0.276 a | 4.753 ± 0.441 a | 3.729 ± 0.388 a | 3.788 ± 0.258 a | 3.576 ± 0.335 a | NS |
| Aspect Ratio | 1.594 ± 0.059 a | 1.687 ± 0.066 a | 1.907 ± 0.159 a | 1.677 ± 0.130 a | 1.842 ± 0.168 a | NS |
| Sphericity | 0.430 ± 0.038 a | 0.402 ± 0.047 a | 0.350 ± 0.054 a | 0.414 ± 0.048 a | 0.365 ± 0.052 a | NS |
| Convexity | 0.824 ± 0.026 a | 0.817 ± 0.027 a | 0.864 ± 0.039 a | 0.829 ± 0.030 a | 0.873 ±0.025 a | NS |
| Hypocotyls—Stele | ||||||
|---|---|---|---|---|---|---|
| Parameters | Ct | 1C | 10C | 1Ti | 10Ti | S |
| Parenchyma Cells | ||||||
| Area (µm) | 1337 ± 125.1 a | 1291 ± 72.56 a | 1329 ± 124.6 a | 1321 ± 114.7 a | 1702 ± 171.6 a | NS |
| Feret max (µm) | 48.04 ± 2.332 b | 47.12 ± 1.362 b | 51.75 ± 1.595 ab | 47.27 ± 1.986 b | 55.02 ± 2.614 a | * |
| Feret mean (µm) | 42.85 ± 2.026 a | 42.61 ± 1.186 a | 44.26 ± 1.722 a | 43.04 ± 1.806 a | 48.68 ± 2.309 a | NS |
| Feret min (µm) | 36.40 ± 1.730 a | 38.24 ± 1.161a | 35.51 ± 1.957 a | 37.75 ± 1.767 a | 41.34 ± 2.140 a | NS |
| Aspect Ratio | 1.297 ± 0.031 b | 1.220 ± 0.022 b | 1.491 ± 0.065 a | 1.248 ± 0.035 b | 1.328 ± 0.035 b | *** |
| Sphericity | 0.642 ± 0.033 ab | 0.726 ± 0.030 a | 0.506 ± 0.041 c | 0.677 ± 0.032 ab | 0.586 ± 0.026 bc | *** |
| Convexity | 0.939 ± 0.008 a | 0.943 ± 0.008 a | 0.927 ± 0.010 a | 0.932 ± 0.007 a | 0.940 ± 0.018 a | NS |
| Starch (%) | 1.598 ± 0.007 c | 1.240 ± 0.019 c | 2.708 ± 0.014 c | 10.58 ± 0.033 a | 5.581 ± 0.023 b | *** |
| Amyloplasts (n°) | 4.380 ± 0.584 b | 5.080 ± 0.742 b | 6.000 ± 1.006 b | 12.57 ± 1.155 a | 10.33 ± 1.103 a | *** |
| Amyl. diameter (µm) | 2.935 ± 0.144 c | 2.662 ± 0.109 c | 2.394 ± 0.136 c | 4.383 ± 0.287 a | 3.504 ± 0.204 b | *** |
| TRS (µm) | 283.2 ± 27.67 b | 282.0 ± 11.97 b | 279.7 ± 14.41 b | 319.6 ± 11.12 ab | 356.6 ± 13.03 a | * |
| TA (µm) | 180.3 ± 38.67 bc | 173.8 ± 38.44 bc | 136.3 ± 34.33 c | 275.8 ± 26.48 ab | 326.3 ± 24.79 a | ** |
| TA/TRS Ratio | 0.715 ± 0.083 ab | 0.826 ± 0.096 a | 0.504 ± 0.126 b | 0.862 ± 0.074 a | 0.907 ± 0.048 a | * |
| Intercellular Spaces | ||||||
| Area (µm) | 20.03 ± 2.552 a | 24.33 ± 5.209 a | 22.90 ± 4.320 a | 22.36 ± 3.965 a | 31.84 ± 4.413 a | NS |
| Feret max (µm) | 7.356 ± 0.574 a | 7.764 ± 0.617 a | 8.624 ± 1.070 a | 8.920 ± 1.445 a | 10.43 ± 0.834 a | NS |
| Feret mean (µm) | 6.231 ± 0.460 a | 6.576 ± 0.519 a | 7.076 ± 0.828 a | 7.232 ± 1.013 a | 8.417 ± 0.626 a | NS |
| Feret min (µm) | 4.640 ± 0.341 a | 4.893 ± 0.366 a | 5.020 ± 0.433 a | 5.074 ± 0.436 a | 5.712 ± 0.428 a | NS |
| Aspect Ratio | 1.605 ± 0.083 a | 1.610 ± 0.074 a | 1.681 ± 0.073 a | 1.641 ± 0.087 a | 1.865 ± 0.138 a | NS |
| Sphericity | 0.469 ± 0.050 a | 0.465 ± 0.043 a | 0.392 ± 0.043 a | 0.395 ± 0.045 a | 0.381 ± 0.052 a | NS |
| Convexity | 0.870 ± 0.026 a | 0.861 ± 0.021 a | 0.815 ± 0.031 a | 0.829 ± 0.033 a | 0.809 ± 0.023 a | NS |
| Chlorophyll and Carotenoid Content | ||||||
|---|---|---|---|---|---|---|
| Parameters | Ct | 1C | 10C | 1Ti | 10Ti | S |
| Chl a (mg/g) | 1.504 ± 0.081 a | 1.293 ± 0.036 b | 0.701 ± 0.033 d | 1.072 ± 0.030 c | 1.278 ± 0.071 b | *** |
| Chl b (mg/g) | 0.652 ± 0.048 a | 0.596 ± 0.068 a | 0.353 ± 0.024 b | 0.602 ± 0.021 a | 0.548 ±0.035 a | ** |
| Chl a + b (mg/g) | 2.157 ± 0.117 a | 1.889 ± 0.101 b | 1.054 ± 0.024 c | 1.673 ± 0.015 b | 1.826 ± 0.095 b | *** |
| x + c (mg/g) | 0.342 ± 0.037 a | 0.233 ± 0.024 b | 0.128 ±0.008 c | 0.231 ± 0.024 b | 0.282 ± 0.032 ab | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Micco, V.; De Francesco, S.; Amitrano, C.; Arena, C. Comparative Analysis of the Effect of Carbon- and Titanium-Ions Irradiation on Morpho-Anatomical and Biochemical Traits of Dolichos melanophthalmus DC. Seedlings Aimed to Space Exploration. Plants 2021, 10, 2272. https://doi.org/10.3390/plants10112272
De Micco V, De Francesco S, Amitrano C, Arena C. Comparative Analysis of the Effect of Carbon- and Titanium-Ions Irradiation on Morpho-Anatomical and Biochemical Traits of Dolichos melanophthalmus DC. Seedlings Aimed to Space Exploration. Plants. 2021; 10(11):2272. https://doi.org/10.3390/plants10112272
Chicago/Turabian StyleDe Micco, Veronica, Sara De Francesco, Chiara Amitrano, and Carmen Arena. 2021. "Comparative Analysis of the Effect of Carbon- and Titanium-Ions Irradiation on Morpho-Anatomical and Biochemical Traits of Dolichos melanophthalmus DC. Seedlings Aimed to Space Exploration" Plants 10, no. 11: 2272. https://doi.org/10.3390/plants10112272
APA StyleDe Micco, V., De Francesco, S., Amitrano, C., & Arena, C. (2021). Comparative Analysis of the Effect of Carbon- and Titanium-Ions Irradiation on Morpho-Anatomical and Biochemical Traits of Dolichos melanophthalmus DC. Seedlings Aimed to Space Exploration. Plants, 10(11), 2272. https://doi.org/10.3390/plants10112272