
Col11a1 Regulates Bone Microarchitecture during Embryonic Development



Abstract
:1. Introduction
2. Experimental Section
2.1. Mice
2.2. Micro-CT Analysis
2.3. Trichrome Stain
2.4. Data Analysis
3. Results and Discussion
3.1. Changes to Embryonic Skeleton in the Absence of Col11a1 Expression
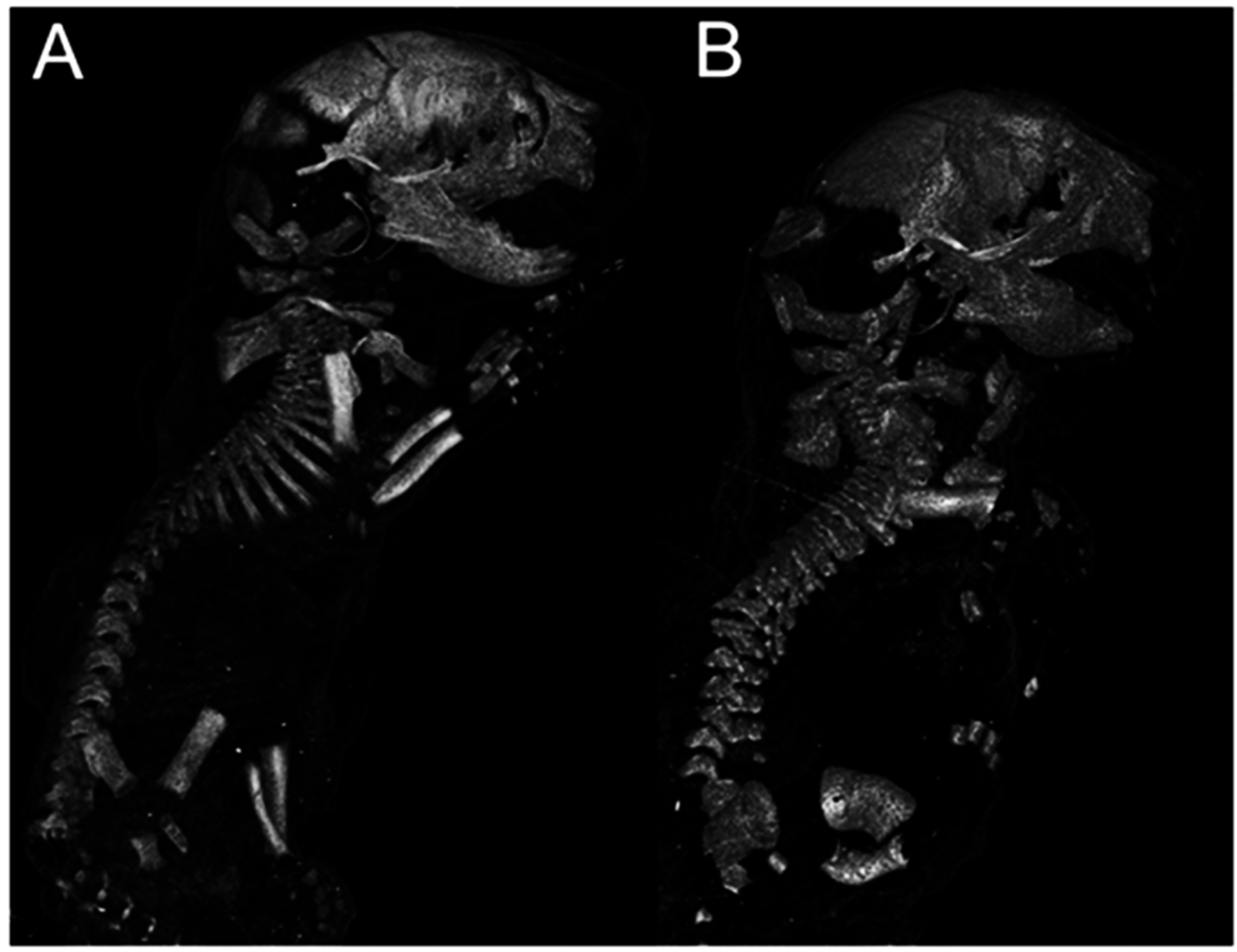
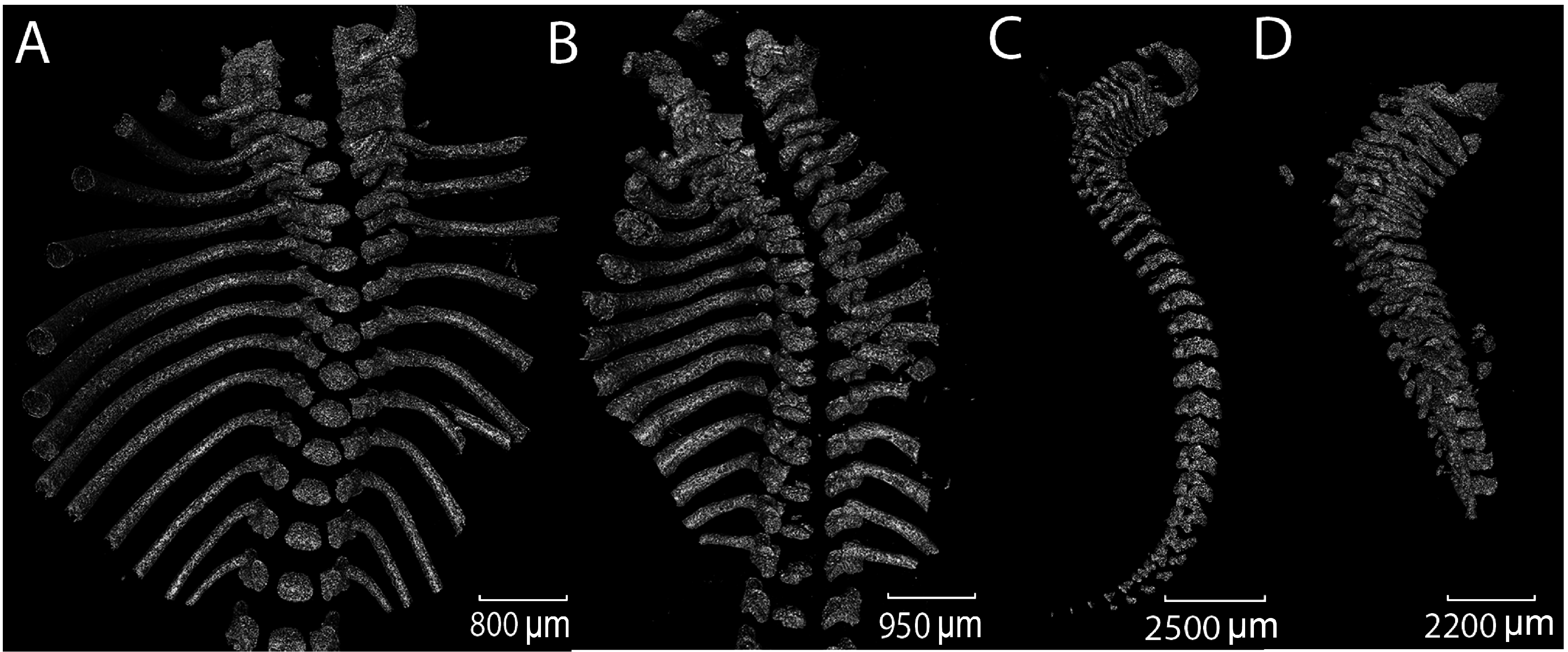
3.2. Analysis of Vertebrae

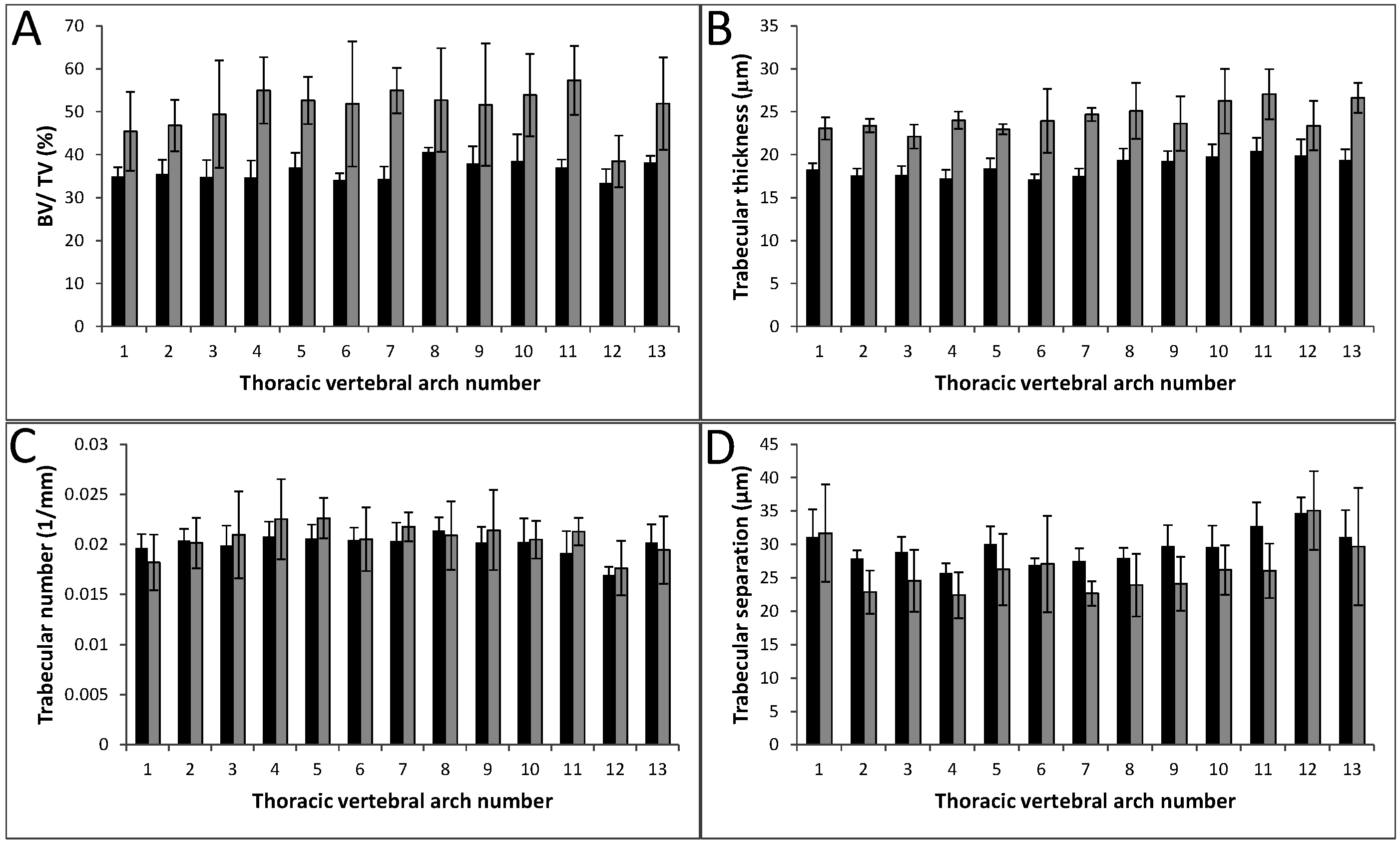
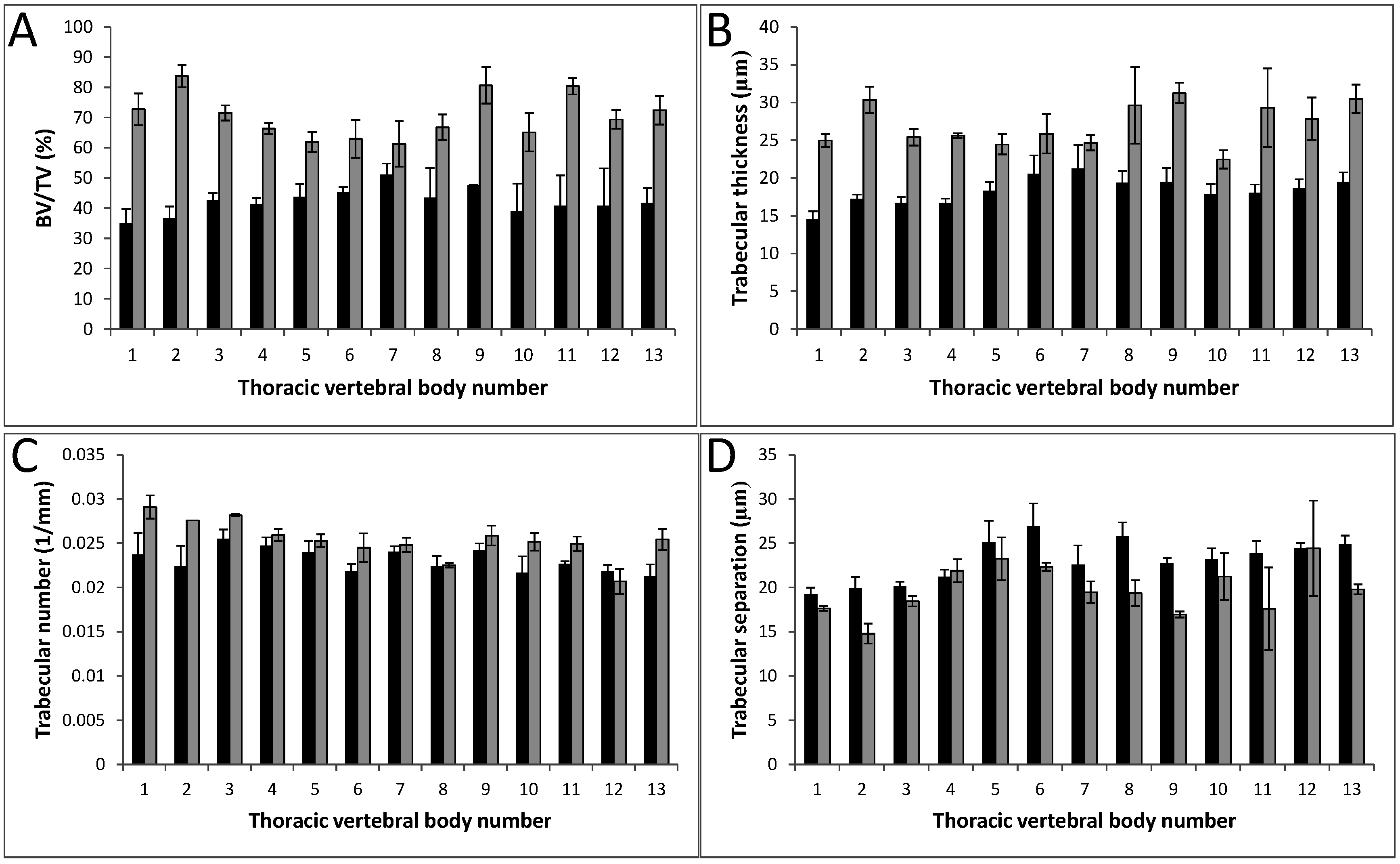
3.3. Bone Microarchitectural Parameters Dependent upon the Expression of Col11a1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BV/TV (%) | Tb.Th (µm) | Tb.N (1/mm) | Tb.Sp (µm) | |
|---|---|---|---|---|
| % difference | +31.7 | +32.8 | −1.01 | +0.03 |
| SD | 3.9 | 3.4 | 1.0 | 1.0 |
| p < 0.05 | p < 0.05 | ns | ns |
| BV/TV (%) | Tb.Th (µm) | Tb.N (1/mm) | Tb.Sp (µm) | |
|---|---|---|---|---|
| % difference | +80.4 | +67.2 | +8.26 | −17 |
| SD | 8.5 | 7.9 | 1.0 | 2.0 |
| p < 0.05 | p < 0.05 | ns | p < 0.05 |
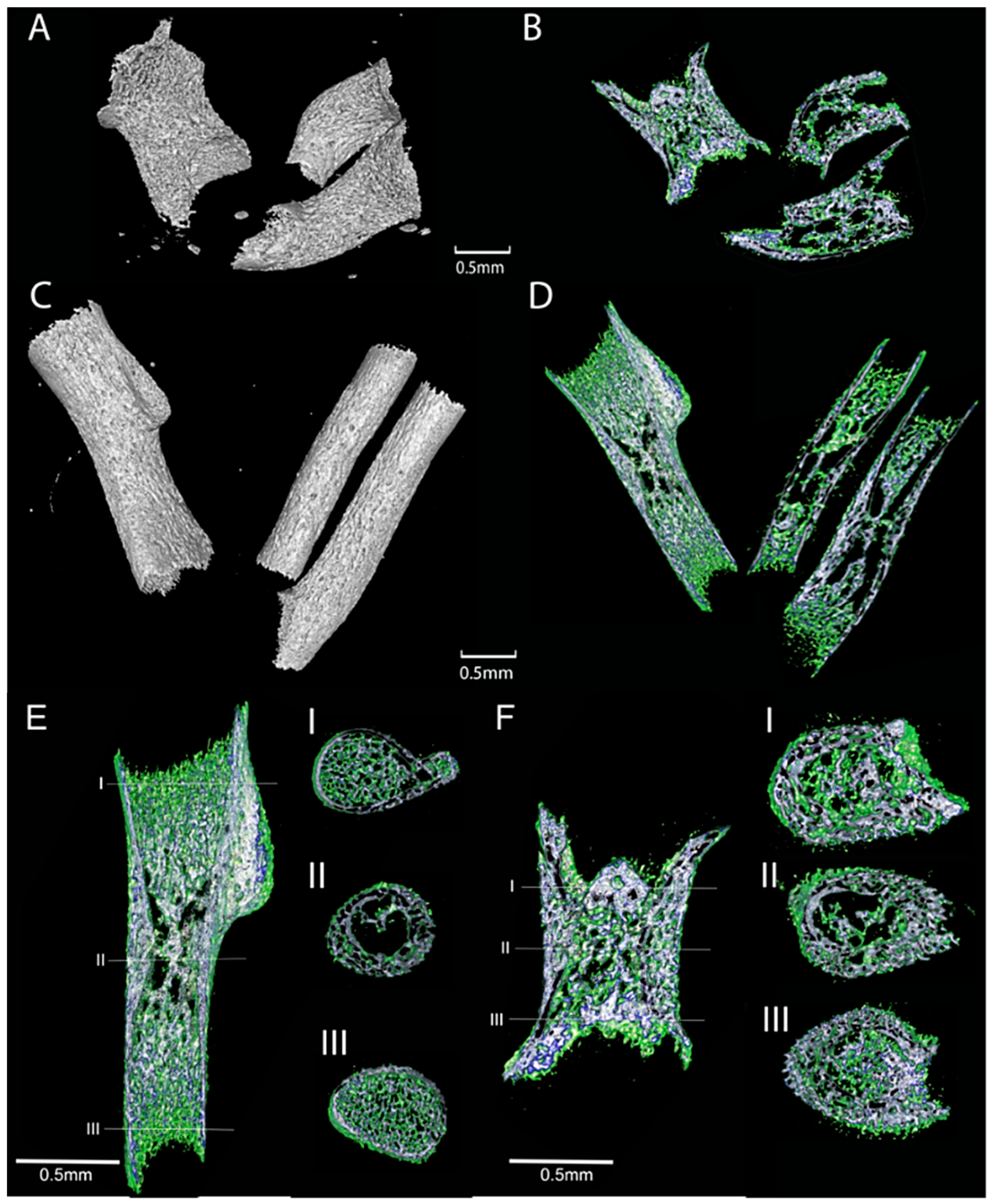
3.4. Col11a1-dependent Changes in the Ribs
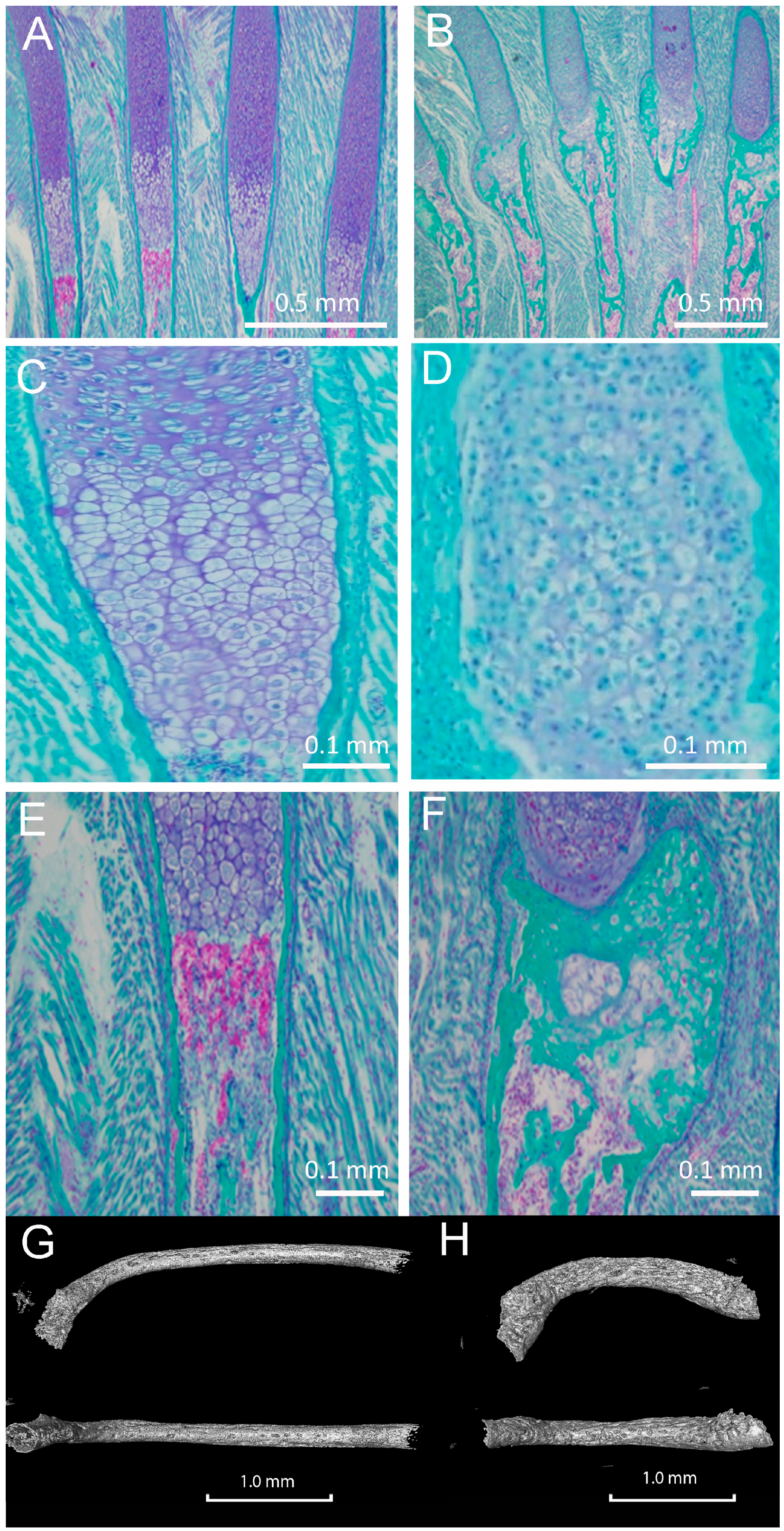
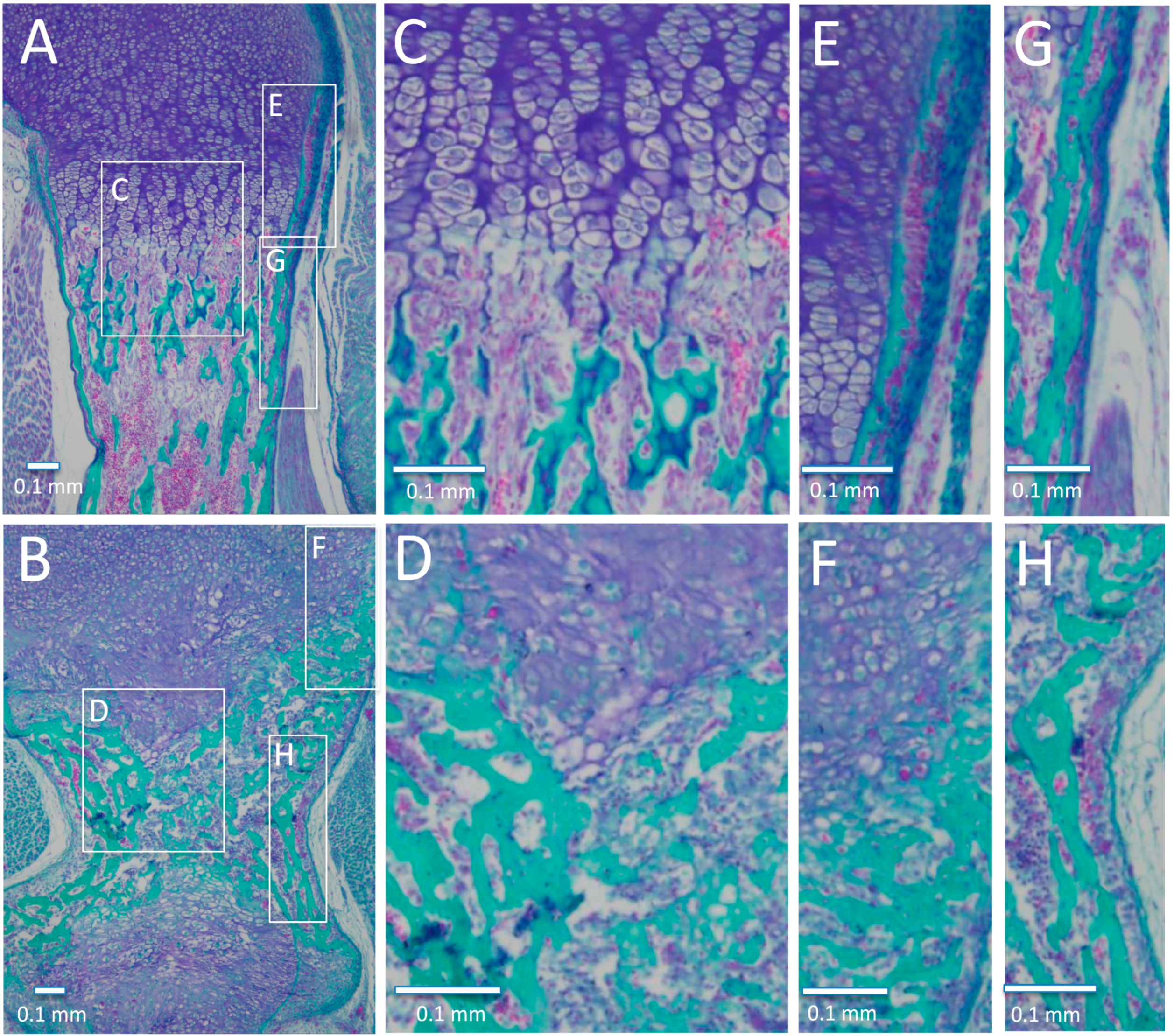
3.5. Histological Analysis of Embryonic Long Bone Formation
3.6. Metaphyses, Diaphysis, and Cross-Sectional Area of the Col11a1-Deficient Forelimbs
| Genotype | Length (µm) | Proximal Metaphysis Diameter (µm) | Diaphysis Diameter (µm) | Distal Metaphysis Diameter (µm) |
|---|---|---|---|---|
| WT | 2409 ± 33.6 | 806 ± 16.9 | 592 ± 11.6 | 528 ± 15.2 |
| Col11a1−/− | 1422 ± 65.5 | 930 ± 38.4 | 735 ± 37.4 | 778 ± 86.1 |
| % difference | 41.0 | 15.4 | 24.2 | 47.4 |
| p < 0.0001 | p < 0.05 | p < 0.05 | p < 0.05 |
| Genotype | BV/TV (%) | SMI | DA | Tb.Th (µm) | Tb.N (1/mm) | Tb.Sp (µm) |
|---|---|---|---|---|---|---|
| WT | 25.3 ± 2.9 | 2.1 ± 0.17 | 0.90 ± 0.025 | 18.32 ± 0.22 | 13.8 ± 1.5 | 33.2 ± 1.81 |
| Col11a1−/− | 43.7 ± 4.1 | 1.87 ± 0.033 | 0.84 ± 0.082 | 35.3 ± 3.7 | 12.4 ± 0.81 | 38.9 ± 1.19 |
| % difference | 72.7 | 11.0 | 0.1 | 91.6 | 10.2 | 17.2 |
| p < 0.05 | ns | ns | p < 0.05 | ns | p < 0.05 |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jacenko, O.; Roberts, D.W.; Campbell, M.R.; McManus, P.M.; Gress, C.J.; Tao, Z. Linking hematopoiesis to endochondral skeletogenesis through analysis of mice transgenic for collagen X. Am. J. Pathol. 2002, 160, 2019–2034. [Google Scholar] [CrossRef]
- Horton, W.A. The biology of bone growth. Growth Genet. Horm. 1990, 6, 21–23. [Google Scholar]
- Kronenberg, H.M. Developmental regulation of the growth plate. Nature 2003, 423, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Ornitz, D.M.; Marie, P.J. FGF signaling pathways in endochondral and intramembranous bone development and human genetic disease. Genes Dev. 2002, 16, 1446–1465. [Google Scholar] [CrossRef] [PubMed]
- Olsen, B.R.; Reginate, A.M.; Wang, W. Bone development. Annual Rev. Cell. Dev. Biol. 2000, 16, 191–220. [Google Scholar] [CrossRef] [PubMed]
- Bruder, S.P.; Caplan, A.I. Cellular and molecular events during embryonic bone development. Connect. Tissue Res. 1989, 20, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Hatori, M.; Klatte, K.J.; Teixeira, C.C.; Shapiro, I.M. End labeling studies of fragmented DNA in the avian growth plate: Evidence of apoptosis in terminally differentiated chondrocytes. J. Bone Miner. Res. 1995, 10, 1960–1968. [Google Scholar] [CrossRef] [PubMed]
- Kahn, A.J.; Simmons, D.J. Investigation of cell lineage in bone using a chimera of chick and quail embryonic tissue. Nature 1975, 258, 258–325. [Google Scholar] [CrossRef]
- Manolagas, S.C.; Jilka, R.L. Bone marrow, cytokines, and bone remodelling. Emerging insights into the pathophysiology of osteoporosis. New Engl. J. Med. 1995, 332, 305–311. [Google Scholar] [PubMed]
- Ash, P.; Loutit, J.F.; Townsend, K.M.S. Osteoclasts derived from haemoatopoietic stem cells. Nature 1980, 283, 669–670. [Google Scholar] [CrossRef] [PubMed]
- Blair, H.C.; Kahn, A.J.; Crouch, E.C.; Jeffrey, J.J.; Teitelbaum, S.L. Isolated osteoclasts resorb the organic and inorganic components of bone. J. Cell Biol. 1986, 102, 1164–1172. [Google Scholar] [CrossRef] [PubMed]
- Van der Rest, M.; Garrone, R. Collagen family of proteins. FASEB J. 1991, 5, 2814–2823. [Google Scholar] [PubMed]
- Holmes, D.F.; Kadler, K.E. The 10+4 microfibril structure of thin cartilage fibrils. Proc. Natl. Acad. Sci. USA 2006, 103, 17429–17254. [Google Scholar] [CrossRef] [PubMed]
- Blaschke, U.K.; Eikenberry, E.F.; Hulmes, D.J.; Galla, H.J.; Bruckner, P. Collagen XI nucleates self-assembly and limits lateral growth of cartilage fibrils. J. Biol. Chem. 2000, 275, 10370–10378. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.E.; Oxford, J.T.; Chen, Y.; Gambee, J.E.; Gygi, S.P.; Aebersold, R.; Neame, P.J.; Mechling, D.E.; Bächinger, H.P.; Morris, N.P. Structural organization of distinct domains within the non-collagenous N-terminal region of collagen type XI. J. Biol. Chem. 2000, 275, 11498–11506. [Google Scholar] [CrossRef] [PubMed]
- Morris, N.P.; Bachinger, H.P. Type XI collagen is a heterotrimers with the composition (1 alpha, 2 alpha, 3 alpha) retaining non-triple-helical domains. J. Biol. Chem. 1987, 262, 11345–11350. [Google Scholar] [PubMed]
- Fisher, H.; Stenling, R.; Rubio, C.; Lindblom, A. Colorectal carcinogenesis is associated with stromal expression of COL11A1 and COL5A2. Carcinogenesis 2001, 22, 875–878. [Google Scholar] [CrossRef]
- Niyibizi, C.; Eyre, D.R. Identification of the cartilage α1(XI) chain in type V collagen form bovine bone. FEBS Lett. 1989, 242, 314–318. [Google Scholar] [CrossRef]
- Mayne, R.; Brewton, R.G.; Mayne, P.M.; Baker, J.R. Isolation and characterization of the chains of type V/type XI collagen present in bovine vitreous. J. Biol. Chem. 1993, 268, 9381–9386. [Google Scholar] [PubMed]
- Sandberg, M.M.; Hirvonen, H.E.; Elima, K.J.M.; Vuorio, E.I. Co-expression of collagens II and XI and alternative splicing of exon 2 of collagen II in several developing human tissues. Biochem. J. 1993, 294, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.E.; Lawrence, R.; Sonenshein, G.E. Concerted modulation of α1(XI) and α2(V) collagen mRNAs in bovine vascular smooth muscle cells. J. Biol. Chem. 1991, 266, 23268–23273. [Google Scholar] [PubMed]
- Yamazaki, M.; Majeska, R.J.; Yoshioka, H.; Moriya, H.; Einhorn, T.A. Spatial and temporal expression of fibril-forming minor collagen genes (types V and XI) during fracture healing. J. Orthop. Res. 1997, 15, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, H.; Ramirez, F. Pro-α1 (XI) collagen Structure of the amino-terminal propeptide and expression of the gene in tumor cell lines. J. Biol. Chem. 1990, 265, 6423–6426. [Google Scholar] [PubMed]
- Kleman, J.P.; Hartmann, D.J.; Ramirez, F.; van der Rest, M. The human rhabdomyosarcoma cell line A204 lays down a highly insoluble matrix composed mainly of α1 type-XI and α2 type-V collagen chains. Eur. J. Biochem. 1992, 210, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Medeck, R.J.; Sosa, S.; Morris, N.P.; Oxford, J.T. BMP-1-mediated proteolytic processing of alternatively spliced isoforms of collagen type XI. Biochem. J. 2003, 376, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Davies, G.B.M.; Oxford, J.R.T.; Hausafus, L.C.; Smoody, B.F.; Morris, N.P. Temporal and spatial expression of spliceforms of the a1(XI) collagen gene in fetal rat cartilage. Dev. Dynam. 1998, 213, 13–26. [Google Scholar] [CrossRef]
- Zhidkova, N.I.; Justice, S.K.; Mayne, R. Alternative mRNA processing occurs in the variable region of the pro-alpha 1(XI) and pro-alpha 2(XI) collagen chains. J. Biol. Chem. 1995, 270, 9486–9493. [Google Scholar] [PubMed]
- Oxford, J.T.; Doege, K.J.; Morris, N.P. Alternative exon splicing within the amino terminal-nonhelical domain of the rat pro α1(XI) collagen chain generates multiple forms of the mRNA transcript which exhibit tissue dependent variation. J. Biol. Chem. 1995, 270, 9478–9485. [Google Scholar] [PubMed]
- Ryan, M.C.; Sandell, L.J. Differential expression of a cysteine-rich domain in the amino-terminal propeptide of Type II (cartilage) procollagen by alternative splicing of mRNA. J. Biol. Chem. 1990, 265, 10334–10339. [Google Scholar] [PubMed]
- Morris, N.P.; Oxford, J.T.; Davies, G.B.; Smoody, B.F.; Keene, D.R. Developmentally regulated alternative splicing of the α1(XI) collagen chain: spatial and temporal segregation of isoforms in the cartilage of fetal rat long bones. J. Histochem. Cytochem. 2000, 48, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lacerda, D.A.; Warman, M.L.; Beier, D.R.; Yoshioka, H.; Ninomiya, Y.; Oxford, J.T.; Morris, N.P.; Andrikopoulos, K.; Ramirez, F.; et al. A fibrillar collagen gene Col11a1, is essential for skeletal morphogenesis. Cell 1995, 80, 423–430. [Google Scholar] [CrossRef]
- Seegmiller, R.E.; Fraser, F.C.; Sheldon, H. A new chondrodystrophic mutant in mice. J. Cell. Biol. 1971, 48, 580–593. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.J.; Weis, M.; Scott, M.A.; Seegmiller, R.E.; Eyre, D.R. Collagen XI chain misassembly in cartilage of the chondrodysplasia (cho) mouse. Matrix Biol. 2007, 26, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Seegmiller, R.E.; Cooper, C.A.; Houghton, M.J.; Carey, J.C. Pulmonary hypoplasia in chondrodystrophic mice. Teratology 1986, 33, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Hepworth, W.B.; Seegmiller, R.E.; Carey, J.C. Thoracic volume reduction as a mechanism for pulmonary hypoplasia in chondrodystrophic mice. Pediatr. Pathol. 1990, 10, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Monson, C.B.; Seegmiller, R.E. Ultrastructural studies of cartilage matrix in mice homozygous for chondrodysplasia. J. Bone Joint Surg. 1981, 63, 637–644. [Google Scholar] [PubMed]
- Seegmiller, R.E.; Myers, R.A.; Dorfman, A.; Horwitz, A.L. Structure and associative properties of cartilage matrix constituents in mice with hereditary chondrodysplasia. Connect. Tissue Res. 1981, 9, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Szymko-Bennett, Y.M.; Kurima, K.; Olsen, B.; Seegmiller, R.; Griffith, A.J. Auditory function associated with Col11a1 haploinsufficiency in chondrodysplasia (cho) mice. Hear. Res. 2003, 175, 178–182. [Google Scholar] [CrossRef]
- Stephens, T.D.; Seegmiller, R.E. Normal production of cartilage glycosaminoglycan in mice homozygous for the chondrodysplasia gene. Teratology 1976, 13, 317–326. [Google Scholar] [CrossRef]
- Parfitt, A.M.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R. Bone Histomorphometry: Standardization of nomenclature, symbols and units. J. Bone Miner. Res. 1987, 2, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, D.; van Rietbergen, B.; Laib, A.; Rüegsegger, P. The ability of three-dimensional structural indices to reflect mechanical aspects of trabecular bone. Bone 1999, 25, 55–60. [Google Scholar] [CrossRef]
- Hildebrand, T.; Ruegsegger, P. A new method for the model independent assessment of thickness in three dimensional images. J. Microsc. 1997, 185, 67–75. [Google Scholar] [CrossRef]
- Lorensen, W.E.; Cline, H.E. Marching cubes: A high resolution 3D surface construction algorithm. Comput. Graph. 1987, 21, 163–169. [Google Scholar] [CrossRef]
- Carson, F.L.; Hladak, C. Histotechnology: A Self-Instructional Text, 3rd ed.; American Society for Clinical Pathology Press: Chicago, IL, USA, printed in Hong Kong, China; 2009. [Google Scholar]
- Gomori, G. A rapid one-step trichrome stain. Am. J. Clin. Pathol. 1950, 20, 661–664. [Google Scholar] [PubMed]
- Goldstein, I.; Makhoul, I.R.; Weissman, A.; Drugan, A. Hemivertebra: Prenatal diagnosis, incidence and characteristics. Fetal Diag. Ther. 2005, 20, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Kahler, R.A.; Yingst, S.M.; Hoeppner, L.H.; Jensen, E.D.; Krawczak, D.; Oxford, J.T.; Westendorf, J.J. Collagen 11a1 is indirectly activated by lymphocyte enhancer-binding factor 1(Lef1) and negatively regulated osteoblast maturation. Matrix Biol. 2008, 27, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Spranger, J. The type XI collagenopathies. Pediatr. Radiol. 1998, 28, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Yamada, Y. Chondrodysplasia of gene knockout mice for aggrecan and link protein. Glycoconj. J. 2003, 19, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Klüppel, M.; Wight, T.N.; Chan, C.; Hinek, A.; Wrana, J.L. Maintenance of chondroitin sulfonation balance by chondroitin-4-sulfotransferase-1 is required for chondrocyte development and growth factor signaling during cartilage morphogenesis. Development 2005, 132, 3989–4003. [Google Scholar] [CrossRef] [PubMed]
- Razzaque, M.S.; Soegiarto, D.W.; Chang, D.; Long, F.; Lanske, B. Conditional deletion of Indian hedgehog from collagen type 2alpha1-expressing cells results in abnormal endochondral bone formation. J. Pathol. 2005, 207, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Delezoide, A.L.; Benoist-Lasselin, C.; Legeai-Mallet, L.; le Merrer, M.; Munnich, A.; Vekemans, M.; Bonaventure, J. Spatio-temoral expression of FGFR 1, 2, 3 genes during human embryo-fetal ossification. Mech. Dev. 1998, 77, 19–30. [Google Scholar] [CrossRef]
- Okamoto, M.; Murai, J.; Yoshikawa, H.; Tsumaki, N. Bone morphogenetic proteins in bone stimulate osteoclasts and osteoblasts during bone development. J. Bone Mineral. Res. 2006, 21, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Seegmiller, R.E.; Monson, C.B. Scanning electron microscopy of cartilage in mice with hereditary chondrodysplasia. Scanning Electron Microsc. 1982, 111, 1259–1267. [Google Scholar]
- Tompson, S.W.; Bacino, C.A.; Safina, N.P.; Bober, M.B.; Proud, V.K.; Funari, T.; Wangler, M.F.; Nevarez, L.; Ala-Kokko, L.; Wilcox, W.R.; et al. Fibrochondrogenesis results from mutations in the COL11A1 type XI collagen gene. Am. J. Human Genet. 2010, 87, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Akawi, N.A.L.; Al-Gazali, L.; Ali, B.R. Clinical and molecular analysis of UAE fibrochondrogenesis patients expands the phenotype and reveals two Col11a1 homozygous null mutations. Clin. Genet. 2012, 82, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Mio, F.; Chiba, K.; Hirose, Y.; Kawaguchi, Y.; Mikami, Y.; Oya, T.; Mori, M.; Kamata, M.; Matsumoto, M.; Ozaki, K.; et al. A functional polymorphism in Col11a1, which encodes the alpha 1 chain of type XI collagen, is associated with susceptibility to lumbar disc degeneration. Am. J. Hum. Genet. 2007, 81, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hafez, A.; Squires, R.; Pedracini, A.; Joshi, A.; Seegmiller, R.E.; Oxford, J.T. Col11a1 Regulates Bone Microarchitecture during Embryonic Development. J. Dev. Biol. 2015, 3, 158-176. https://doi.org/10.3390/jdb3040158
Hafez A, Squires R, Pedracini A, Joshi A, Seegmiller RE, Oxford JT. Col11a1 Regulates Bone Microarchitecture during Embryonic Development. Journal of Developmental Biology. 2015; 3(4):158-176. https://doi.org/10.3390/jdb3040158
Chicago/Turabian StyleHafez, Anthony, Ryan Squires, Amber Pedracini, Alark Joshi, Robert E. Seegmiller, and Julia Thom Oxford. 2015. "Col11a1 Regulates Bone Microarchitecture during Embryonic Development" Journal of Developmental Biology 3, no. 4: 158-176. https://doi.org/10.3390/jdb3040158
APA StyleHafez, A., Squires, R., Pedracini, A., Joshi, A., Seegmiller, R. E., & Oxford, J. T. (2015). Col11a1 Regulates Bone Microarchitecture during Embryonic Development. Journal of Developmental Biology, 3(4), 158-176. https://doi.org/10.3390/jdb3040158