Abstract
The stomach has been a highly conserved organ throughout vertebrate evolution; however, there are now over 20 lineages composed of monotremes, lungfish and teleost fish displaying a secondary loss of stomach function and morphology. This “agastric phenotype” has evolved convergently and is typified by a loss of gastric glands and gastric acid secretion and a near-to-complete loss of storage capacity of the stomach. All agastric species have lost the genes for gastric enzymes (Pga and Pgc) and proton pump subunits (Atp4a and Atp4b), and gastrin (Gast) has been lost in monotremes. As a key gastric hormone, the conservation of gastrin has not yet been investigated in the lungfish or agastric teleosts, and it is unclear how the loss of gastrin affects the evolution and selection of the native receptor (Cckbr), gastrin-releasing peptide (Grp) and gastrin-releasing peptide receptor (Grpr) in vertebrates. Furthermore, there are still many genes implicated in gastric development and function which have yet to be associated with the agastric phenotype. We analysed the evolution, selection and conservation of the gastrin pathway and a novel gastric gene repertoire (Gkn1, Gkn2, Tff1, Tff2, Vsig1 and Anxa10) to determine the correlation with the agastric phenotype. We found that the loss of gastrin or its associated genes does not correlate with the agastric phenotype, and their conservation is due to multiple pleiotropic roles throughout vertebrate evolution. We found a loss of the gastric gene repertoire in the agastric phenotype, except in the echidna, which retained several genes (Gkn1, Tff2 and Vsig1). Our findings suggest that the gastrin physiological pathway evolved differently in pleiotropic roles throughout vertebrate evolution and support the convergent evolution of the agastric phenotype through shared independent gene-loss events.
1. Introduction
The stomach is a highly conserved and integral organ throughout jawed vertebrate (Gnathostomata) evolution, as evidenced by conserved developmental pathways found in extant representatives of basal Gnathastomes [1] and even dating back to protochordate species [2,3]. Though morphological features can vary widely between taxa, comparable components include storage compartments (enlargement of the lumen) and acid-secreting glandular epithelium [4,5]. Despite gastric features being nearly ubiquitous, a unique gastric phenotype has evolved over 15 times independently in ray-finned fish (Actinopterygii) and at least twice in lobe-finned fish (Sarcopterygii): once in lungfish and once in monotremes. This gastric phenotype—hereby known as agastric—is typified by an absence of acidic luminal contents, loss of glandular epithelium and a significant reduction in overall size and gross morphology [6,7]. These convergent morphologies only derive slightly in monotremes, where the echidna has retained the pyloric sphincter and some storage capacity [8].
In line with convergent phenotypes, agastric genotypes also exhibit similarities: agastric species share the concomitant loss of genes for gastric enzymes (Pga and Pgc) and proton pump subunits (Atp4a and Atp4b). Previously, it was found that monotremes also have lost gastrin (Gast): a key gastric hormone regulated by the upstream neuropeptide gastrin-releasing peptide (Grp) and its native receptor (Grpr) [7]. When released from gastric G cells, gastrin binds to the native cholecystokinin B receptor (Cckbr) on parietal cells, thereby potentiating gastric acid secretion [9,10,11,12]. Given the heterogeneity of the gastric epithelium in cell type (e.g., parietal cells, mucous cells and stem cells) and function (e.g., acidic digestion, immunity and hormone release) and various physiological and intracellular signalling pathways regulating gastric development and function, we expect many more genes to be correlated with the drastic shift in gastric anatomy and physiology [13,14,15].
Recent discoveries in vertebrate developmental genetics and the availability of high-quality whole genomes have expanded the repertoire of genes implicated in gastric development and function in various species. These have included the constituents of the gastrokine and trefoil factor families (Gkn1, Gkn2, Gkn3, Tff1 and Tff2), and secreted products of gastric mucosal epithelia with both independent and synergistic anti-inflammatory tumour-suppressing and anti-apoptotic roles [16,17]. In addition, Vsig1—a junction adhesion molecule expressed highly in gastric epithelia—and annexin A10 (Anxa10)—a component of the phospholipid-binding protein annexin superfamily—have displayed restricted gastric epithelial expression and have been implicated in squamous versus glandular epithelial differentiation [18,19,20]. Despite some extensive work on expression patterns (particularly in gastrokine and trefoil factor families) and recent work by Kato et al., (2024) [21]—which noted the absence of Vsig1 in agastric teleost lineages and the platypus—important questions remain. In particular, it is still unclear what emergent properties (i.e., gene ontologies) arise from the list of lost genes and whether these additional genes are conserved in agastric vertebrate lineages [22,23].
To better understand the evolution of the agastric phenotype in the Actinopterygii and Sarcopterygii clades, we explored the evolution, selection and loss of the gastrin physiological pathway along with the conservation of novel gastric gene sets in the genomes of gastric and agastric taxa.
2. Materials and Methods
2.1. Sequence Retrieval and Selection of Gastric Genes
A total of 21 chromosome-level genome assemblies from evolutionary representative taxa of tetrapods (placental mammals, marsupials, monotremes and reptiles), Sarcopterygii (lobe-finned fish) and Actinopterygii (ray-finned fish) were downloaded from NCBI Genbank (https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 5 April 2023) with a scaffold N50 filter of above 10 Mb. Aside from gastrin physiological pathway constituents, gastric genes were chosen for analyses by implication in gastric expression, digestion, gastric mucosal maintenance or gastric development from the previous literature on vertebrates in combination with manual searching through the gene ontology resource and KEGG pathway database [7,9,14,24,25]. From this list, genes were selected for gene-loss analyses if NCBI Genbank contained no annotated gene or protein in one or more agastric species.
Annotated coding DNA sequences (CDSs), exon sequences and protein sequences for gastrin pathway constituents (GAST, CCKBR, GRPR and GRP) and other gastric genes (GKN1, GKN2, TFF1, TFF2, VSIG1 and ANXA10) for these species were downloaded from NCBI Genbank (https://www.ncbi.nlm.nih.gov/genbank/). Where annotated, protein sequences were absent, CDSs from genomic sequences were uploaded to Geneious 11.0.14+1 for manual translation and alignment. See Supplementary Table S1 for sequence and genome accession numbers for all species used.
2.2. Gene-Loss Analysis
To establish gene orthology and loss, we took a modified approach of existing bioinformatic pipelines [26,27]. We firstly established genomic neighbourhoods (synteny) for the target gene through the identification of consistent flanking orthologs between species. Where synteny was conserved (i.e., flanking genes were present with no gaps), the region between orthologs was used as a query for a tblastx search with the CDS from a closely related species [28]. Where one or no flanking genes were conserved, the entire genome was used as a query for the tblastx search. To validate these sequences, the top results from BLAST 2.16.0 were then downloaded and used as a query against raw reads in the sequence read archive (SRA) BLAST function [29]. The top result was then imported into Geneious and aligned with the query CDS using Clustal Omega 1.2.2 on default settings. Inactivating mutations (e.g., premature stop codon, frameshift and deletion) were then manually annotated from these alignments. Where no sequence was returned from the BLAST search but synteny was conserved, the sequence was assumed to be entirely eroded.
2.3. Sequence Alignments and Phylogenies
Multiple sequence alignments were performed using the Clustal Omega plugin in Geneious 11.0.14+1, except for deleted/pseudogenised monotreme genes [30]. Protein maximum likelihood phylogenies were then constructed using the IQ-TREE web server (http://iqtree.cibiv.univie.ac.at/ (accessed on 12 September 2023)) tree inference tool with standard settings and 1000 bootstrap replicates [31]. Substitution models derived from the Bayesian information criterion and alignment outgroups can be found in Table S1.
2.4. Selection Analysis (Ka/Ks Ratios)
To detect signatures of selection, amino acid alignments and corresponding open reading frame DNA alignments were converted into PAML format codon alignments using PAL2NAL v 14 [32]. Codon alignments along with their corresponding phylogenies in Newick format were then input into EasyCodeML v1.41 using the preset branch model with multiples models of foreground branch selection for each lineage and likelihood ratio tests (LRTs) to assess model significance [33]. EasyCodeML outputs can be found in Table S2.
3. Results
3.1. Conservation and Loss of the Gastrin Release Pathway and Novel Gastric Gene Repertoire in Gastric and Agastric Jawed Vertebrates (Gnathostomata)
Synteny analysis of gastrin (Gast) and the native gastrin receptor (Cckbr), as well as the gastrin-releasing peptide (Grp) and its receptor (Grpr), returned conserved syntenic blocks in most ray- and lobe-finned fish species with recurring differences of flanking loci between lineages (Figure 1A,B). Our genome search returned no orthologous sequences for gastrin in the genomes of the agastric pufferfish or West African lungfish, indicating near-total erosion in these genomes. In the agastric zebrafish, an orthologous sequence in the conserved syntenic region which could be validated against raw sequence reads was identified. The highly conserved syntenic blocks of tetrapod species corroborated the previously noted gene losses of gastrin in both monotreme species (Ordoñez et al., 2008 [9]), with our analyses indicating the loss of Grpr in the platypus through deletion of the latter two exons (Figure 1C).
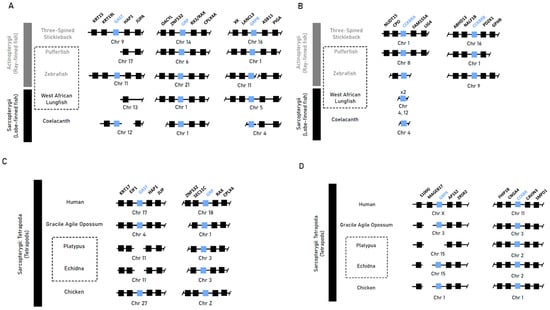
Figure 1.
Syntenic analysis of gastrin (Gast), gastrin-releasing peptide (Grp), gastrin-releasing peptide receptor (Grpr) and cholecystokinin receptor B (Cckbr) in select vertebrate species. (A,B) Synteny plot of gastric genes lost or pseudogenised in the agastric species of the zebrafish (D. rerio) and West African lungfish (P. annectens) compared to the gastric three-spined stickleback (G. aculeatus) and coelacanth (L. chalumnae). (C,D) Synteny plot of the short-beaked echidna (T. aculeatus) and platypus (O. anatinus) when compared to humans (H. sapiens/placental mammals), gracile agile opossums (G. agilis/marsupials) and chickens (G. gallus/reptiles). Class-level taxonomy (Sarcopterygii vs Actinopterygii) is displayed on the left, boxes with dotted lines indicate agastric species, coloured boxes indicate genes of interest, numbers within a box indicate presence of only that family member, and breaks or missing boxes in the syntenic block indicate gene absence.
These findings suggest that the agastric phenotype does not correlate with the loss of gastrin. Furthermore, the absence of gastrin is not consistently associated with the loss of its indirect upstream regulator, Grpr.
We then explored the conservation and loss of genes implicated in gastric epithelial maintenance and development in jawed vertebrates. A BLAST search and synteny analysis of the sequences bearing gastrokine family homology (Gkn1, Gkn2 and Gkn3), trefoil factor homology (Tff1 and Tff2), as well as Anxa10 and Vsig1 returned no orthologous sequences in agastric Actinopterygii taxa. Conversely, in the gastric grey bichir and spotted gar, there was a conservation of the syntenic neighbourhood with an orthologous sequence conserved (Figure 2A).
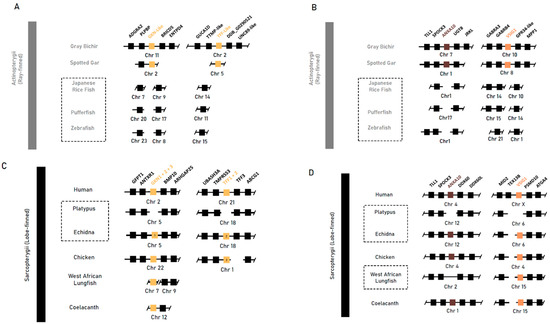
Figure 2.
Syntenic analysis of genes implicated in gastric mucosal maintenance (Gkn1 and Gkn2) and gastric development (Vsig1) and unclear roles in gastric mucosal maintenance, growth and differentiation (Anxa10) in select vertebrate species. (A,B) Syntenic analyses of Actinopterygii taxa: the agastric teleost species of the Japanese rice fish (O. latipes), pufferfish (T. rubripes) and zebrafish (D. rerio) compared to the gastric grey bichir (P. senegalus) and spotted gar (L. oculatus). (C,D) Syntenic analyses of Sarcopterygii taxa: the agastric short-beaked echidna (T. aculeatus), platypus (O. anatinus) and West African lungfish (P. annectens) compared to the gastric human (H. sapien), chicken (G. gallus) and coelacanth (L. chalumnae). Class-level taxonomy (Sarcopterygii vs Actinopterygii) is displayed on the left, boxes with dotted lines indicate agastric species, coloured boxes indicate genes of interest, numbers within a box indicate presence of only that family member, and breaks or missing boxes in the syntenic block indicate gene absence.
In Sarcopterygii, gastrokine family paralogs (Gkn1, Gkn2 and Gkn3) were mainly conserved together within a syntenic block flanked by Antxr1 and Bmp10. Gastrokine paralogs 2 and 3 were lost in both monotreme species, but Gkn1 was retained in the echidna. In the platypus, Gkn1 was found to be inactivated through the accumulation of missense and nonsense mutations throughout the CDS. The three Gastrokine paralogs found in the West African lungfish were not placed within the conserved genomic region, and further phylogenetic analysis of these sequences shows that basal Sarcopterygii sequences (coelacanth, West African lungfish) were placed in a clade separate from all three vertebrate paralogs. Furthermore, the echidna Gkn1 sequence has higher similarity to Gkn3 sequences and was placed within the vertebrate Gkn3 clade with 100 percent bootstrap support (Figures S1 and S2).
Trefoil factor paralogs 1 and 2 (Tff1 and Tff2) were absent in the coelacanth and West African lungfish and found only in tetrapod species in a highly conserved syntenic block between Tff3 and Spock3, except in the platypus—which was missing both—and the echidna, which was missing Tff1. Genomic sequences of Anxa10 and Vsig1 were found in all Sarcopterygii taxa but not in the platypus, supporting the findings by Kato et al., [21], which associate the loss with the agastric phenotype (Figure 2B). These findings suggest that these genes were likely present in early gnathastome evolution, prior to the divergence of major jawed vertebrate lineages, and were later lost independently.
3.2. Sequence Evolution and Selection Pressures in the Gastrin Release Pathway Between Gastric and Agastric Jawed Vertebrates (Gnathostomata)
We then investigated sequence evolution and Ka/Ks ratios in the gastrin release pathway in gastric and agastric vertebrates to see whether gastrin loss affected the evolution or selection of other pathway components.
The Actinopterygii clade of gastrin sequences displayed minimal differences in substitution per site between the gastric three-spined stickleback (0.48) and agastric Japanese rice fish (0.34) (Figure 3A). Branches leading to monotreme and therian mammal CCKBR clades displayed identical lengths (0.17 substitutions per site) but differed more in post-divergence in the mouse and wombat (0.41 and 0.25 substitutions per site) when compared to the agastric platypus and echidna (0.07 and 2 × 10−6 substitutions per site). Actinopterygii CCKBRA sequences had similar substitutions per site within the clade of the pufferfish (0.15), the Japanese rice fish (0.15) and the three-spined stickleback (0.12). The pufferfish CCKBRB sequence contained a larger number of substitutions per site (0.2) when compared to three-spined stickleback (0.07) and Japanese rice fish (0.13) (Figure 3B).
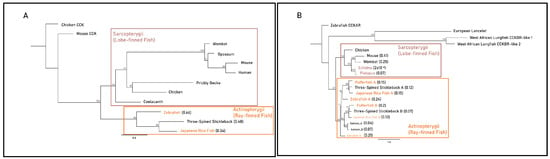
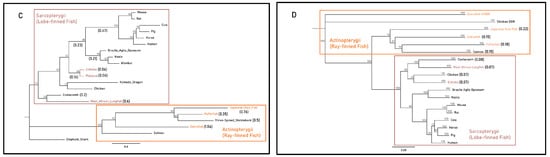
Figure 3.
Maximum-likelihood amino acid phylogenies of the gastrin release pathway constituents (A) gastrin (GAST), (B) cholecystokinin B receptor (CCKBR), (C) gastrin-releasing peptide (GRP) and (D) gastrin-releasing peptide receptor (GRPR) for select gastric and agastric vertebrate species constructed by the IQ-TREE web server with 1000 bootstrap replicates. Taxa, substitution models as chosen by the Bayesian information criterion (BIC), outgroups and sequence references are in Supplementary File. Paralogues of the CCKBR receptor for teleost lineages are identified with the letters A and B. Branch labels denote the bootstrap percentage (%), coloured tip labels represent agastric lineages, coloured boxes outline class-level taxonomic groupings and branch length, and scale bar and bracketed numbers measure substitutions per site to the nearest bifurcation.
In the Sarcopterygii clade, Grp sequences from the coelacanth and West African lungfish displayed large variations in substitutions per site (0.2 vs. 0.6). Though echidna and platypus sequences displayed identical estimated substitutions per site (0.06), there was a substantial difference in branch length relative to the root of mammalian sequences between monotremes (0.16), marsupials (0.44) and placental mammals (0.7). In the Actinopterygii clade, branch lengths differed relative to the root of the clade with the Japanese rice fish (0.76), pufferfish (0.41) and three-spined stickleback (0.55) (Figure 3C). Sarcopterygii and Actinopterygii Grpr sequences exhibited little difference in substitutions per site between gastric and agastric species, as shown by comparisons between the salmon (0.15) and the pufferfish (0.18) and comparisons of the coelacanth (0.08) and West African lungfish (0.07) (Figure 3D). Together, these findings suggest that the presence or absence of a stomach does not strictly correlate with accelerated molecular evolution or pseudogenization of these genes across vertebrate lineages. Instead, some agastric species retain high sequence conservation, implying possible pleiotropic roles or evolutionary constraints unrelated to gastric function.
Finally, we calculated the ratio of synonymous to non-synonymous substitutions (Ka/Ks) using maximum-likelihood branch model selection analyses. Overall, we found that the Gast, Cckbr, Grp and Grpr genes displayed several bouts of purifying selection (Table S1). Foreground scenarios with significant likelihood-ratio tests (LRTs) in the Cckbr phylogeny included branches leading to the clade with Japanese rice fish, pufferfish and three-spined stickleback Cckbrb (p = 0.04, ω1 = 0.19 and ω0 = 0.08), the three-spined stickleback Cckbra (p = 0.01, ω1 = 3 × 10−3 and ω0 = 0.07) and Cckbrb sequences (p = 1.12 × 10−5, ω1 = 0.33 and ω0 = 0.08). In the Grp phylogeny, significant LRTs included branches leading to the tetrapod clade (p = 2.17 × 10−5, ω1 = 8.9 × 10−4 and ω0 = 0.28), the mammalian clade (p = 0.03, ω1 = 0.02 and ω0 = 0.24) and the mouse sequence (p = 0.04, ω1 = 0.02, and ω0 = 0.24). The Grpr phylogeny included two significant LRTs: the branch leading to the pufferfish sequence (p = 0.02, ω1 = 0.11 and ω0 = 0.04) and to the Sarcopterygii clade (p = 8.93 × 10−7, ω1 = 1.94 × 10−3 and ω0 = 0.05) (Table 1). These findings suggest that independent bursts of sequence evolution in the gastrin release pathway were primarily characterised by conservative mutations resulting in the maintenance of functional integrity. Furthermore, these molecular changes appeared to have no direct correlation with variation in gastric phenotypes.

Table 1.
Summary of significant findings (p < 0.05) from CodeML branch model selection analysis of Cckbr, Grp and Grpr sequences with columns for the foreground branch (taxonomic grouping or species), the model-fitting metric (likelihood-ratio test p) and the Ka/Ks ratio under differing foreground scenarios (foreground = ω1 and background = ω0).
4. Discussion
Stomach anatomy in vertebrates can be broadly described as gastric or agastric. The latter phenotype lacks gross morphology, glandular epithelium and gastric acid secretion, as has been described in independent teleost lineages and monotremes. Changes in stomach morphology have been correlated with the loss of gastric enzymes (Pga and Pgc), proton pump subunits (Atp4a and Atp4b) and the gastrin hormone (Gast) in monotremes [7,9,11,21]. Due to the complex constitution and function of the stomach—with a large complement of cell types, signalling pathways and physiological functions—we hypothesised that the gene losses associated with the agastric phenotype extended beyond the aforementioned genes. To test this, we investigated the evolution, selection and loss of the gastrin physiological pathway along with genes implicated in gastric development and functionality in both gastric and agastric Sarcopterygii and Actinopterygii taxa.
4.1. Gastrin Physiological Pathway: Correlations with Gastric Phenotype, Known Functions and Genetic Dispensability
We determined loss of gastrin in four of the five agastric species presented (platypus, echidna, pufferfish and West African lungfish) but no substantial differences in substitution rate or selection pressures of the gastrin physiological pathway constituents (Gast, Cckbr, Grp and Grpr) between gastric and agastric species or between species who had retained or lost gastrin. These findings demonstrate that gastrin loss is not likely associated with the agastric phenotype and that loss of gastrin does not correlate with subsequent sequence evolution, gene loss or selection in other gastrin release pathway constituents.
These findings are surprising given both the conserved role in gastric acid secretion and localised expression of gastric mucosa in vertebrate species [34,35]. However, gastrin and the closely related cholecystokinin have been shown to be expressed in the stomach and the central nervous system (CNS) in vertebrates [36], and our analyses here provide further evidence of pleiotropic roles of these genes.
Recent research has found novel roles of gastrin and its related isoforms (progastrin and the glycine-extended gastrins) in cell proliferation, apoptosis and extracellular remodelling in colonic epithelium in a suite of vertebrate models [37]. Furthermore, mouse gastrin knockout models present with heavily deleterious gastric phenotypes: achlorydic gastric environments with high incidences of gastric metaplasia, bacterial overgrowth and tumours [38]. These unique roles and deleterious knockout phenotypes raise questions as to how the monotreme ancestor and potentially other fish lineages could have lost gastrin without a subsequent substantial reduction in fitness.
4.2. Evolution of Gastric Genes Implicated in the Secondary Loss of Gastric Phenotype in Jawed Vertebrates (Gnathostomata)
We noted the absence of gastrokine (Gkn1 and Gkn2) and trefoil factor homologs (Tff1 and Tff2), Vsig1 and Anxa10 in all agastric taxa with the exception of gastrokine-like sequences and Vsig1 in the West African lungfish. Convergent loss between agastric Actinopterygii taxa, the West African lungfish and the platypus suggests a role of these genes in the secondary loss of stomach features, in line with the loss of gastric enzymes, proton pump subunits and gastrin [7,9,11,32]. Furthermore, the different inactivating mutations displayed between independent agastric taxa highlight the independent nature of these loss events.
Our findings are not consistent for the echidna, which has an agastric phenotype and has retained Gkn1, Tff2, Vsig1 and Anxa10. These findings align with our recent works in which the common monotreme ancestor likely lost antral glandular epithelium and pyloric restriction through Nkx3.2 pseudogenisation and subsequent shifts in developmental processes. However, these findings also point to a retention or re-establishment of pyloric-like restriction in the echidna lineage through potential compensation or an additional evolutionary event [39].
The presence of trefoil factor and gastrokine homologs in Actinopterygii taxa suggests the evolution of these genes likely occurred early in Gnathostome evolution, prior to the divergence of major jawed vertebrate lineages. Furthermore, presence of a homologous trefoil factor-like sequence in the vase tunicate (Ciona intestinalis) provides evidence of gene evolution, in line with the advent of the alimentary canal in early ascidians (Table S1), [3]. Recent research has implicated these genes in the differentiation of squamous and glandular epithelium in the mammalian hindstomach as well as mucosal proliferation/differentiation and protection to chemical insults [40,41,42]. The gastrokines and trefoil factor homologs, in particular, are known for their potent anti-tumorigenic properties and are implicated in several gastrointestinal cancers [43,44,45]). Further in vivo experimentation and comparative genomics in a variety of biological models could determine whether these features represent ancestral traits or lineage-specific innovations.
Moreover, this study highlights that Anxa10 and Vsig1 arose prior to the divergence of jawed vertebrates. Previous characterisations of Anxa10 have outlined its likely role as a calcium-dependent phospholipid-binding protein and its expression in both healthy and cancerous (i.e., carcinoma) gastric tissues in humans, but this study is the first to implicate the gene in function in a wide range of vertebrate species [19,46,47,48]. Similarly, Vsig1 has been implicated to regulate the development of glandular gastric epithelium in mice models, but our findings, along with those from Kato et al. (2024), show an ancestral role in gastric development in jawed vertebrates [20].
5. Conclusions
Here, we investigated the evolution, selection and conservation of the gastrin physiological pathway and other genes involved in gastric function in gastric and agastric jawed vertebrates in order to further understand the independent evolutionary trajectories of the agastric phenotype. Interestingly, we discovered that the absence of a gastric gene repertoire (Gkn1, Gkn2, Tff1, Tff2, Vsig1 and Anxa10)—which evolved prior to the divergence of major jawed vertebrate lineages—correlates with the agastric phenotype, with the exception of the echidna and West African lungfish, which may have retained some aspects of gastric functionality (Figure 4). We found that the loss of Gast and Grpr does not correlate with the gastric or agastric phenotype. These findings differentiate aspects of the agastric phenotype and reveal convergent gene loss and retention in agastric species separated by more than 500 million years of evolution.
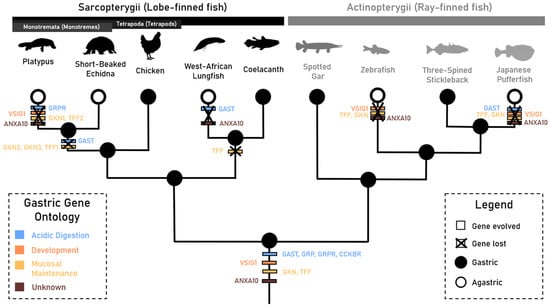
Figure 4.
Phylogeny outlining the proposed gastric gene evolution, duplication and loss events between taxa displaying the gastric and agastric phenotype in Sarcopterygii (lobe-finned fish) and Actinopterygii (ray-finned fish) clades. Agastric taxa are denoted by hollow branch tips, gastric gene ontologies are outlined by gene colouration, and gene evolution and loss events are signified by white and black borders, respectively. Branches are not drawn to scale.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jdb13030027/s1, Figure S1: Clustal Omega amino acid alignment of vertebrate gastrokine sequences (GKN1, GKN2, GKN3, GKN-like) with sequence consensus metrics. Dark-shaded boxes indicate 100% sequence identity, whereas lighter-shaded gray boxes indicate lower sequence identity metrics; Figure S2: Maximum-likelihood phylogeny of vertebrate gastrokinelike amino acid sequences constructed by the IQ-Tree web server using the JTT + G4 substitution model and 1000 bootstrap replicates. Branch values indicate bootstrap percentage, the scale represents substitutes per site and groupings of gastrokine-like sequences are superimposed on the right; Table S1: Sequence and genome NCBI accession numbers as well as substitution models and phylogenetics settings; Table S2: EasyCodeML outputs.
Author Contributions
Conceptualization, J.D.; methodology, J.D. and F.G.; software, J.D.; validation, J.D. and F.G.; formal analysis, J.D.; investigation, J.D.; resources, F.G.; data curation, J.D.; writing—original draft preparation, J.D.; writing—review and editing, F.G.; visualization, J.D.; supervision, F.G.; project administration, J.D. and F.G.; funding acquisition, F.G. All authors have read and agreed to the published version of the manuscript.
Funding
J.D. was supported by a University of Adelaide Research Scholarship.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequence data are openly available through NCBI GenBank (https://www.ncbi.nlm.nih.gov/genbank/), otherwise all data necessary to evaluate results and conclusions are available within the paper, references and supplementary materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gonçalves, O.; Freitas, R.; Ferreira, P.; Araujo, M.; Zhang, G.; Mazan, S.; Cohn, M.J.; Castro, L.F.C.; Wilson, J.M. Molecular ontogeny of the stomach in the castshark Scyliorhinus canicular. Sci. Rep. 2019, 9, 586. [Google Scholar] [CrossRef]
- Nakazawa, K.; Yamazawa, T.; Moriyama, Y.; Ogura, Y.; Kawai, N.; Sasakura, Y.; Saiga, H. Formation of the digestive tract in Ciona intestinalis includes two distinct morphogenic processes between its anterior and posterior parts. Dev. Dyn. 2013, 242, 1172–1183. [Google Scholar] [CrossRef]
- Nakayama, S.; Ogasawara, M. Compartmentalized expression patterns of pancreatic- and gastric-related genes in the alimentary canal of the ascidian Ciona intestinalis: Evolutionary insights into the functional regionality of the gastrointestinal tract in Olfactores. Cell Tissue Res. 2017, 370, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Koelz, H.R. Gastric Acid in Vertebrates. Scand. J. Gastroenterol. 1992, 27, 2–6. [Google Scholar] [CrossRef]
- Karasov, W.H.; Douglas, A.E.; Ronald, T. Comparative Digestive Physiology. Compr. Physiol. 2013, 3, 741–783. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Castro, L.F.C. The Multifunctional Gut of Fish; Academic Press: London, UK, 2010. [Google Scholar]
- Castro, L.F.C.; Gonçalves, O.; Mazan, S.; Tay, B.H.; Venkatesh, B.; Wilson, J.M. Recurrent gene loss correlates with the evolution of stomach phenotypes in gnathostome history. Proc. R. Soc. B 2014, 281, 20132669. [Google Scholar] [CrossRef]
- Griffiths, M. The Biology of Monotremes; Academic Press: New York, NY, USA, 1978; pp. 77–81. [Google Scholar]
- Ordoñez, G.R.; Hillier, L.D.W.; Warren, W.C.; Grützner, F.; López-Otín, C.; Puente, X.S. Loss of genes implicated in gastric function during platypus evolution. Genome Biol. 2008, 9, R81. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.L.; Peura, D.A. Control of Gastric Acid Secretion in Health and Disease. Gastroenterology 2008, 134, 1842–1860. [Google Scholar] [CrossRef]
- Zhou, Y.; Shearwin-Whyatt, L.; Li, J.; Song, Z.; Hayakawa, T.; Stevens, D.; Fenelon, J.C.; Peel, E.; Cheng, Y.; Pajpach, F.; et al. Platypus and echidna genomes reveal mammalian biology and evolution. Nature 2021, 592, 756–762. [Google Scholar] [CrossRef]
- Duan, S.; Rico, K.; Merchant, J.L. Gastrin: From Physiology to Gastrointestinal Malignancies. Function 2022, 3, zqab062. [Google Scholar] [CrossRef]
- Kim, T.H.; Shivdasani, R.A. Stomach development, stem cells and disease. Development 2016, 143, 554–565. [Google Scholar] [CrossRef]
- McCracken, K.W.; Wells, J.M. Mechanisms of embryonic stomach development. Semin. Cell Dev. Biol. 2017, 66, 36–42. [Google Scholar] [CrossRef]
- Agace, W.W.; McCoy, K.D. Regionalized Development and Maintenance of the Intestinal Adaptive Immune Landscape. Immunity 2017, 46, 532–548. [Google Scholar] [CrossRef] [PubMed]
- Menheniott, T.R.; Kurklu, B.; Giraud, A.S. Gastrokines: Stomach-specific proteins with putative homeostatic and tumor suppressor roles. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G109–G121. [Google Scholar] [CrossRef]
- Hoffmann, W. Trefoil factor family (TFF) peptides and their diverse molecular functions in mucus barrier protection and more: Changing the paradigm. Int. J. Mol. Sci. 2020, 21, 4535. [Google Scholar] [CrossRef] [PubMed]
- Gerke, V.; Creutz, C.E.; Moss, S.E. Annexins: Linking Ca2+ signalling to membrane dynamics. Nat. Rev. Mol. Cell Biol. 2005, 6, 449–461. [Google Scholar] [CrossRef]
- Lu, S.H.; Chen, Y.L.; Shun, C.T.; Lai, J.N.; Peng, S.Y.; Lai, P.L.; Hsu, H.C. Expression and prognostic significance of gastric-specific annexin A10 in diffuse- and intestinal-type gastric carcinoma. J. Gastroenterol. Hepatol. 2011, 26, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Oidovsambuu, O.; Nyamsuren, G.; Liu, S.; Goring, W.; Engel, W.; Adham, I.M.; Gires, O. Adhesion protein VSIG1 is required for the proper differentiation of glandular gastric epithelia. PLoS ONE 2011, 6, e25908. [Google Scholar] [CrossRef]
- Kato, A.; Pipil, S.; Ota, C.; Kusakabe, M.; Watanabe, T.; Nagashima, A.; Chen, A.P.; Islam, Z.; Hayashi, N.; Wong, M.K.S.; et al. Convergent gene losses and pseudogenizations in multiple lineages of stomachless fishes. Commun. Biol. 2024, 7, 408. [Google Scholar] [CrossRef]
- Jiang, Z.; Lossie, A.C.; Applegate, T.J. Evolution of Trefoil Factor(s): Genetic and Spatio-Temporal Expression of Trefoil Factor 2 in the Chicken (Gallus Gallus Domesticus). PLoS ONE 2011, 6, e22691. [Google Scholar] [CrossRef]
- Geahlen, J.H.; Lapid, C.; Thorell, K.; Nikolskiy, I.; Huh, W.J.; Oates, E.L.; Lennerz, J.K.M.; Tian, X.; Weis, V.G.; Khurana, S.S.; et al. Evolution of the human gastrokine locus and confounding factors regarding the pseudogenicity of GKN3. Physiol. Genom. 2013, 45, 667–683. [Google Scholar] [CrossRef]
- Carbon, S.; Ireland, A.; Mungall, C.J.; Shu, S.; Marshall, B.; Lewis, S.; AmiGO Hub; Web Presence Working Group. AmiGO: Online access to ontology and annotation data. Bioinformatics 2009, 25, 288–289. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 1, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Hecker, N.; Roscito, J.G.; Foerster, L.; Langer, B.; Hiller, M. A genomics approach reveals insights into the importance of gene losses for mammalian adaptations. Nat. Commun. 2018, 9, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Marín, B.; Rakijas, J.B.; Tyagi, A.; Pandey, A.; Hanschen, E.R.; Anderson, J.; Heffel, M.G.; Platt, T.G.; Olson, B.J.S.C. Gene loss during a transition to multicellularity. Sci. Rep. 2023, 13, 5268. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 12, 421. [Google Scholar] [CrossRef]
- Katz, K.; Shutov, O.; Lapoint, R.; Kimelman, M.; Brister, J.R.; O’Sullivan, C. The Sequence Read Archive: A decade of more explosive growth. Nucleic Acids Res. 2022, 50, D387–D390. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haasler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Mikita, S.; Torrents, D.; Bork, P. PAL2NAL: Robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res. 2006, 34, W609–W612. [Google Scholar] [CrossRef]
- Gao, F.; Chen, C.; Arab, D.A.; Du, Z.; He, Y.; Ho, S.Y.W. EasyCodeML: A visual tool for analysis of selection using CodeML. Ecol. Evol. 2019, 9, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, A.H. Phylogeny of the Cholecystokinin/Gastrin Family. Front. Neuroendocr. 1998, 19, 73–99. [Google Scholar] [CrossRef]
- Cui, T.; Wang, J.; Hu, Z.; Chen, X. Expression of gastrin and cholecystokinin B receptor in Lateolabrax maculatus. Aquac. Fish. 2023, 8, 492–497. [Google Scholar] [CrossRef]
- Nässel, D.R.; Wu, S.F. Cholecystokinin/sulfakinin peptide signaling: Conserved roles at the intersection between feeding, mating and aggression. Cell. Mol. Life Sci. 2022, 79, 188. [Google Scholar] [CrossRef]
- Dimaline, R.; Varro, A. Novel roles of gastrin. J. Physiol. 2014, 592, 2951–2958. [Google Scholar] [CrossRef]
- Friis-Hansen, L. Lessons from the gastrin knockout mice. Regul. Pept. 2007, 139, 5–22. [Google Scholar] [CrossRef]
- Dann, J.; Qu, Z.; Shearwin-Whyatt, L.; van der Ploeg, R.; Grützner, F. Pseudogenization of NK3 homeobox 2 (Nkx3.2) in monotremes provides insight into unique gastric anatomy and physiology. Open Biol. 2024, 14, 240071. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.R.; Xu, S.H.; Tan, Z.L.; Yin, X.F.; He, Q.Y. Proteomics characterization of gastrokine 1-induced growth inhibition of gastric cancer cells. Proteomics 2011, 11, 3657–3664. [Google Scholar] [CrossRef]
- Kim, O.; Yoon, J.H.; Choi, W.S.; Ashktorab, H.; Smoot, D.T.; Nam, S.W.; Lee, J.Y.; Park, W.S. GKN2 Contributes to the Homeostasis of Gastric Mucosa by Inhibiting GKN1 Activity. J. Cell. Physiol. 2013, 229, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, W. TFF2, a MUC6-binding lectin stabilizing the gastric mucus barrier and more (Review). Int. J. Oncol. 2015, 47, 806–816. [Google Scholar] [CrossRef]
- Jahan, R.; Shah, A.; Kisling, S.G.; Macha, M.A.; Thayer, S.; Batra, S.K.; Sukhwinder, K. Odyssey of trefoil factors in cancer: Diagnostic and therapeutic implications. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188362. [Google Scholar] [CrossRef]
- Soutto, M.; Belkhiri, A.; Piazuelo, M.B.; Blanca, M.; Schneider, B.G.; Peng, D.; Jiang, A.; Washington, M.K.; Kokoye, Y.; Crowe, S.E.; et al. Loss of TFF1 is associated with activation of NF-κB-mediated inflammation and gastric neoplasia in mice and humans. J. Clin. Investig. 2011, 121, 1753–1767. [Google Scholar] [CrossRef]
- Moss, S.E.; Morgan, R.O. The annexins. Genome Biol. 2004, 5, 219–735. [Google Scholar] [CrossRef]
- Kim, J.K.; Kim, P.J.; Jung, K.H.; Noh, J.H.; Eun, J.W.; Bae, H.J.; Xie, H.J.; Shan, J.M.; Ping, W.Y.; Park, W.S.; et al. Decreased expression of Annexin A10 in gastric cancer and its overexpression in tumor cell growth expression. Oncol. Rep. 2010, 24, 607–612. [Google Scholar] [PubMed]
- Galura, G.M.; Chavez, L.O.; Robles, A.; McCallum, R. Gastroduodenal Injury: Role of Protective Factors. Curr. Gastroenterol. Rep. 2019, 21, 34. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.H.; Lin, Y.L.; Cheng, Y.C.; Chen, C.C.; Lin, L.I.; Tseng, L.H.; Cheng, M.L.; Liau, J.Y.; Jeng, Y.M. Aberrant expression of annexin A10 is closely related to gastric phenotype in serrated pathway to colorectal carcinoma. Mod. Pathol. 2015, 28, 268–278. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).