
Crucial Convolution: Genetic and Molecular Mechanisms of Coiling during Epididymis Formation and Development in Embryogenesis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. The Need for and the Mechanisms of Coiling and Folding across Development
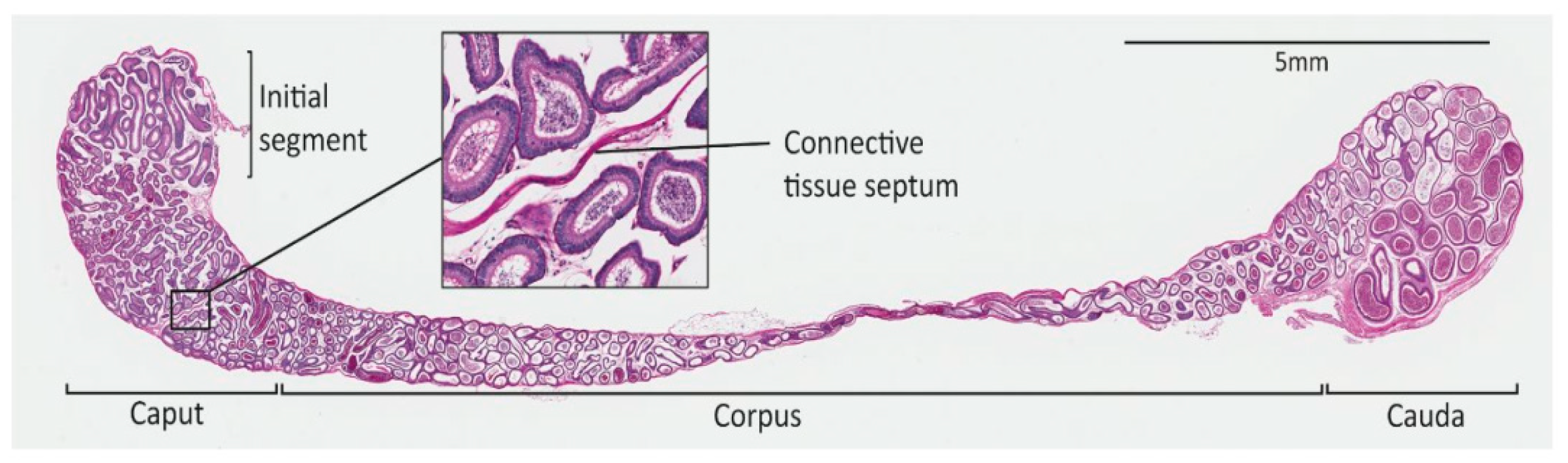
1.1.1. The Epididymis as a Model for Understanding Tissue Coiling
1.1.2. Androgen Signalling
1.1.3. WNT Signalling
1.1.4. Activin A
1.1.5. A Role for the Extracellular Matrix?
1.2. Seminiferous Tubules
Intestine
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinzel, B.; Pikiolek, M.; Orsini, V.; Sprunger, J.; Isken, A.; Zietzling, S.; Desplanches, M.; Dubost, V.; Breustedt, D.; Valdez, R.; et al. Functional roles of Lgr4 and Lgr5 in embryonic gut, kidney and skin development in mice. Dev. Biol. 2014, 390, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Hinton, B.T.; Galdamez, M.M.; Sutherland, A.; Bomgardner, D.; Xu, B.; Abdel-Fattah, R.; Yang, L. How Do You Get Six Meters of Epididymis Inside a Human Scrotum? J. Androl. 2011, 32, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Combes, A.N.; Lesieur, E.; Harley, V.R.; Sinclair, A.H.; Little, M.H.; Wilhelm, D.; Koopman, P. Three-dimensional visualization of testis cord morphogenesis, a novel tubulogenic mechanism in development. Dev. Dyn. 2009, 238, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Ritsche, I.S.; Fahlke, J.M.; Wieder, F.; Hilger, A.; Manke, I.; Hampe, O. Relationships of cochlear coiling shape and hearing frequencies in cetaceans, and the occurrence of infrasonic hearing in Miocene Mysticeti. Foss. Rec. 2018, 21, 33–45. [Google Scholar] [CrossRef]
- Dworkin, S.; Jane, S.M. Novel mechanisms that pattern and shape the midbrain-hindbrain boundary. CMLS 2013, 70, 3365–3374. [Google Scholar] [CrossRef]
- Männer, J. The anatomy of cardiac looping: A step towards the understanding of the morphogenesis of several forms of congenital cardiac malformations. Clin. Anat. 2009, 22, 21–35. [Google Scholar] [CrossRef]
- Sullivan, R.; Mieusset, R. The human epididymis: Its function in sperm maturation. Hum. Reprod. Updat. 2016, 22, 574–587. [Google Scholar] [CrossRef]
- Nerurkar, N.L.; Mahadevan, L.; Tabin, C.J. BMP signaling controls buckling forces to modulate looping morphogenesis of the gut. Proc. Natl. Acad. Sci. USA 2017, 114, 2277–2282. [Google Scholar] [CrossRef]
- Sela-Donenfeld, D.; Kayam, G.; Wilkinson, D.G. Boundary cells regulate a switch in the expression of FGF3 in hindbrain rhombomeres. BMC Dev. Biol. 2009, 9, 16. [Google Scholar] [CrossRef]
- Nakata, H. Morphology of mouse seminiferous tubules. Anat. Sci. Int. 2019, 94, 1–10. [Google Scholar] [CrossRef]
- Lewis, S.; Chen, L.; Raghuram, V.; Khundmiri, S.J.; Chou, C.-L.; Yang, C.-R.; Knepper, M.A. “SLC-omics” of the kidney: Solute transporters along the nephron. Am. J. Physiol. Cell Physiol. 2021, 321, C507–C518. [Google Scholar] [CrossRef] [PubMed]
- Andrew, D.J.; Ewald, A.J. Morphogenesis of epithelial tubes: Insights into tube formation, elongation, and elaboration. Dev. Biol. 2010, 341, 34–55. [Google Scholar] [CrossRef] [PubMed]
- Lubarsky, B.; Krasnow, M.A. Tube Morphogenesis: Making and Shaping Biological Tubes. Cell 2003, 112, 19–28. [Google Scholar] [CrossRef]
- Iruela-Arispe, M.L.; Beitel, G.J. Tubulogenesis. Development 2013, 140, 2851–2855. [Google Scholar] [CrossRef]
- Soffers, J.H.; Hikspoors, J.P.; Mekonen, H.K.; Koehler, S.E.; Lamers, W.H. The growth pattern of the human intestine and its mesentery. BMC Dev. Biol. 2015, 15, 31. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Kikuchi, K.; Hochgreb, T.; Poss, K.D.; Stainier, D.Y. Hand2 Regulates Extracellular Matrix Remodeling Essential for Gut-Looping Morphogenesis in Zebrafish. Dev. Cell 2010, 18, 973–984. [Google Scholar] [CrossRef]
- Onouchi, S.; Ichii, O.; Nakamura, T.; Elewa, Y.H.A.; Kon, Y. Spatiotemporal distribution of extracellular matrix changes during mouse duodenojejunal flexure formation. Cell Tissue Res. 2016, 365, 367–379. [Google Scholar] [CrossRef]
- Savin, T.; Kurpios, N.A.; Shyer, A.E.; Florescu, P.; Liang, H.; Mahadevan, L.; Tabin, C.J. On the growth and form of the gut. Nature 2011, 476, 57–61. [Google Scholar] [CrossRef]
- Campione, M.; Steinbeisser, H.; Schweickert, A.; Deissler, K.; van Bebber, F.; Lowe, L.; Nowotschin, S.; Viebahn, C.; Haffter, P.; Kuehn, M.; et al. The homeobox gene Pitx2: Mediator of asymmetric left-right signaling in vertebrate heart and gut looping. Development 1999, 126, 1225–1234. [Google Scholar] [CrossRef]
- Tsuda, T.; Majumder, K.; Linask, K.K. Differential expression of flectin in the extracellular matrix and left-right asymmetry in mouse embryonic heart during looping stages. Genesis 1998, 23, 203–214. [Google Scholar] [CrossRef]
- Eshi, Y.; Eyao, J.; Young, J.M.; Fee, J.A.; Eperucchio, R.; Taber, L.A. Bending and twisting the embryonic heart: A computational model for c-looping based on realistic geometry. Front. Physiol. 2014, 5, 297. [Google Scholar]
- Le Garrec, J.-F.; Domínguez, J.N.; Desgrange, A.; Ivanovitch, K.D.; Raphaël, E.; Bangham, J.A.; Torres, M.; Coen, E.; Mohun, T.J.; Meilhac, S.M. A predictive model of asymmetric morphogenesis from 3D reconstructions of mouse heart looping dynamics. eLife 2017, 6, e28951. [Google Scholar] [CrossRef] [PubMed]
- Ocaña, O.H.; Coskun, H.; Minguillón, C.; Murawala, P.; Tanaka, E.M.; Galceran, J.; Muñoz-Chápuli, R.; Nieto, M.A. A right-handed signalling pathway drives heart looping in vertebrates. Nature 2017, 549, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Linask, K.K.; Han, M.; Cai, D.H.; Brauer, P.R.; Maisastry, S.M. Cardiac morphogenesis: Matrix metalloproteinase coordination of cellular mechanisms underlying heart tube formation and directionality of looping. Dev. Dyn. 2005, 233, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Linask, K.K.; VanAuker, M. A Role for the Cytoskeleton in Heart Looping. Sci. World J. 2007, 7, 280–298. [Google Scholar] [CrossRef]
- Hinton, B.T.; Murashima, A.; Xu, B. Understanding normal and abnormal development of the Wolffian/epididymal duct by using transgenic mice. Asian J. Androl. 2015, 17, 749–755. [Google Scholar] [CrossRef]
- França, L.R.; Avelar, G.F.; Almeida, F.F. Spermatogenesis and sperm transit through the epididymis in mammals with emphasis on pigs. Theriogenology 2005, 63, 300–318. [Google Scholar] [CrossRef]
- Dacheux, J.-L.; Dacheux, F. New insights into epididymal function in relation to sperm maturation. Reproduction 2014, 147, R27–R42. [Google Scholar] [CrossRef] [PubMed]
- Guyonnet, B.; Dacheux, F.C.; Dacheux, J.; Gatti, J.-L. The Epididymal Transcriptome and Proteome Provide Some Insights Into New Epididymal Regulations. J. Androl. 2011, 32, 651–664. [Google Scholar] [CrossRef]
- Joseph, A.; Yao, H.; Hinton, B.T. Development and morphogenesis of the Wolffian/epididymal duct, more twists and turns. Dev. Biol. 2009, 325, 6–14. [Google Scholar] [CrossRef]
- McArdle, C.A.; Roberson, M.S.; Plant, T.M.; Zeleznik, A.J. Orgebin-Crist, The epididymis. In Knobil and Neill’s Physiology of Reproduction, 4th ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 1071–1148. [Google Scholar]
- Ferreira, L.G.; Nishino, F.A.; Fernandes, S.G.; Ribeiro, C.M.; Hinton, B.T.; Avellar, M.C.W. Epididymal embryonic development harbors TLR4/NFKB signaling pathway as a morphogenetic player. J. Reprod. Immunol. 2022, 149, 103456. [Google Scholar] [CrossRef] [PubMed]
- Jelinsky, S.A.; Turner, T.T.; Bang, H.J.; Finger, J.N.; Solarz, M.K.; Wilson, E.; Brown, E.L.; Kopf, G.S.; Johnston, D.S. The Rat Epididymal Transcriptome: Comparison of Segmental Gene Expression in the Rat and Mouse Epididymides1. Biol. Reprod. 2007, 76, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Damdimopoulos, A.; Damdimopoulou, P.; Gasperoni, J.G.; Tran, S.C.; Grommen, S.V.; De Groef, B.; Dworkin, S. Transcriptome analysis of the epididymis from Plag1 deficient mice suggests dysregulation of sperm maturation and extracellular matrix genes. Dev. Dyn. 2020, 249, 1500–1513. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Juma, A.R.; Tran, S.C.; Gasperoni, J.G.; Grommen, S.V.H.; De Groef, B. Deficiency of the transcription factor PLAG1 results in aberrant coiling and morphology of the epididymis. Asian J. Androl. 2020, 22, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Murashima, A.; Kishigami, S.; Thomson, A.; Yamada, G. Androgens and mammalian male reproductive tract development. Biochim. Biophys. Acta 2015, 1849, 163–170. [Google Scholar] [CrossRef]
- Hannema, S.E.; Hughes, I.A. Regulation of Wolffian Duct Development. Horm. Res. Paediatr. 2007, 67, 142–151. [Google Scholar] [CrossRef]
- Shaw, G.; Renfree, M.B. Wolffian Duct Development. Sex. Dev. 2014, 8, 273–280. [Google Scholar] [CrossRef]
- Menad, R.; Fernini, M.; Lakabi, L.; Smaï, S.; Gernigon-Spychalowicz, T.; Farida, K.; Bonnet, X.; Moudilou, E.; Exbrayat, J.-M. Androgen and estrogen receptors immunolocalization in the sand rat (Psammomys obesus) cauda epididymis. Acta Histochem. 2021, 123, 151683. [Google Scholar] [CrossRef]
- Welsh, M.; Saunders, P.; Marchetti, N.I.; Sharpe, R.M. Androgen-Dependent Mechanisms of Wolffian Duct Development and Their Perturbation by Flutamide. Endocrinology 2006, 147, 4820–4830. [Google Scholar] [CrossRef][Green Version]
- Welsh, M.; Sharpe, R.M.; Walker, M.; Smith, L.B.; Saunders, P.T.K. New Insights into the Role of Androgens in Wolffian Duct Stabilization in Male and Female Rodents. Endocrinology 2009, 150, 2472–2480. [Google Scholar] [CrossRef]
- Ribeiro, C.M.; Ferreira, L.G.; Thimoteo, D.S.; Smith, L.B.; Hinton, B.T.; Avellar, M.C.W. Novel androgen-induced activity of an antimicrobial β-defensin: Regulation of Wolffian duct morphogenesis. Mol. Cell. Endocrinol. 2017, 442, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Björkgren, I.; Alvarez, L.; Blank, N.; Balbach, M.; Turunen, H.; Laajala, T.D.; Toivanen, J.; Krutskikh, A.; Wahlberg, N.; Huhtaniemi, I.; et al. Targeted inactivation of the mouse epididymal beta-defensin 41 alters sperm flagellar beat pattern and zona pellucida binding. Mol. Cell. Endocrinol. 2016, 427, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Diao, R.; Fok, K.L.; Chen, H.; Yu, M.K.; Duan, Y.; Chung, C.M.; Li, Z.; Wu, H.; Li, Z.; Zhang, H.; et al. Deficient human β-defensin 1 underlies male infertility associated with poor sperm motility and genital tract infection. Sci. Transl. Med. 2014, 6, 108–249. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.S.; Webb, S.; Lettice, L.; Tardif, S.; Kilanowski, F.; Tyrrell, C.; MacPherson, H.; Semple, F.; Tennant, P.; Baker, T.; et al. Partial Deletion of Chromosome 8 β-defensin Cluster Confers Sperm Dysfunction and Infertility in Male Mice. PLoS Genet. 2013, 9, e1003826. [Google Scholar] [CrossRef]
- Zhou, C.X.; Zhang, Y.-L.; Xiao, L.; Zheng, M.; Leung, K.M.; Chan, M.Y.; Lo, P.S.; Tsang, L.L.; Wong, H.Y.; Ho, L.S.; et al. An epididymis-specific β-defensin is important for the initiation of sperm maturation. Nat. Cell Biol. 2004, 6, 458–464. [Google Scholar] [CrossRef]
- Osterhoff, C.; Kirchhoff, C.; Krull, N.; Ivell, R. Molecular Cloning and Characterization of a Novel Human Sperm Antigen (HE2) Specifically Expressed in the Proximal Epididymis. Biol. Reprod. 1994, 50, 516–525. [Google Scholar] [CrossRef]
- Ribeiro, C.M.; Queiróz, D.B.; Patrão, M.T.; Denadai-Souza, A.; Romano, R.M.; Silva, E.J.; Avellar, M.C.W. Dynamic changes in the spatio-temporal expression of the β-defensin SPAG11C in the developing rat epididymis and its regulation by androgens. Mol. Cell. Endocrinol. 2015, 404, 141–150. [Google Scholar] [CrossRef]
- Murashima, A.; Miyagawa, S.; Ogino, Y.; Nishida-Fukuda, H.; Araki, K.; Matsumoto, T.; Kaneko, T.; Yoshinaga, K.; Yamamura, K.-I.; Kurita, T.; et al. Essential Roles of Androgen Signaling in Wolffian Duct Stabilization and Epididymal Cell Differentiation. Endocrinology 2011, 152, 1640–1651. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, C.J.; Liu, Y. Wnt Signaling in Kidney Development and Disease. Prog. Mol. Biol. Transl. Sci. 2017, 153, 181–207. [Google Scholar]
- Carroll, T.J.; Park, J.-S.; Hayashi, S.; Majumdar, A.; McMahon, A.P. Wnt9b Plays a Central Role in the Regulation of Mesenchymal to Epithelial Transitions Underlying Organogenesis of the Mammalian Urogenital System. Dev. Cell 2005, 9, 283–292. [Google Scholar] [CrossRef]
- Koch, S.; Acebron, S.P.; Herbst, J.; Hatiboglu, G.; Niehrs, C. Post-transcriptional Wnt Signaling Governs Epididymal Sperm Maturation. Cell 2015, 163, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Syed, S.M.; Taketo, M.M.; Tanwar, P.S. Epithelial Wnt/βcatenin signalling is essential for epididymal coiling. Dev. Biol. 2016, 412, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Okazawa, M.; Murashima, A.; Harada, M.; Nakagata, N.; Noguchi, M.; Morimoto, M.; Kimura, T.; Ornitz, D.M.; Yamada, G. Region-specific regulation of cell proliferation by FGF receptor signaling during the Wolffian duct development. Dev. Biol. 2015, 400, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Warr, N.; Siggers, P.; Bogani, D.; Brixey, R.; Pastorelli, L.; Yates, L.; Dean, C.H.; Wells, S.; Satoh, W.; Shimono, A.; et al. Sfrp1 and Sfrp2 are required for normal male sexual development in mice. Dev. Biol. 2009, 326, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Yun, K.; Ajima, R.; Sharma, N.; Costantini, F.; Mackem, S.; Lewandoski, M.; Yamaguchi, T.P.; Perantoni, A.O. Non-canonical Wnt5a/Ror2 signaling regulates kidney morphogenesis by controlling intermediate mesoderm extension. Hum. Mol. Genet. 2014, 23, 6807–6814. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Washington, A.M.; Domeniconi, R.F.; Souza, A.C.F.; Lu, X.; Sutherland, A.; Hinton, B.T. Protein tyrosine kinase 7 is essential for tubular morphogenesis of the Wolffian duct. Dev. Biol. 2016, 412, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Karner, C.; Wharton, K.A.; Carroll, T.J. Planar cell polarity and vertebrate organogenesis. Semin. Cell Dev. Biol. 2006, 17, 194–203. [Google Scholar] [CrossRef]
- Lee, J.; Andreeva, A.; Sipe, C.W.; Liu, L.; Cheng, A.; Lu, X. PTK7 Regulates Myosin II Activity to Orient Planar Polarity in the Mammalian Auditory Epithelium. Curr. Biol. 2012, 22, 956–966. [Google Scholar] [CrossRef][Green Version]
- Andreeva, A.; Lee, J.; Lohia, M.; Wu, X.; Macara, I.G.; Lu, X. PTK7-Src Signaling at Epithelial Cell Contacts Mediates Spatial Organization of Actomyosin and Planar Cell Polarity. Dev. Cell 2014, 29, 20–33. [Google Scholar] [CrossRef]
- Mendis, S.H.; Meachem, S.J.; Sarraj, M.A.; Loveland, K.L. Activin A Balances Sertoli and Germ Cell Proliferation in the Fetal Mouse Testis. Biol. Reprod. 2010, 84, 379–391. [Google Scholar] [CrossRef]
- Wijayarathna, R.; de Kretser, D.M.; Meinhardt, A.; Middendorff, R.; Ludlow, H.; Groome, N.P.; Loveland, K.A.; Hedger, M.P. Activin over-expression in the testis of mice lacking the inhibin α-subunit gene is associated with androgen deficiency and regression of the male reproductive tract. Mol. Cell. Endocrinol. 2017, 470, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Miles, D.C.; Wakeling, S.I.; Stringer, J.M.; Bergen, J.A.V.D.; Wilhelm, D.; Sinclair, A.H.; Western, P.S. Signaling through the TGF Beta-Activin Receptors ALK4/5/7 Regulates Testis Formation and Male Germ Cell Development. PLoS ONE 2013, 8, e54606. [Google Scholar] [CrossRef] [PubMed]
- Meehan, T.; Schlatt, S.; O’Bryan, M.; de Kretser, D.M.; Loveland, K. Regulation of Germ Cell and Sertoli Cell Development by Activin, Follistatin, and FSH. Dev. Biol. 2000, 220, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Bilezikjian, L.M.; Blount, A.L.; Donaldson, C.J.; Vale, W.W. Pituitary actions of ligands of the TGF-β family: Activins and inhibins. Reproduction 2006, 132, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Cui, X.; Ge, J.; Li, J.; Niu, L.; Liu, H.; Qi, Y.; Liu, Z.; Wang, Y. Activin A inhibits activities of lipopolysaccharide-activated macrophages via TLR4, not of TLR2. Biochem. Biophys. Res. Commun. 2013, 435, 222–228. [Google Scholar] [CrossRef]
- Nicolas, N.; Muir, J.A.; Hayward, S.; Chen, J.L.; Stanton, P.G.; Gregorevic, P.; de Kretser, D.M.; Loveland, K.L.; Bhushan, S.; Meinhardt, A.; et al. Induction of experimental autoimmune orchitis in mice: Responses to elevated circulating levels of the activin-binding protein, follistatin. Reproduction 2017, 154, 293–305. [Google Scholar] [CrossRef]
- Hedger, M.P.; Winnall, W.R. Regulation of activin and inhibin in the adult testis and the evidence for functional roles in spermatogenesis and immunoregulation. Mol. Cell. Endocrinol. 2012, 359, 30–42. [Google Scholar] [CrossRef]
- Muttukrishna, S.; Farouk, A.; Sharma, S.K.; Evans, L.; Groome, N.; Ledger, W.; Sathanandan, M. Serum activin A and follistatin in disorders of spermatogenesis in men. Eur. J. Endocrinol. 2001, 144, 425–429. [Google Scholar] [CrossRef][Green Version]
- Ritvos, O.; Tuuri, T.; Erämaa, M.; Sainio, K.; Hilden, K.; Saxén, L.; Gilbert, S.F. Activin disrupts epithelial branching morphogenesis in developing glandular organs of the mouse. Mech. Dev. 1995, 50, 229–245. [Google Scholar] [CrossRef]
- Cancilla, B.; Jarred, R.A.; Wang, H.; Mellor, S.L.; Cunha, G.R.; Risbridger, G. Regulation of Prostate Branching Morphogenesis by Activin A and Follistatin. Dev. Biol. 2001, 237, 145–158. [Google Scholar] [CrossRef]
- Tomaszewski, J.; Joseph, A.; Archambeault, D.; Yao, H.H. Essential roles of inhibin beta A in mouse epididymal coiling. Proc. Natl. Acad. Sci. USA 2007, 104, 11322–11327. [Google Scholar] [CrossRef] [PubMed]
- Wijayarathna, R.; Sarraj, M.A.; Genovese, R.; Girling, J.; Michel, V.; Ludlow, H.; Loveland, K.L.; Meinhardt, A.; De Kretser, D.M.; Hedger, M.P. Activin and follistatin interactions in the male reproductive tract: Activin expression and morphological abnormalities in mice lacking follistatin 288. Andrology 2017, 5, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Winnall, W.R.; Wu, H.; Sarraj, M.; Rogers, P.A.W.; De Kretser, D.M.; Girling, J.; Hedger, M. Expression patterns of activin, inhibin and follistatin variants in the adult male mouse reproductive tract suggest important roles in the epididymis and vas deferens. Reprod. Fertil. Dev. 2013, 25, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Daley, W.P.; Yamada, K.M. ECM-modulated cellular dynamics as a driving force for tissue morphogenesis. Curr. Opin. Genet. Dev. 2013, 23, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, E.; Mayor, R. Collective cell migration in development. J. Cell Biol. 2016, 212, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Wight, T.N.; Potter-Perigo, S. The extracellular matrix: An active or passive player in fibrosis? Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, 950–955. [Google Scholar] [CrossRef]
- Long, K.B.; Artlett, C.M.; Blankenhorn, E.P. Tight skin 2 mice exhibit a novel time line of events leading to increased extracellular matrix deposition and dermal fibrosis. Matrix Biol. 2014, 38, 91–100. [Google Scholar] [CrossRef]
- Karsdal, M.A.; Manon-Jensen, T.; Genovese, F.; Kristensen, J.H.; Nielsen, M.J.; Sand, J.M.B.; Hansen, N.-U.B.; Bay-Jensen, A.-C.; Bager, C.L.; Krag, A.; et al. Novel insights into the function and dynamics of extracellular matrix in liver fibrosis. Am. J. Physiol. Liver Physiol. 2015, 308, G807–G830. [Google Scholar] [CrossRef]
- Dai, L.; Zhang, X.; Hu, X.; Liu, Q.; Man, Z.; Huang, H.; Meng, Q.; Zhou, C.; Ao, Y. Silencing of miR-101 Prevents Cartilage Degradation by Regulating Extracellular Matrix–related Genes in a Rat Model of Osteoarthritis. Mol. Ther. 2015, 23, 1331–1340. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, X.; Dai, L.; Hu, X.; Zhu, J.; Li, L.; Zhou, C.; Ao, Y. Long Noncoding RNA Related to Cartilage Injury Promotes Chondrocyte Extracellular Matrix Degradation in Osteoarthritis. Arthritis Rheumatol. 2014, 66, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Troeberg, L.; Nagase, H. Proteases involved in cartilage matrix degradation in osteoarthritis. Biochim. Biophys. Acta 2012, 1824, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Hoshii, T.; Takeo, T.; Nakagata, N.; Takeya, M.; Araki, K.; Yamamura, K.-I. LGR4 Regulates the Postnatal Development and Integrity of Male Reproductive Tracts in Mice1. Biol. Reprod. 2007, 76, 303–313. [Google Scholar] [CrossRef]
- Obara-Ishihara, T.; Kuhlman, J.; Niswander, L.; Herzlinger, D. The surface ectoderm is essential for nephric duct formation in intermediate mesoderm. Development 1999, 126, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Arend, L.J. Pkd1 is required for male reproductive tract development. Mech. Dev. 2013, 130, 567–576. [Google Scholar] [CrossRef]
- Chauvet, V.; Qian, F.; Boute, N.; Cai, Y.; Phakdeekitacharoen, B.; Onuchic, L.F.; Attié-Bitach, T.; Guicharnaud, L.; Devuyst, O.; Germino, G.G.; et al. Expression of PKD1 and PKD2 Transcripts and Proteins in Human Embryo and during Normal Kidney Development. Am. J. Pathol. 2002, 160, 973–983. [Google Scholar] [CrossRef]
- Nie, X.; Arend, L.J. Novel roles of Pkd2 in male reproductive system development. Differentiation 2014, 87, 161–171. [Google Scholar] [CrossRef]
- Hu, J.; Chen, Y.-X.; Wang, D.; Qi, X.; Li, T.-G.; Hao, J.; Mishina, Y.; Garbers, D.L.; Zhao, G.-Q. Developmental expression and function of Bmp4 in spermatogenesis and in maintaining epididymal integrity. Dev. Biol. 2004, 276, 158–171. [Google Scholar] [CrossRef]
- Zhao, G.; Liaw, L.; Hogan, B. Bone morphogenetic protein 8A plays a role in the maintenance of spermatogenesis and the integrity of the epididymis. Development 1998, 125, 1103–1112. [Google Scholar] [CrossRef]
- Zhao, G.-Q.; Chenb, Y.X.; Liub, X.M.; Xuab, Z.; Qiab, X. Mutation in Bmp7 Exacerbates the Phenotype of Bmp8a Mutants in Spermatogenesis and Epididymis. Dev. Biol. 2001, 240, 212–222. [Google Scholar] [CrossRef]
- Kato, S.; Matsubara, M.; Matsuo, T.; Mohri, Y.; Kazama, I.; Hatano, R.; Umezawa, A.; Nishimori, K. Leucine-Rich Repeat-Containing G Protein-Coupled Receptor-4 (LGR4, Gpr48) Is Essential for Renal Development in Mice. Nephron Exp. Nephrol. 2006, 104, e63–e75. [Google Scholar] [CrossRef] [PubMed]
- Mohri, Y.; Oyama, K.; Akamatsu, A.; Kato, S.; Nishimori, K. Lgr4-deficient mice showed premature differentiation of ureteric bud with reduced expression of Wnt effector Lef1 and Gata3. Dev. Dyn. 2011, 240, 1626–1634. [Google Scholar] [CrossRef] [PubMed]
- Mendive, F.; Laurent, P.; van Schoore, G.; Skarnes, W.; Pochet, R.; Vassart, G. Defective postnatal development of the male reproductive tract in LGR4 knockout mice. Dev. Biol. 2006, 290, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Cool, J.; Carmona, F.; Szucsik, J.; Capel, B. Peritubular Myoid Cells Are Not the Migrating Population Required for Testis Cord Formation in the XY Gonad. Sex. Dev. 2008, 2, 128–133. [Google Scholar] [CrossRef]
- Coveney, D.; Cool, J.; Oliver, T.; Capel, B. Four-dimensional analysis of vascularization during primary development of an organ, the gonad. Proc. Natl. Acad. Sci. USA 2008, 105, 7212–7217. [Google Scholar] [CrossRef]
- Svingen, T.; Koopman, P. Building the mammalian testis: Origins, differentiation, and assembly of the component cell populations. Genes Dev. 2013, 27, 2409–2426. [Google Scholar] [CrossRef]
- Chen, S.-R.; Liu, Y.-X. Testis Cord Maintenance in Mouse Embryos: Genes and Signaling1. Biol. Reprod. 2016, 94, 42. [Google Scholar] [CrossRef]
- Nel-Themaat, L.; González, G.; Akiyama, H.; Behringer, R.R. Illuminating Testis Morphogenesis in the Mouse. J. Androl. 2010, 31, 5–10. [Google Scholar] [CrossRef]
- Archambeault, D.R.; Tomaszewski, J.; Joseph, A.; Hinton, B.T.; Yao, H.H.-C. Epithelial-mesenchymal crosstalk in Wolffian duct and fetal testis cord development. Genesis 2009, 47, 40–48. [Google Scholar] [CrossRef][Green Version]
- Cool, J.; DeFalco, T.; Capel, B. Testis formation in the fetal mouse: Dynamic and complex de novo tubulogenesis. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 847–859. [Google Scholar] [CrossRef]
- Archambeault, D.R.; Yao, H.H.-C. Activin A, a product of fetal Leydig cells, is a unique paracrine regulator of Sertoli cell proliferation and fetal testis cord expansion. Proc. Natl. Acad. Sci. USA 2010, 107, 10526–10531. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R. Intestinal atresia and stenosis: A review comparing its etiopathogenesis. Veter. Res. Commun. 1986, 10, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Shaw-Smith, C. Review of genetic factors in intestinal malrotation. Pediatr. Surg. Int. 2010, 26, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Iwuagwu, O.; Deans, G.T. Small bowel volvulus: A review. J. R. Coll. Surg. Edinb. 1999, 44, 150–155. [Google Scholar]
- Chinya, A.; Naranje, K.; Mandelia, A. Situs inversus abdominalis, polysplenia, complex jejunal atresia and malrotation in a neonate: A rare association. Int. J. Surg. Case Rep. 2019, 56, 93–95. [Google Scholar] [CrossRef]
- Davis, N.M.; Kurpios, N.A.; Sun, X.; Gros, J.; Martin, J.F.; Tabin, C.J. The Chirality of Gut Rotation Derives from Left-Right Asymmetric Changes in the Architecture of the Dorsal Mesentery. Dev. Cell 2008, 15, 134–145. [Google Scholar] [CrossRef]
- Shiratori, H.; Sakuma, R.; Watanabe, M.; Hashiguchi, H.; Mochida, K.; Sakai, Y.; Nishino, J.; Saijoh, Y.; Whitman, M.; Hamada, H. Two-Step Regulation of Left–Right Asymmetric Expression of Pitx2: Initiation by Nodal Signaling and Maintenance by Nkx2. Mol. Cell 2001, 7, 137–149. [Google Scholar] [CrossRef]
- Welsh, I.C.; Thomsen, M.; Gludish, D.W.; Alfonso-Parra, C.; Bai, Y.; Martin, J.F.; Kurpios, N.A. Integration of Left-Right Pitx2 Transcription and Wnt Signaling Drives Asymmetric Gut Morphogenesis via Daam2. Dev. Cell 2013, 26, 629–644. [Google Scholar] [CrossRef]
- Shiratori, H.; Yashiro, K.; Shen, M.M.; Hamada, H. Conserved regulation and role of Pitx2 in situs-specific morphogenesis of visceral organs. Development 2006, 133, 3015–3025. [Google Scholar] [CrossRef]
- Cervantes, S.; Yamaguchi, T.P.; Hebrok, M. Wnt5a is essential for intestinal elongation in mice. Dev. Biol. 2009, 326, 285–294. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, J.; Gasperoni, J.; Fuller, J.; Grommen, S.V.H.; De Groef, B.; Hogarth, C.; Dworkin, S. Crucial Convolution: Genetic and Molecular Mechanisms of Coiling during Epididymis Formation and Development in Embryogenesis. J. Dev. Biol. 2022, 10, 25. https://doi.org/10.3390/jdb10020025
Wong J, Gasperoni J, Fuller J, Grommen SVH, De Groef B, Hogarth C, Dworkin S. Crucial Convolution: Genetic and Molecular Mechanisms of Coiling during Epididymis Formation and Development in Embryogenesis. Journal of Developmental Biology. 2022; 10(2):25. https://doi.org/10.3390/jdb10020025
Chicago/Turabian StyleWong, Joanne, Jemma Gasperoni, Jarrad Fuller, Sylvia V. H. Grommen, Bert De Groef, Cathryn Hogarth, and Sebastian Dworkin. 2022. "Crucial Convolution: Genetic and Molecular Mechanisms of Coiling during Epididymis Formation and Development in Embryogenesis" Journal of Developmental Biology 10, no. 2: 25. https://doi.org/10.3390/jdb10020025
APA StyleWong, J., Gasperoni, J., Fuller, J., Grommen, S. V. H., De Groef, B., Hogarth, C., & Dworkin, S. (2022). Crucial Convolution: Genetic and Molecular Mechanisms of Coiling during Epididymis Formation and Development in Embryogenesis. Journal of Developmental Biology, 10(2), 25. https://doi.org/10.3390/jdb10020025