
Male Infertility: The Effect of Natural Antioxidants and Phytocompounds on Seminal Oxidative Stress

Abstract
:1. Introduction
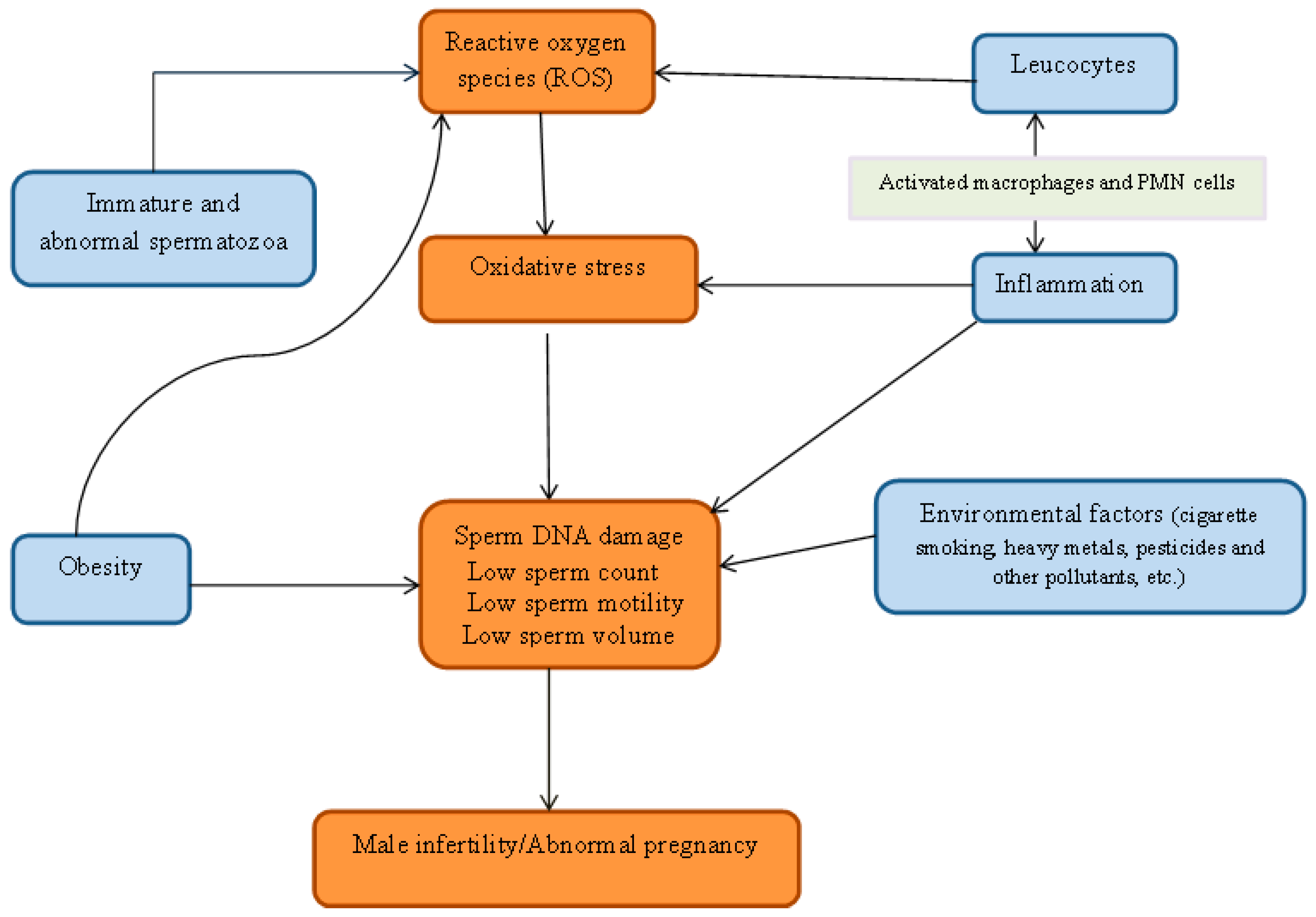
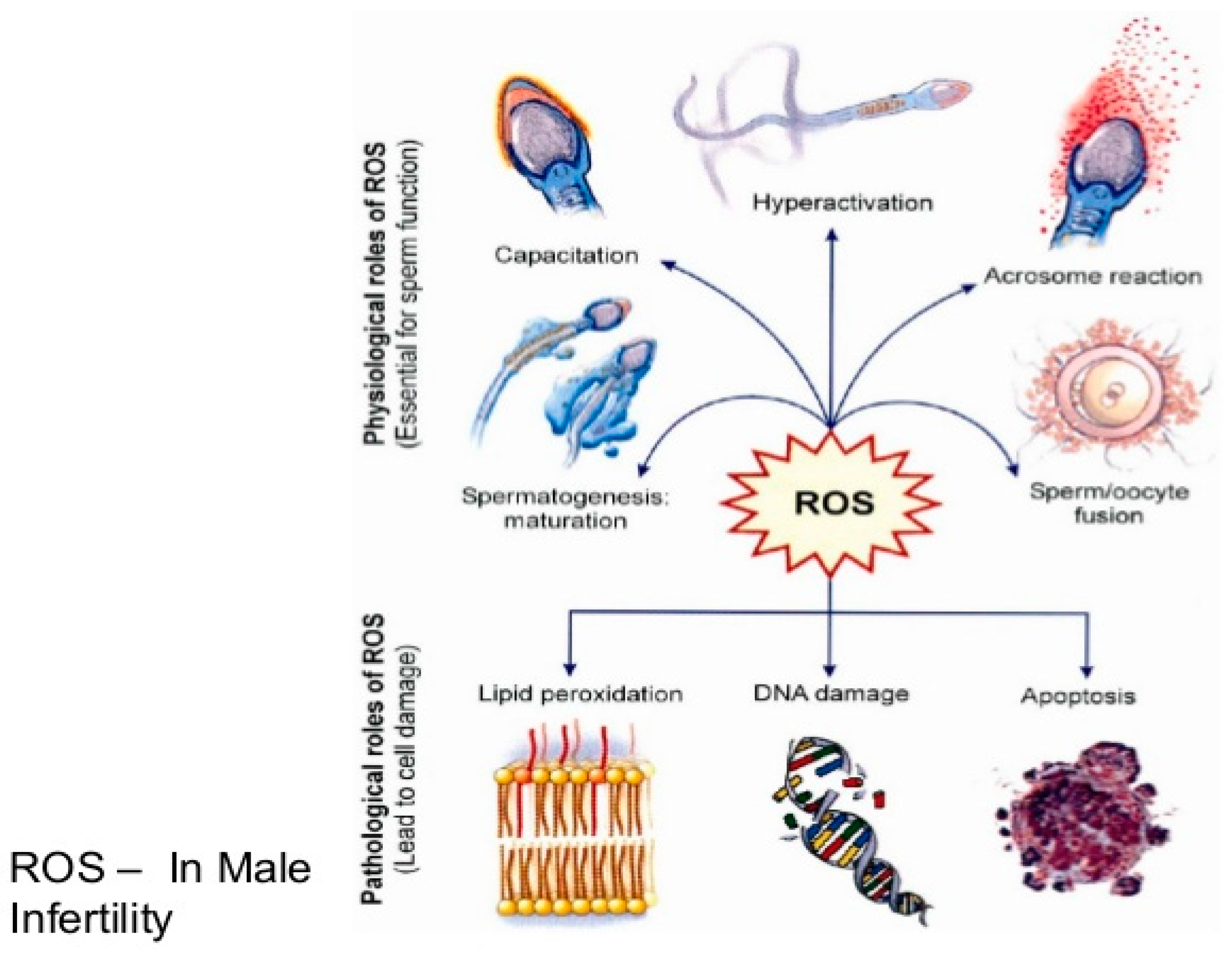
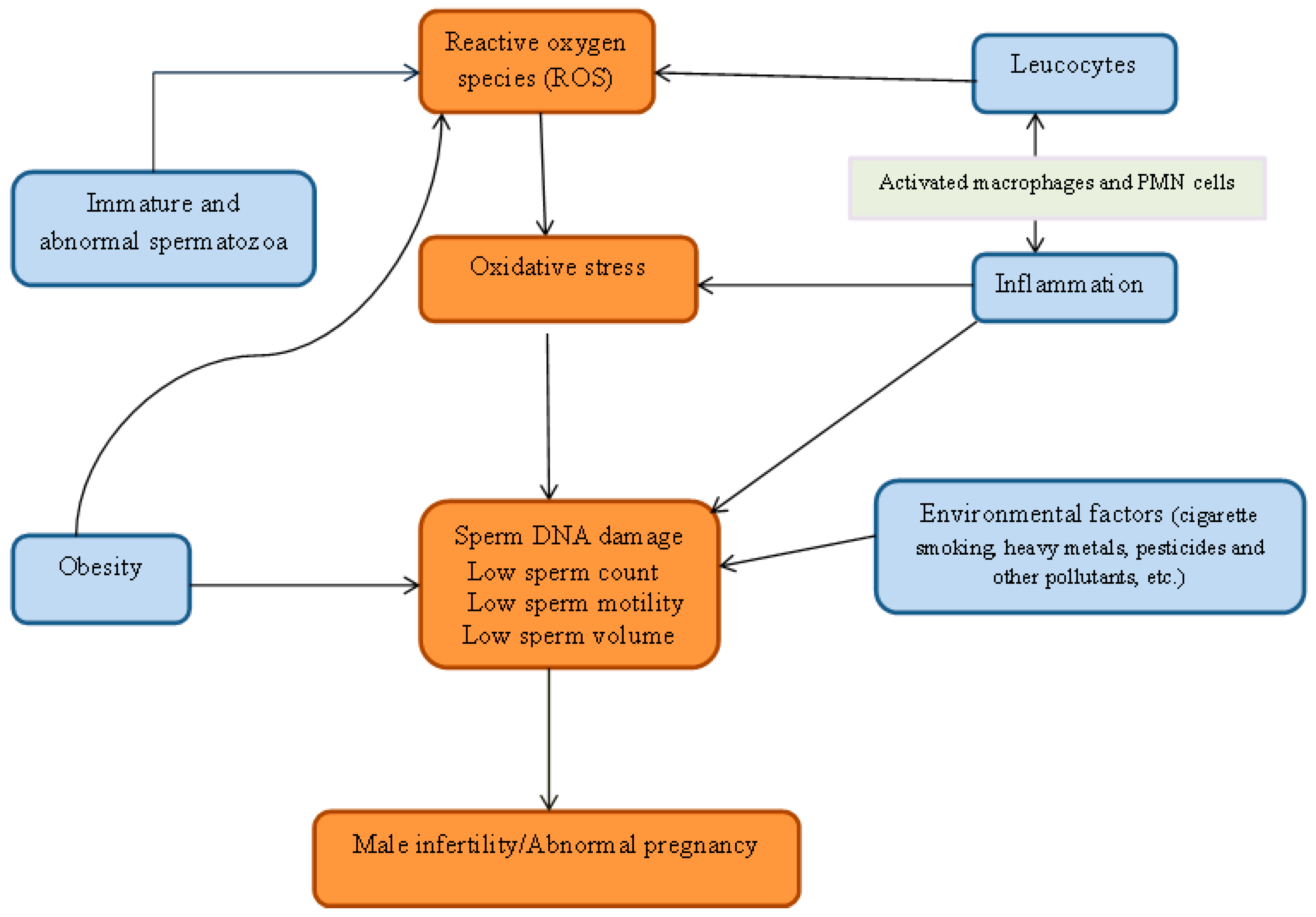
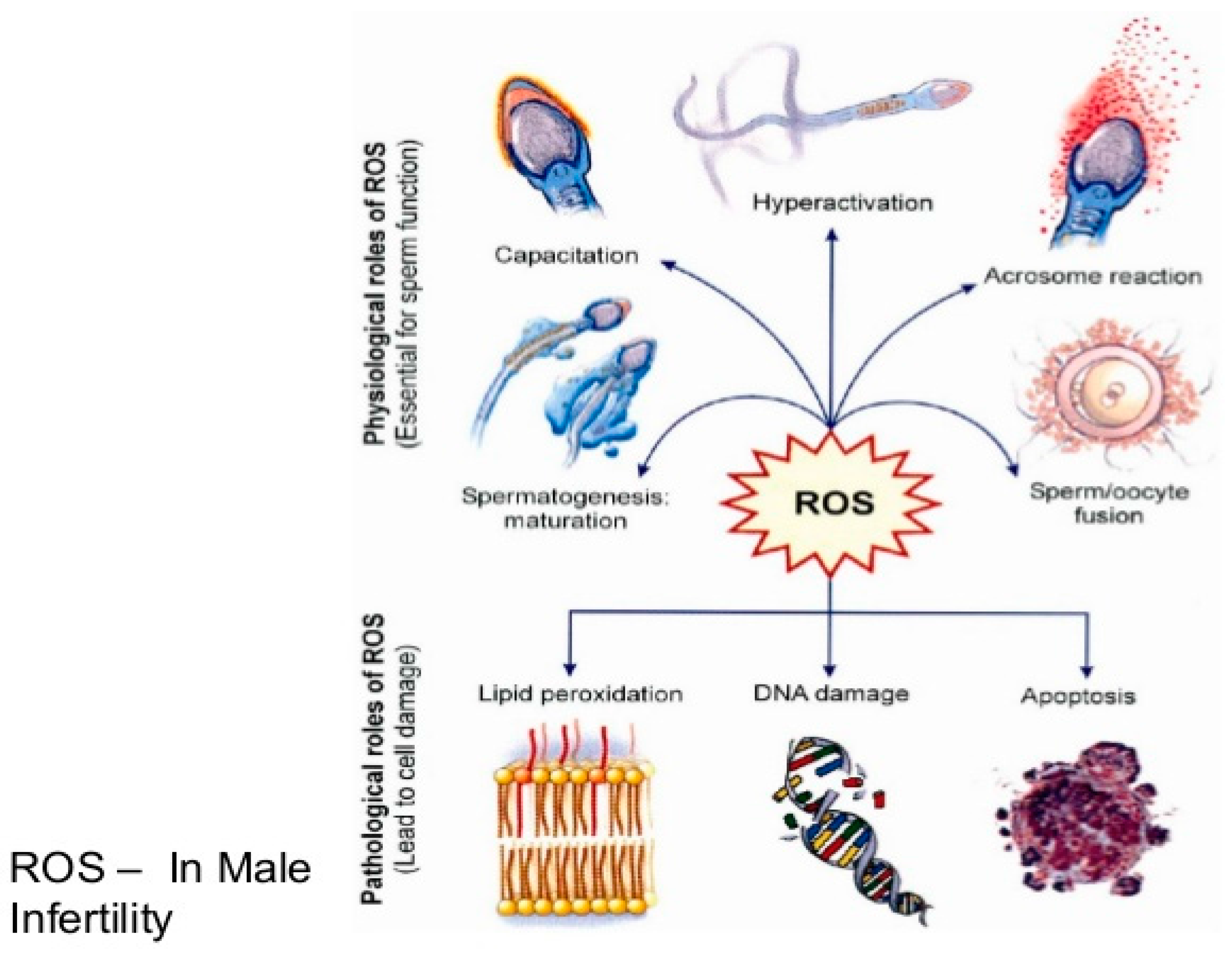
2. Oxidative Stress
3. Origin of ROS
4. Environmental Pollutants
5. Inflammation, Oxidative Stress and Male Fertility
6. Obesity and Infertility
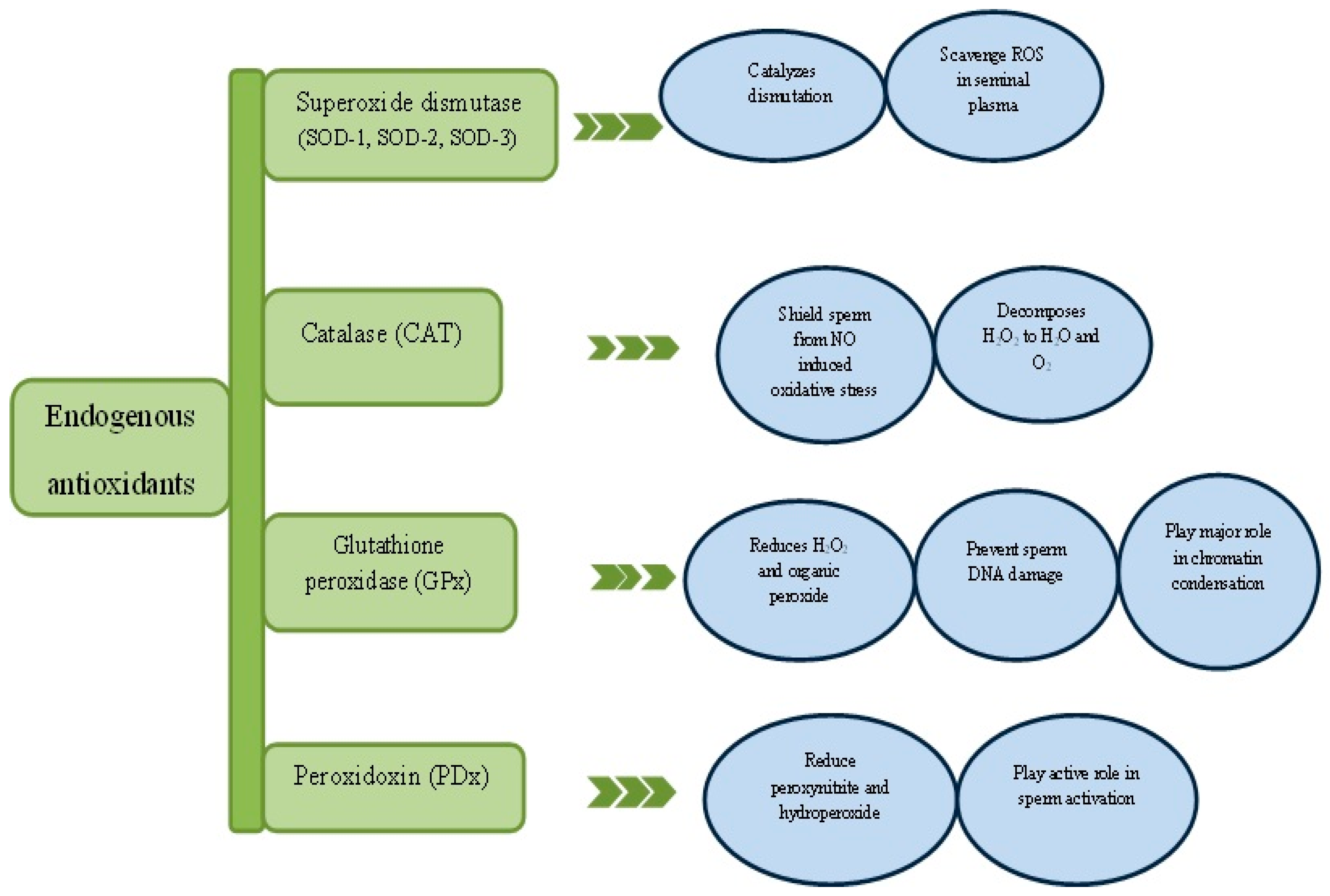
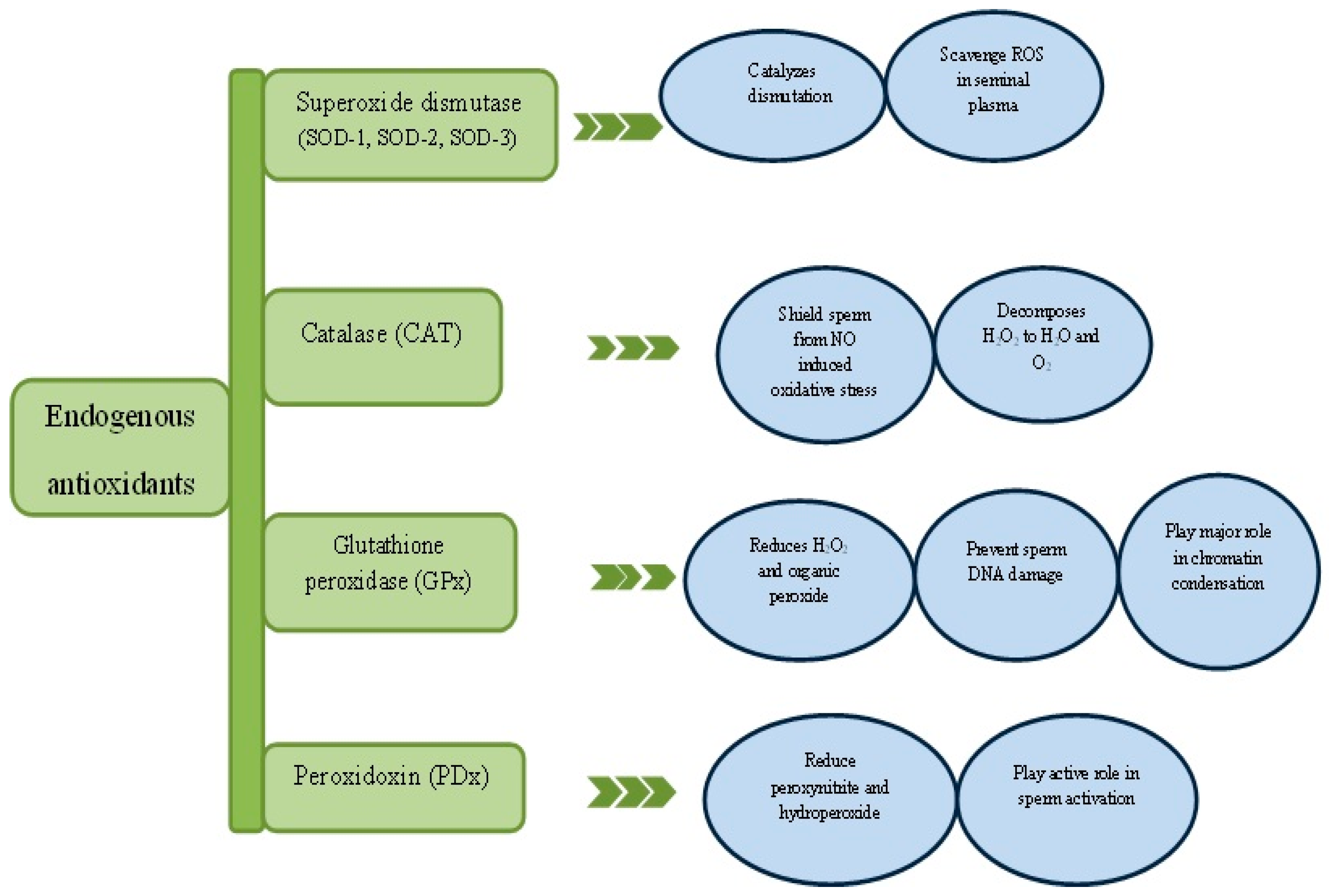
7. Natural Antioxidants and Spermatogenesis
8. Peroxidoxins
9. Exogenous Antioxidants
9.1. Carnitines
9.2. Vitamin E
9.3. Vitamin C
9.4. Carotenoids
10. Herbal Remedy for Male Infertility
10.1. Eurycoma longifolia Jack
10.2. Cardiospermum halicacabum
10.3. Grape Seed Extract
10.4. Marjoram Essential Oil
10.5. Syzygium aromaticum
10.6. Nigella sativa
10.7. Lycium barbarum
10.8. Tribulus terrestris
10.9. Asteracantha longifolia
10.10. Polycarpea corybosa
11. Discussion
12. Conclusions
Conflicts of Interest
References
- Alkhedaide, A.; Alshehri, Z.S.; Sabry, A.; Abdel-Ghaffar, T.; Soliman, M.M.; Attia, H. Protective effect of grape seed extract against cadmium-induced testicular dysfunction. Mol. Med. Rep. 2016, 13, 3101–3109. [Google Scholar] [CrossRef] [PubMed]
- Sigman, M.; Lipshultz, L.; Howards, S. Evaluation of the subfertile male. In Infertility in the Male; Lipshultz, L.A., Howards, S.S., Eds.; Chuchill Livingstone: London, UK, 1991. [Google Scholar]
- Brake, A.; Krause, W. Decreasing quality of semen. BMJ Br. Med. J. 1992, 305, 1498. [Google Scholar] [CrossRef]
- Sikka, S.C.; Hellstrom, W.J.G.; Naz, R.K. Pentoxifylline: Role in management of male infertility/mechanisms of action. Mol. Androl. 1993, 5, 220–231. [Google Scholar]
- World Health Organisation. WHO Laboratory Manual for the Examination of Human Semen and Sperm-Cervical Mucus Interaction, 3rd ed.; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- World Health Organization. WHO Monographs on Selected Medicinal Plants (Volume 2); World Health Organization: Geneva, Switzerland, 1999. [Google Scholar]
- Sabra, S.M.; Al-Harbi, M.S. An influential relationship of seminal fluid microbial infections and infertility, Taif Region, KSA. World J. Med. Sci. 2014, 10, 32–37. [Google Scholar]
- Kumar, N.; Singh, A.K. Trends of male factor infertility, an important cause of infertility: A review of literature. J. Hum. Reprod. Sci. 2015, 8, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Men’s Health 2014, 32, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fujii, J.; Imai, H. Redox reactions in mammalian spermatogenesis and the potential targets of reactive oxygen species under oxidative stress. Spermatogenesis 2014, 4, e979108. [Google Scholar] [CrossRef] [PubMed]
- Azantee, Y.A.W.; Lokman, M.I. The future of Azoospermic Patients: In vitro spermatogenesis. Andrology 2015, 4, 1000143. [Google Scholar]
- Gharagozloo, P.; Aitken, R.J. The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Hum. Reprod. 2011, 26, 1628–1640. [Google Scholar] [CrossRef] [PubMed]
- Makker, K.; Agarwal, A.; Sharma, R. Oxidative stress & male infertility. Indian J. Med. Res. 2009, 129, 357–367. [Google Scholar] [PubMed]
- Tay, P.Y.; Tan, C.P.; Abas, F.; Yim, H.S.; Ho, C.W. Assessment of extraction parameters on antioxidant capacity, polyphenol content, epigallocatechin gallate (EGCG), epicatechin gallate (ECG) and iriflophenone 3-C-β-glucoside of agarwood (Aquilaria crassna) young leaves. Molecules 2014, 19, 12304–12319. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.J.; Kim, J.H.; Ryu, C.S.; Lee, J.Y.; Park, J.S.; Chung, D.Y.; Choi, S.Y.; Kim, M.H.; Chun, E.K.; Roh, S.I. Protective effect of antioxidant supplementation in sperm-preparation medium against oxidative stress in human spermatozoa. Hum. Reprod. 2008, 23, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Bakos, H.W.; Henshaw, R.C.; Mitchell, M.; Lane, M. Paternal body mass index is associated with decreased blastocyst development and reduced live birth rates following assisted reproductive technology. Fertil. Steril. 2011, 95, 1700–1704. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.O.; Bakos, H.W.; Fullston, T.; Lane, M. Impact of obesity on male fertility, sperm function and molecular composition. Spermatogenesis 2012, 2, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Azantee, Y.A.W.; Lokman, M.I.; Roszaman, R. Spermatogonial Stem Cells Protein Identification in In Vitro Culture from Non-Obstructive Azoospermia Patient. Malays. J. Med. Sci. 2016, 23, 40–48. [Google Scholar]
- Aitken, R.J. Free radicals, lipid peroxidation and sperm function. Reprod. Fertil. Dev. 1995, 7, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Buckingham, D.W.; Brindle, J.; Lanzafame, F.; Irvine, D.S.; Aitken, R.J. Development of an image analysis system to monitor the retention of residual cytoplasm by human spermatozoa: Correlation with biochemical markers of the cytoplasmic space, oxidative stress, and sperm function. J. Androl. 1996, 17, 276–287. [Google Scholar] [PubMed]
- Aitken, R.J.; Fisher, H.M.; Fulton, N.; Gomez, E.; Knox, W.; Lewis, B.; Irvine, S. Reactive oxygen species generation by human spermatozoa is induced by exogenous NADPH and inhibited by the flavoprotein inhibitors diphenylene iodonium and quinacrine. Mol. Reprod. Dev. 1997, 47, 468–482. [Google Scholar] [CrossRef]
- Aitken, R.J.; Buckingham, D.W.; West, K.M. Reactive oxygen species and human spermatozoa: Analysis of the cellular mechanisms involved in luminol- and lucigenin-dependent chemiluminescence. J. Cell. Physiol. 1992, 151, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Gavella, M.; Lipovac, V. NADH-dependent oxidoreductase (diaphorase) activity and isozyme pattern of sperm in infertile men. Arch. Androl. 1992, 28, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P.; Darzynkiewicz, Z.; Melamed, M.R. Simultaneous measurement by flow cytometry of sperm cell viability and mitochondrial membrane potential related to cell motility. J. Histochem. Cytochem. 1982, 30, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Pasqualotto, F.F.; Sharma, R.K.; Potts, J.M.; Nelson, D.R.; Thomas, A.J.; Agarwal, A. Seminal oxidative stress in patients with chronic prostatitis. Urology 2000, 55, 881–885. [Google Scholar] [CrossRef]
- Plante, M. Reactive oxygen species, ROS, released by activated neutrophils, but not by deficient spermatozoa, are sufficient to affect normal sperm motility. Fertil. Steril. 1994, 62, 387–393. [Google Scholar] [CrossRef]
- Rahman, M.S.; Kwon, W.S.; Lee, J.S.; Yoon, S.J.; Ryu, B.Y.; Pang, M.G. Bisphenol-A affects male fertility via fertility-related proteins in spermatozoa. Sci. Rep. 2015, 5, 9169. [Google Scholar] [CrossRef] [PubMed]
- Close, C.E.; Roberts, P.L.; Berger, R.E. Cigarettes, alcohol and marijuana are related to pyospermia in infertile men. J. Urol. 1990, 144, 900–903. [Google Scholar] [PubMed]
- Potts, R.J.; Newbury, C.J.; Smith, G.; Notarianni, L.J.; Jefferies, T.M. Sperm chromatin damage associated with male smoking. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1999, 423, 103–111. [Google Scholar] [CrossRef]
- Sheweita, S.A.; Tilmisany, A.M.; Al-Sawaf, H. Mechanisms of male infertility: Role of antioxidants. Curr. Drug Metabol. 2016, 6, 495–501. [Google Scholar] [CrossRef]
- Kvietys, P.R.; Granger, D.N. Role of reactive oxygen and nitrogen species in the vascular responses to inflammation. Free Radic. Biol. Med. 2012, 52, 556–592. [Google Scholar] [CrossRef] [PubMed]
- Adewoyin, M.; Mohsin, S.M.N.; Arulselvan, P.; Hussein, M.Z.; Fakurazi, S. Enhanced anti-inflammatory potential of cinnamate-zinc layered hydroxide in lipopolysaccharide-stimulated RAW 264.7 macrophages. Drug Des. Dev. Ther. 2015, 9, 2475–2484. [Google Scholar]
- O’Bryan, M.K.; Schlatt, S.; Gerdprasert, O.; Phillips, D.J.; de Kretser, D.M.; Hedger, M.P. Inducible nitric oxide synthase in the rat testis: Evidence for potential roles in both normal function and inflammation-mediated infertility. Biol. Reprod. 2000, 63, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, O.; Bahrainwala, J.; Chandrasekaran, S.; Kothari, S.; Mathur, P.P.; Agarwal, A. Impact of inflammation on male fertility. Front. Biosci. (Elite Ed.) 2010, 3, 89–95. [Google Scholar]
- Hales, D.B. Interleukin-1 inhibits Leydig cell steroidogenesis primarily by decreasing 17 alpha-hydroxylase/C17-20 lyase cytochrome P450 expression. Endocrinology 1992, 131, 2165–2172. [Google Scholar] [PubMed]
- Liew, S.H.; Meachem, S.J.; Hedger, M.P. A stereological analysis of the response of spermatogenesis to an acute inflammatory episode in adult rats. J. Androl. 2007, 28, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Tremellen, K. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod. Update 2008, 14, 243–258. [Google Scholar] [CrossRef] [PubMed]
- D’agata, R.; Vicari, E.; Moncada, M.L.; Sidoti, G.; Calogero, A.E.; Fornito, M.C.; Polosa, P. Generation of reactive oxygen species in subgroups of infertile men. Int. J. Androl. 1990, 13, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.D.; Yuan, W.; Zhou, W.J.; Cui, Y.Q.; Wu, J.Q.; Gao, E.S. Do reproductive hormones explain the association between body mass index and semen quality? Asian J. Androl. 2007, 9, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.; Romijn, J.C.; Wildhagen, M.F.; Weber, R.F.; Dohle, G.R. Sperm chromatin structure is associated with the quality of spermatogenesis in infertile patients. Fertil. Steril. 2010, 94, 1748–1752. [Google Scholar] [CrossRef] [PubMed]
- Akpinar, E.; Bashan, I.; Bozdemir, N.; Saatci, E. Which is the best anthropometric technique to identify obesity: Body mass index, waist circumference or waist-hip ratio? Coll. Antropol. 2007, 31, 387–393. [Google Scholar] [PubMed]
- Cabler, S.; Agarwal, A.; Flint, M.; du Plessis, S.S. Obesity: Modern man’s fertility nemesis. Asian J. Androl. 2010, 12, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Kashou, A.H.; Plessis, S.S.; Agarwal, A. The Role of Obesity in ROS Generation and Male Infertility. In Studies on Men’s Health and Fertility; Humana Press: New York, NY, USA, 2012; pp. 571–590. [Google Scholar]
- Crha, I.; Hruba, D.; Ventruba, P.; Fiala, J.; Totusek, J.; Visnova, H. Ascorbic acid and infertility treatment. Cent. Eur. J. Public Health 2003, 11, 63–67. [Google Scholar] [PubMed]
- Kehrer, J.P.; Smith, C.V. Free radicals in biology: Sources, reactivities, and roles in the etiology of human diseases. In Natural Antioxidants in Human Health and Disease; Academie Press: San Diego, CA, USA, 1994; pp. 25–62. [Google Scholar]
- Jacob, R.A. The integrated antioxidant system. Nutr. Res. 1995, 15, 755–766. [Google Scholar] [CrossRef]
- Walczak-Jedrzejowska, R.; Wolski, J.K.; Slowikowska-Hilczer, J. The role of oxidative stress and antioxidants in male fertility. Cent. Eur. J. Urol. 2013, 66, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.; Kurpisz, M. The redox system in human semen and peroxidative damage of spermatozoa. Postepy Higieny i Medycyny Doswiadczalnej (Online) 2004, 59, 523–534. [Google Scholar]
- Gałecka, E.; Jacewicz, R.; Mrowicka, M.; Florkowski, A.; Gałecki, P. Antioxidative enzymes—Structure, properties, functions. Polski Merkuriusz Lekarski: Organ Polskiego Towarzystwa Lekarskiego 2008, 25, 266–268. [Google Scholar]
- Kaur, P.; Bansal, M.P. Influence of selenium induced oxidative stress on spermatogenesis and lactate dehydrogenase-X in mice testis. Asian J. Androl. 2004, 6, 227–232. [Google Scholar]
- Aitken, R.J.; Roman, S.D. Antioxidant systems and oxidative stress in the testes. Oxid. Med. Cell. Longev. 2008, 1, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Scibior, D.; Czeczot, H. Catalase: Structure, properties, functions. Postepy Hig. Med. Dosw. 2006, 60, 170–180. [Google Scholar]
- Wood, Z.A.; Schröder, E.; Harris, J.R.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef]
- Kang, S.W.; Chae, H.Z.; Seo, M.S.; Kim, K.; Baines, I.C.; Rhee, S.G. Mammalian peroxiredoxin isoforms can reduce hydrogen peroxide generated in response to growth factors and tumor necrosis factor-α. J. Biol. Chem. 1998, 273, 6297–6302. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, C. The Enzymatic Antioxidant System of Human Spermatozoa. Adv. Androl. 2014, 2014, 626374. [Google Scholar]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [PubMed]
- Banmeyer, I.; Marchand, C.; Verhaeghe, C.; Vucic, B.; Rees, J.F.; Knoops, B. Overexpression of human peroxiredoxin 5 in subcellular compartments of chinese hamster ovary cells: Effects on cytotoxicity and DNA damage caused by peroxides. Free Radic. Biol. Med. 2004, 36, 65–77. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, C. Peroxiredoxins: Hidden players in the antioxidant defence of human spermatozoa. Basic Clin. Androl. 2014, 24, 4. [Google Scholar] [CrossRef] [PubMed]
- Bøhmer, T.; Hoel, P.; Purvis, K.; Hansson, V. Carnitine levels in human accessory sex organs. Arch. Androl. 1978, 1, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Bieber, L.L. Carnitine. Annu. Rev. Biochem. 1988, 57, 261–283. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Said, T.M. Carnitines and male infertility. Reprod. Biomed. Online 2004, 8, 376–384. [Google Scholar] [CrossRef]
- Palamanda, J.R.; Kehrer, J.P. Involvement of vitamin E and protein thiols in the inhibition of microsomal lipid peroxidation by glutathione. Lipids 1993, 28, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, S.A.; Elamin Ali, M.; Zaki, Z.M.S.; El-Malik, E.M.A.; Nasr, M.A. Lipid peroxidation and human sperm motility: Protective role of vitamin E. J. Androl. 1996, 17, 530–537. [Google Scholar] [PubMed]
- Moslemi, M.K.; Tavanbakhsh, S. Selenium-vitamin E supplementation in infertile men: Effects on semen parameters and pregnancy rate. Int. J. Gen. Med. 2011, 4, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Vézina, D.; Mauffette, F.; Roberts, K.D.; Bleau, G. Selenium-vitamin E supplementation in infertile men. Biol. Trace Elem. Res. 1996, 53, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, S. Male infertility: Nutritional and environmental considerations. Altern. Med. Rev. 2000, 5, 28–38. [Google Scholar] [PubMed]
- Subramanian, V.S.; Marchant, J.S.; Reidling, J.C.; Said, H.M. N-Glycosylation is required for Na+-dependent vitamin C transporter functionality. Biochem. Biophys. Res. Commun. 2008, 374, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Khan, A.; Khattak, M.M.A.K. Biological significance of ascorbic acid (Vitamin C) in human health—A review. Pak. J. Nutr. 2004, 3, 5–13. [Google Scholar]
- Kallner, A.; Hartmann, D.; Hornig, D. Steady-state turnover and body pool of ascorbic acid in man. Am. J. Clin. Nutr. 1979, 32, 530–539. [Google Scholar] [PubMed]
- Angulo, C.; Maldonado, R.; Pulgar, E.; Mancilla, H.; Córdova, A.; Villarroel, F.; Castro, M.A.; Concha, I.I. Vitamin C and oxidative stress in the seminiferous epithelium. Biol. Res. 2011, 44, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Brzozowski, T.; Kwiecien, S.; Konturek, P.C.; Konturek, S.J.; Mitis-Musiol, M.; Duda, A.; Hahn, E.G. Comparison of nitric oxide-releasing NSAID and vitamin C with classic NSAID in healing of chronic gastric ulcers; involvement of reactive oxygen species. Med. Sci. Monit. 2001, 7, 592–599. [Google Scholar] [PubMed]
- Guaiquil, V.H.; Farber, C.M.; Golde, D.W.; Vera, J.C. Efficient transport and accumulation of vitamin C in HL-60 cells depleted of glutathione. J. Biol. Chem. 1997, 272, 9915–9921. [Google Scholar] [PubMed]
- Dawson, E.B.; Harris, W.A.; Rankin, W.E.; Charpentier, L.A.; McGANITY, W.J. Effect of ascorbic acid on male fertility. Ann. N. Y. Acad. Sci. 1987, 498, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.A.; Pianalto, F.S.; Agee, R.E. Cellular ascorbate depletion in healthy men. J. Nutr. 1992, 122, 1111–1118. [Google Scholar] [PubMed]
- Fraga, C.G.; Motchnik, P.A.; Shigenaga, M.K.; Helbock, H.J.; Jacob, R.A.; Ames, B.N. Ascorbic acid protects against endogenous oxidative DNA damage in human sperm. Proc. Natl. Acad. Sci. USA 1991, 88, 11003–11006. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.E.; Sterling, E.S.L.; Young, I.S.; Thompson, W. Comparison of individual antioxidants of sperm and seminal plasma in fertile and infertile men. Fertil. Steril. 1997, 67, 142–147. [Google Scholar] [CrossRef]
- Acharya, U.R.; Mishra, M.; Patro, J.; Panda, M.K. Effect of vitamins C and E on spermatogenesis in mice exposed to cadmium. Reprod. Toxicol. 2008, 25, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.V.; Ray, M.R.; Rao, L.G. Lycopene. Adv. Food Nutr. Res. 2006, 51, 99–164. [Google Scholar] [PubMed]
- Atasoy, N. Biochemistry of lycopene. J. Anim. Vet. Adv. 2012, 11, 2605–2610. [Google Scholar] [CrossRef]
- Rao, A.V.; Agarwal, S. Role of lycopene as antioxidant carotenoid in the prevention of chronic diseases: A review. Nutr. Res. 1999, 19, 305–323. [Google Scholar] [CrossRef]
- Zini, A.; San Gabriel, M.; Libman, J. Lycopene supplementation in vitro can protect human sperm deoxyribonucleic acid from oxidative damage. Fertil. Steril. 2010, 94, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- Palan, P.; Naz, R. Changes in various antioxidant levels in human seminal plasma related to immunoinfertility. Arch. Androl. 1996, 36, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Mangiagalli, M.G.; Martino, P.A.; Smajlovic, T.; Guidobono Cavalchini, L.; Marelli, S.P. Effect of lycopene on semen quality, fertility and native immunity of broiler breeder. Br. Poult. Sci. 2010, 51, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D.; Agarwal, A.; Ong, C.; Prashast, P. Lycopene and male infertility. Asian J. Androl. 2014, 16, 420–425. [Google Scholar] [PubMed]
- Miyake, K.; Tezuka, Y.; Awale, S.; Li, F.; Kadota, S. Quassinoids from Eurycoma longifolia. J. Nat. Prod. 2009, 2, 2135–2140. [Google Scholar] [CrossRef] [PubMed]
- Ang, H.H.; Hitotsuyanagi, Y.; Fukaya, H.; Takeya, K. Quassinoids from Eurycoma longifolia. Phytochemistry 2002, 59, 833–837. [Google Scholar] [CrossRef]
- Bedir, E.; Abou-Gazar, H.; Ngwendson, J.N.; Khan, I.A. Eurycomaoside: A new quassinoid-type glycoside from the roots of Eurycoma longifolia. Chem. Pharm. Bull. 2003, 51, 1301–1303. [Google Scholar] [CrossRef] [PubMed]
- Sholikhah, E.N.; Wijayanti, M.A.; Nurani, L.H. Stage specificity of pasak bumi root (Eurycoma longifolia Jack) isolate on Plasmodium falciparum cycles. Med. J. Malays. 2008, 63, 98–99. [Google Scholar]
- Chan, K.L.; Low, B.S.; Teh, C.H.; Das, P.K. The effect of Eurycoma longifolia on sperm quality of male rats. Nat. Prod. Commun. 2009, 4, 1331–1336. [Google Scholar] [PubMed]
- Tambi, M.I.B.M.; Imran, M.K.; Henkel, R.R. Standardised water-soluble extract of Eurycoma longifolia, Tongkat Ali, as testosterone booster for managing men with late-onset hypogonadism? Andrologia 2012, 44, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Solomon, M.C.; Erasmus, N.; Henkel, R.R. In vivo effects of Eurycoma longifolia Jack (Tongkat Ali) extract on reproductive functions in the rat. Andrologia 2014, 46, 339–348. [Google Scholar]
- Ali, J.M.; Saad, J.M. Biochemical Effect of Eurycoma longifolia Jack on the Sexual Behavior, Fertility, Sex Hormone, and Glycolysis. Ph.D. Dissertation, Department of Biochemistry, University Malaya, Malaysia of Malaysia, Malaysia, 1993. [Google Scholar]
- Androherb 2017. Available online: http://www.androherb.com/tongkat-ali (accessed on 15 February 2017).
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, regional, and global trends in infertility prevalence since 1990: A systematic analysis of 277 health surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed]
- Peiris, L.D.C.; Dhanushka, M.A.T.; Jayathilake, T.A.H.D.G. Evaluation of Aqueous Leaf Extract of Cardiospermum halicacabum (L.) on Fertility of Male Rats. BioMed Res. Int. 2015, 2015, 175726. [Google Scholar] [CrossRef] [PubMed]
- Altrogiornale 2017. Available online: http://www.altrogiornale.org/wp-content/uploads/2015/05/3-CARDIOSPERMUM-HALICACABUM.jpg (accessed on 15 February 2017).
- Shenoy, S.F.; Keen, C.L.; Kalgaonkar, S.; Polagruto, J.A. Effects of grape seed extract consumption on platelet function in postmenopausal women. Thromb. Res. 2007, 121, 431–432. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Yu, J.; Pohorly, J.E.; Kakuda, Y. Polyphenolics in grape seeds-biochemistry and functionality. J. Med. Food 2003, 6, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Tyagi, A.K.; Dhanalakshmi, S.; Agarwal, R.; Agarwal, C. Grape seed extract inhibits advanced human prostate tumor growth and angiogenesis and upregulates insulin-like growth factor binding protein-3. Int. J. Cancer 2004, 108, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Hala, A.H.; Khattab, Z.A.; Abdallah, G.; Kamel, M. Grape seed extract alleviate reproductive toxicity caused by aluminium chloride in male rats. J. Am. Sci. 2010, 6, 352–361. [Google Scholar]
- Bayatli, F.; Akkuş, D.; Kilic, E.; Saraymen, R.; Sönmez, M.F. The protective effects of grape seed extract on MDA, AOPP, apoptosis and eNOS expression in testicular torsion: An experimental study. World J. Urol. 2013, 31, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Amazing Olive 2017. Available online: http://www.amazingolive.com/uploads/7/2/4/2/72424671/s328690282648775400_p46_i1_w1500.jpeg (accessed on 15 February 2017).
- Skidmore-Roth, L. Mosby’s Handbook of Herbs and Natural Supplements; Mosby. Inc.: St. Louis, MO, USA, 2001; p. 369. [Google Scholar]
- El-Wakf, A.M.; Elhabibi, E.S.M.; El-Ghany, E.A. Preventing male infertility by marjoram and sage essential oils through modulating testicular lipid accumulation and androgens biosynthesis disruption in a rat model of dietary obesity. Egypt. J. Basic Appl. Sci. 2015, 2, 167–175. [Google Scholar] [CrossRef]
- El-Ashmawy, I.M.; Saleh, A.; Salama, O.M. Effects of marjoram volatile oil and grape seed extract on ethanol toxicity in male rats. Basic Clin. Pharm. Toxicol. 2007, 101, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Rocket Gardens 2017. Available online: https://www.rocketgardens.co.uk/growing-guides/marjoram-2/ (accessed on 15 February 2017).
- Gurib-Fakim, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Mol. Asp. Med. 2006, 27, 1–93. [Google Scholar] [CrossRef] [PubMed]
- Boudou, F.; Berroukche, A.; Bendahmane-Salmi, M.; Kandouci, B.A.; Tou, N. Ameliorative Effects of Syzygium aromaticum Essential Oil on Fertility in Male Rats Exposed to Manganese. Adv. Sex. Med. 2013, 3, 37412. [Google Scholar]
- Trade Winds 2017. Available online: http://www.tradewindsfruit.com/content/clove.htm (accessed on 15 February 2017).
- Haseena, S.; Aithal, M.; Das, K.K.; Saheb, S.H. Phytochemical Analysis of Nigella sativa and it’s effect on reproductive system. J. Pharm. Sci. Res. 2015, 7, 514–517. [Google Scholar]
- Pure Life 2017. Available online: http://www.purelife-eg.net/product-details.aspx?id=58 (accessed on 15 February 2017).
- Gao, X.M.; Xu, Z.M.; Li, Z.W. Traditional Chinese Medicines; People’s Health Publishing House: Beijing, China, 2000. [Google Scholar]
- Proven Winners 2017. Available online: https://www.provenwinners.com/plants/lycium/big-lifeberry-goji-berry-lycium-barbarum (accessed on 15 February 2017).
- Luo, Q.; Li, Z.; Huang, X.; Yan, J.; Zhang, S.; Cai, Y.Z. Lycium barbarum polysaccharides: Protective effects against heat-induced damage of rat testes and H2O2-induced DNA damage in mouse testicular cells and beneficial effect on sexual behavior and reproductive function of hemicastrated rats. Life Sci. 2006, 79, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Nair, V.; Gupta, Y.K. Evaluation of the aphrodisiac activity of Tribulus terrestris Linn. in sexually sluggish male albino rats. J. Pharmacol. Pharmacother. 2012, 3, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Guan, Y.; Liu, J.; Zhai, F.; Zhang, X.; Guan, L. Cellular and molecular mechanisms in vascular smooth muscle cells by which total saponin extracted from Tribulus terrestris protects against artherosclerosis. Cell. Physiol. Biochem. 2013, 32, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Elahi, R.K.; Asl, S.; Shahian, F. Study on the effects of various doses of Tribulus terrestris extract on epididymal sperm morphology and count in rat. Glob. Vet. 2013, 10, 13–17. [Google Scholar]
- Shalaby, M.; Hammouda, A. Assessment of Protective and Antioxidant Properties of Tribulus terrestris Fruits against Testicular Toxicity in Rats. J. Intercult. Ethnopharmacol. 2014, 3, 113. [Google Scholar] [CrossRef] [PubMed]
- The Supplement Reviews 2017. Available online: http://thesupplementreviews.org/testosterone-boosters/ingredients/the-benefits-and-side-effects-of-tribulus-terrestris/ (accessed on 15 February 2017).
- Kirtikar, K.R.; Basu, B.D. Terminalia chebula. Indian Med. Plants 1935, 1, 1020–1023. [Google Scholar]
- Chauhan, N.S.; Sharma, V.; Dixit, V.K. Effect of Asteracantha longifolia seeds on the sexual behaviour of male rats. Nat. Prod. Res. 2011, 15, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.K.; Ghandare, B. Effect of Hygrophila spinosa on reproductive function of male albino rats. J. Complement. Integr. Med. 2010, 7. [Google Scholar] [CrossRef]
- Ayurvediccommunity 2017. Available online: http://www.ayurvediccommunity.com/Botany.asp?Botname=Asteracantha%20longifolia (accessed on 15 February 2017).
- Damodharan, R.; Chandrasekharan, C.C.; Quereshi, M.I.; Shankar, B.P.; Kumara, D.S. Studies on the Effect of Asteracantha longifolia Seed Powder on Cadmium Chloride Induced Testicular (Micrometry) Changes in Albino Rats. Vet. Res. Int. 2015, 3, 7–11. [Google Scholar]
- Lafuente, A.; Márquez, N.; Pérez-Lorenzo, M.; Pazo, D.; Esquifino, A.I. Pubertal and postpubertal cadmium exposure differentially affects the hypothalamic–pituitary–testicular axis function in the rat. Food Chem. Toxicol. 2000, 38, 913–923. [Google Scholar] [CrossRef]
- Pillai, A.; Rawal, A.; Gupta, S. Effect of low level exposure of lead and cadmium on hepatic estradiol metabolism in female rats. Indian J. Exp. Biol. 2002, 40, 807–811. [Google Scholar] [PubMed]
- Balamurugan, K.; Nishanthini, A.; Mohan, V.R. Anticancer activity of ethanol extract of Polycarpaea corymbosa (L.) Lam whole plant against Dalton Ascites Lymphoma. Int. J. Pharm. Biol. Sci. 2013, 4, 296–303. [Google Scholar]
- Mohan, V.R.; Balamurugan, K.; Sakthidevi, G. Fertility enhancement of corymbosa (L.) Lam (Caryophyllaceae) whole plant on male albino rats. Asian J. Pharm. Clin. Res. 2013, 6 (Suppl. 5), 151–155. [Google Scholar]
- Wikiwand 2017. Available online: http://www.wikiwand.com/es/Polycarpaea_corymbosa (accessed on 15 February 2017).
- Nantia, E.A.; Moundipa, P.F.; Monsees, T.K.; Carreau, S. Medicinal plants as potential male anti-infertility agents: A review. Basic Clin. Androl. 2009, 19, 148–158. [Google Scholar] [CrossRef]
- Elumalai, P.; Krishnamoorthy, G.; Selvakumar, K.; Arunkumar, R.; Venkataraman, P.; Arunakaran, J. Studies on the protective role of lycopene against polychlorinated biphenyls (Aroclor 1254)-induced changes in StAR protein and cytochrome P450 scc enzyme expression on Leydig cells of adult rats. Reprod. Toxicol. 2009, 27, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, P.; Muthusamy, T.; Balasubramanian, K.; and Arunakaran, J. Effects of vitamins C and E on steroidogenic enzymes mRNA expression in polychlorinated biphenyl (Aroclor 1254) exposed adult rat Leydig cells. Toxicology 2007, 232, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Mennella, M.R.F.; Jones, R. Properties of spermatozoal superoxide dismutase and lack of involvement of superoxides in metal-ion-catalysed lipid-peroxidation and reactions in semen. Biochem. J. 1980, 191, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Sanocka, D.; Miesel, R.; Jędrzejczak, P.; CHEŁMONSKA-SOYTA, A.; Kurpisz, M. Effect of reactive oxygen species and the activity of antioxidant systems on human semen; association with male infertility. Int. J. Androl. 1997, 20, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Said, T.M.; Aziz, N.; Sharma, R.K.; Lewis-Jones, I.; Thomas, A.J.; Agarwal, A. Novel association between sperm deformity index and oxidative stress-induced DNA damage in infertile male patients. Asian J. Androl. 2005, 7, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Deepinder, F.; Cocuzza, M.; Agarwal, A. Should seminal oxidative stress measurement be offered routinely to men presenting for infertility evaluation? Endocr. Pract. 2008, 14, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Kwon, W.S.; Oh, S.A.; Kim, Y.J.; Rahman, M.S.; Park, Y.J.; Pang, M.G. Proteomic approaches for profiling negative fertility markers in inferior boar spermatozoa. Sci. Rep. 2015, 5, 13821. [Google Scholar] [CrossRef] [PubMed]
- Sikka, S.C. Relative impact of oxidative stress on male reproductive function. Curr. Med. Chem. 2001, 8, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Jahan, N.; Radhakrishnan, G.; Banerjee, B.D. To Evaluate the Efficacy of Combination Antioxidant Therapy on Oxidative Stress Parameters in Seminal Plasma in the Male Infertility. J. Clin. Diagn. Res. JCDR 2016, 10, QC14–QC17. [Google Scholar] [CrossRef] [PubMed]
- Kwon, W.S.; Rahman, M.S.; Lee, J.S.; Yoon, S.J.; Park, Y.J.; Pang, M.G. Discovery of predictive biomarkers for litter size in boar spermatozoa. Mol. Cell. Proteom. 2015, 14, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Kwon, W.S.; Rahman, M.S.; Pang, M.G. Diagnosis and prognosis of male infertility in mammal: The focusing of tyrosine phosphorylation and phosphotyrosine proteins. J. Proteome Res. 2014, 13, 4505–4517. [Google Scholar] [CrossRef] [PubMed]
- Karande Vishvanath 2017. Available online: http://www.inviafertility.com/blog/blog/infertility/drvkarande/anti-oxidants-and-male-fertility/ (accessed on 15 February 2017).
- Bello, N.B.; Rabiu, A.M.; Odeh, S.O. The Effect of Aqueous Leave Extract of Nicotiana Tabacum (Tobacco) on Some Reproductive Parameters and Micro-Anatomical Architecture of the Testis In Male Albino Wistar Rats. Natu. Sci. Res. 2013, 5, 137–143. [Google Scholar]
- Rehman, S.U.; Choe, K.; Yoo, H.H. Review on a traditional herbal medicine, Eurycoma longifolia jack (Tongkat Ali): Its traditional uses, chemistry, evidence-based pharmacology and toxicology. Molecules 2016, 21, 331. [Google Scholar] [CrossRef] [PubMed]
- Low, B.S.; Das, P.K.; Chan, K.L. Standardized quassinoid-rich Eurycoma longifolia extract improved spermatogenesis and fertility in male rats via the hypothalamic–pituitary–gonadal axis. J. Ethnopharmacol. 2013, 145, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.S.; Sharma, V.; Dixit, V.K.; Thakur, M. A review on plants used for improvement of sexual performance and virility. BioMed Res. Int. 2014, 2014, 868062. [Google Scholar] [CrossRef] [PubMed]
- Kotta, S.; Ansari, S.H.; Ali, J. Exploring scientifically proven herbal aphrodisiacs. Pharmacogn. Rev. 2013, 7, 1–10. [Google Scholar] [PubMed]
- Venkatachalapathi, A.; Sangeeth, T.; Ali, M.A.; Tamilselvi, S.S.; Paulsamy, S. Ethnomedicinal assessment of Irula tribes of Walayar valley of Southern Western Ghats, India. Saudi J. Biol. Sci. 2016, in press. [Google Scholar] [CrossRef]
- Hafsa, S.H.A.; Ayman, H.A.; Helham, A.S.; Popescu, A.; Mocuta, D. Grape Seed (Vitis vinifera L.) alleviate reproductive toxicity caused by lindane in new zeeland white male rabbits. In Scientific Papers Series Management, Economic Engineering in Agriculture and Rural Development; RAWEX-COMS DOMINOR: Bucharest, Romania, 2016; Volume 16. [Google Scholar]
- Mishra, R.K.; Singh, S.K. Biphasic effect of Syzygium aromaticum flower bud on reproductive physiology of male mice. Andrologia 2016, 48, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.K.; Singh, S.K. Reproductive effects of lipid soluble components of Syzygium aromaticum flower bud in male mice. J. Ayurveda Integr. Med. 2013, 4, 94. [Google Scholar] [PubMed]
- Mahdavi, R.; Heshmati, J.; Namazi, N. Effects of black seeds (Nigella sativa) on male infertility: A systematic review. J. Herb. Med. 2015, 5, 133–139. [Google Scholar] [CrossRef]
- Parandin, R.; Yousofvand, N.; Ghorbani, R. The enhancing effects of alcoholic extract of Nigella sativa seed on fertility potential, plasma gonadotropins and testosterone in male rats. Iran. J. Reprod. Med. 2012, 10, 355–362. [Google Scholar] [PubMed]
- Hamady, J.J.; Ganim, K.G.; Ali, Z.S. Effect of Methanolic and Phenolic Extracts of Nigella sativa Seeds on Testicular Expression Levels of inhibin alpha and beta Genes in Mature Male Wistar Rats. J. Appl. Pharm. Sci. 2016, 6, 118–121. [Google Scholar] [CrossRef]
- Cho Ping, N.; Hashim, N.H.; Hasan Adli, D.S. Effects of Nigella sativa (Habbatus sauda) oil and nicotine chronic treatments on sperm parameters and testis histological features of rats. Evid. Based Complement. Altern. Med. 2014, 2014, 218293. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhou, Z.W.; Sheng, H.P.; He, L.J.; Fan, X.W.; He, Z.X.; Sun, T.; Zhang, X.; Zhao, R.J.; Gu, L.; et al. An evidence-based update on the pharmacological activities and possible molecular targets of Lycium barbarum polysaccharides. Drug Des. Dev. Ther. 2015, 9, 33–78. [Google Scholar]
- Shi, G.; Zheng, J.; Wu, J.; Qiao, H.; Chang, Q.; Niu, Y.; Sun, T.; Li, Y.; Yu, J. Beneficial effects of Lycium barbarum polysaccharide on spermatogenesis by improving antioxidant activity and inhibiting apoptosis in streptozotocin-induced diabetic male mice. Food Funct. 2017. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Huang, Q.; Zhao, K.; Shang, P. Biological activities and potential health benefit effects of polysaccharides isolated from Lycium barbarum L. Int. J. Boil. Macromol. 2013, 54, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.W.M.; Chia-Di Lee, J.; Li, Y.; Fung, S.M.Y.; Sang, Y.H.; Shen, J.; Chang, R.C.C.; So, K.F. Polysaccharides from wolfberry prevents corticosterone-induced inhibition of sexual behavior and increases neurogenesis. PLoS ONE 2012, 7, e33374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, A.; Sun, X.; Li, X.; Zhao, X.; Li, S.; Ma, A. Protective effects of Lycium barbarum polysaccharides on testis spermatogenic injury induced by bisphenol A in mice. Evid. Based Complement. Altern. Med. 2013, 2013, 690808. [Google Scholar]
- Mohaned, M.M. Effects of Tribulus terrestris ethanolic extract in male rats & cocks fertility. J. Pharm. Biomed. Sci. 2013, 30, S13–S18. [Google Scholar]
- Singh, S.; Gupta, Y.K. Aphrodisiac activity of Tribulus terrestris Linn. in experimental models in rats. J. Men’s Health 2011, 8, S75–S77. [Google Scholar] [CrossRef]
- Biswas, T.K.; Pandit, S.; Jana, U. In Search of Spermatogenetic and Virility Potential Drugs of Ayurvedic Leads: A Review. Andrology (Los Angel) 2015, 4, 148. [Google Scholar] [CrossRef]
- Tag, H.M.; Abdelazek, H.M.; Mahoud, Y.S.; EL-Shenawy, N.S. Efficacy of Tribulus terrestris extract and metformin on fertility indices and oxidative stress of testicular tissue in streptozotocin-induced diabetic male rats. Afr. J. Pharm. Pharmacol. 2015, 9, 1088–1098. [Google Scholar]
- Moghaddam, M.H.G.; Khalili, M.; Maleki, M.; Abadi, M.E.A. The effect of oral feeding of Tribulus terrestris L. on sex hormone and gonadotropin levels in addicted male rats. Int. J. Fertil. Steril. 2013, 7, 57–62. [Google Scholar]
- Singh, R.; Ali, A.; Jeyabalan, G.; Semwal, A. An overview of the current methodologies used for evaluation of aphrodisiac agents. J. Acute Dis. 2013, 2, 85–91. [Google Scholar] [CrossRef]
- Vyas, N.Y.; Raval, M.A. Aphrodisiac and spermatogenic potential of alkaloidal fraction of Hygrophila spinosa T. Ander in rats. J. Ethnopharmacol. 2016, 194, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.A.; Singh, S.A.; Jeyabalan, G.; Ali, A.S.; Semwal, A.L. Medicinal plants used to treat sexual dysfunction: A review. Int. J. Rec. Adv. Pharm. Res. 2013, 3, 29–35. [Google Scholar]
- Chandra, S.; Rawat, D.S. Medicinal plants of the family Caryophyllaceae: A review of ethno-medicinal uses and pharmacological properties. Integr. Med. Res. 2015, 4, 123–131. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Botanical Name | Effect on Fertility | Other Medicinal Applications | References |
|---|---|---|---|---|
| Tongkat Ali | E. longifolia | Prevents hypogonadism, stimulates biosynthesis of androgens | antimalarial, anticancer, antibacterial | [90,91,144,145,146,147] |
| Baloon vine | C. halicacabum | Enhances caput and epididymal sperm count, sperm motility and serum testosterone | Rheumatism, bleeding piles and snake bite | [94,95,148] |
| Grapevine | V. vinifera | Improves sperm profile; Protect sperm from DNA damage; alleviate apoptosis of germ cell | Antiinflammatory, antioxidant, antimicrobial, hepatoprotective | [100,101,149] |
| Marjoram | O. marojana | Increases sperm cells and androgens; protect sex organs; increases serum level of T | For treating cramps, coughs, dizziness, depression etc. | [104,105] |
| Clove | S. aromaticum | Cure for sexual dysfunction and low libido | Treatment of dental disorder, headache and respiratory diseases, | [107,108,150,151] |
| Black seed | N. sativa | Enhances levels of T and L hormone | For cooking, antiviral, antiinflammatory, immunomodulatory | [110,152,153,154,155] |
| Wolfberry | L. barbarum | Protect sperm cells from DNA damage; increases serum T | General medicine | [156,157,158,159,160,161] |
| Puncture vine | T. terrestris | Increases weight of testis and seminal vesicle and serum level of T | Cure for inflammation, edema, ascite | [117,118,147,162,163,164,165] |
| Hygrophila | A. longifolia | Increases weight of sertoli cells; enhances micrometric measurement of spermatogonia, spermatocytes and spermatids | Cure for diarrhea, dysentery and anemia | [121,122,124,166,167] |
| Pallipoondu | P. carymbosa | Increases levels of T and L hormone; enhances weights of testis, epididymis and vas deferens | Antiinflammatory, hepatoprotective | [128,168,169] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adewoyin, M.; Ibrahim, M.; Roszaman, R.; Isa, M.L.M.; Alewi, N.A.M.; Rafa, A.A.A.; Anuar, M.N.N. Male Infertility: The Effect of Natural Antioxidants and Phytocompounds on Seminal Oxidative Stress. Diseases 2017, 5, 9. https://doi.org/10.3390/diseases5010009
Adewoyin M, Ibrahim M, Roszaman R, Isa MLM, Alewi NAM, Rafa AAA, Anuar MNN. Male Infertility: The Effect of Natural Antioxidants and Phytocompounds on Seminal Oxidative Stress. Diseases. 2017; 5(1):9. https://doi.org/10.3390/diseases5010009
Chicago/Turabian StyleAdewoyin, Malik, Muhammad Ibrahim, Ramli Roszaman, Muhammad Lokman Md Isa, Nur Aizura Mat Alewi, Ainin Azwani Abdul Rafa, and Mohd Nur Nasyriq Anuar. 2017. "Male Infertility: The Effect of Natural Antioxidants and Phytocompounds on Seminal Oxidative Stress" Diseases 5, no. 1: 9. https://doi.org/10.3390/diseases5010009
APA StyleAdewoyin, M., Ibrahim, M., Roszaman, R., Isa, M. L. M., Alewi, N. A. M., Rafa, A. A. A., & Anuar, M. N. N. (2017). Male Infertility: The Effect of Natural Antioxidants and Phytocompounds on Seminal Oxidative Stress. Diseases, 5(1), 9. https://doi.org/10.3390/diseases5010009