
Melatonin as Immune Potentiator for Enhancing Subunit Vaccine Efficacy against Bovine Viral Diarrhea Virus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus, Plasmids, and Cells
2.3. Cell Proliferation Test
2.4. Cell Culture
2.5. Protein Extraction and Western Blotting
2.6. RNA Extraction and RT-qPCR
2.7. Animal Design and Sampling
2.8. Immunohistochemistry
2.9. Histology
2.10. Flow Cytometry
2.11. RNA Extraction and RT-qPCR
2.12. Protein Extraction and Western Blot
2.13. Statistical Analysis
3. Results
3.1. Effects of MT on MDBK Cell Proliferation
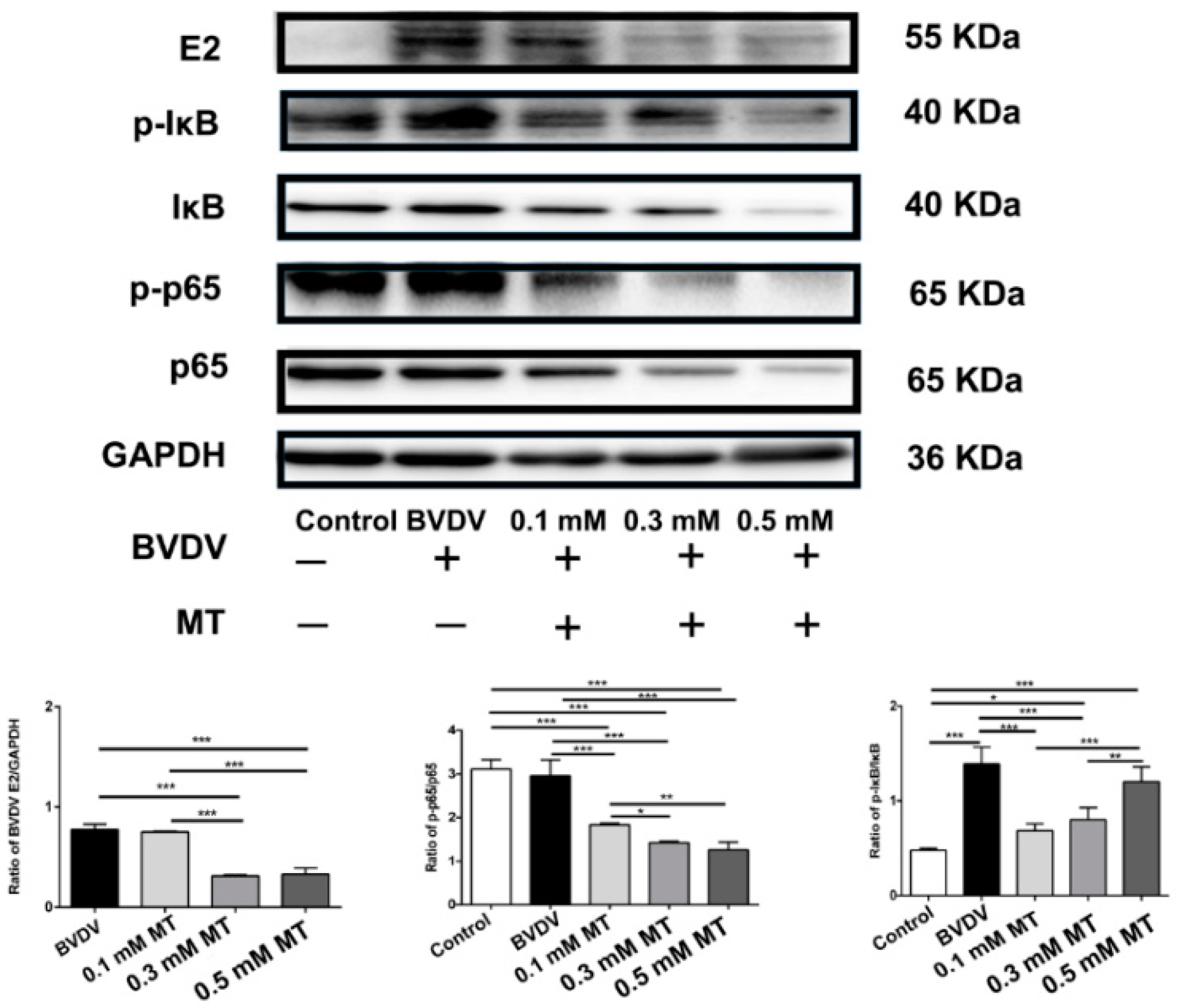
3.2. Effects of MT on the NF-κB Signal Pathway of MDBK Cells
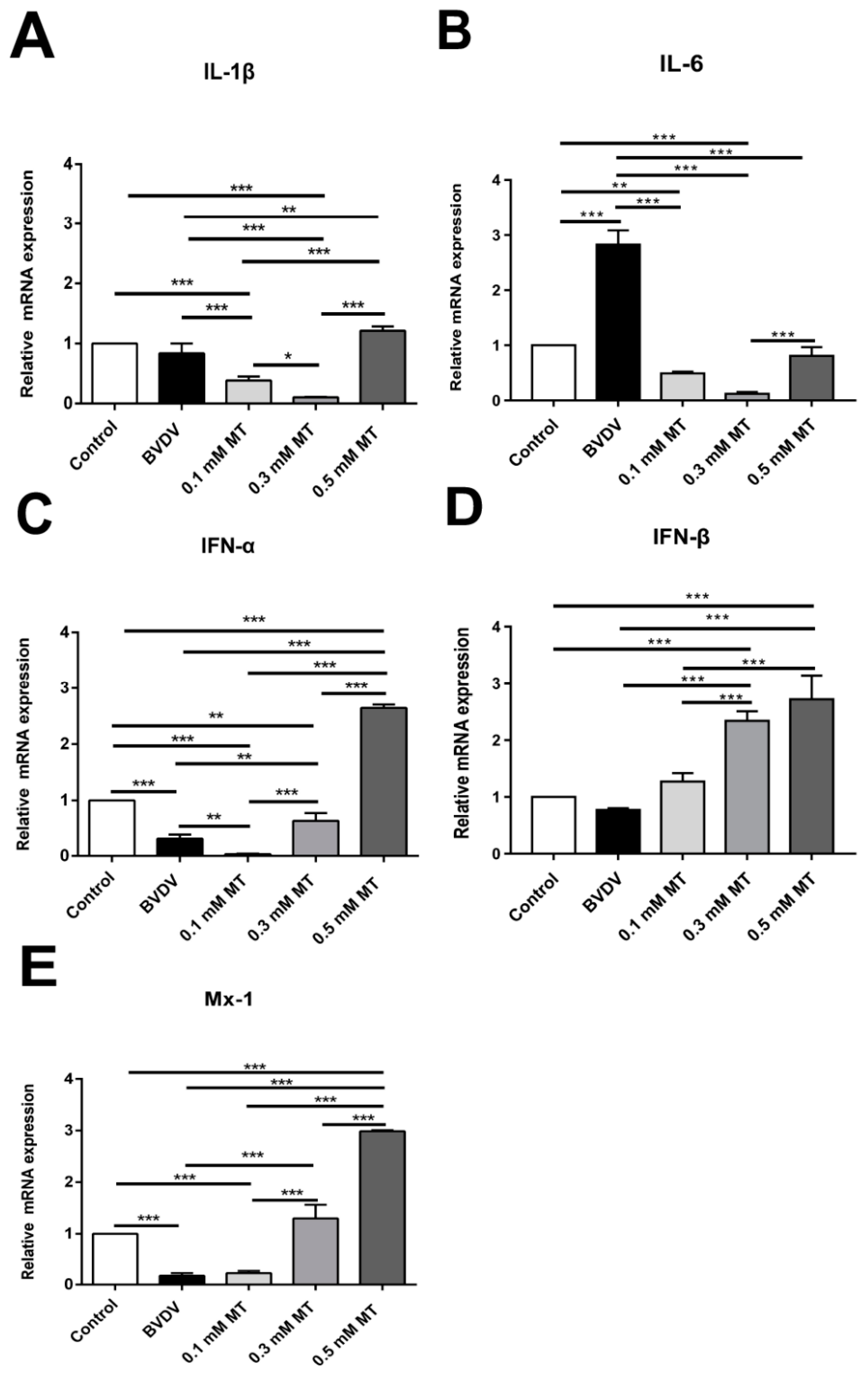
3.3. Effects of MT on the Levels of Inflammation-Mediating Cytokine in MDBK Cells
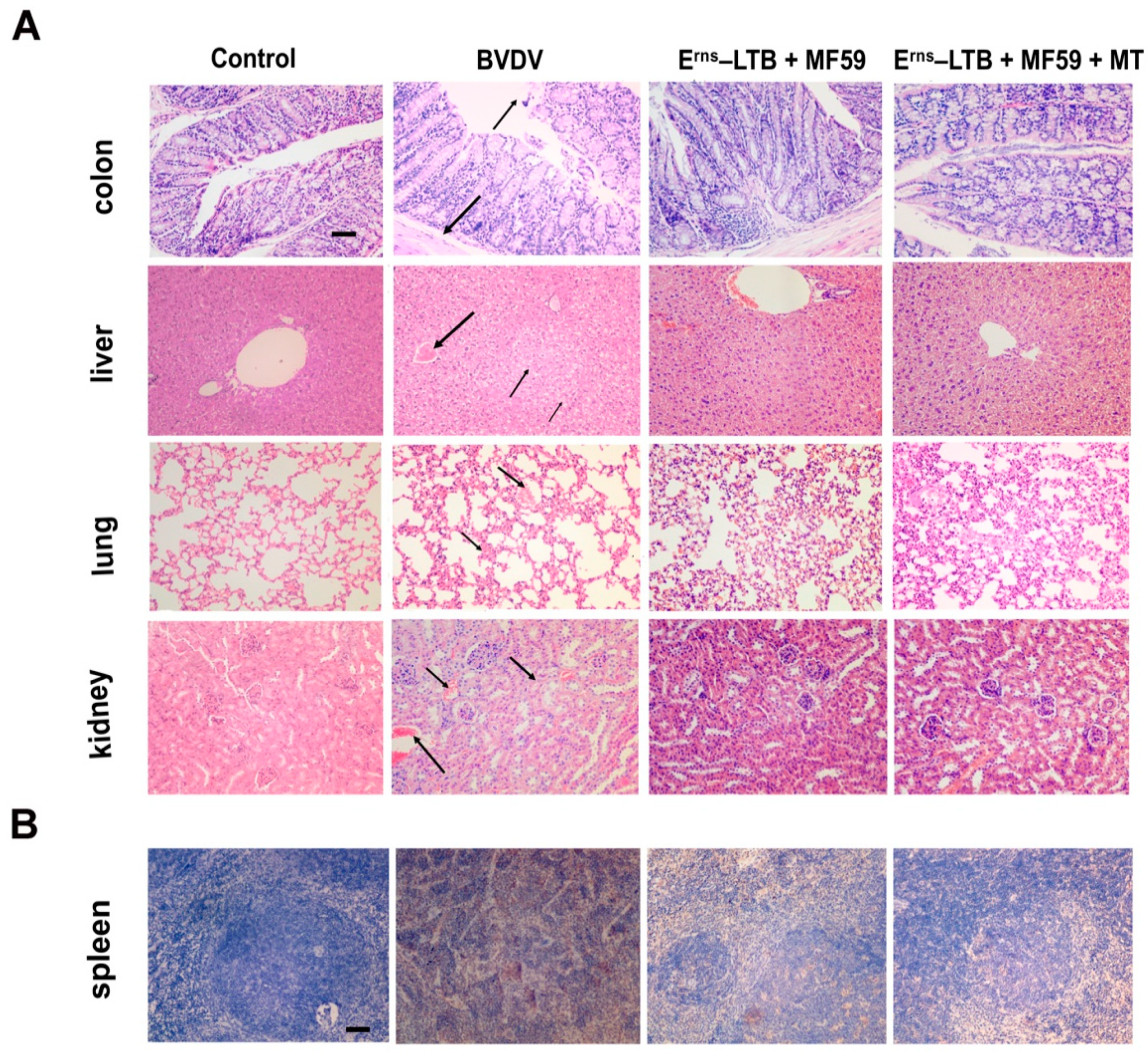
3.4. Histopathology and Immunohistochemistry
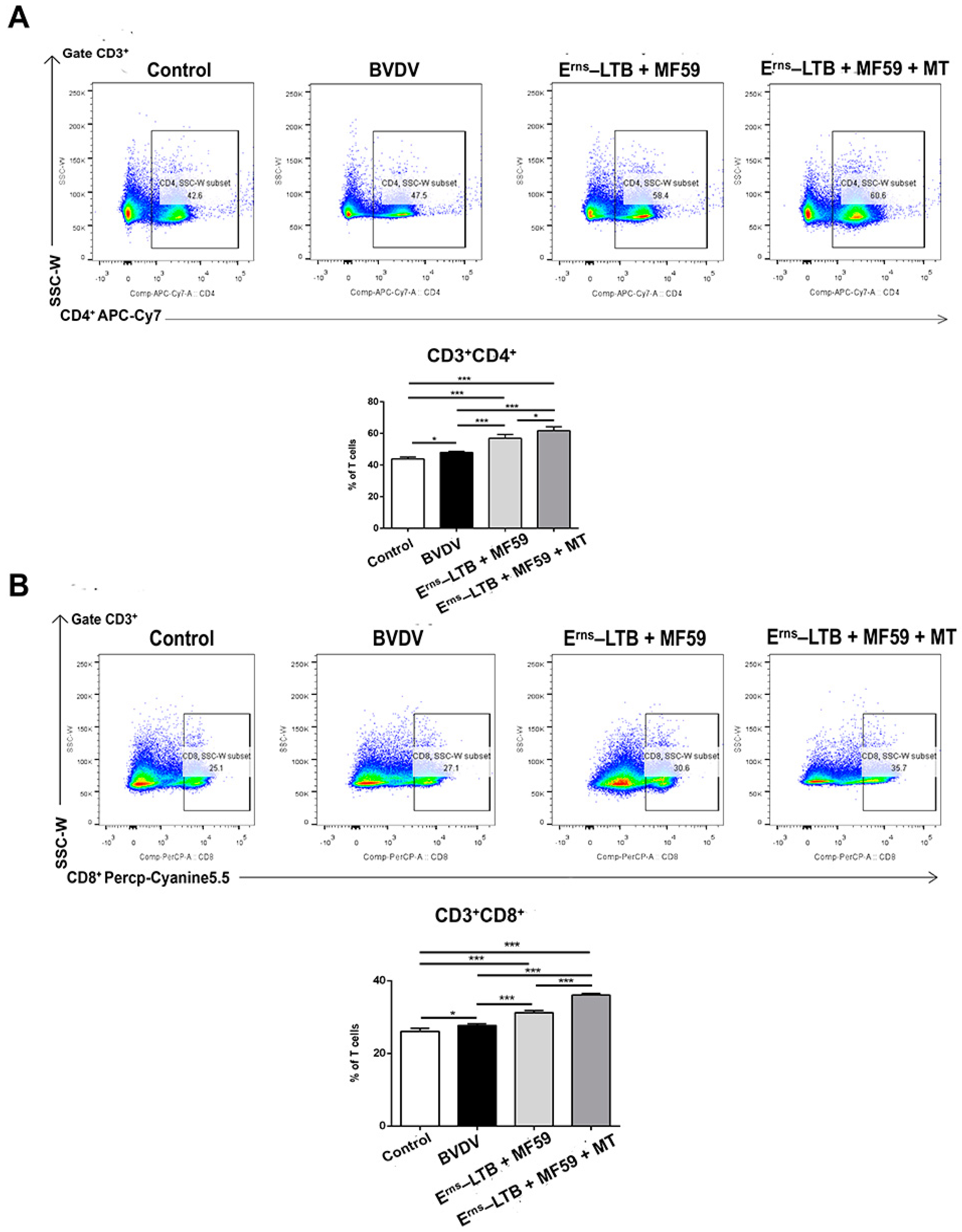
3.5. T Cell Responses Following Immunization
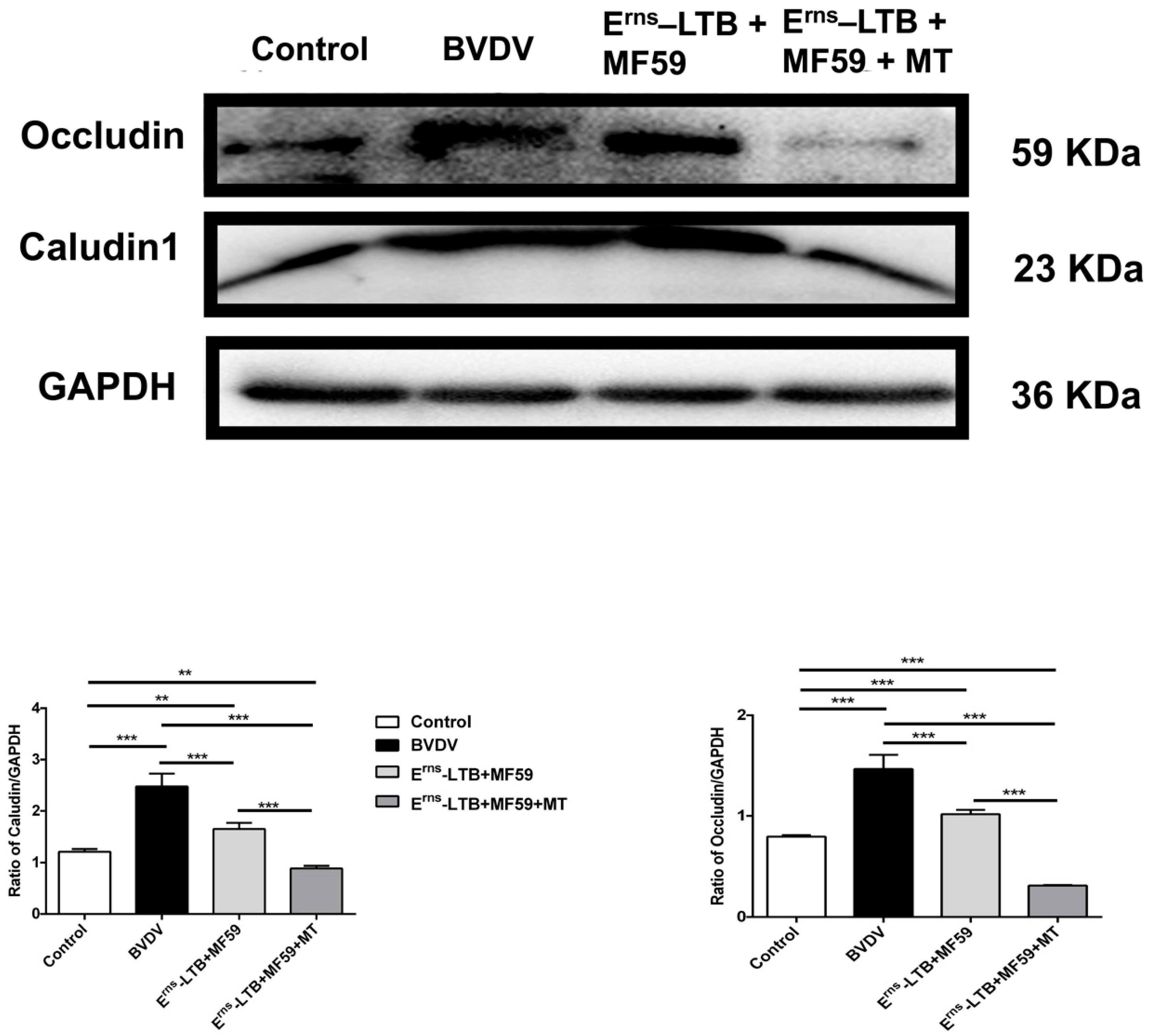
3.6. Colon Permeability
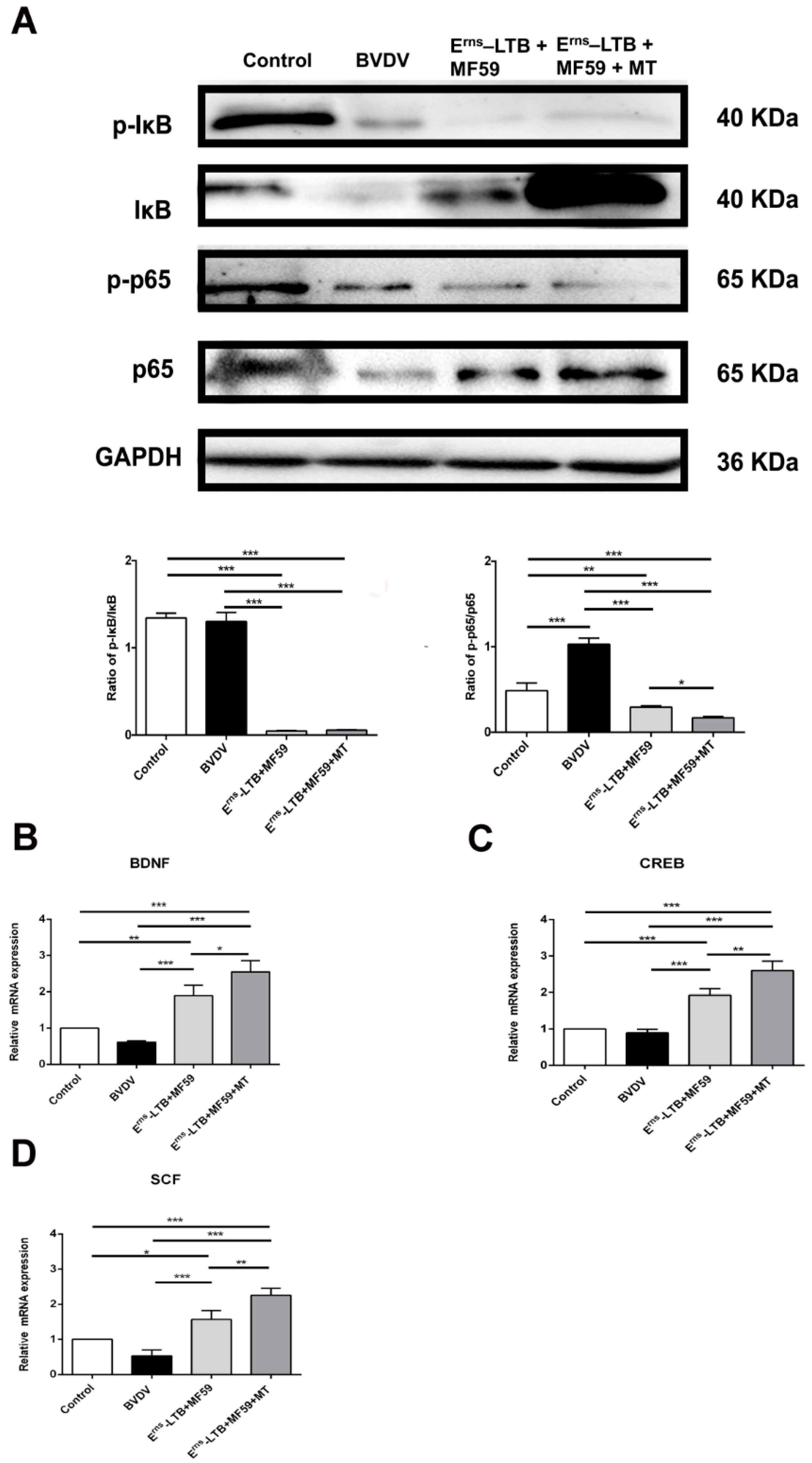
3.7. MT Inhibits the NF-κB Signaling Pathway in the Hippocampus
3.8. MT Repaired the Recognition System in the Hippocampus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garoussi, M.T.; Mehrzad, J.; Nejati, A. Investigation of Persistent Infection of Bovine Viral Diarrhea Virus (BVDV) in Holstein Dairy Cows. Trop. Anim. Health. Prod. 2019, 51, 853–858. [Google Scholar] [CrossRef]
- Kottwitz, J.J.; Ortiz, M. Bovine Viral Diarrhea Virus in Zoos: A Perspective from the Veterinary Team. Front. Microbiol. 2015, 6, 1496. [Google Scholar] [CrossRef]
- Quintana, M.E.; Barone, L.J.; Trotta, M.V.; Turco, C.; Mansilla, F.C.; Capozzo, A.V.; Cardoso, N.P. In-Vivo Activity of IFN-λ and IFN-α against Bovine-Viral-Diarrhea Virus in a Mouse Model. Front. Vet. Sci. 2020, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Knapek, K.J.; Georges, H.M.; Van, C.H.; Bishop, J.V.; Bielefeldt-Ohmann, H.; Smirnova, N.P.; Hansen, T.R. Fetal Lymphoid Organ Immune Responses to Transient and Persistent Infection with Bovine Viral Diarrhea Virus. Viruses 2020, 12, 816. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhang, S.; Gao, X.T.; Guo, X.Y.; Xin, T.; Zhu, H.F.; Jia, H.; Hou, S.H. Experimental Immunization of Mice with a Recombinant Bovine Enterovirus Vaccine Expressing BVDV Erns Protein Elicits a Long-Lasting Serologic Response. Virol. J. 2020, 17, 88. [Google Scholar] [CrossRef]
- Gallina, L.; Koch, M.C.; Gentile, A.; Treglia, I.; Bombardi, C.; Mandrioli, L.; Bolcato, M.; Scagliarini, A.; Drögemüller, C.; Seuberlich, T.; et al. Bovine Viral Diarrhoea Virus 1b Infection Associated with Congenital Tremor and Hypomyelination in Holstein Calves. Vet. Microbiol. 2021, 256, 109047. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Yang, G.H.; Nie, J.W.; Yang, R.J.; Du, M.Z.; Su, J.H.; Wang, J.; Wang, J.F.; Zhu, Y.H. Recombinant Erns-E2 Protein Vaccine Formulated with MF59 and CPG-ODN Promotes T Cell Immunity against Bovine Viral Diarrhea Virus Infection. Vaccine 2020, 38, 3881–3891. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Yang, G.H.; Nie, J.W.; Wang, J.; Wang, Y.X.; Du, M.Z.; Guo, L.; Yang, R.J.; Zhu, Y.H. Immunization with Recombinant E-LTB Fusion Protein Elicits Protective Immune Responses against Bovine Viral Diarrhea Virus. Vet. Microbiol. 2021, 259, 109084. [Google Scholar] [CrossRef]
- Torres, F.J.M.; Arias, C.F. Tight Junctions Go Viral! Viruses 2015, 7, 5145–5154. [Google Scholar] [CrossRef]
- Montgomery, D.L.; Van, O.A.; Van, C.H.; Hansen, T.R. The Fetal Brain in Bovine Viral Diarrhea Virus-Infected Calves: Lesions, Distribution, and Cellular Heterogeneity of Viral Antigen at 190 Days Gestation. Vet. Pathol. 2008, 45, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Więckowska, G.A.; Mietelska, P.A.; Wydrych, M.; Wojda, U. Western Diet as a Trigger of Alzheimer’s Disease: From Metabolic Syndrome and Systemic Inflammation to Neuroinflammation and Neurodegeneration. Ageing Res. Rev. 2021, 70, 101397. [Google Scholar] [CrossRef]
- Villalba, M.; Fredericksen, F.; Otth, C.; Olavarría, V.H. Molecular Characterization of the Bovine IER3 Gene: Down-Regulation of IL-8 by Blocking NF-κB Activity Mediated by IER3 Overexpression in MDBK Cells Infected with Bovine Biral Diarrhea Virus-1. Mol. Immunol. 2017, 92, 169–179. [Google Scholar] [CrossRef]
- Maldonado, N.; Fredericksen, F.; Espiñeira, C.; Toledo, C.; Oltra, J.; La Barra, V.D.; Fernandez, C.; Salvador, J.; Villalba, M.; Olavarría, V.H. BVDV-1 Induces Interferon-Beta Gene Expression through a Pathway Involving IRF1, IRF7, and NF-κB Activation. Mol. Immunol. 2020, 128, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Riitho, V.; Strong, R.; Larska, M.; Graham, S.P.; Steinbach, F. Bovine Pestivirus Heterogeneity and Its Potential Impact on Vaccination and Diagnosis. Viruses 2020, 12, 1134. [Google Scholar] [CrossRef]
- McMorrow, C.; Gunn, A.J.; Khalfan, S.; Hernandez-Jover, M.; Brookes, V.J. Veterinarians’ Knowledge, Attitudes and Practices Associated with Bovine Viral Diarrhoea Virus Control and Prevention in South-East Australia. Animals 2020, 10, 1630. [Google Scholar] [CrossRef]
- Sangewar, N.; Hassan, W.; Lokhandwala, S.; Bray, J.; Reith, R.; Markland, M.; Sang, H.; Yao, J.X.; Fritz, B.; Waghela, S.D.; et al. Mosaic Bovine Viral Diarrhea Virus Antigens Elicit Cross-Protective Immunity in Calves. Front. Immunol. 2020, 11, 589537. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, S.; Liu, R.X.; Hao, G.Y. Genetic Diversity of Bovine Viral Diarrhea Virus Infection in Goats in Southwestern China. J. Vet. Med. 2018, 2018, 8274397. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.G.; Yuan, Y.; Ankenbauer, R.G.; Nelson, L.D.; Witte, S.B.; Jackson, J.A.; Welch, S.W. Construction of Chimeric Bovine Viral Diarrhea Viruses Containing Glycoprotein Erns of Heterologous Pestiviruses and Evaluation of the Chimeras as Potential Marker Vaccines against BVDV. Vaccine 2012, 30, 3843–3848. [Google Scholar] [CrossRef]
- Duan, Q.; Zhang, W. Genetic Fusion Protein 3 × STa-Ovalbumin is an Effective Coating Antigen in ELISA to Titrate Anti-STa Antibodies. Microbiol. Immunol. 2017, 61, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Ruan, A.; Zhou, J.; Huang, L.W.; Zhang, X.Z.; Ma, Y.F.; Wang, Q.F. Cin namic Aldehyde Inhibits Lipopolysaccharide-Induced Chondrocyte Inflammation and Reduces Cartilage Degeneration by Blocking the Nuclear Factor-Kappa B Signaling Pathway. Front. Pharmacol. 2020, 11, 949. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, A.; Liu, X.Y.; Ma, Y.; Zhao, F.F.; Wang, M.Z.; Loor, J.J.; Wang, H.G. Melatonin Ameliorates Ochratoxin A Induced Liver Inflammation, Oxidative Stress and Mitophagy in Mice Involving in Intestinal Microbiota and Restoring the Intestinal Barrier Function. J. Hazard. Mater. 2021, 407, 124489. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.P.; Liu, J.X.; Gu, J.Y.; Liu, F.; Li, J.H.; Yang, B.; Zheng, Y.; Li, J.; Wu, S.H.; Wu, Q.H.; et al. Sarcandra GlabraCombination Effect of Three Main Constituents from Inhibits Oxidative Stress in the Mice Following Acute Lung Injury: A Role of MAPK-NF-κB Pathway. Front. Pharmacol. 2020, 11, 580064. [Google Scholar] [CrossRef]
- Veneroso, C.; Tuñón, M.J.; González-Gallego, J.; Collado, P.S. Melatonin Reduces Cardiac Inflammatory Injury Induced by Acute Exercise. J. Pineal Res. 2009, 47, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.H.; Song, J.; Zhang, H.; Zhang, F.; Liu, H.; Li, L.; Zhang, Z.G.; Chen, L.S.; Zhang, M.; Lin, D.C.; et al. Melatonin Mediated Foxp3-Downregulation Decreases Cytokines Production via the TLR2 and TLR4 Pathways in H. pylori Infected Mice. Int. Immunopharmacol. 2018, 64, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.G.R.; Kuehn, C.C.; Dos Santos, C.D.; Miranda, M.A.; Da Costa, C.M.B.; Mendonça, V.J.; Do Prado, J.C., Jr. Protective Actions of Melatonin against Heart Damage during Chronic Chagas Disease. Acta. Trop. 2013, 128, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ji, H.; Wang, Y.; Gu, C.; Gu, W.; Hu, L.; Zhu, L. Melatonin Alleviates Radiation-Induced Lung Injury via Regulation of MiR-30e/NLRP3 Axis. Oxidative Med. Cell. Longev. 2019, 2019, 4087298. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.C.; Peng, C.K.; Liao, W.I.; Pao, H.P.; Huang, K.L.; Chu, S.J. Melatonin Re ceptor Agonist Protects against Acute Lung Injury Induced by Ventilator through up-Regulation of IL-10 Production. Respir. Res. 2020, 21, 65. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wang, X.B.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Liu, C.W.; Reiter, R.J. COVID-19: Melatonin as a Potential Adjuvant Treatment. Life Sci. 2020, 250, 117583. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Aneja, S.; Kohli, K. Add-on Melatonin Improves Sleep Behavior in Children with Epilepsy: Randomized, Double-Blind, Placebo-Controlled Trial. J. Child. Neuro. 2005, 20, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Regodón, S.; Ramos, A.; Morgado, S.; Tarazona, R.; Martín, P.P.; Rosado, J.A.; Míguez, M.D.P. Melatonin Enhances the Immune Response to Vaccination against A1 and C Strains of Dichelobacter Nodosus. Vaccine 2009, 27, 1566–1570. [Google Scholar] [CrossRef]
- Cao, Y.; Zhu, X.; Hossen, M.N.; Kakar, P.; Zhao, Y.; Chen, X. Augmentation of Vaccine Induced Humoral and Cellular Immunity by a Physical Radiofrequency Adjuvant. Nat. Commun. 2018, 9, 3695. [Google Scholar] [CrossRef]
- Mosca, F.; Tritto, E.; Muzzi, A.; Monaci, E.; Bagnoli, F.; Iavarone, C. Molecular and Cellular Signatures of Human Vaccine Adjuvants. Proc. Natl. Acad. Sci. USA 2008, 105, 10501–10506. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine Adjuvants: Understand ing the Structure and Mechanism of Adjuvanticity. Vaccine 2019, 37, 3141–3250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Suo, M.Y.; Liu, L.X.; Qi, Y.; Zhang, C.; Xie, L.; Zheng, X.H.; Ma, C.; Li, J.Y.; Yang, J.M.; et al. Melatonin Alleviates Contrast-Induced Acute Kidney Injury by Activation of Sirt3. Oxid. Med. Cell. Longev. 2021, 2021, 6668887. [Google Scholar] [CrossRef]
- Song, Q.J.; Weng, X.G.; Cai, D.J.; Zhang, W.; Wang, J.F. Forsythoside A Inhibits BVDV Replication via TRAF2-Dependent CD28-4-1BB Signaling in Bovine PBMCs. PLoS ONE 2016, 11, e0162791. [Google Scholar] [CrossRef] [Green Version]
- Hibaoui, Y.; Reutenauer-Patte, J.; Patthey-Vuadens, O.; Ruegg, U.T.; Dorchies, O.M. Melatonin Improves Muscle Function of the Dystrophic Mdx5Cv Mouse, a Model for Duchenne Muscular Dystrophy. J. Pineal Res. 2011, 51, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.S.; Sun, Y.; Xin, J.G.; Zhang, T.; Sun, N.; Ni, X.Q.; Zeng, D.; Bai, Y. Lacto bacillus Johnsonii BS15 Prevents Psychological Stress-Induced Memory Dysfunction in Mice by Modulating the Gut-Brain Axis. Front. Microbiol. 2020, 11, 1941. [Google Scholar] [CrossRef]
- Xia, D.Y.; Yang, L.; Li, Y.; Chen, J.Y.; Zhang, X.F.; Wang, H.; Zhai, S.S.; Jiang, X.Z.; Meca, G.; Wang, S.X.; et al. Melatonin Alleviates Ochratoxin A-Induced Liver Inflammation Involved Intestinal Microbiota Homeostasis and Microbiota-Independent Manner. J. Hazard. Mater. 2021, 413, 125239. [Google Scholar] [CrossRef] [PubMed]
- Bonmati-Carrion, M.A.; Tomas-Loba, A. Melatonin and Cancer: A Polyhedral Net Work Where the Source Matters. Antioxidants 2021, 10, 210. [Google Scholar] [CrossRef]
- Zhao, C.N.; Wang, P.; Mao, Y.M.; Dan, Y.L.; Wu, Q.; Li, X.M.; Wang, D.G.; Davis, C.; Hu, W.B.; Pan, H.F. Potential Role of Melatonin in Autoimmune Diseases. Cytokine Growth Factor Rev. 2019, 48, 1–10. [Google Scholar] [CrossRef]
- Patterson, R.; Nerren, J.; Kogut, M.; Court, P.; Villarreal-Ramos, B.; Seyfert, H.M.; Dalby, P.; Werling, D. Yeast-Surface Expressed BVDV E2 Protein Induces a Th1/Th2 Response in Naive T Cells. Dev. Comp. Immunol. 2012, 37, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, D.J.; Song, Q.J.; Duan, C.; Wang, S.H.; Wang, J.F.; Zhu, Y.H. Enhanced Im mune Responses to E2 Protein and DNA Formulated with ISA 61 VG Administered as a DNA Prime-Protein Boost Regimen against Bovine Viral Diarrhea Virus. Vaccine 2018, 36, 5591–5599. [Google Scholar] [CrossRef] [PubMed]
- Sharif, M.; Baek, Y.B.; Naveed, A.; Stalin, N.; Kang, M.I.; Park, S.I.; Soliman, M.; Cho, K.O. Porcine Sapovirus-Induced Tight Junction Dissociation via Activation of RhoA/ROCK/MLC Signaling Pathway. J. Virol. 2021, 95, e00051-21. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Direction a | Sequence (5′ → 3′) |
|---|---|---|
| IL-6 | F | GCTGAATCTTCCAAAAATGGAGG |
| R | GCTTCAGGATCTGGATCAGTG | |
| IL-1β | F | CCTCGGTTCCATGGGAGATG |
| R | AGGCACTGTTCCTCAGCTTC | |
| IFN-α | F | GTGAGGAAATACTTCCACAGACTCACT |
| R | TGAGGAAGAGAAGGCTCTCATGA | |
| IFN-β | F | CCTGTGCCTGATTTCATCATGA |
| R | GCAAGCTGTAGCTCCTGGAAAG | |
| Mx 1 | F | GTACGAGCCGAGTTCTCCAA |
| R | ATGTCCACAGCAGGCTCTTC | |
| GAPDH | F | AAAGTGGACATCGTCGCCAT |
| R | CCGTTCTCTGCCTTGACTGT |
| Primers Name | Direction a | Sequence (5′ → 3′) |
|---|---|---|
| BDNF | F | GCGCCCATGAAAGAAGTAAA |
| R | TCGTCAGACCTCTCGAACCT | |
| CREB | F | CCAGTTGCAAACATCAGTGG |
| R | TTGTGGGCATGAAGCAGTAG | |
| SCF | F | CCTTATGAAGAAGACACAAACTTGG |
| R | CCATCCCGGCGACATAGTTGAGGG | |
| β-actin | F | GCTCTTTTCCAGCCTTCCTT |
| R | GATGTCAACGTCACACTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-X.; Yang, G.-H.; Zhang, L.-L.; Wang, J.; Wang, J.-F. Melatonin as Immune Potentiator for Enhancing Subunit Vaccine Efficacy against Bovine Viral Diarrhea Virus. Vaccines 2021, 9, 1039. https://doi.org/10.3390/vaccines9091039
Wang Y-X, Yang G-H, Zhang L-L, Wang J, Wang J-F. Melatonin as Immune Potentiator for Enhancing Subunit Vaccine Efficacy against Bovine Viral Diarrhea Virus. Vaccines. 2021; 9(9):1039. https://doi.org/10.3390/vaccines9091039
Chicago/Turabian StyleWang, Yi-Xuan, Guang-Hui Yang, Lin-Lin Zhang, Jing Wang, and Jiu-Feng Wang. 2021. "Melatonin as Immune Potentiator for Enhancing Subunit Vaccine Efficacy against Bovine Viral Diarrhea Virus" Vaccines 9, no. 9: 1039. https://doi.org/10.3390/vaccines9091039
APA StyleWang, Y.-X., Yang, G.-H., Zhang, L.-L., Wang, J., & Wang, J.-F. (2021). Melatonin as Immune Potentiator for Enhancing Subunit Vaccine Efficacy against Bovine Viral Diarrhea Virus. Vaccines, 9(9), 1039. https://doi.org/10.3390/vaccines9091039